-
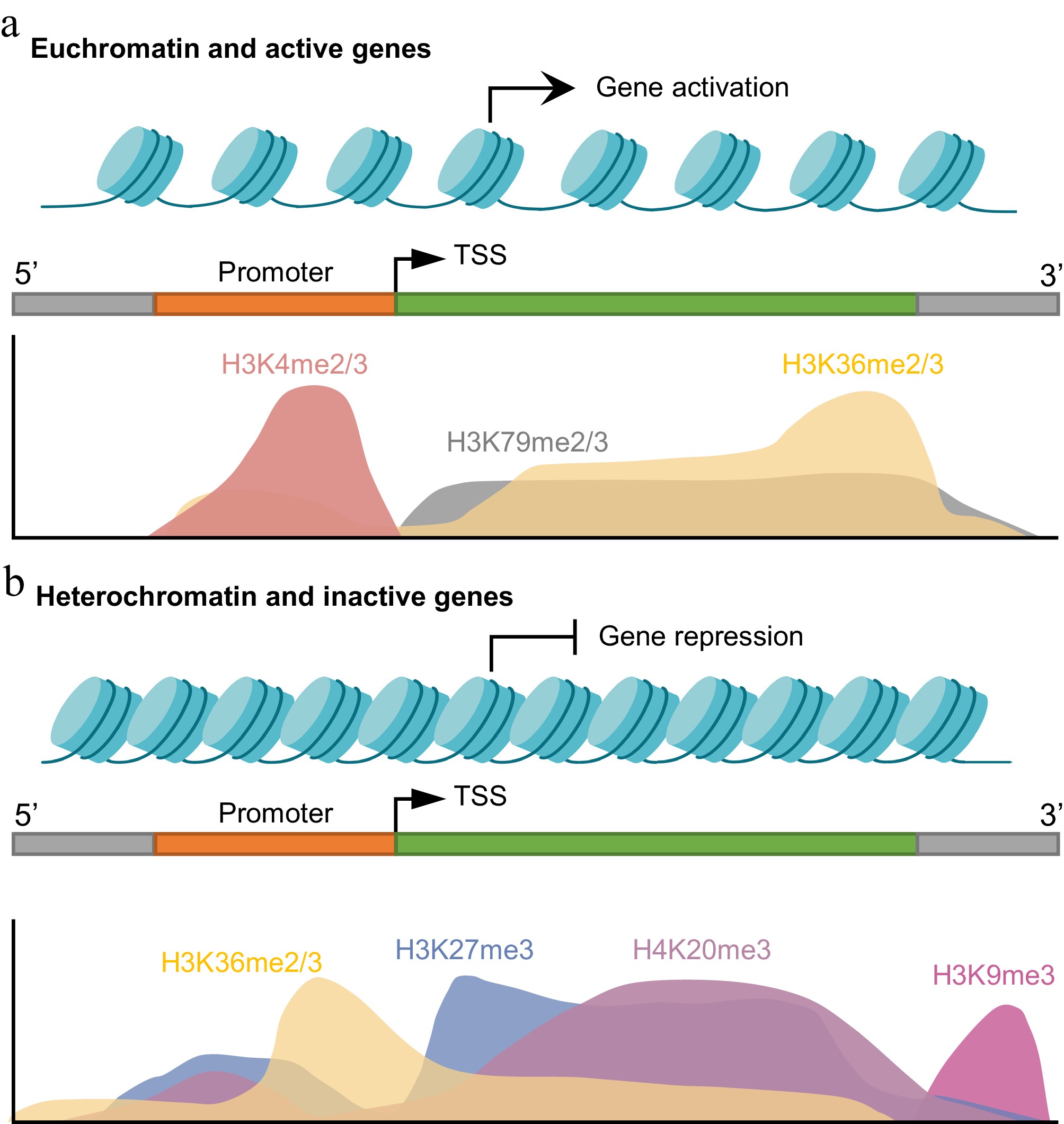
Figure 1.
