








-
The genus Pholiota (Basidiomycota) was proposed by Kummer[1] in 1871 for taxa having yellow or brown pileus with scaly and dry to slimy caps, smooth spores having a small germ pore at the apex, and rusty-brown to yellow-brown spore deposits[2−4]. It is a well-defined genus with about 157 species recorded worldwide[4]. The type species of this genus is Pholiota squarrosa (Vahl) P. Kumm.[2,3] . The species of Pholiota are saprophytic that rely mainly on dead wood and play a significant role as wood decomposers in the forest ecosystem[4]. It is also regarded as the major genus of wood − rotting agarics having colored spores[2].
A major feature of Pholiota is the diversity in cystidial content, the presence or absence of pleurocystidia and cheilocystidia, wall thickness, and coloring, all of which have been utilized as important features to distinguish various Pholiota species[2,4]. However, depending on the prevalent environmental factors, macro-morphological characteristics of Pholiota species can vary, even among the same species, so morphological characteristics alone are insufficient to distinguish the species[2,4]. In order to properly identify fungal species, further identification using molecular analysis has taken the lead[4,5].
The Pholiota species have been assigned to the Strophariaceae family based on phylogenetic analyses but also form a paraphyletic clade with Hypholoma and Stropharia species[3−4]. The genus Pholiota has been subdivided into a number of subgenera, however in-depth DNA analyses of the genus are still needed to ascertain the phylogenetic relation within the Pholiota[3,6]. Pholiota has been well examined and documented from North America by Overholts[7] and Smith & Hesler[1].
Complete documentation of Pholiota species from India has not yet been recorded[6]. Only 21 species of Pholiota have been reported from India[6, 8−12]. A detailed morphological and molecular study revealed the collected sample as P. multicingulata Horak, which has not been previously reported from India and is presented here.
-
Fresh fruiting bodies of the specimen were collected from the Kalimpong District, West Bengal, India, in May 2018. The specimen was photographed in the field using a digital camera and macro-morphological details of the collected specimen were noted before drying. Methuen Handbook of color[13] was followed for color terminology. To observe the micro-morphological features, free-hand sections were prepared from the dried basidiocarp and mounted in a 5% aqueous KOH solution and the section were examined under a compound microscope after being stained with Congo red. At least 30 basidiospores were measured per specimen and mean values were provided. Q value refers to the length/width ratio of individual basidiospore; Qm represents the mean of Q value. The voucher specimen examined was preserved[14] and added to the Calcutta University Herbarium (CUH) with the accession number CUHAM763.
DNA extraction, PCR amplification and sequencing
-
Genomic DNA was isolated from dried basidiocarp using the XcelGen Fungal gDNA Kit (Xcelris Genomics, Ahmedabad, India) following the manufacturers protocol. The nuclear ribosomal internal transcribed spacer (nrDNA ITS) region was amplified using the primer pair ITS1 and ITS4[15,16]. PCR amplification of desired region was carried out according to Dutta et al.[17]. The QIAquick® Gel Extraction Kit was used to purify the amplified PCR products before they were used for automated DNA sequencing on the ABI3730xl DNA Analyzer (Applied Biosystems, USA) with a primer that was identical to that used for amplification of the nrDNA region. The chromatograms of freshly generated sequences were examined and manually modified using Bioedit sequence alignment editor version 7.0.5 software[18]. The BLASTn tool were used (NCBI) to compare the query sequence obtained in this study to homologous sequences in GenBank[19]. The newly generated sequence was submitted with the accession number OM428169 into the GenBank nucleotide database (
www.ncbi.nlm.nih.gov ).Phylogenetic analyses
-
The newly generated sequence along with the sequences obtained from GenBank by BLAST search were used for conducting the phylogenetic analyses (Table 1). The Agrocybe species was chosen as outgroup taxa following, Lee et al.[4]. A final dataset of 38 nrDNA ITS sequences was aligned using MUSCLE[20] and further manual alignment correction was carried out in MEGA v.7.0[21]. An appropriate model of sequence evolution for phylogenetic analysis was performed using the best-fit substitution model (TPM2uf+G with BIC of 9282.555780) from jModeltest 2.1.10 v.20160303 (Darriba et al.[22] in the CIPRES web portal (
www.phylo.org/portal2 ). Maximum likelihood bootstrapping (MLBS) analysis was performed using RAxMLv.8.2.12[23] with 1000 bootstrap replicates using the default parameters as implemented on the CIPRES NSF XSEDE resource. MrBayes v. 3.2.7[24] software was employed to perform Bayesian analysis (BI) using Markov chain Monte Carlo (MCMC) methods[25] based on the TPM2uf+G substitution model. The Markov chain was run with default parameters as described by Vishal et al.[26] for 106 generations. Phylogenetic trees were opened and edited in FigTree v.1.4.4[27]. The resultant tree displays the maximum likelihood bootstrap (MLBS) and Bayesian posterior probabilities (PP) values over 50% and 0.50.Table 1. Names, voucher numbers, geographic origins and GenBank accession numbers of the taxa used in the phylogenetic analyses.
