-
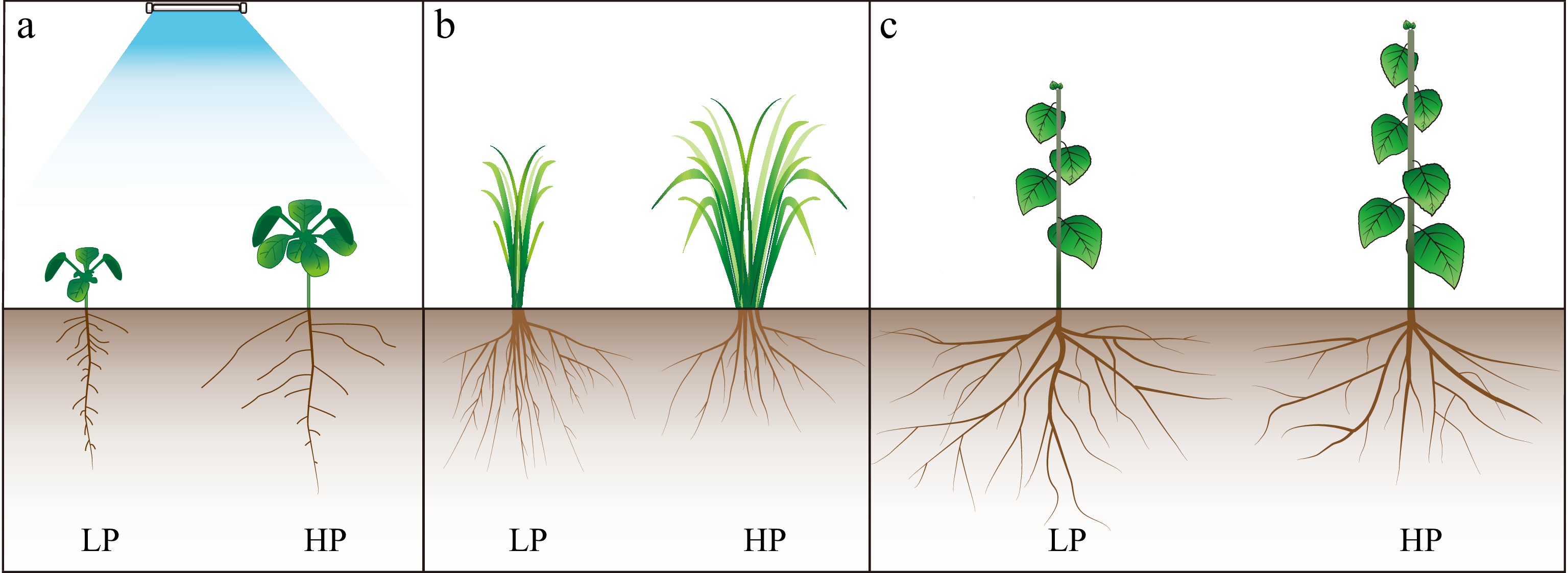
Figure 1.
Plant phenotype response in different phosphorus statuses. Phosphorus is one of the critical macronutrients for the growth and development of plants. When phosphorus availability varies, it triggers a cascade of physiological and morphological responses. These responses are manifested in different plant phenotypes. Under phosphorus-deficient conditions (LP), plants exhibit an adaptive strategy by increasing root length, density, and branching to produce more lateral roots and longer root hairs to improve their ability to take up more phosphorus from the soil. In addition, plant growth is often stunted with reduced shoot biomass and smaller leaves, limiting photosynthesis. Stem elongation is also affected, resulting in shorter plants, and there is a tendency for plants to increase the ratio of root-to-shoot (such as LP in (a)−(c)). In contrast, when phosphorus is abundant or in high status, the root system tends to develop more normally, without the exaggerated expansion that occurs under deficiency conditions. Roots and shoots are kept in equilibrium. It will show vigorous vegetation with larger leaves, more leaves, and normal stem length (such as HP in (a)−(c)).
