-
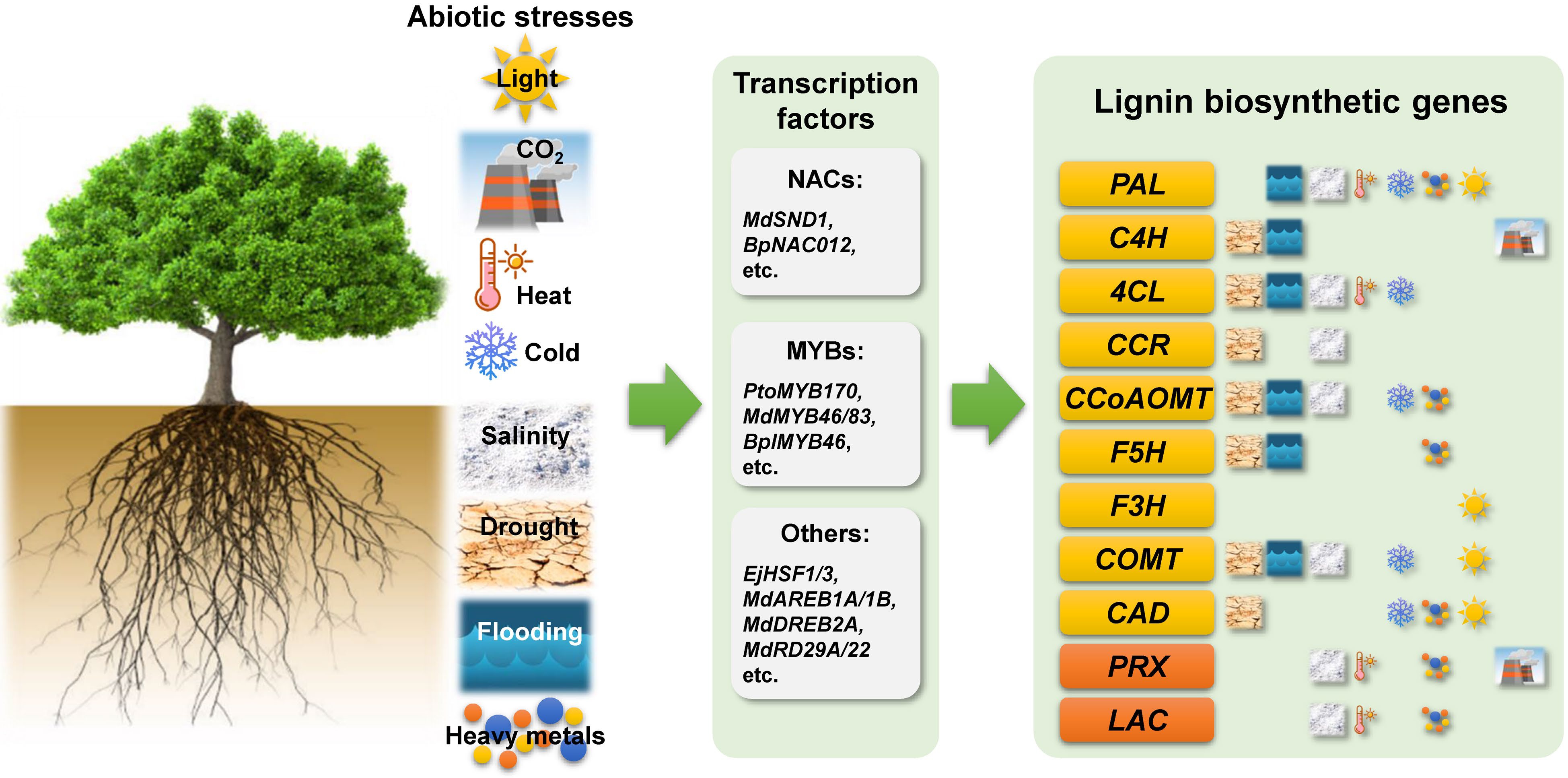
Figure 1.
