









-
Chrysanthemum is a perennial ornamental flower with various shapes and colors and is commercially popular as potted plants and cut flowers. Chrysanthemum white rust is an autoecious rust fungus, caused by Puccinia horiana Hennings[1]. It is one of the most destructive diseases of chrysanthemum worldwide and could be called 'the cancer' of the chrysanthemum. The disease first appeared in Japan in 1895 and eventually spread to China and other Asian countries[2−4]. Recent studies revealed the life cycle of this pathogen on a single host by light microscopy[5], scanning and transmission electron microscopy[6], and in situ hybridization[7]. Bonde et al. also revealed the overwinter ability of chrysanthemum white rust[6]. The common approach for chrysanthemum white rust is biological control; Cladosporium cladosporioides and Cladosporium pseudocladosporioides are potential new fungal antagonists[8], while the liquid formulation of Bacillus spp. is a bacterial control agent[9]. Previously, Alaei et al. used a molecular method based on rDNA internal transcribed spacer (ITS) sequence analysis to detect white rust[10]. As a research method, single-nucleotide polymorphisms (SNPs) are widely used to analyze the phylogenetic relationship among different pathogens[11]. A multiplex real-time PCR assay has also been used to detect P. horiana[12]. To identify potential genetic sources of resistance to white rust in chrysanthemum, disease classification standards have been developed[3,13−17]. Recently, an effective DNA marker related to P. horiana resistance locus was developed by genome-wide association study method[18]. However, the genes related to resistance were barely reported, except the WRKY15-1, a transcription factor worked on the salicylic acid signaling pathway, which could promote the resistance to the fungus[19, 20].
Plants have evolved a variety of strategies that counteract pathogen attack[21−23]. Defense strategies include physical barriers, antimicrobial compounds, pattern recognition receptors (PRRs), and resistance genes (R genes) along with the immune system[24−26]. Previous research has established that R gene mainly encodes nucleotide-binding and leucine-rich repeat (NB-LRR) proteins[27−29]. They interact with Avr effector proteins to induce a rapid and intense resistance response, called effector-triggered immunity (ETI). The ETI response is usually associated with the hypersensitive response (HR). In HR, localized programmed cell death limits the growth of pathogens in plant cells[24]. The N-terminal Toll-like interleukin-1 receptor (TIR) and a coiledcoil (CC) domains have the function of activating the defense signal[29−32]. The central NB-ARC domain consists of three subdomains: NB, ARC1, and ARC2. The ARC domain is named based upon its presence in APAF-1 (apoptotic protease-activating factor-1), R proteins and CED-4 (Caenorhabditis elegans death-4 protein)[33−34]. The NB-ARC domain acts as a nucleotide-binding pocket to hydrolyze Adenosine triphosphate (ATP) and induce the conformational change of R protein[27,31]. Several studies have shown that R genes respond to fungi, oomycetes, and bacteria in crops and horticultural plants including tomato (Solanum lycopersicum)[35], wheat (Triticum aestivum)[36,37], rice (Oryza sativa)[38], cucumber (Cucumis sativus)[39], brown mustard (Brassica juncea)[40], chickpea (Cicer arietinum)[41], apple (Malus domestica)[42], and melon (Cucumis melo)[43], sunflower (Helianthus annuus)[44−46], and rose (Rosa rugosa)[47]. However, it is unknown whether R genes are related to resistance to chrysanthemum white rust.
In this study, we identified and isolated the potential resistance chrysanthemum white rust gene in four chrysanthemum cultivars with different susceptibilities to chrysanthemum white rust. We predicted the conserved domain sequence and carried out a phylogenetic analysis. Then we explored the gene expression levels in fresh leaves of two cultivars after inoculation with P. horiana. Finally, we illustrated the function of CmCC-NB-ARC by constructing the overexpression vector and transformed into the susceptible chrysanthemum cultivar. This work would lay a theoretical foundation for analyzing the involvement of CC-NB-ARC family genes in chrysanthemum white rust resistance, and might contributed to the development of new resistant cultivars.
-
The four chrysanthemum cultivars 'C029', 'Fenhuaxiaori', 'LZ08-61', and 'Jinba' were used as specific experimental materials and planted in a greenhouse at 25 ºC ± 2 ºC, 16 h light and 8 h dark from Shenyang Agricultural University, Shenyang, China, since 2018. The disease index was properly evaluated according to the method created by Zhu et al.[3].
The tissue culture seedlings C029 and LZ08-61 for fungal inoculation and real-time PCR were grown under a specific condition of 16 h/8 h photoperiod at 24 ºC. For inoculation, it was taken from a telium of P. horiana on the leaf of an LZ08-61 seedling and resuspended in 0.05% Tween-20 sterile water. Then sprayed on the backs of the both cultivars' leaves fully dabbed with sterile water in advance. The plants were grown at 17–20 ºC in the dark for 3 d and then cultured under a 16 h/8 h photoperiod. Humidity was kept above 95%.
Nucleic acid isolation and quality assessment
-
Genomic DNA was extracted from fresh leaves by the CTAB method[48]. The method of DNA quality detection was conducted by Feng et al[49].
After that, we used the EASYspin Plus Complex Plant RNA Kit (Aidlab, Beijing, China) to extract total RNA from fresh leaves collected at the seedling stage (starting two months after cutting propagation). Assessment of RNA integrity and synthesis of first-strand complementary DNA referred to the description of Feng et al.[49].
Cloning and analysis of the gene potentially related to chrysanthemum white rust
-
We used the sequence of the conserved NBS domain from the five plants Artemisia annua (accession no. PWA98207)[50], Lactuca sativa (accession no. PLY80179)[51], Helianthus annuus (accession no. AY490793)[52], Triticum aestivum (accession no. ABS29034)[53], and Arabidopsis thaliana (accession no. At1g56510)[54], which are resistant to rust and other diseases, to search the chrysanthemum reference database (
www.amwayabrc.com/zh-cn/index.html )[55].We identified the gene CHR00059759 and amplified its coding and genomic sequences with the primers F-‘ATGACATACACTGGCTTGGAA’, R-‘TCAGAACGGGAATATCTTGATATATAAACATA’ from the highly resistant cultivar C029 and susceptible cultivars Fenhuaxiaori, LZ08-61, and Jinba. Specific fragments clone and domains prediction of the gene was established according to the methods of Feng et al.[56].