-
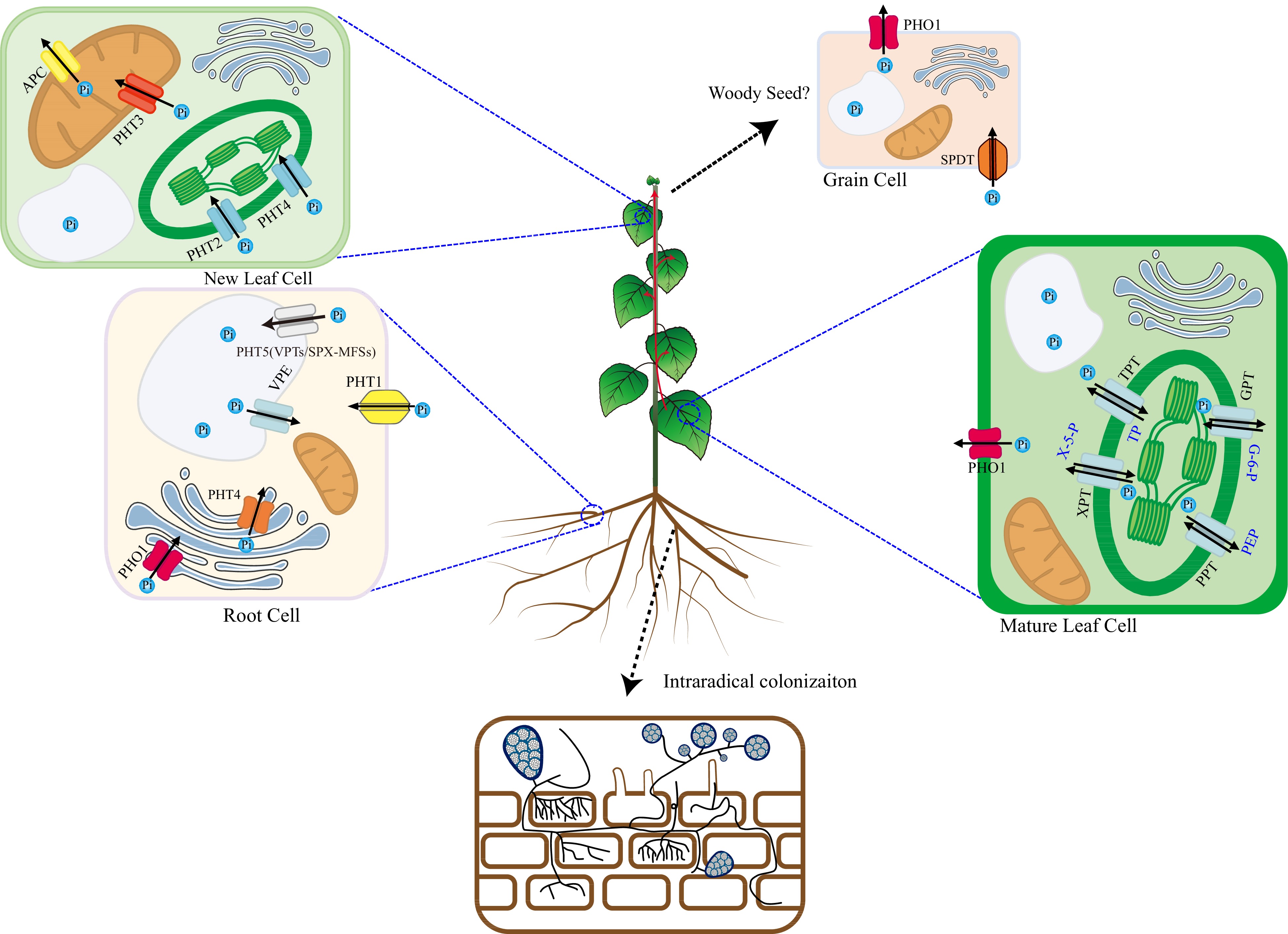
Figure 2.
Pi uptake, transport, and remobilization transporters in plants. In plants, several types of transporters are involved in the complex uptake, transport, and remobilization of inorganic phosphate (Pi), the primary form of phosphorus taken up by roots. Intracellular Transport: At the root-soil interface, Pi is taken up from the soil solution by high-affinity transporters, mainly PHT1 family proteins. Once inside the cytoplasm, Pi is a central hub for exchanging Pi between different organelles. Several transporters shuttle Pi across organellar membranes. For example, PHT/VPE transporters can move Pi into vacuoles for storage or release it when needed. Other transporters manage Pi fluxes into and out of chloroplasts (TPT, PHT2, PHT4, XPT, PPT, GPT), mitochondria (PHT3, APC), and the Golgi (PHT4, PHO1) apparatus to support essential metabolic processes. Long-Distance Transport: The long-distance transport of inorganic phosphate (Pi) by the PHO1 transporter, primarily localizing at cellular membranes, is a critical process that ensures the efficient distribution of this essential nutrient from roots to shoots and the developing tissues. PHO1 facilitates the loading of Pi into the xylem sap for its systemic movement through the vascular system of the plant. Pi Remobilization in Mature Leaves: During senescence or phosphorus deficiency, Pi can be remobilized in mature leaves. PHO1 and possibly PHT1 transporters help to mobilize stored or excess Pi back into the cytoplasm. This prepares it for export to younger leaves or developing grains. Pi transport in Grains: Developing grains requires significant Pi to grow and mature. Specific Pi transporters, such as SPDT, can mediate the uptake of external Pi into grain cells. And PHO1 plays a role in expelling Pi from grains when necessary.