Schematic diagram summarizing the various abiotic stresses affecting lignin biosynthesis in woody plants. Phenylalanine ammonia lyase (PAL), cinnamate 4-hydroxylase (C4H), 4-coumarate:coenzyme A ligase (4CL), cinnamoyl-CoA reductase (CCR), caffeoyl-CoA O-methyltransferase (CCoAOMT), ferulate 5-hydroxylase (F5H), flavonoid 3-hydroxylase (F3H), caffeic acid O-methyltransferase (COMT), cinnamyl alcohol dehydrogenase (CAD), peroxidase (PRX), and laccase (LAC).
-
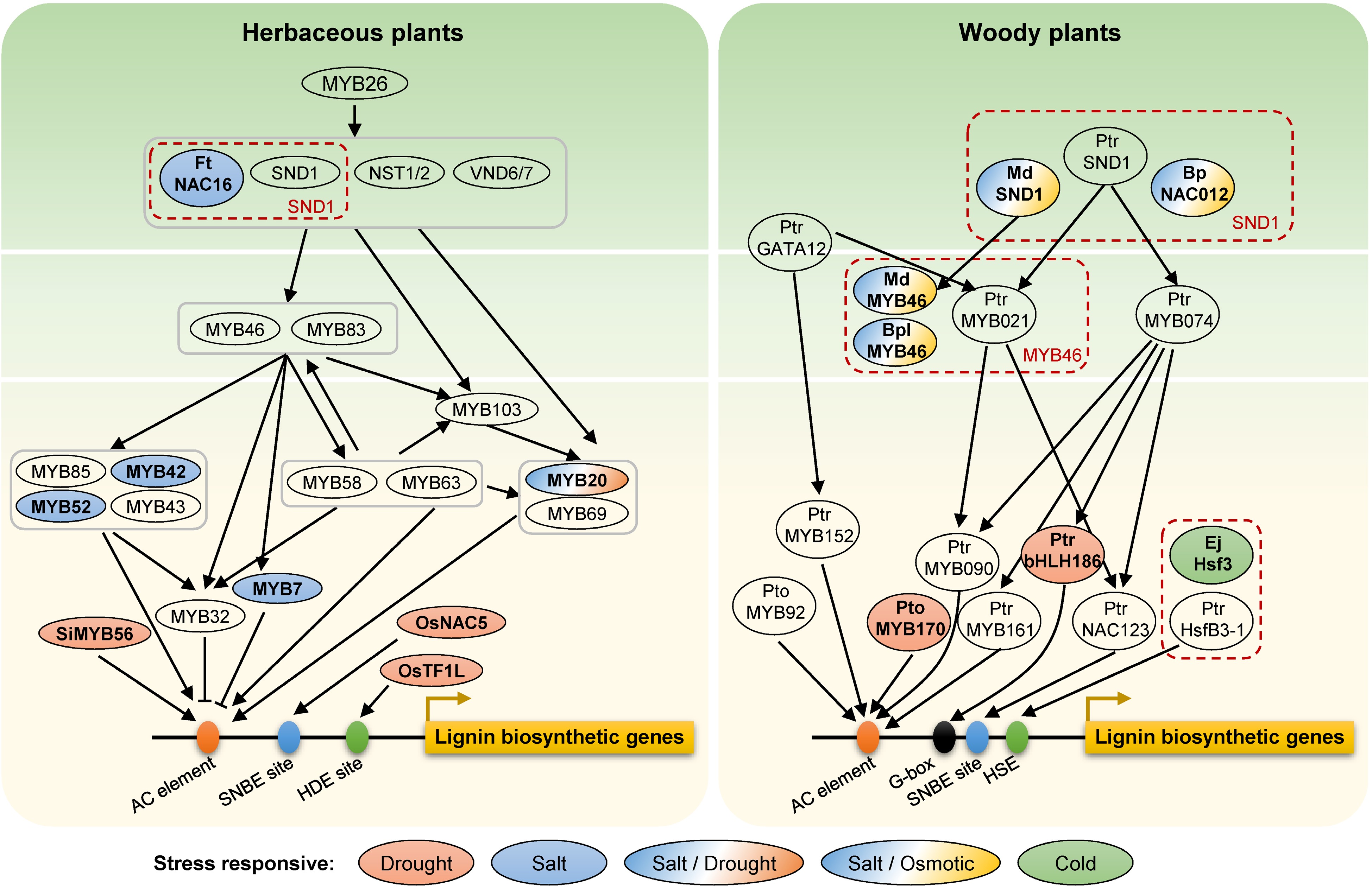
Figure 2.