Spatial distribution of histone lysine methylation marks in active and repressed genes. (a) In euchromatic regions linked to active transcription, H3K4me2/3 is enriched at the promoters and transcription start sites (TSSs), whereas H3K79me2/3 and H3K36me2/3 are found across gene bodies and facilitate transcriptional elongation. (b) Transcriptionally repressed regions in heterochromatin are marked by broad domains of H3K27me3 and H4K20me3, with H3K9me3 being primarily concentrated in constitutive heterochromatin, including telomeric and repeat-rich regions. In these repressive regions, H3K36me2/3 levels are reduced across gene bodies but may be locally enriched near the TSS. Created in BioRender. Xingmin, H. (2026)
https://BioRender.com/807zrla . -
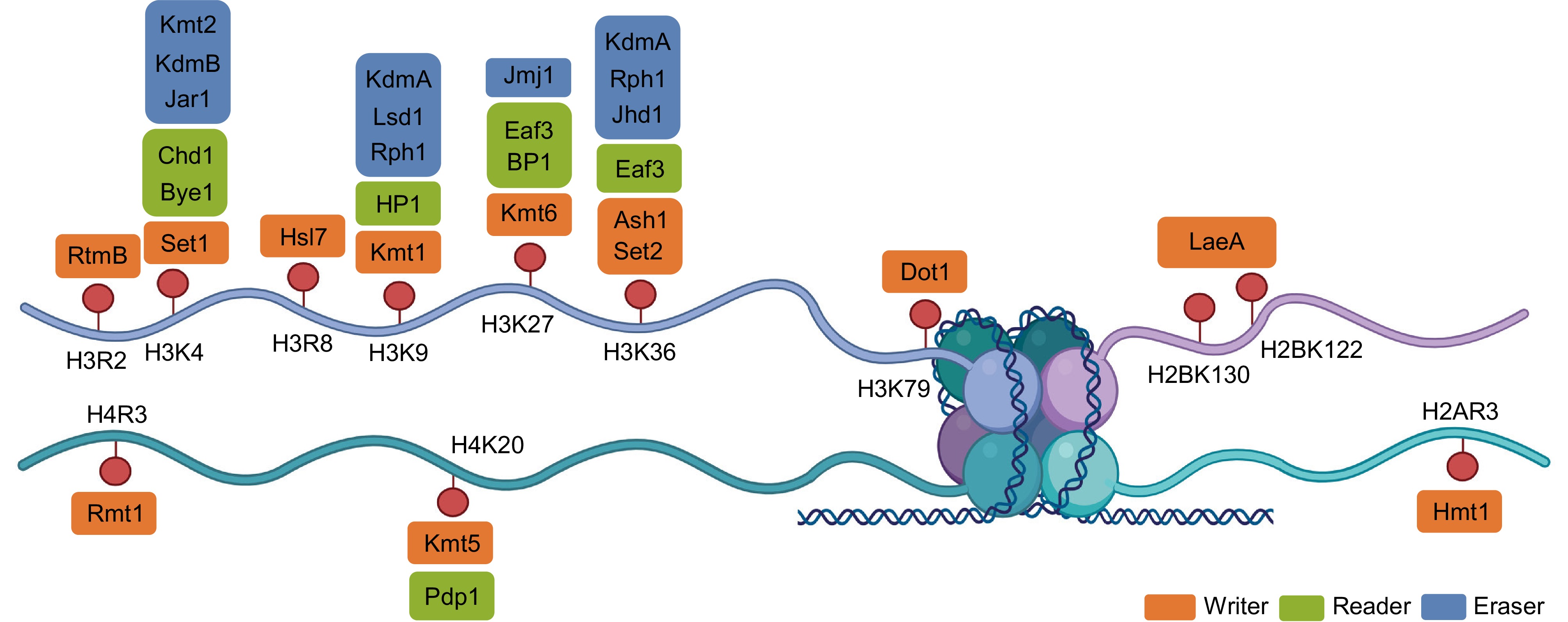
Figure 2.
