-
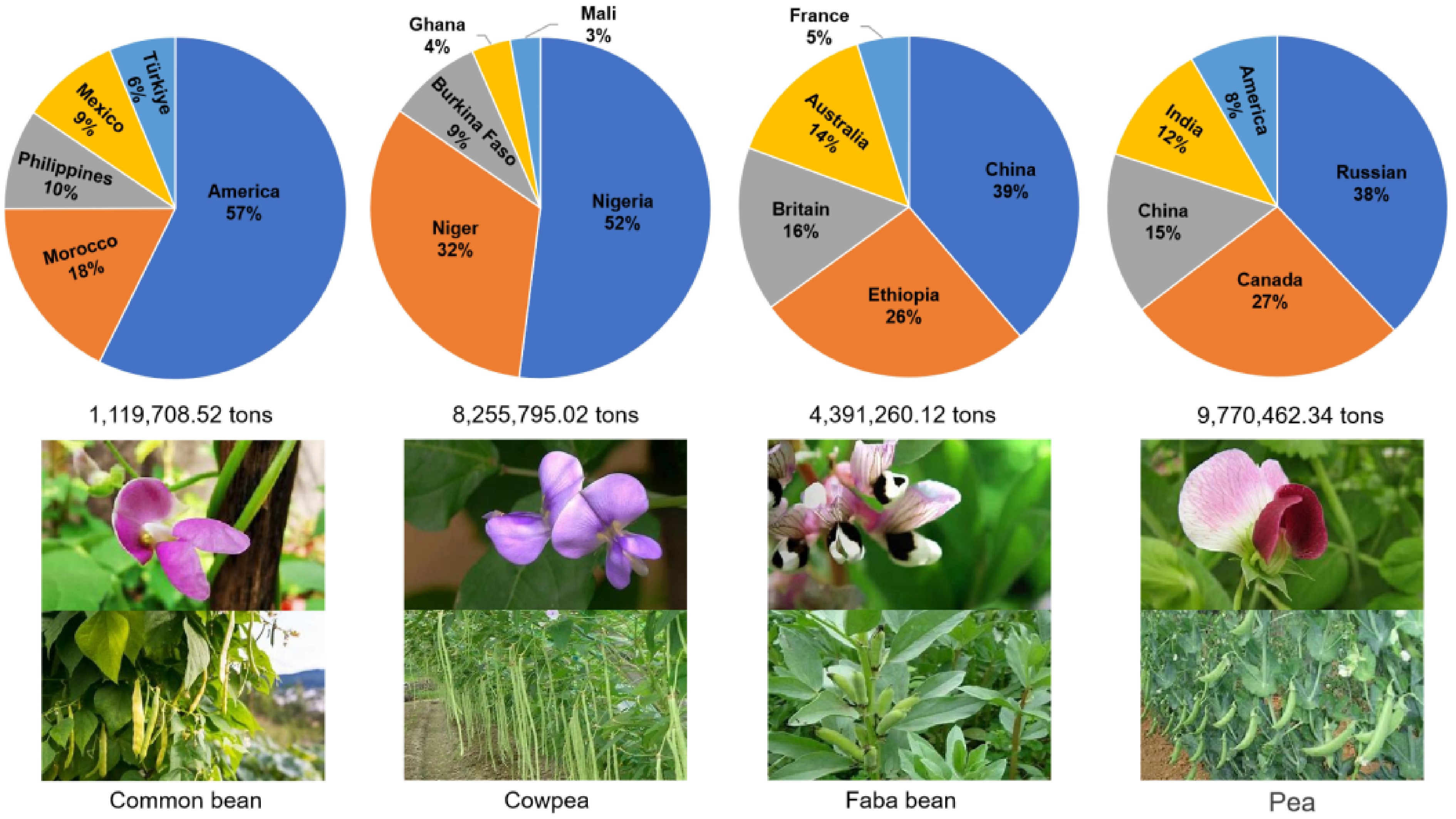
Figure 1.