Homologous sequence analysis was performed through the BLAST tool in the NCBI database (
www.ncbi.nlm.nih.gov/blast ). The open reading frame was searched via the ORF Finder panel in the NCBI database (www.ncbi.nlm.nih.gov/orffinder/ ). Conserved domains of proteins were identified in the NCBI Conserved Domain Search online tool (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ). Coding sequences and amino acid sequences were aligned using DNAman software (Lynnon Biosoft, USA).Phylogenetic analysis of CmCC-NB-ARC
-
According to the protein sequences CmCC-NB-ARC, the homologies of other plants were downloaded from NCBI by a BLAST-search to construct a phylogenetic tree. Sequences from 15 species were identified: Actinidia chinensis var. chinensis (accession no. PSR88102.1), Artemisia annua (accession no. PWA98207.1), Camellia sinensis (accession no. XP_028096865.1), Camellia sinensis var. Sinensis (accession no. THG00403.1), Coffea arabica (accession no. XP_027097933.1), Coffea canephora (accession no. CDP05558.1), Coffea eugenioides (accession no. XP_027153382.1), Cynara cardunculus var. scolymus (accession no. XP_024991410.1), Helianthus annuus (accession no. XP_022025247.1), Lactuca saligna (accession no. CAB4075315.1), Lactuca sativa (accession no. XP_023766728.1), Mikania micrantha (accession no. KAD7117470.1), Nyssa sinensis (accession no. KAA8535375.1), Solanum tuberosum (accession no. XP_006366308.1), and Tanacetum cinerariifolium (accession no. GEV88943.1). ClustalW (EMBL, Cambridge, UK). These were used for amino acid sequence alignment. Besides, the neighbor-joining tree method[57] and 1,000 bootstrap replications[58] were used to construct the phylogenetic tree by MEGA 7.0 software[59].
Quantitative real-time PCR analysis of CmCC-NB-ARC
-
We collected leaves from tissue culture seedlings of chrysanthemum cultivars inoculated with the fungus resuspended in 0.05 % Tween-20. We took one fresh leaf from each variety and used three leaves as a sample. Leaves of the same variety were inoculated with sterile water as a blank control.
The reagents and cycling conditions for quantitative real-time PCR were carried out as per the methods of Feng et al.[56]. Each experiment had three biological and three technical replicates. CmACTIN (CHR00061013) was used as a reference gene. The expression levels of the gene CHR00059759 (F-‘GTTCCCTGCCCGATGGTTGATTAG’, R-‘TGCCGACTAGGTCCATTAGGTACG’) and CmACTIN (F-‘TCCGTTGCCCTGAGGTTCT’, R-‘GATTTCCTTGCTCATCCTGTCA’) in chrysanthemum cultivars were determined using the 2−ΔΔCᴛ method[60].
Overexpression vector construction and transformation
-
To determine the function of CmCC-NB-ARC, we constructed an overexpression vector according to the method described by Feng et al.[56]. The coding region of the gene was amplified from the resistant cultivar C029 with the primers F 5′-GAACACGGGGGACTCTTGACATGACATACACTGGCTTG-3′ and R 5′-TGGTCACCTGTAATTCACACTCAGAACGGGAATATCTTG-3′, which were designed on NEB online software (http://www.neb-china.com/). Then the PCR product was fused into the plant binary expression vector pCAMBIA3301 that was precisely excised by double digestion with NcoⅠ endonuclease and PmlⅠ endonuclease (New England Biolabs NEB, USA). The recombinant expression vector was transformed into Agrobacterium tumefaciens strain GV3101 through freeze-thaw method.
The strain GV3101, which contained recombinant plasmid, was cultured in Luria Bertani medium, and then resuspended in the Murashige and Skoog (MS) liquid medium to OD600 value of 0.8. The leaf discs (1.0 cm × 1.0 cm) of the aseptic seedlings ‘Jinba’ were pre-cultured on MS + 1 mg/L 6-BA + 0.1 mg/L NAA for 2 d. Subsequently, leaf discs were cultured on MS + 1 mg/L 6-BA + 0.1 mg/L NAA + 200 mg/L cephalosporin for 3 d after immersion in Agrobacterium. Finally, they were transferred to MS + 1mg/L 6-BA + 0.1 mg/L NAA + 200 mg/L cephalosporin + 10 mg/L kanamycin for 2 d. The resistant seedlings rooted on MS + 0.1 mg/L NAA.
Identification of transgenic plants
-
To confirm the integration of the overexpression cassette, we developed a cleaved amplified polymorphic sequence (CAPS) marker according to the description in Feng et al.[59]. Using the same qRT-PCR method mentioned above, the relative transcript levels of CmCC-NB-ARC were measured in all transgenic plants at 48 h after inoculation. After being inoculated for 15 d, the phenotypes of the transgenic plants were compared with the wild type cultivars 'C029' and 'Jinba'. The microscopic investigation of the diseased spots was performed according to the method described by Zhu et al[3].
-
After years of observation, 'C029' plants were robust and without any disease spots on the abaxial leaf surfaces (Fig. 1a). In 'Fenhuaxiaori' plants, the abaxial leaf surface was covered with a large number of milky yellow or pale pink diseased spots, accounting for about four-fifths of the leaf area (Fig. 1b). In 'LZ08-61' and 'Jinba' plants, diseased spots accounted for less than half of the leaf dorsal area (Fig. 1c, d). Thus, the cultivar 'C029' is highly resistant to chrysanthemum white rust, 'Fenhuaxiaori' and 'LZ08-61' are susceptible to this disease, and 'Jinba' is moderately susceptible.

Figure 1.
