-
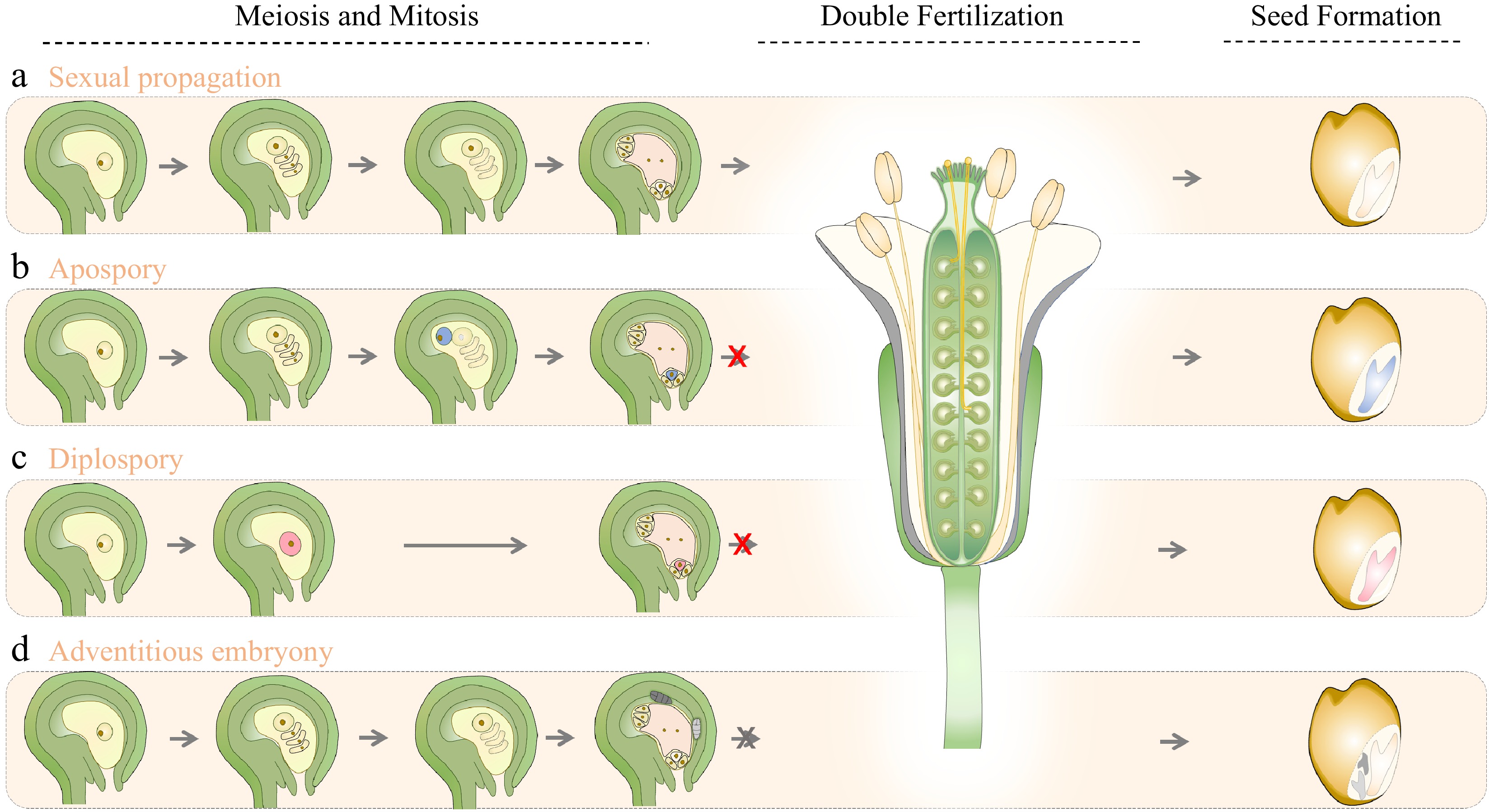
Figure 1.
Schematic representation of sexual and apomictic seed formation in flowering plants. Apomixis deviates from sexual propagation mainly in meiosis, mitosis and double fertilization, which are the primary developmental stages in forming seeds. This diagram depicts the (a) normal sexual reproduction process, and the asexual reproduction processes of apomixis, (b) including apospory, (c) diplospory, and (d) adventitious embryony in flowering plants. Arrows indicate the developmental sequences for seed production. Red crosses mark the bypass of double fertilization in gametophytic apomixis (apospory and diplospory). In gametophytic apomixis (b & c), diplospory produces diploid embryo sac through mitotic division from MMC (red), while the unreduced embryo sac in apospory initiates from a different ovule precursor cell (blue) that divides mitotically, and the unreduced egg cell (b & c) can further develop into embryo without gamete fusion. However, the endosperm formation of apospory and diplospory may need central cell fertilization. In (d) adventitious embryony, embryos initiate directly from somatic cells of the nucellus or integument that are adjacent to the embryo sac, and endosperm develops from the fertilized central cell.