Species Voucher no. Geographic origin GenBank accession no. Pholiota multicingulata CUHAM763 India OM428169 Pholiota multicingulata SFC20180907-142 South Korea MT879450 Pholiota multicingulata Strain 1215 New Zealand MH409971 Pholiota multicingulata TENN:074783 (HMJAU37414) China MN209761 Pholiota multicingulata STDS-1-6 Japan LC098735 Pholiota terrestris UC 1859859 USA KC122896 Pholiota terrestris KA15_0175 South Korea MT626081 Pholiota terrestris SFC20151120-02 Korea KX773888 Pholiota microspora CBS 360.51 Japan MH856901 Pholiota microspora NIBRFG0000103779 South Korea MT626085 Pholiota abietis SFC20121009-35 Korea KJ609166 Pholiota abietis − China JF961360 Pholiota adiposa CBS 279.29 Canada: Ontario MH855073 Pholiota adiposa isolate ZBH(6) China HQ436122 Pholiota aurivella CBS 118.18 Germany MH854669 Pholiota aurivella CBS 262.32 Netherland MH855317 Pholiota squarrosoides TENN61728 USA FJ596877 Pholiota squarrosoides TENN61692 USA FJ596859 Pholiota limonella SFC20150707-19 Korea KX773882 Pholiota limonella KUC20130923-06 Korea KM496470 Pholiota spumosa voucher 3533 Italy JF908577 Pholiota spumosa voucher 5509 Russia MH930222 Pholiota spumosa PRM:897147 Czech Republic HG007981 Pholiota lubrica PRM:857179 Slovakia HG007984 Pholiota lubrica NBRC 32453 Japan AB301612 Pholiota lenta R. Tuomikoski s.n. (H, IBUG) Finland AY281022 Pholiota lenta 7100 Italy JF908582 Pholiota squarrosa PBM 2735 (CUW) Colorado USA DQ494683 Pholiota squarrosa − Germany: Leer FR686575 Pholiota scamba 13482 Italy JF908585 Pholiota chocenensis − Czech Republic HG007985 Pholiota highlandensis Mushroom Observer 366219 USA MN832738 Pholiota highlandensis MES-2478 Chile MH930366 Pholiota brunnescens TENN:074791 (HMJAU37361) China MN209732 Pholiota brunnescens TENN:074782 (HMJAU37363) China MN209733 Pholiota brunnescens TFB5897 USA KF871789 Agrocybe pediades PBM 2080(WTU) California DQ484057 Agrocybe praecox PBM 2310 USA AY818348 Agrocybe smithii PBM 2298 Washington DQ484058 Phylogenetic analyses
-
Phylogenetic analyses of chosen species were performed using the combined ITS nrDNA dataset of 38 sequences to evaluate the uniqueness of collection in relation to the existing Pholiota species. Bayesian analyses after 106 generations attained a standard deviation of split frequencies of 0.008223 and the primary 25% of recovered trees were considered as the burn-in. The topology of the phylogenetic tree obtained from the Bayesian analysis and the Maximum Likelihood (ML) analysis was identical. Hence, the phylogenetic tree generated using Bayesian analysis has been displayed in Fig. 1.
The phylogenetic analyses based on nrITS region sequence data clustered the present Indian collection along with the sequence of the same taxon reported from Korea, USA, Japan, and New Zealand with significant support value (97% BS and 1.00 PP).