-
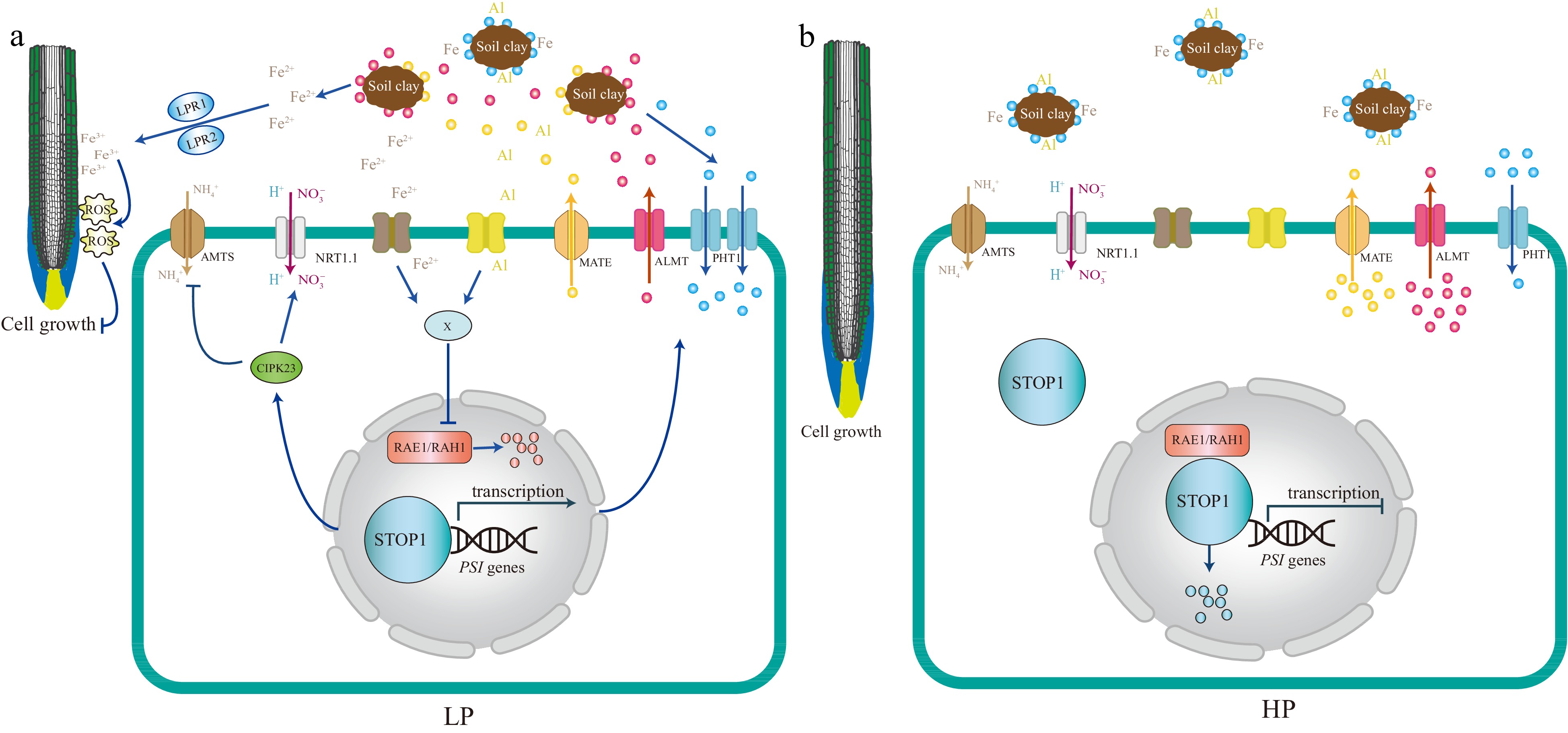
Figure 3.