The phenotype of the abaxial of the leaves of Chrysanthemum in the stage of symptom appearance. (a)−(d) showed the highly resistant cultivar 'C029' and susceptible cultivars 'Fenhuaxiaori', 'LZ08-61', and 'Jinba', respectively. The scale bar represents 1 cm. 0 is immune, represents no visible symptoms; 5 is highly susceptible, represents yellowish necrosis and clear telia were same as the third-class, but telia crowded together, telia was more 1/2 of the whole leaf; 3 is susceptible, represents yellowish necrosis and clear telia on the back, larger and less, or smaller and more, total telia area was no more than 1/4 of the whole leaf.
Identification of the potential gene related to resistance to chrysanthemum white rust
-
R genes containing NBS domains are commonly reported to confer resistance to fungi and oomycetes[35,36,38−43]. Among the five plants, Artemisia annua, Lactuca sativa, and Helianthus annuus all belonged to the same family Asteraceae. The pathogens of wheat leaf rust and chrysanthemum white rust were both Puccinia fungi. The fifth plant was a model plant Arabidopsis thaliana. It is feasible to find a chrysanthemum resistance gene by a blast-search against the chrysanthemum reference database according to the five resistance gene analogs. We compared the sequences of NBS domains in Artemisia annua, Lactuca sativa, Helianthus annuus, Arabidopsis thaliana, and Triticum aestivum, it showed 50.28 % similarity between the CHR00059759 and the conserved domain of the five genes. The P-loop, Kinase-2a, Kinase-3a, and GLPL core motifs were conserved (Fig. 2). Using the conversed sequence among the five plants as a query, we conducted a BLAST search for homologous sequences in the chrysanthemum reference database (
www.amwayabrc.com/zh-cn/index.html ). We achieved several homologous amino acid sequences containing the conserved motifs, among these CHR00059759 showed high similarity with the above NBS domains. Hence, we suspected that CHR00059759 might be a potential gene conferring the resistance to chrysanthemum white rust.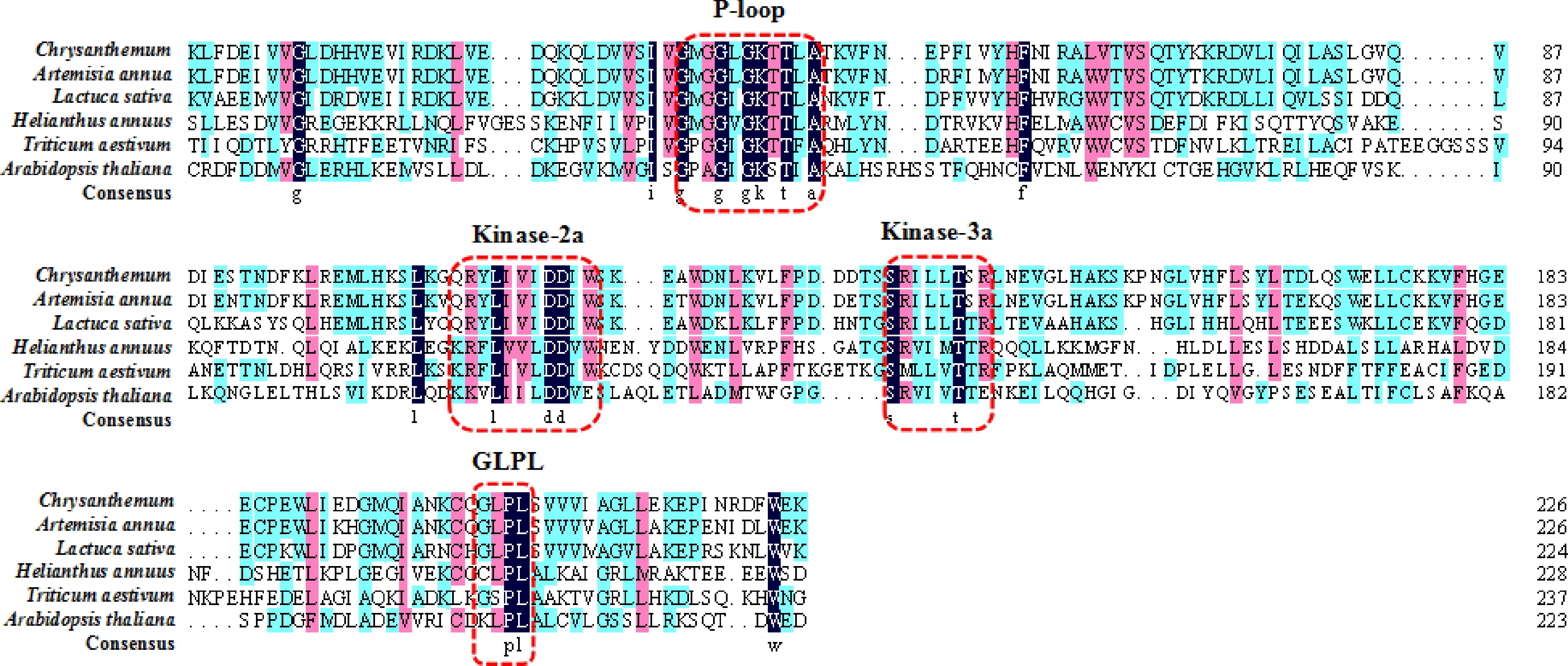
Figure 2.
The NBS conserved domain in different plants. Conserved domains are marked with dashed red lines.
Isolation of CHR00059759 in highly resistant and susceptible chrysanthemum cultivars
-
We isolated the genomic sequences and coding sequences of CHR00059759 from the highly resistant- and susceptible- cultivars (Fig. 3a, b). The open reading frames (ORF) of 'C029', 'Fenhuaxiaori', 'LZ08-61', and 'Jinba' were 1,612 bp, 1,618 bp, 1,615 bp, and 1,615, respectively (Supplemental Fig. S1). The corresponding coding sequences were the same size with Open Reading Frames.