Global yield percentages of the major vegetable legumes by country. The value under each pie chart represents the total production of this legume across the top five countries. Values in each pie chart represent the production percentage of these five countries. The global production data of common bean, cowpea, faba bean, and pea were sourced from FAOSTAT 2023 (
www.fao.org/faostat/en/#data/QCL ). -
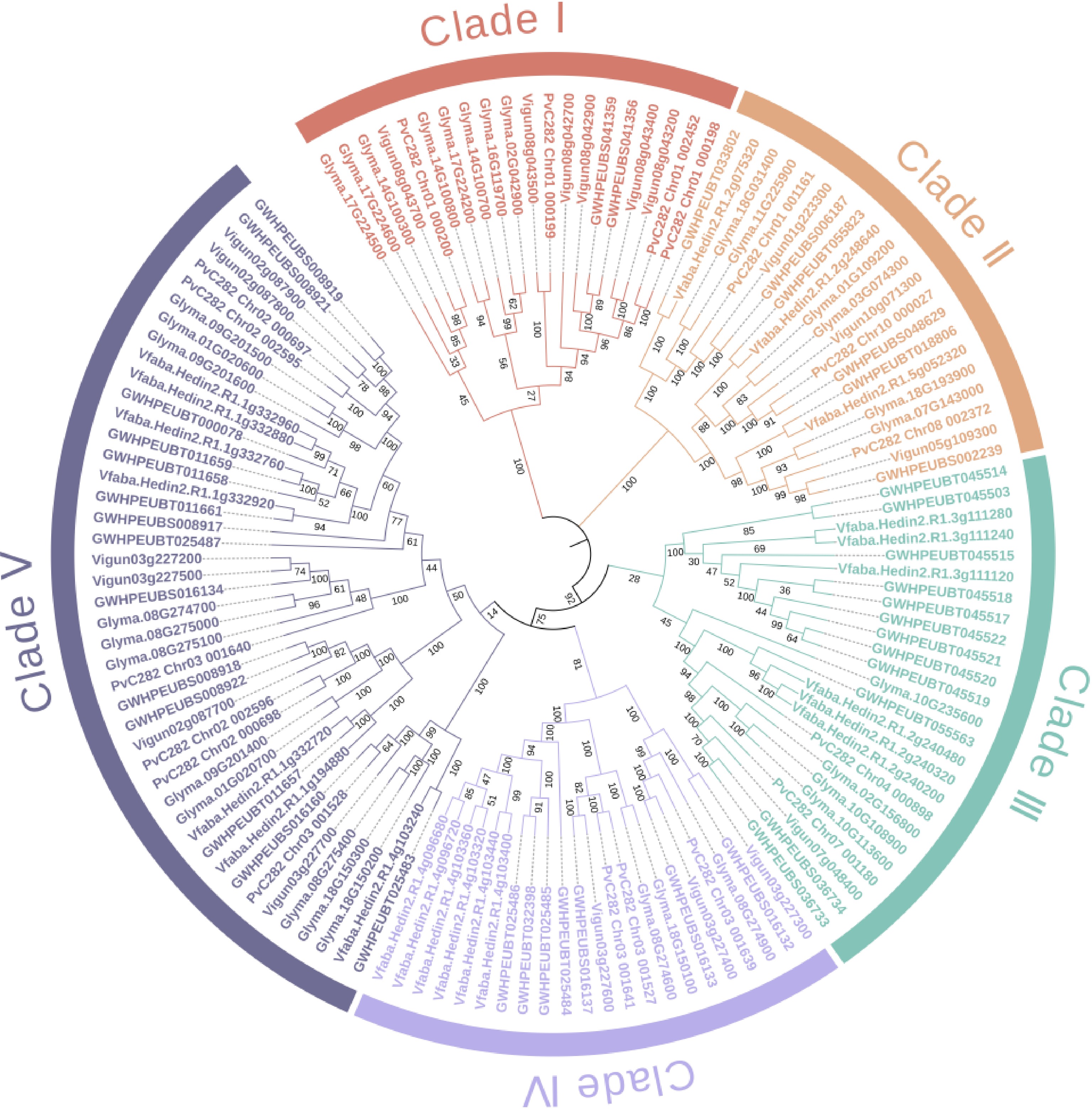
Figure 2.