A model based on the current understanding of the local Pi signaling pathway. STOP1 acts as a master regulator integrating multiple facets of local Pi signaling, including ion homeostasis, nutrient mobilization, and tolerance to environmental stressors such as pH changes and toxic ions, particularly Iron (Fe) and Aluminum (Al). Under low Pi conditions (LP), STOP1 is primarily activated by Fe. Fe acts similarly to Al under moderately acidic conditions. It is hypothesized that extracellular Fe (or Al) may induce the accumulation of an unknown compound X in the cell, preventing the degradation of STOP1, which is controlled by the F-box proteins RAE1 and RAH1. The STOP1 activity is upregulated, which induces the expression of ALMT1, an aluminum-activated malate transporter, releasing malate into the rhizosphere. This exuded malate interacts with the ferroxidases LPR1 and LPR2 and promotes the aggregation of Fe3+ in the apoplast of root cells, which stimulates the formation of reactive oxygen species (ROS). The accumulation of ROS negatively impacts primary root growth by preventing elongation. In addition, ammonium transporters of the AMT1 family (AMTs in the figure), which are up-regulated by low Pi, are involved in local Pi signaling. Ammonium uptake via AMTs involves proton extrusion, which acidifies the rhizosphere. High rhizosphere acidity is sufficient to induce STOP1 activation, but Fe greatly enhances the level of STOP1 activation. STOP1 prevents ammonium toxicity by upregulating CIPK23 and then inhibiting AMT transporter activity. Furthermore, STOP1 directly upregulates the NRT1.1, which co-transport nitrate and protons, providing a mechanism to increase rhizosphere tolerance to low pH. Under Pi-sufficient conditions (HP), root growth is generally normal because the excess phosphate provides an adequate supply for essential cellular processes. Excess phosphorus may interact with metal ions such as Aluminum (Al) and Iron (Fe), forming complexes that make these metals less bioavailable to plants. Blue dot: Pi, Red dot: malate, Yellow dot: citrate.
-
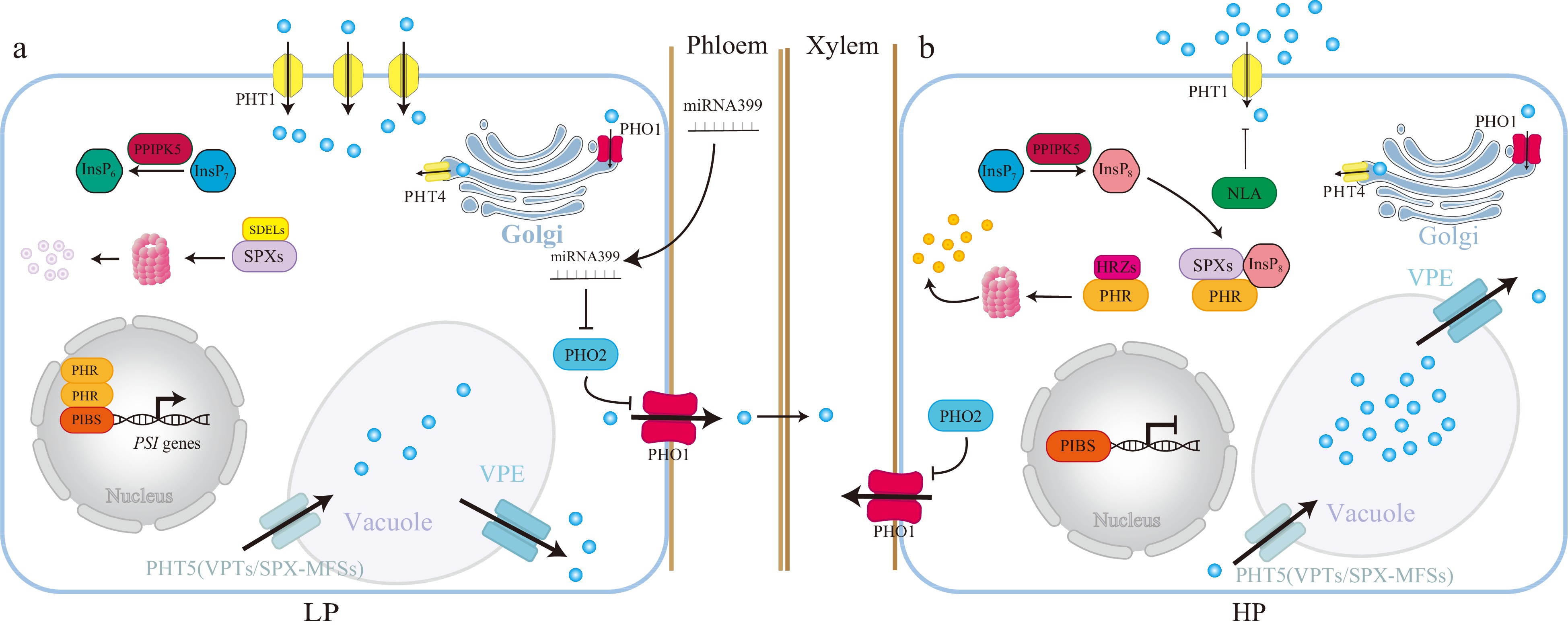
Figure 4.