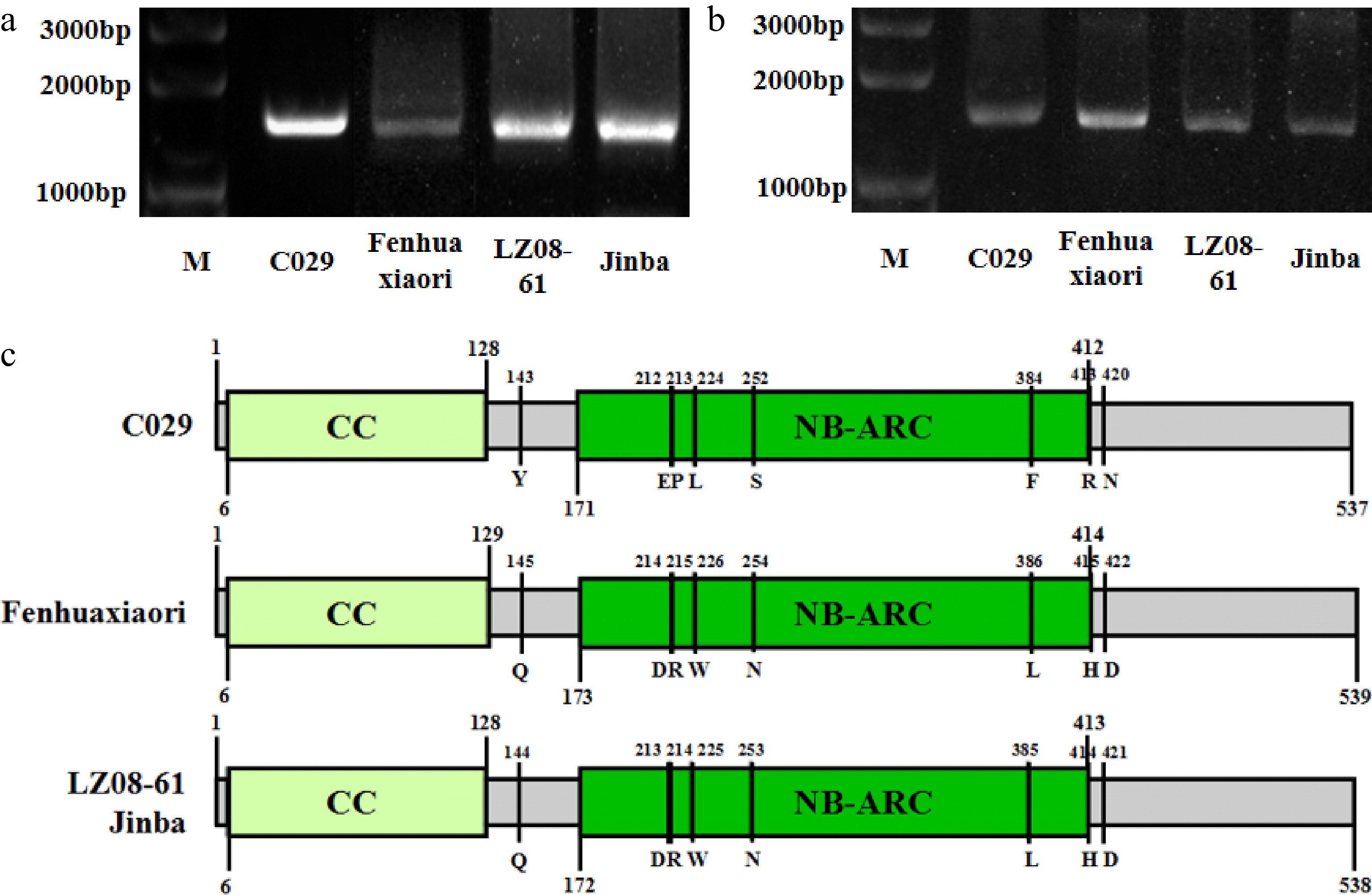
Figure 3.
Amplification and conserved domain prediction of CHR00059759 in highly resistant and susceptible cultivars. (a) PCR amplification of the genomic sequence of CHR00059759 in cultivars 'C029', 'Fenhuaxiaori', 'LZ08-61', and 'Jinba', respectively. (b) Amplification of the coding sequence of CHR00059759 in cultivars 'C029', 'Fenhuaxiaori', 'LZ08-61', and 'Jinba', respectively. M, DNA marker. (c) The structure of CHR00059759 in the four cultivars. Gray boxes indicate exons of CHR00059759 gene. Yellow and green indicate the CC and NB-ARC conserved domain, respectively. Several unique variations between the highly resistant- and susceptible- cultivars were omitted.
Comparing the coding sequence of CHR00059759 to the NCBI Conserved Domain Database indicated that the gene belongs to the RX-CC_like superfamily and contains an NB-ARC conserved domain. We therefore named the gene CmCC-NB-ARC. Compared with cultivar C029, cultivar Fenhuaxiaori had 111 SNP variations, including 38 nonsynonymous mutations raised by 49 SNPs, and cultivars LZ08-61 and Jinba had 29 bp variations including 10 nonsynonymous mutations raised by 12 SNPs (Supplemental Fig. S1). Among these differences, there were 17 coincident different SNP sites between the highly resistant cultivar and three susceptible cultivars, among which nine SNP differences resulted in nonsynonymous mutations of eight common amino acids (Supplemental Fig. S2). There was no difference in the CC domain. There are five coincident differences in the NB-ARC domain: glutamic acid (E), proline (P), leucine (L), serine (S), and phenylalanine (F) in the resistant cultivar C029 changed to aspartic acid (D), arginine (R), tryptophan (W), asparagine (N), and leucine (L), respectively in the susceptible cultivars (Fig. 3c). Several studies showed that the mutations occurred in conserved domain may response to resistance of R genes[37−40]. According to the variations in the NB-ARC conserved domain among the highly resistant and susceptible cultivars, we speculate that the gene might be resistant to chrysanthemum white rust.
Phylogenetic tree of CmCC-NB-ARC
-
To better elaborate the structure and evolution of CmCC-NB-ARC and the relationships among its homologous proteins in other 15 species, we constructed a phylogenetic tree using the amino acid sequences of Chrysanthemum morifolium and 15 homologous proteins (Fig. 4). CmCC-NB-ARC had a certain degree of conservation relative to the homologous proteins in other species, indicating that the function of this protein was conserved among different plants. In Arabidopsis and other plants, the NB-ARC domain confers resistance against pathogens[33,34]. Therefore, we speculated that the CmCC-NB-ARC might have resistant function.