Structural organization of histone methylation sites and the associated writer–reader–eraser proteins in fungi. Schematic representation of the core histone tails (H3, H4, H2A, and H2B) showing major lysine and arginine methylation sites, including H3R2, H3K4, H3K9, H3K27, H3K36, H3K79, H4K20, H2BK122/K130, and H2AR3. Each modification is presented alongside representative fungal methyltransferases (writers), demethylases (erasers), and reader proteins. Writer proteins (orange) include Set1 (H3K4), Kmt1 (H3K9), Kmt6 (H3K27), Set2 and Ash1 (H3K36), Dot1 (H3K79), Kmt5 (H4K20), LaeA (H2B), and Rmt1/Hmt1 (arginine). Reader proteins (green), such as HP1, Chd1, Bye1, Eaf3, and BP1, recognize methylated histones to mediate downstream chromatin remodeling and transcriptional changes. Erasers (blue) comprise Lsd1 and JmjC-domain-containing demethylases acting on distinct lysine residues. This diagram summarizes the coordinated protein machinery that maintains the state of the fungal epigenome. Created in BioRender. Xingmin, H. (2026)
https://BioRender.com/3r8vxo7 . -
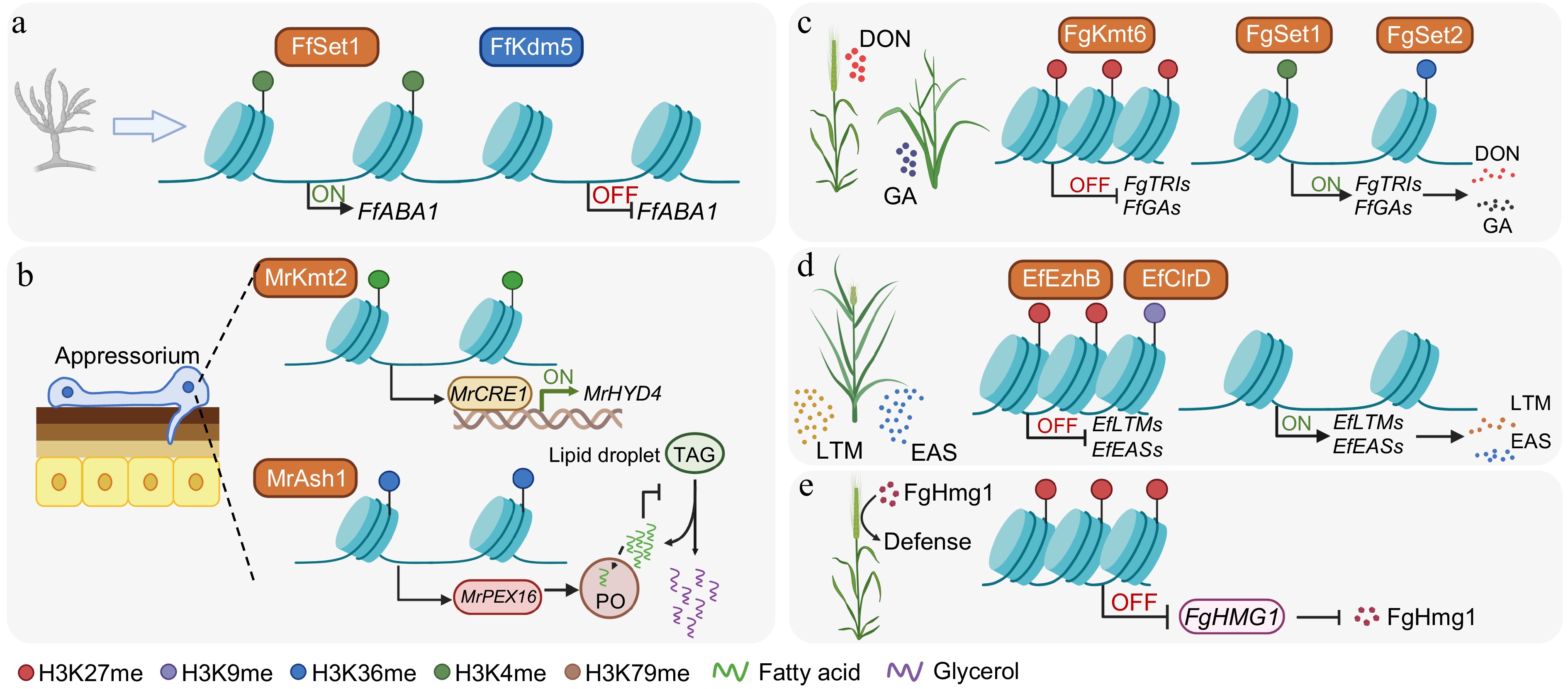
Figure 3.