A model depicts the InsPs-SPXs-PHRs-centered systematic Pi signaling in response to Pi deficiency. Under Pi deficiency, the Pi and ATP levels are lower, and PPIP5K converts the InsP7 to InsP6, leading to a decrease in the InsP8 level. The SPXs-PHRs dimer is dissociated, and SPXs are degraded by SDELs[127]. Thus, PHRs are free to activate the transcription of PSI genes (PHT1s, microRNA827, microRNA399 and PHT5). PHRs activate the Pi uptake by up-regulating the transcription of PHT1 transporters such as PHT1;1[142]. PHRs further increase the plasma-membrane localized PHT1;1 by increasing the transcription of microRNA827 (miR827), which targets and degrades NLA, a ubiquitin E3 ligase of PHT1[143]. PHRs reduce the Pi storage in the vacuole by increasing the transcription of miR827, which also targets and degrades PHT5, a vacuole Pi influx[53]. PHRs stimulate the expression of the vacuole Pi efflux VPE via the P1BS motif[54]. Thus, PHRs reduce the Pi storage and increase Pi activation by increasing the expression of Pi influx but reducing the expression of Pi efflux in the vacuole, the Pi storage organelle. PHR1 also activates the transcription of microRNA399 (miR399), and miR399 moves from shoot to root upon Pi deficiency to target and degrade the mRNA of a ubiquitin-conjugating E2 genes-PHO2[144−148]. PHO2 degrades the PHO1 and disrupts the xylem loading of Pi from root to shoot[148]. Therefore, PHR1 activates miR399 to repress PHO2 and thus ultimately leads to activating the PHO1-mediated Pi allocation from root to shoot upon Pi deficiency. Conversely, under Pi-sufficient conditions, the PPIP5K converts the InsP7 to InsP8, and InsP8-SPXs block the activity of PHRs. The PHRs are also degraded by HRZs[149], and PSI genes are less activated by PHRs. The intracellular Pi is more likely to be stored in a vacuole, and Pi uptake is also reduced, ultimately reducing Pi activation by increasing the Pi storage. Thus, the PHR-centered systematic Pi signaling is not activated.