Transcription factors in the lignin regulatory network involved in abiotic stress responses in herbaceous and woody plants. Genes in red dashed boxes represent orthologous genes in different species. Different colors show different abiotic stresses. Ft, Fagopyrum tataricum; Si, Setaria italic; Os, Oryza sativa; Ptr, Populus trichocarpa; Pto, Populus tomentosa; Md, Malus × domestica; Bp or Bpl, Betula platyphylla; Ej, Eriobotrya japonica; No prefix, Arabidopsis. AC element, SNBE site, HDE site, G-box, and HSE located on the promoter region of lignin biosynthetic genes represent the cis-acting elements for the binding of MYB, NAC, TF1L, bHLH and HSF transcrition factors, respectively.
-
Stress Species Tissue Treatment Main effects Reference Drought Populus tomentosa PtoMYB170-overexpressing Arabidopsis Desiccated without watering for 2 weeks and then rewatered Promoted lignin deposition by activating CCR2, COMT, CCoAOMT1 and C4H2 Xu et al.[91] Populus trichocarpa 4-month-old OE-PtrbHLH186-L4 plants Soil relative water content was reduced from 60% to 13% by withholding water Transregulated many monolignol genes and increased in G subunits Liu et al.[85] Leucaena leucocephala Stem and root 1% mannitol (w/v) Increased lignification and corresponding CCR protein accumulation Srivastava et al.[16] Eucalyptus urograndis Basal and apical regions of the stem Not irrigated until wilt Promoted lignin deposition and reduced the S/G ratio in the basal regions Moura-Sobczak et al.[12] Populus trichocarpa Young shoot tissues (top 3 cm) Supplying 150 ml water once per week for 4 weeks Decreased the lignin S/G ratio in young shoots Hori et al.[17] Flooding Populus × canescens Root Water level of the containers exceeded the soil surface by 2 to 3 cm Down-regulated lignin biosynthesis genes Kreuzwieser et al.[20] Syzygium samarangense Root The flooded pots maintained a depth of several centimeters of water in a plastic dish placed under the pots Lignin accumulation in epidermis and cortex in both normal and flooded batches Tuladhar et al.[13] Salt Populus canescens and Populus euphratica Leaf, stem, root 25 or 100 mM NaCl Increased in the lignin: carbohydrate ratio in both species. Janz et al.[21] Betula platyphylla Leaf of BpNAC012-overexpressing lines 200 mM NaCl Elicited higher expression levels of lignin biosynthetic genes and elevated lignin accumulation in BpNAC012-overexpressing lines Hu et al.[79] Betula platyphylla BplMYB46-overexpressing lines 200 mM NaCl for 10 d BplMYB46 improved salt and osmotic tolerance by increasing lignin deposition Guo et al.[80] Malus × domestica Leaf of MdSND1-overexpressing and MdSND1-RNAi lines 200 mM NaCl MdSND1 is directly involved in the regulation of lignin biosynthesis. Chen et al.[78] Simmondsia chinensis Leaf 50, 100 or 200 mM NaCl Reduced lignin production Alghamdi et al.[23] Coffea arabica Leaf 50, 100 or 150 mM NaCl Cell walls of coffee leaves have undergone changes in the polysaccharide and lignin composition de Lima et al.[14] Tamarix hispida Root 400 mM NaCl Two genes SAMS and COMT involved in lignin synthesis were highly responsive to NaCl stress Li et al.[22] Heat Eriobotrya japonica Fruit 40 °C EjHSF1 trans-activated the lignin biosynthesis-related genes Zeng et al.