Figure 4.
The phylogenetic tree of the CmCC-NB-ARC. The phylogenetic tree was constructed by the neighbor-joining method[[57]]. The percentage of replicate trees in which the associated taxa clustered together in a bootstrap test (1000 replications) is shown next to the branches[[58]]. In the guided test (1000 replicates), the percentage of replicate trees with related taxa clustered together is shown next to the branch. The evolutionary distances were computed using the Poisson correction method and given in units of amino acid substitutions per site[[61]]. All ambiguous positions for each sequence pair were removed.
Relative expression of CmCC-NB-ARC in plants inoculated with P. horiana
-
We compared the expression of CmCC-NB-ARC in leaves of highly resistant cultivar 'C029' after fungus inoculation at different stages, using sterile water as the control. The same method was used in the susceptible cultivar 'LZ08-61', either. Compared with controls, the relative expression level of CmCC-NB-ARC decreased slightly at 6 h after inoculation both in 'C029' and 'LZ08-61' (Fig. 5a, b). The expression level increased at 12 h, decreased significantly at 18 h, and then continuously increased exponentially at 24, 36, 48, and 72 h in 'C029' (Fig. 5a). However, in 'LZ08-61', the relative expression level decreased at 12 h, increased at 18 h and 24 h, peaked at 36 h, then gradually decreased at 48 h and 72 h (Fig. 5b). Combined with the difference in relative expression level bwtween the highly resistant cultivar 'C029' and the susceptible cultivar 'LZ08-61', the variation in expression at different time points suggests that CmCC-NB-ARC expression varied in response to the infection with the fungus. At 30 d after inoculation, the abaxial surfaces of leaves of cultivar 'C029' showed no disease spots (Fig. 5c), but disease spots appeared on those of leaves of cultivar 'LZ08-61' (Fig. 5d), suggesting that the infection cycle of the fungus spores had been completed. In consequence, we infer that CmCC-NB-ARC made a response to chrysanthemum white rust.
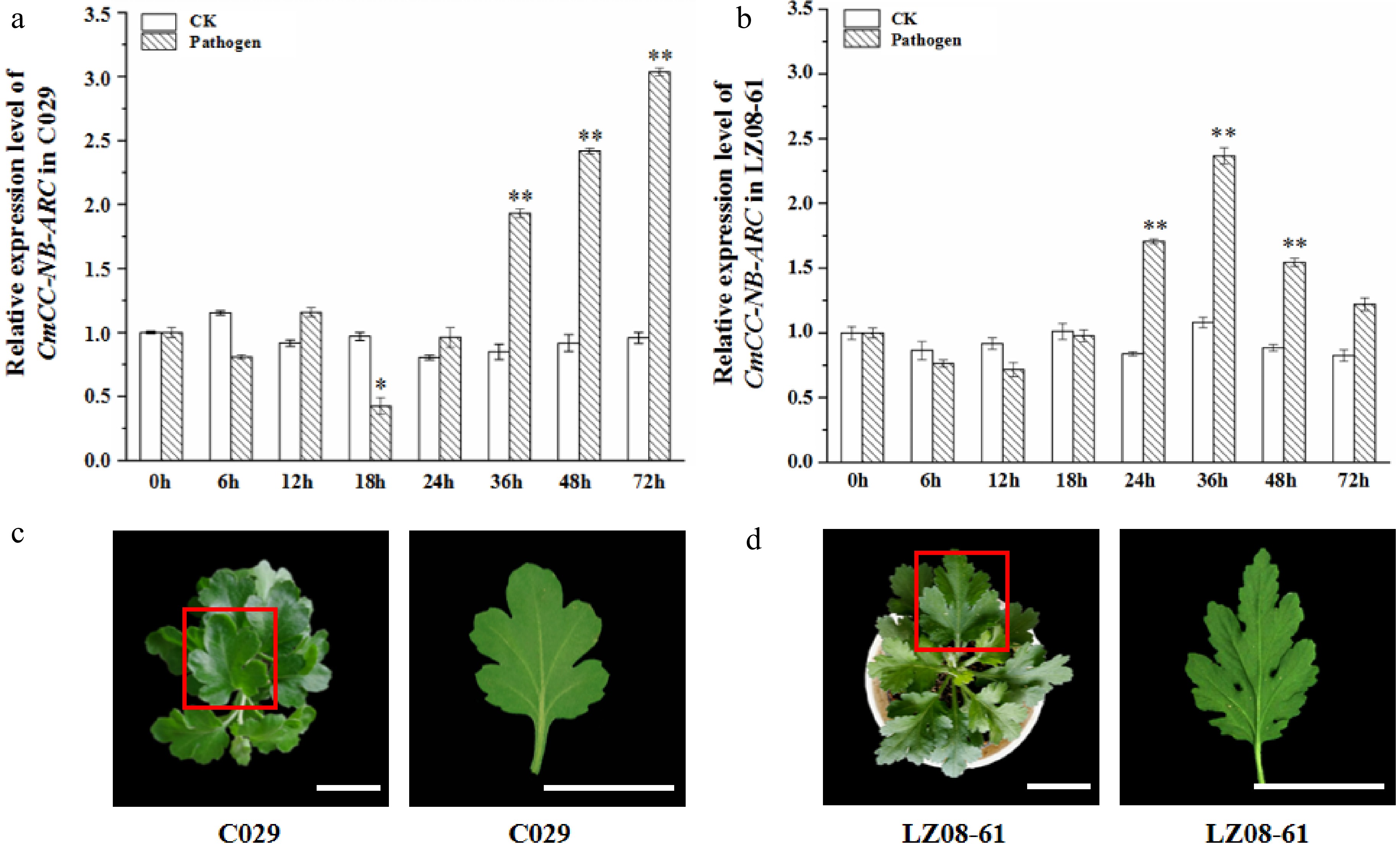
Figure 5.