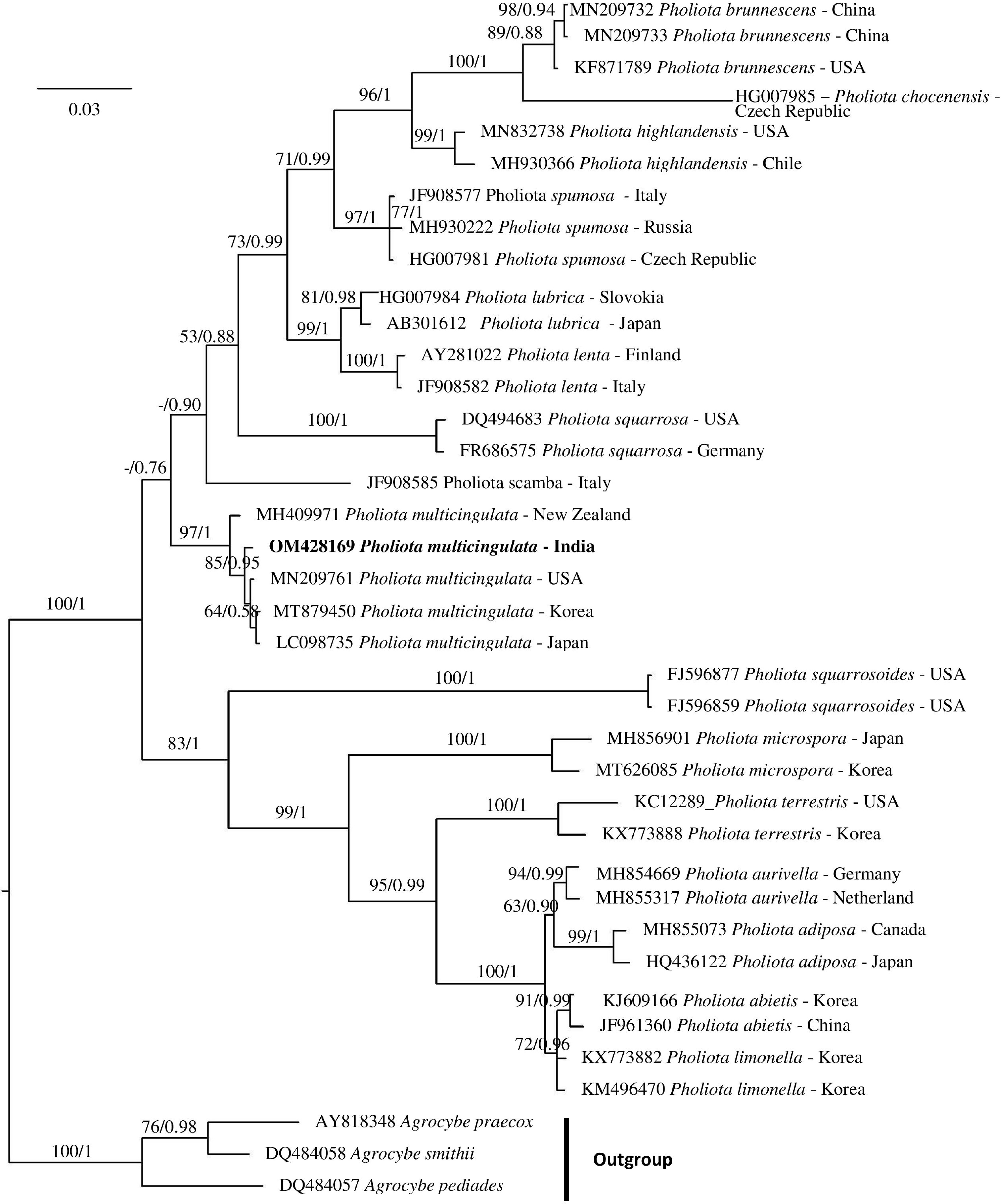
Figure 1.
Phylogenetic analyses of P. multicingulata inferred from nrITS sequences. ML bootstrap percentages (MLBS) are shown to the left of /, and Bayesian posterior probabilities (PP) are shown to the right. MLBS values ≥ 50% and PP values ≥ 0.50 are displayed at the nodes. Collection from West Bengal, India is shown in bold. Scale bar depicts the expected changes per site.
Taxonomy
-
Pholiota multicingulata Horak, Aust. J. Bot. Suppl. 10: 33 (1983) Figs 2 & 3

Figure 2.
Pholiota multicingulata Horak. (a)−(d) Fresh basidioma in field. Scale bars = 10 mm.