Neighbor-joining tree of the legume-type lectin proteins in six vegetable legume crops. The identifier names of soybean, pea, faba bean, common bean, mung bean, and cowpea start with 'Glyma', 'GWHPEUBT', 'Vfaba', 'PvC282', 'GWHPEUBS', and 'Vigun', respectively. The lectin sequences were retrieved from the reference genomes: Glycine max Wm82.a2.v1 (soybean), Pisum sativum Pea_genome_GWHEUBT00000000.1 (pea), Vicia faba v1.1 (faba bean), common bean (in-house unpublished, available upon request), Vigna radiata Mungbean_genome_GWHEUBS00000000.1 (mung bean) and Vigna unguiculata v1.1 (cowpea), which are deposited in Phytozome (
https://phytozome-next.jgi.doe.gov/ ) or China National Center for Bioinformation (www.cncb.ac.cn ). In total, 132 legume lectin genes were identified across the six species, with the following distribution: soybean (32), pea (24), faba bean (22), common bean (18), mung bean (17), and cowpea (19). Their corresponding protein sequences are listed in Supplementary Table S1. A neighbor-joining phylogenetic tree was constructed based on these sequences (Fig. 2), resolving them into five major clades (Clades I−V), comprising 20, 21, 25, 22, and 44 genes, respectively. -
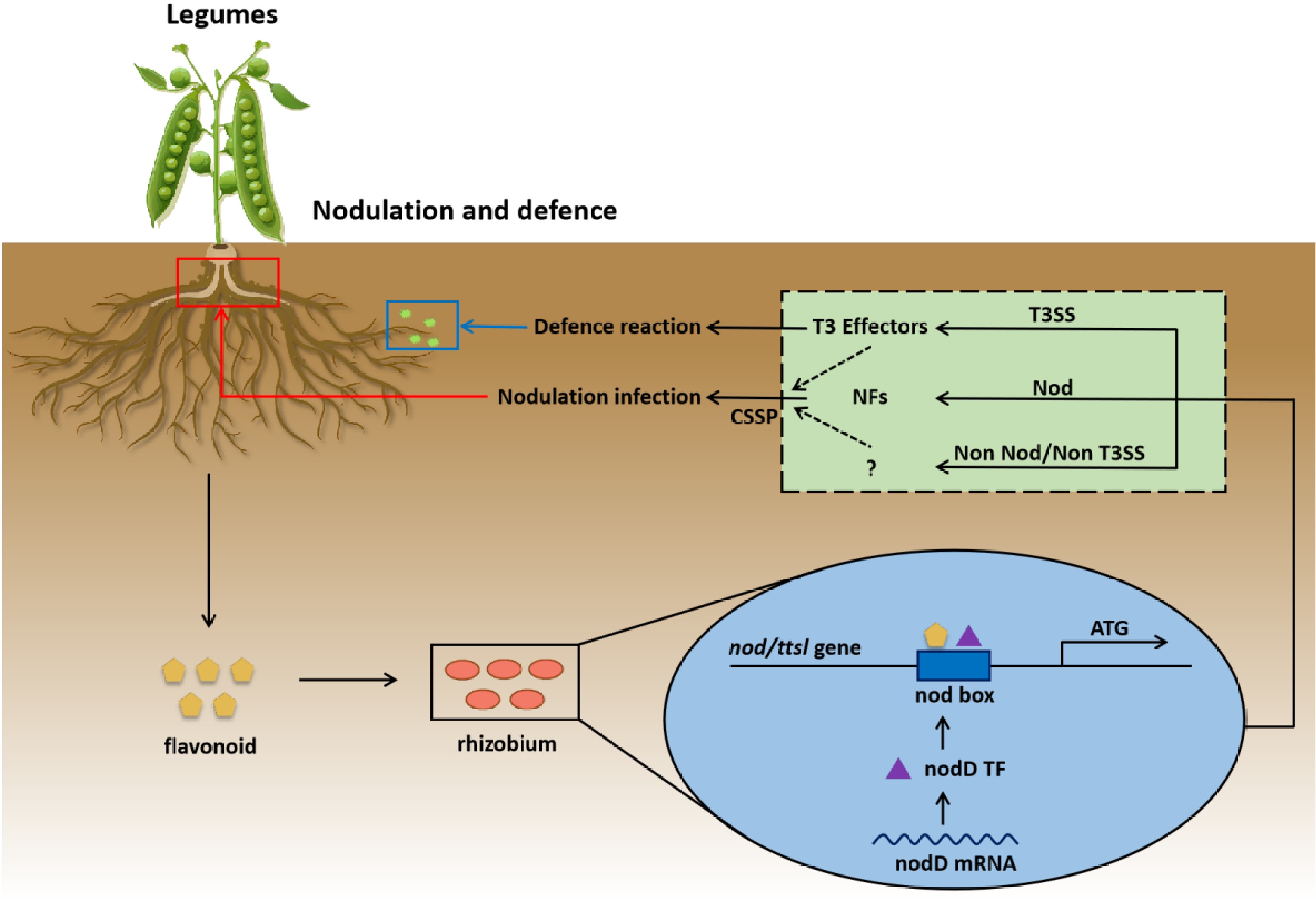
Figure 3.