-
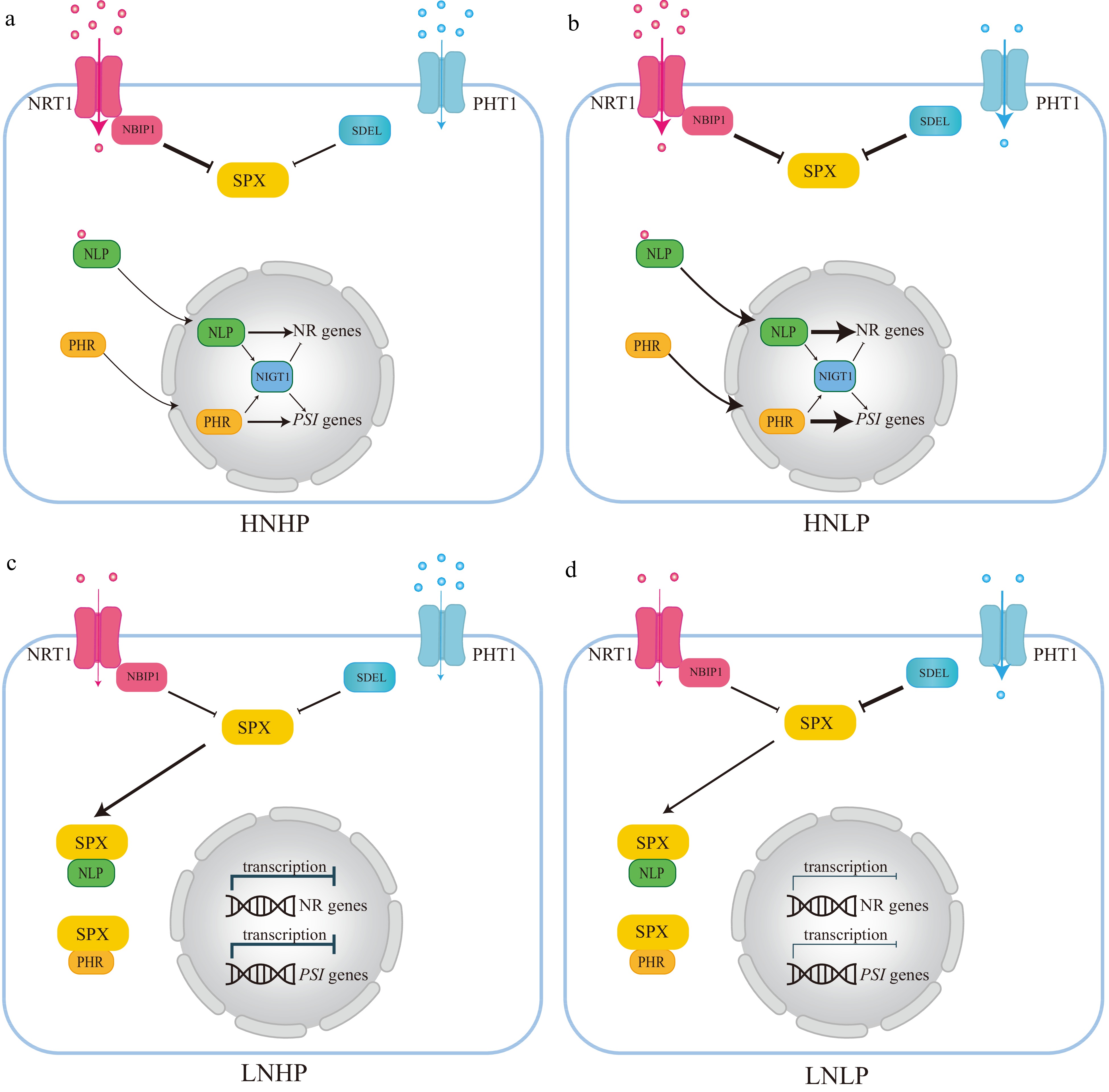
Figure 5.
The integration network of nitrogen (N) and phosphorus (P) interactions based on Arabidopsis and rice. (a) Under high nitrate and high phosphate (HNHP) conditions, SPX protein, through NRT1.1-NBIP1, undergoes partial degradation and releases PHRs and NLPs from the cytoplasm into the nuclei to activate both PSI and nitrate-responsive gene expression. (b) Under high nitrate and low phosphate conditions (HNLP), NRT1,1-NBIP1 and SDELs mediate the degradation of SPX protein, leading to a significant reduction in SPX protein levels, thereby retaining PHR and NLP in the cytoplasm and repressing the expression of both PSI and nitrate-responsive genes. (c) and (d) SPX proteins tend to accumulate heavily in the cytoplasm in both high phosphate (HP) and low phosphate (LP) nitrate-limited environments (LNHP or LNLP). This can also lead to the retention of key regulatory factors like PHR1 and NLPs, and repression of the expression of both PSI and nitrate-responsive genes. Blue dot: Pi, Red dot: Nitrate.
-
Species Root Shoot Mycorrhizal symbiosis Primary
rootLateral
rootShoot/root
ratioLeaf number Leaf growth Leaf angle Height Photosynthetic Shoot biomass AM ECM Arabidopsis Inhibit Inhibit Reduce Reduce Inhibit – Reduce Inhibit Reduce No No Rice Enhance Enhance Reduce Reduce Inhibit Reduce Reduce Inhibit Reduce Colonize No Poplar Enhance Enhance Reduce Reduce Inhibit – Reduce Inhibit Reduce Colonize Colonize AM: Arbuscular Mycorrhizal fungi; EM: Ectomycorrhizal fungi; '–' represents unexplored. Table 1.
The effects of phosphorus deficiency on growth and mycorrhizal fungal symbiosis in Arabidopsis, rice, and poplar.
Figures
(5)
Tables
(1)