[94] Cold Eriobotrya japonica Fruit 0 or 5 °C Chilling condition during postharvest storage lead to increased expression levels of PAL, 4CL and CAD, and resulted in increased lignin content Liu et al.[32] Populus tremula × P. tremuloides cv. Muhs1 Stem 10 °C Increased lignin contents in cuttings Hausman et al.[33] Heavy metals Pinus sylvestris Needles, stem, and roots 0.5, 1, 2, or 4 mM Al(NO3)3 Roots affected by Al showed deformation in cell walls and higher lignification and suberization. Oleksyn et al.[41] Citrus sinensis and Citrus grandis Leaf, stem, root 1.0 mM AlCl3·6H2O DCBC protein promoted the synthesis of lignin to achieve the ability to immobilize Al Wu et al.[45] Nine Myrtaceae species roots 1.0 mM AlCl3·6H2O Lignin was formed only in the root tips in Melaleuca bracteata Tahara et al.[40] Camellia sinensis Root, leaf Al2(SO4)3·18H2O The down-regulation of F5H and the binding of Al and phenolic acids reduced the accumulation of lignin Xu et al.[44] Camellia sinensis Roots or in cultured cells 400 µM AlCl3 Reduced the activities of PAL and POD, and lignin content Ghanati et al.[43] Camellia sinensis Callus Cultures 106 µM Cd(NO3)2 Increased the lignin content in the root and stem calli Zagoskina et al.[46] Avicennia schaueriana Leaf, stem, root 0, 5, 15, 30, or 45 mg·L−1 CdCl2·5/2H2O Induced lignin deposition in xylem cells of all vegetative organs Garcia et al.[47] Populus × canescens Leaf, wood, root 50 µM CdSO4 Increased in GH3 activities and thereby shunted the metabolism to enhanced formation of lignin. Elobeid et al.[48] Scots pine (Pinus sylvestris) Root 5 or 50 µM Cd 50 µM Cd resulted in significant increases in lignin content Schützendübel et al.[49] Two Salix caprea isolates Root 0.5 mg·L−1 CdNO3·4H2O KH21 delayed the development of apoplastic barriers Vaculík et al.[53] Bruguiera gymnorrhiza and Rhizophora stylosa Root, leaf 100, 200, 300, or 400 mg·kg−1 CuCl2 Increased lignification in mangroves roots Cheng et al.[60] Six species of mangroves Root 400 µg·kg−1 ZnCl2,
200 µg·kg−1 CuCl2Three high metal-tolerant rhizophoraceous species exhibited a thick exodermis with high lignification Cheng et al.[61] Light Pinus radiata Callus cultures From the dark to a 16 h photoperiod Enhanced the enzymes CAD and PAL, and increased the amount of lignin Möller et al.[64] Malus domestica Fruit Direct sunlight, shaded, and severe sun damage Induced the expression levels of MdCOMT1 and MdCAD in the flesh, and increased the accumulation of lignin in cell walls of the flesh and skin in sudden exposure Torres et al.[65] Fagus sylvatica Leaf Leaves of the sun and shade crown Induced higher lignin contents in leaves in shade crown than in sun crown under ambient O3 concentrations Jehnes et al.[62] CO2 Populus tremula × alba Stems of young tree Elevated CO2 (800 µL·L−1) Increased lignin content in lower and middle stems Richet et al.[67] Betula pendula Green leaves Elevated CO2 (2 × ambient) Decreased contents of acid-soluble lignin in birch leaves Oksanen et al.[68] Fagus sylvatica Two-year-old beech seedlings Ambient and elevated CO2 Leaves in the elevated CO2 treatment contained less lignin than leaves in the ambient CO2 treatment Blaschke et al.[69] Table 1.
Effects of different abiotic stresses on lignin in woody plants.
Figures
(2)
Tables
(1)