Schematic diagram of the symbiotic nitrogen fixation mechanism with rhizobia. In this process, flavonoids secreted by legume roots act as initial signals and are recognized by rhizobia. Inside the rhizobia cell, flavonoids induce the expression of the nodD gene. The nodD transcription factor (TF), which binds to the nod box adjacent to the nod/ttsl genes, initiates the production of Nod factors (NFs) related to the Nod operon. NFs, likely in association with cell-secreted signaling proteins (CSSP), drive the nodulation infection of the legume root. Concurrently, a parallel pathway involves the type III secretion system (T3SS) in rhizobia. T3SS secretes T3 effectors, which trigger a defense reaction in the legume. Additionally, a 'Non Nod/Non T3SS' pathway exists, which may represent a regulatory mechanism to fine-tune rhizobia-legume interactions. This mechanism helps prevent excessive nodulation or over-activation of the defense response, thus maintaining a balanced interaction and ensuring the efficient progress of symbiotic nitrogen fixation.
-
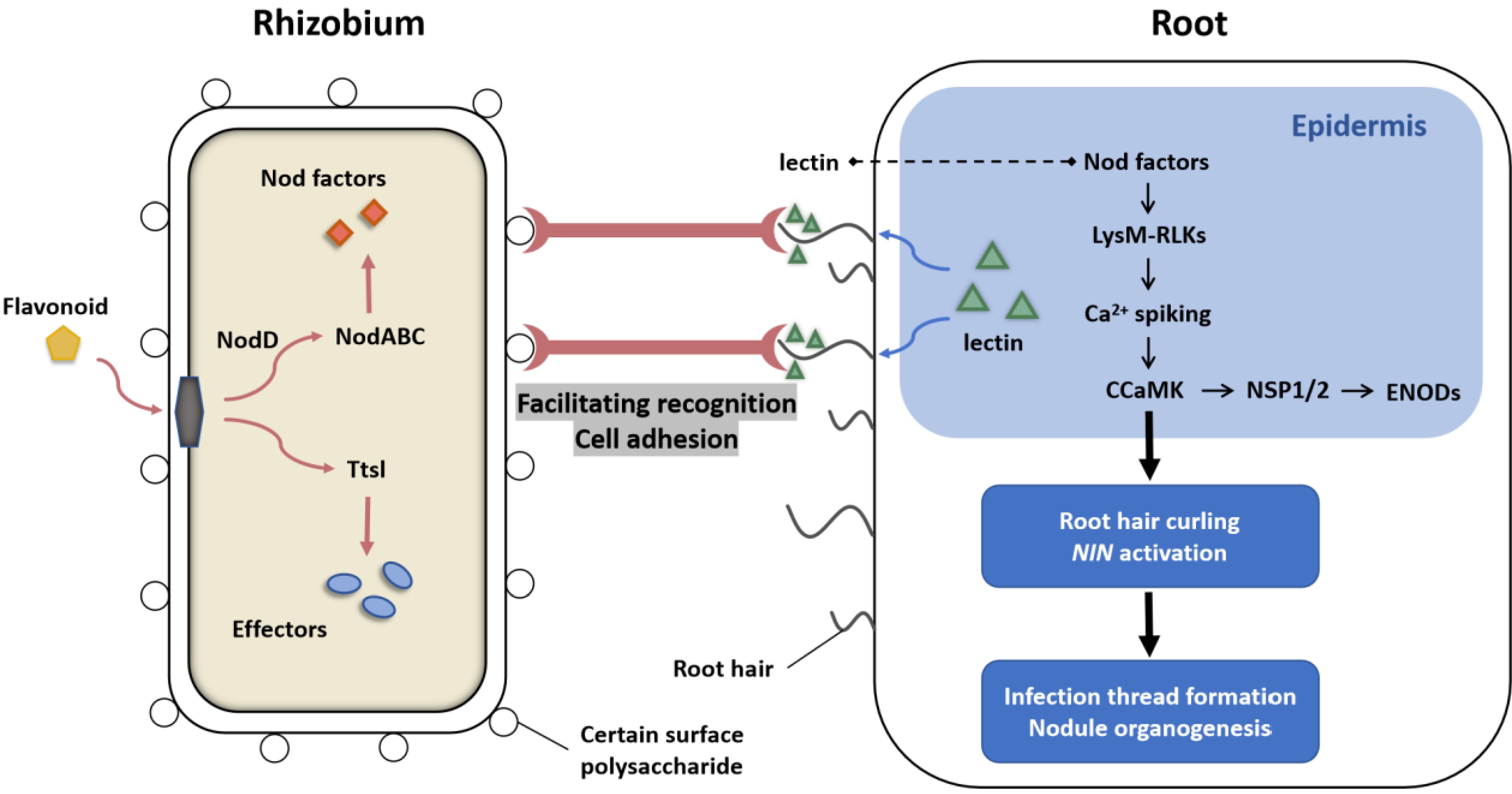
Figure 4.
Mutual recognition mechanism between lectins and rhizobia. Flavonoids interact with rhizobial components such as nodD and induce the expression of nod genes. Lectins, acting as 'molecular sentinels' on the root surface, accumulate at the tips of root hairs-key sites for rhizobial infection. Their carbohydrate-binding domains specifically recognize glycan modifications on the rhizobial surface, promoting bacterial colonization and initial attachment to the host root. This lectin-mediated accumulation enhances local bacterial density on root hairs, creating a favorable micro-environment for subsequent infection. By stabilizing the interaction between rhizobia and root hairs, lectins may also facilitate intercellular communication and signal exchange among rhizobia, increasing the chances of successful symbiosis. Notably, lectins can also recognize specific sugar structures on NFs, possibly through direct binding with the lectin's carbohydrate-binding site and the sugar chains of NFs. The NF signal is subsequently perceived by LysM receptor kinases, such as NFP and LYK3, in the root epidermis. Upon NF recognition, these receptors trigger root hair deformation and intracellular Ca2+ spiking, initiating downstream signaling cascades. This includes the activation of the CCaMK-NSP1/2-ENOD pathway and NIN, ultimately driving infection thread formation and nodule organogenesis. However, some evidence proves that lectins are not indispensable for root nodule nitrogen fixation, suggesting redundant recognition pathways.