Transcription profiling of CmCC-NB-ARC and phenotype of the highly resistant cultivar 'C029' and susceptible cultivar 'LZ08-61' at one month after inoculation with P. horiana. (a), (b) Relative expression level of CmCC-NB-ARC in the highly resistant cultivar 'C029' and susceptible cultivar 'LZ08-61'. The leaf under inoculation for 0 h (before inoculation) is the control. CmACTIN served as an equal loading control. The error bars represent standard errors of three biological replicates. The asterisks on the error bars represent significant differences at the same time point of both under inoculation of with the fungus resuspended in buffer and sterile water. Asterisks represent significant differences using a t-test with three repeats (* p < 0.05, ** p < 0.01). (c), (d), Phenotypes of 'C029' and 'LZ08-61' plants at one month after inoculation with P. horiana. The red box represents the single leaf which was used for morbidity observation. Scale bar = 3 cm.
Overexpression of the gene CmCC-NB-ARC in susceptible cultivar Jinba
-
To verify the function of the gene CmCC-NB-ARC, an overexpression vector was constructed containing the coding sequence of the resistant cultivar 'C029' (Fig. 6a, b). We transferred the recombinant vector 35Spro:CmCC-NB-ARC into the susceptible cultivar Jinba and achieved three transgenic individuals with Agrobacterium-mediated genetic transformation procedure (Fig. 6c−g).
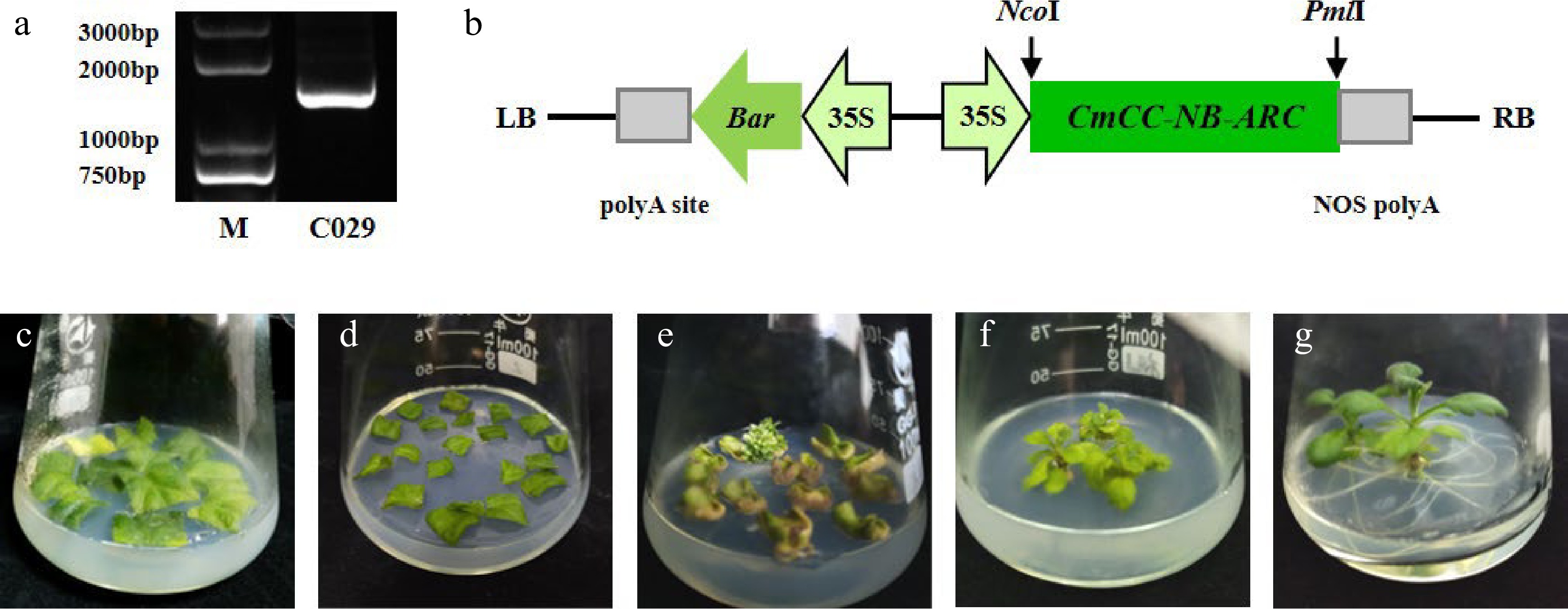
Figure 6.
Construction of the recombinant overexpression vector and genetic transformation. (a) The specific fragment of CmCC-NB-ARC in 'C029' on the agarose gel. (b) Schematic of overexpression cassette of the gene. (c)–(g) The procedure of Agrobacterium-mediated transformation. (c) Leaf discs were pre-cultured. (d) Leaf discs were cultured after being immersed with Agrobacterium. (e) Regenerated kanamycin-resistant buds. (f) Rooting. (g) Seedling.
In order to confirm the presence of the overexpression cassette in transgenic plants, we developed a cleaved amplified polymorphic sequence (CAPS) marker according to the sequence differences of the gene CmCC-NB-ARC between 'C029' and 'Jinba'. The 271-bp PCR product was amplified among the two wild types and the three transgenic strains by the primers F: 5’-AGAAGCAGCTAGATGTCGTCT-3’ and R: 5’-TCAATGACAATCAAATACCTC-3’. As expected, the product was digested by restriction endonuclease MboⅠ into two fragments 194bp and 74bp both in the three transgenic plants and the susceptible wild type 'Jinba'. Whereas the 271bp in the resistant cultivar 'C029' was not digested. So the lack of the restriction enzyme cutting sites in ‘C029’ might be the reason (Fig. 7a). Above all, the coding sequence of CmCC-NB-ARC from 'C029' was successfully integrated into 'Jinba'.