Histone methylation-dependent mechanisms regulating fungal development, metabolism, and pathogenicity. (a) Regulation of asexual development by activating and repressive marks. In F. fujikuroi, Set1 deposits H3K4 methylation at the promoter of FfABA1, a central regulator of conidiation, to promote its transcription. Conversely, the removal of H3K4 methylation by the demethylase Kdm5 reduces FfABA1 expression. (b) Control of the formation of the infection structure and lipid metabolism in M. robertsii. During host infection, Kmt2-catalyzed H3K4me3 activates the transcription factor MrCRE1, which, in turn, induces the hydrophobin gene MrHYD4 to facilitate appressorium formation and surface adhesion. Simultaneously, Ash1-mediated H3K36me2 activates MrPEX16, supporting peroxisome (PO) biogenesis and fatty acid β-oxidation. This process prevents excessive lipid accumulation, promotes triacylglycerol (TAG) hydrolysis within lipid droplets, and supports glycerol production necessary for appressorial turgor generation and penetration. (c) In F. graminearum and F. fujikuroi, Kmt6-dependent H3K27me3 establishes repressive chromatin over the deoxynivalenol (DON) and gibberellic acid (GA) biosynthetic clusters, respectively. This repression is counteracted by Set1-mediated H3K4 methylation and Set2-mediated H3K36 methylation, which sustain the transcriptional activation of these clusters during pathogenesis. (d) Heterochromatin-mediated silencing and host-induced derepression of alkaloid gene clusters in E. festucae. The methyltransferases EzhB and ClrD catalyze H3K27me3 and H3K9me3, respectively, to cooperatively silence the lolitrem (LTM) and ergot alkaloid (EAS) clusters during axenic growth. During symbiosis, this dual heterochromatic repression