-
Lectin Mainly distribution Sugar-binding specificity Function Ref. Phaseolus vulgaris agglutinin (PHA) Seeds N-glycans Insect-resistance, antibacterial, antitumor [10,14] Conconvalin A (ConA) Seeds D-Glucose, D-Mannose Antitumor, promote angiogenesis [15,16] Soybean lectin (SBA) Seeds and roots D-Galactose, N-Acetylgalactosamine (GalNAc) Agglutinate red blood cells, promote lymphocyte division [12,17] Peanut agglutinin (PNA) Seeds and roots D-Galactose, GalNAc Antitumor, antibacterial [17,18] Pisum sativum agglutinin (PSA) Seeds D-Mannose, methyl-α-D-glucoside Antitumor [19,20] Table 1.
Classification and distribution of major plant lectins.
-
Classification Representative strain Host plants Key characteristics Ref. Rhizobium R. leguminosarum Pisum sativum, Vicia faba, Phaseolus vulgaris Fast-growing type, symbiotic with temperate leguminous plants, and high nitrogen-fixing efficiency [26−29] Bradyrhizobium B. japonicum Glycine max, Arachis hypogaea Slow-growing type, acid soil-tolerant, and high nitrogen-fixing efficiency [30,31] Ensifer E. meliloti Medicago sativa, Melilotus officinalis, Phaseolus vulgaris Fast-growing type, strong salt tolerance, and high symbiotic nitrogen-fixing ability [29,32,33] Sinorhizobium S. fredii Glycine max, Vigna radiata, Phaseolus vulgaris Fast-growing type, salt-alkali tolerant, and with a large genome [34] Mesorhizobium M. loti Lotus japonicus, Trifolium repens Moderate growth rate, tolerance to infertile soil, and enhancement of plant resistance to pathogenic bacteria [35,36] Azorhizobium A. caulinodans Sesbania rostrata Moderate growth rate and high nitrogen-fixing efficiency [37] Neorhizobium N. galegae Medicago sativa Fast-growing type, aerobic bacteria, adapted to low-oxygen environments after symbiosis with plants. [38] Pararhizobium P. giardinii Phaseolus vulgari, Lens culinaris Fast-growing type, acid soil-tolerant, and high nitrogen-fixing efficiency [29,39] Allorhizobium A. vitis Vitis vinifera Mainly infect plants as pathogens and do not symbiotically fix nitrogen with leguminous plants [40] Table 2.
Classification and characteristics of major rhizobial genera involved in symbiotic nitrogen fixation.
-
Legume Strain Area Ref. Alfalfa S. meliloti Mexico, Japan,
Northwestern Iran[41,42] S. medicae Mediterranean [36] Soybean B. lingoningense Xinjiang Region [43] S. fredii Xinjiang Region,
Loess Plateau,
Huang-Huai-hai Plain[43,44] B. japonicum Heilongjiang Province [44] Common
beanR. etli Central and South
America, Europe[45] R. leguminosarum Andes, Nepal [46,47] Bradyrhizobium sp. Acidic soil of southern China [48] Pea R. laguerreae Northwestern Spain, Tunisia [49,50] R. leguminosarum and
R. ruizarguesonisTurkey [51] R. anhuiense Shandong Peninsula [52] Peanut B. liaoningense Shandong, Hebei, Jiangsu, Guangdong, and
Guangxi Province[53] B. yuanmingense and the
close relative of B. japonicumHenan and Sichuan
Province[53] B. japonicum and B. elkanii Argentina [54] B. lioningense, B. chanariense, B. aponicum, and B. etae Moroco [55] Cowpea B. elkanii Africa [57] Table 3.
Reported dominant rhizobium strains of legumes in specific areas.
Figures
(4)
Tables
(3)