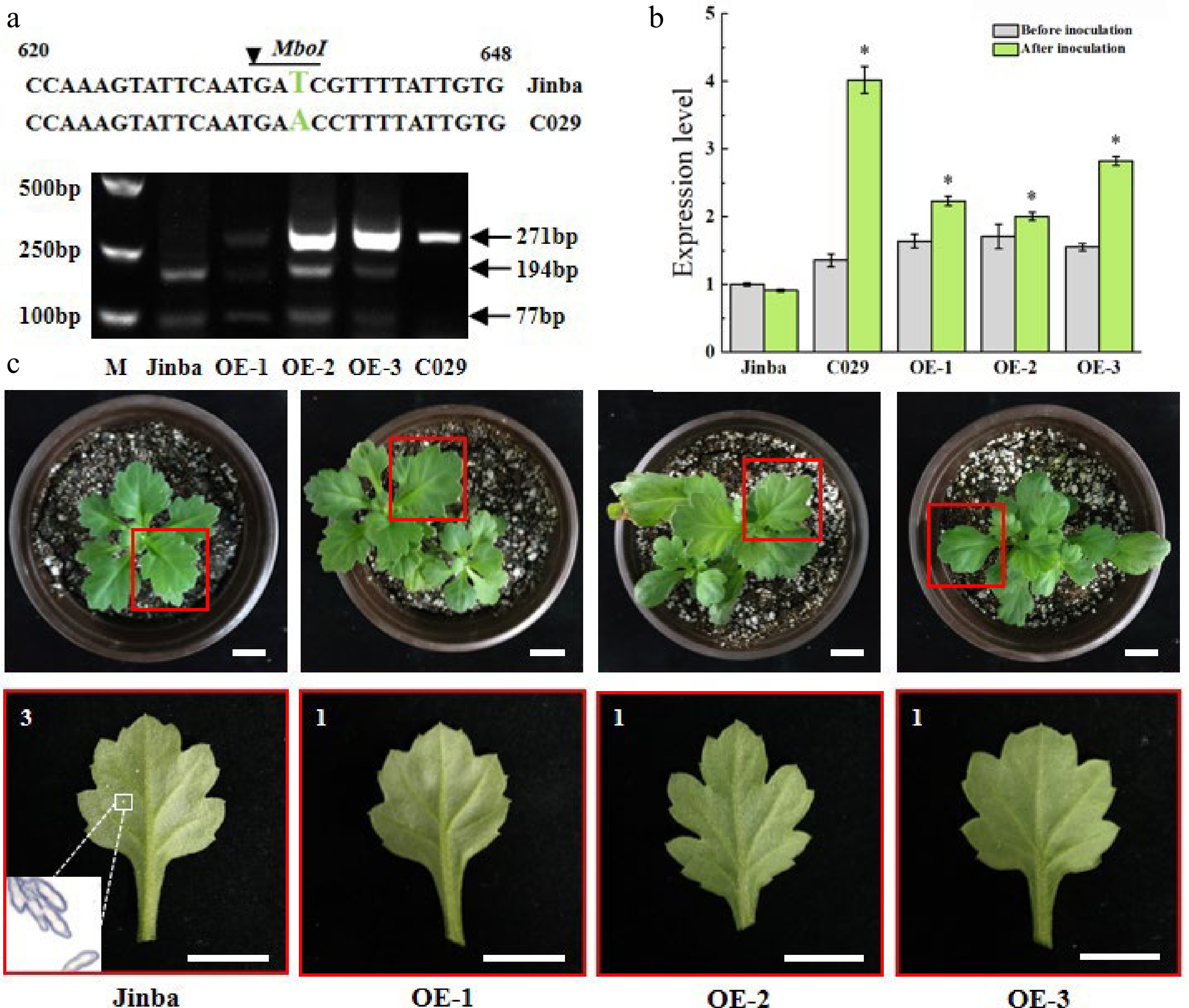
Figure 7.
Characterization of overexpressing plants. (a) The specific fragments indicate the digested PCR products by restriction enzyme MboI in the two wild types and the three transgenic individuals, respectively. Fragments 271bp represent the size of the amplified product without the cutting site of restriction enzyme. Fragments 194bp and 77bp represent the size cleaved by the enzyme. (b) Relative expression levels of CmCC-NB-ARC in the two wild types and the transgenic individuals before inoculation and at 48 h after inoculation, respectively. Asterisks represent a significantly different expression level to that of ‘Jinba’, as determined using a t-test with three repeats. (*p < 0.05). (c) Phenotypes of the transgenic lines at 15 d after inoculation. The red box indicates leaves were selected to be observed in the potted plant. The white box indicates the telia from diseased spots under a microscope of 200×. Scale bars = 2 cm. 3 is susceptible, represents yellowish necrosis and clear telia on the back, larger and less, or smaller and more, total telia area was no more than 1/4 of the whole leaf. 1 is highly resistant, represents rare visible whitish or yellowish necrotic or chlorotic lesions are discernible.
Furthermore, we detected the relative expression levels of CmCC-NB-ARC in the transgenic lines at 48 h after inoculation (Fig. 7b). The relative expression levels were 2.454-fold, 2.205-fold and 3.102-fold higher than Jinba in OE-1, OE-2 and OE-3, respectively. The differences between the transgenic lines and the susceptible cultivar 'Jinba' were statistically significant, whereas there was no difference between the three transgenic plants and the resistant cultivar 'C029'.
Finally, we inspected the phenotypes of the transgenic individuals and wild types after inoculated by the fungus in 15d. We did not find any diseased spots at the abaxial leaf surface of the three transgenic individuals. Nevertheless, several milk white diseased spots appeared distinctly on several leaves of the wild type 'Jinba' (Fig. 7c). The telia of the diseased spot of 'Jinba' can be observed clearly under microscope, while we did not detect teliospore in the three transgenic lines. In summary, we could speculate that the gene CmCC-NB-ARC harbored the resistance to P. horiana.
-
Chrysanthemum white rust is an obligate parasitic fungus that seriously harms the chrysanthemum industry. In this study, we identified cultivar 'C029' as highly resistant to chrysanthemum white rust and cultivars 'Fenhuaxiaori', 'LZ08-61', and 'Jinba' as susceptible over several years’ observation. We found the gene CHR00059759 by BLAST-searching the typical NBS conserved domain in the chrysanthemum reference genome database. The gene comprised a conserved CC-NB-ARC domain, and there were eight coincident nonsynonymous amino acid mutations among the highly resistant and three susceptible cultivars, including five in the NB-ARC conserved domain. The expression level of this gene was different between cultivars 'C029' and 'LZ08-61' after inoculation with chrysanthemum white rust. The overexpression experiments verified the gene CmCC-NB-ARC harboring the resistance to chrysanthemum white rust.