is relieved, leading to the activation of LTM and EAS clusters and the production of protective alkaloids. (e) In F. graminearum, PRC2/FgKMT6-dependent H3K27me3 establishes a facultative repressive chromatin state at the pathogen-associated molecular pattern gene FgHMG1, which encodes a secreted glycoside hydrolase. This H3K27me3-mediated repression reduces host recognition and immune activation, thereby attenuating its immune elicitation and facilitating successful infection. Created in BioRender. Xingmin, H. (2026)
https://BioRender.com/lmnbvna . -
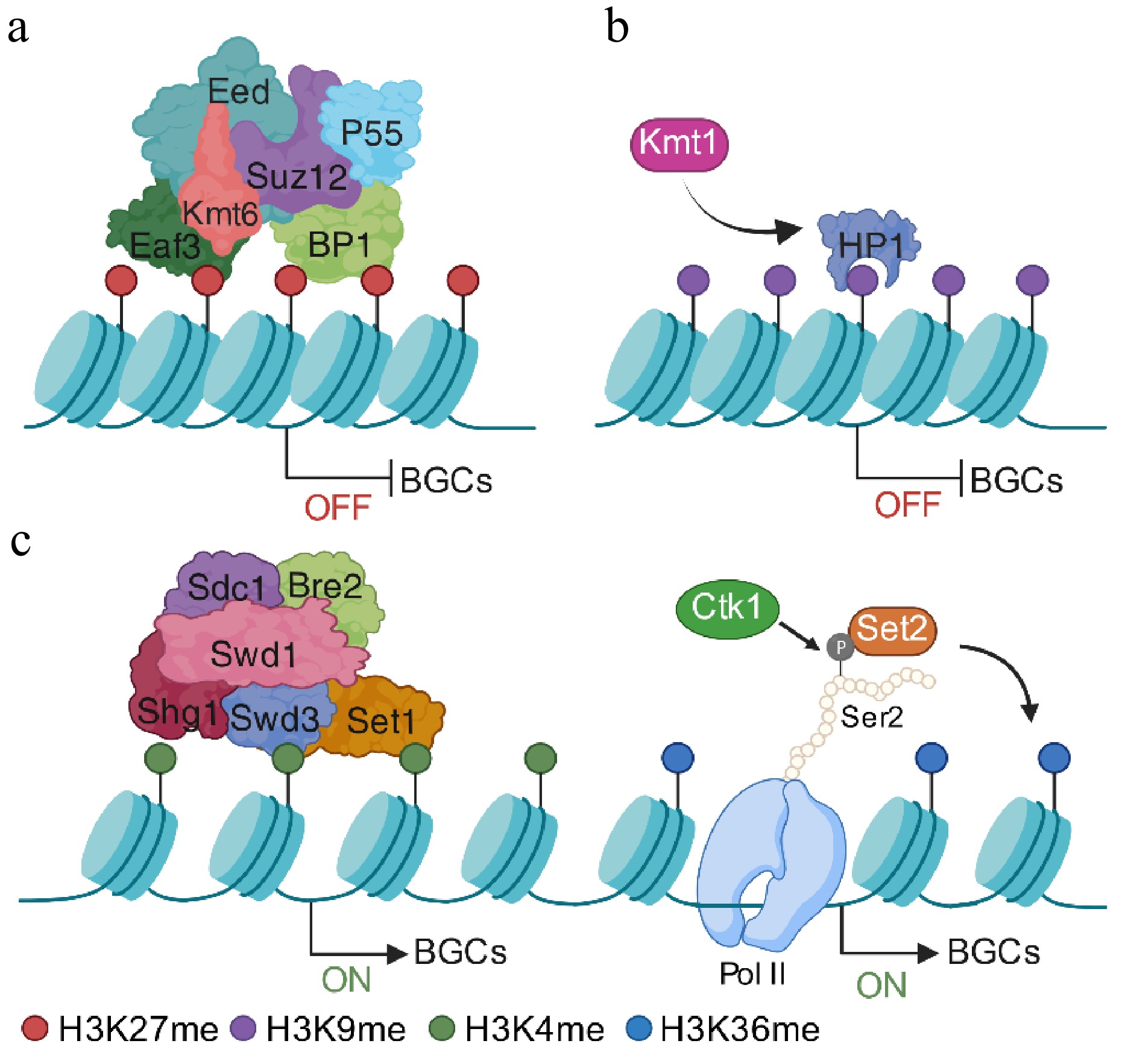
Figure 4.
Epigenetic regulation of fungal secondary metabolites' BGCs via histone methylation. (a) Under noninducing conditions, BGCs are maintained in a transcriptionally silent yet reversible state. In fungi possessing a functional PRC2, the catalytic subunit Kmt6 deposits H3K27me3 to establish facultative heterochromatin over BGC regions. (b) Alternatively, in fungi lacking PRC2, Kmt1-mediated H3K9 methylation cooperates with the reader protein HP1 to stabilize compact chromatin and restrict transcriptional accessibility. (c) Upon pathway induction, the removal or redistribution of repressive markers enables chromatin relaxation. The COMPASS complex, including Set1, deposits H3K4 methylation at BGC promoters to facilitate transcriptional initiation. Following derepression, active BGC expression is sustained by Set2-mediated H3K36 methylation across gene bodies. This mark is coupled to RNA Polymerase II elongation via Ser2 phosphorylation of the C-terminal domain, ensuring efficient transcriptional processivity and robust secondary metabolite production. Created in BioRender. Xingmin, H. (2026)
https://BioRender.com/55u0m5b
Figures
(4)
Tables
(0)