-
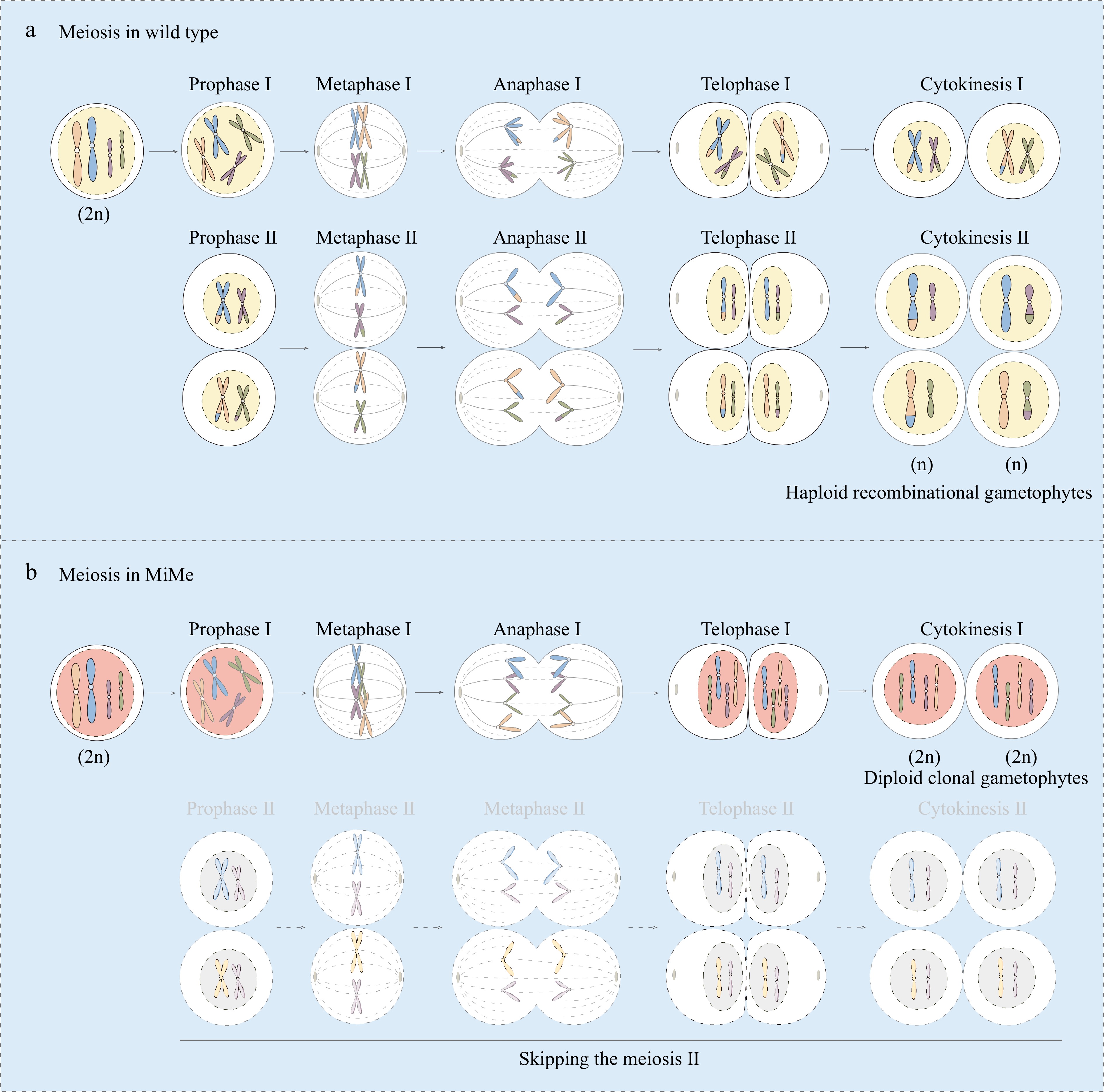
Figure 2.
Schematic representation of meiosis in wild type and MiMe plants. Meiosis in wild type plant is depicted in (a), and (b) exhibits the altered meiosis in MiMe plants. In normal meiosis, the chromosome undergoes one replication, but twice divisions (meiosis I and II). During meiotic prophase I, the homologous recombination takes places with exchange of genetical information between homologous chromosomes. After meiosis II, four haploid recombinational gametes are formed from one megasporocyte (a). However, only two diploid clonal gametes are formed from one megasporocyte in MiMe system without recombination event and meiosis II (b).
-

Figure 3.
Schematic representation of genome elimination and parthenogenesis. The haploid offspring can be induced through crossing plant carrying modified CENH3 gene with (a) wild type pollen, (b) mutating MTL or (c) DMP gene, or (d) nectopic expression of BBMs in the egg cell. In (a), genome elimination occurs in crosses between wild type pollen and CEHN3-based haploid inducer. The chromosome of haploid inducer is unstable during mitotic divisions of the early embryo, and will be lost to produce haploid plants, which are genetically identical to the genome of wild type parental gametophyte. (b) Depicts the haploid induction by chromosome fragmentation event. In mtl mutants, chromosomes undergo fragmentation with varied frequencies from microsporocyte to embryogenesis stage. During double fertilization, DMP is supposed to bridge the gamete fusion, however, mutating DMP results in gamete fusion failure and haploid production (c). Additionally, parthenogenesis can be induced in sexual plants by ectopic expression of BBMs in the egg cell (d).
Figures
(3)
Tables
(0)