Most plant R genes pertain to the NB-LRR superfamily, containing a coiled-coil (CC) domain that acts as a 'switch' that recognizes the exogenous stress signal[62]. The N-terminal TIR and CC domains are involved in the formation of homo-dimers, which are necessary for activating defense signals[29−32]. The NB-ARC domain acts as a nucleotide-binding pocket to hydrolyze Adenosine triphosphate (ATP) and induce the conformational change of R protein[27,31]. Conserved motifs in the NB-ARC domain, including the P-loop, RNBSA to D and methionine-histidine-aspartate (MHD) motifs, play an important role in controlling R gene activation[27,28]. Genes comprising NB-ARC domains were reportedly involved in defense against fungi or oomycetes, such as Puccinia striiformis f.sp tritici in wheat[63], Fusarium oxysporum in chickpea[41], and Phytophthora infestans in tomato[64].
A gene encoding a CC-NBS-LRR protein identified by BLAST-searching the rice sequence was isolated by map-based cloning. The P-loop in the conserved domain was missing and other partial motifs degenerate, and these deficiencies may lead to abnormal resistance[38]. In leaf spot of cucumber, the gene with a nonsynonymous SNP in the NB-ARC domain may alter the function of the conserved NB-ARC motif[39]. In this study, we identified mutations in the NB-ARC domain between highly resistant and susceptible cultivars, but the core motifs were conserved. We speculate that the mutations in the conserved domain might cause differences in resistance to the chrysanthemum white rust in different cultivars.
To defend to exogenous pathogenic invasion, R genes would contend with the pathogies, hence the expression will be along with the infected degree. The relative expression of CC-NB-LRR gene in white rust resistance of Brassica juncea did not change significantly at different time points after infection. Its resistance was expressed at the pre-infection mRNA levels[40]. In our study, the relative expression of CmCC-NB-ARC decreased slightly 6 h after inoculation in 'C029' and 'LZ08-61' plants inoculated with P. horiana compared with controls. Over the next six time points, gene expression showed an increase-decrease-increase trend at 12 h, 18 h, and 24 h − 72 h in ‘C029’, but a decrease-increase-decrease trend at 12 h −18 h, 24 h − 36 h, and 48 h − 72 h in 'LZ08-61'. The expression pattern closely reflected the chrysanthemum white rust invasion pattern. We observed a telium in susceptible culture seedlings, and marked disease spots could be seen on the surface of the adaxial leaves of 'LZ08-61' seedlings, which is the same pathogenesis as observed in previous studies[3,6].
In previous studies, chrysanthemum white rust is usually classified into six levels from 0 to 5 according to the resistance, among them 0−2 and 3−5 mean resistance and susceptiblity, respectively[3, 13−17]. The similar several levels classification method was also used in wheat leaf rust caused by Puccinia recondita[65]. Based on the isolation of CmCC-NB-ARC gene, the coding sequences could be clearly divided into resistant- and susceptible groups. Different resistance levels in the same group may be caused by gene regulation. R genes have been identified in a range of plants by genome sequencing[36,38] and RNA sequencing[43,64]. In future work, we aim to further explore the R genes responding to chrysanthemum white rust by RNA sequencing to elucidate the underlying molecular mechanism.
Data availability
-
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
This research was supported by the National Key R&D Program of China (2018YFD1000400).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Lin Jiang, Xin Feng, Xinyue Chen
- Supplemental Fig. S1 The open reading frames of cultivars 'C029', 'Fenhuaxiaori', 'LZ08-61', and 'Jinba'
- Supplemental Fig. S2 The amino acid sequences of cultivars 'C029', 'Fenhuaxiaori', 'LZ08-61', and 'Jinba'The red brackets represent the CC domain. The yellow brackets represent the NB-ARC domain.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Jiang L, Feng X, Chen X, Yu Y, Mao H, et al. 2023. Cloning and identification of CmCC-NB-ARC, a chrysanthemum white rust resistance gene. Ornamental Plant Research 3:7 doi: 10.48130/OPR-2023-0007 

Cloning and identification of CmCC-NB-ARC,
- Received: 29 September 2022
- Accepted: 21 February 2023
- Published online: 03 April 2023
Abstract: Chrysanthemum are important perennial commercial and ornamental flowers worldwide. Chrysanthemum white rust caused by the fungus Puccinia horiana is a destructive disease. Recent studies have aimed to identify sources of resistance to this disease, whereas genes related to the resistance to the fungus were rarely reported. In this study, we used highly resistant and susceptible chrysanthemum cultivars whose disease sensitivity was confirmed over years of observation. Using a conserved nucleotide binding site (NBS) domain sequence that is associated with resistance to rust in other plants as a query in a BLAST search against the chrysanthemum reference database, we identified the gene CHR00059759. There were eight consistent nonsynonymous mutations between the highly resistant and susceptible cultivars, five of which occurred in the NB-ARC conserved domain. The expression level of CmCC-NB-ARC increased and then decreased in the susceptible cultivar 'LZ08-61', whereas decreased and then increased in the resistant cultivar 'C029' in the eight time-points after inoculation with the fungus. In order to verify the gene function, we constructed an overexpression vector 35Spro:CmCC-NB-ARC and transformed into the susceptible cultivar 'Jinba'. Compared with 'Jinba', the relative expression levels of the gene in the transgenic plants increased significantly at 48h after inoculation. Compared with three overexpression lines, the diseased spots appeared in 'Jinba' at 15d after inoculation. Hence, we speculated the gene CmCC-NB-ARC conferred resistance to chrysanthemum white rust. This study provides insight into the mechanism of resistance to chrysanthemum white rust and might help inform the breeding of resistant cultivars.
-
Key words:
- CC-NB-ARC conserved domain /
- Chrysanthemum /
- Puccinia horiana /
- Overexpression