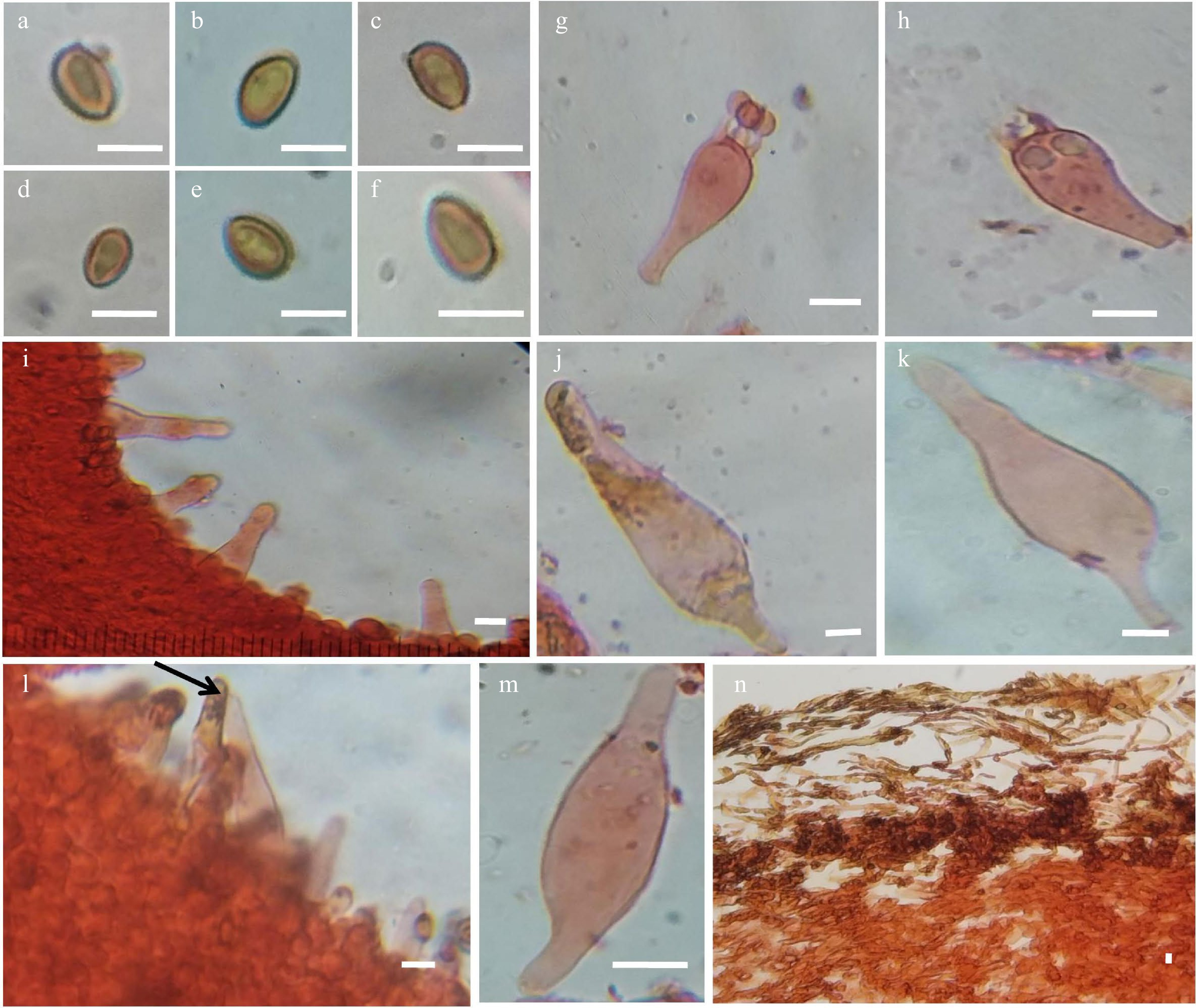
Figure 3.
Pholiota multicingulata Horak. (a)−(f) Basidiospores. (g), (h) Basidium. (i)−(k) Pleurocystidia. (l), (m) Cheilocystidia. (n) Pileipellis. Scale bars = 10 μm
Pileus 17–34 mm. broad, convex when young, subumbonate becoming plano-convex as it matures; surface viscid when moist, with concentric rings of broad appressed scales, variable in color, burnt sienna (7D8) when young to brick red (7D7) at maturity, center reddish-orange(7B8), margin incurved to plane when mature. Lamellae adnate, regular, greyish white to grey (1B1–4B1), close with lamellulae of 4-tier. Stipe 32 mm × 4 mm, central, multiplezonate, with brown fibrillose zone below the veil, well developed, cylindric, hollow with rhizoids at the base. Partial veil present in the young stage.
Basidiospores 6.3–8.5 × 4.8–5.9 µm (Lm = 7.8, Wm = 5.5); Q = 1.27–1.5, Qm = 1.42, smooth, thick-walled, ellipsoid in shape, germ pore minute and inconspicuous. Basidia 4-sterigmate, 21.98–28.26 × 8.47–9.42 µm, clavate or cylindro-clavate. Pleurocystidia 53.38–69.08 × 12.56–18.84 µm, abundant, projecting prominently, fusiform-lageniform, hyaline or with yellow-brown colloidal content or with yellow-brown plug-in neck area, crystalline cap over apex, pedicel long or short. Lamellae-edge sterile. Cheilocystidia 53.38–56.52 × 12.56–15.4 µm, similar to pleurocystidia in shape and content. Subhymenium layer of gelatinized hyphae. Pileipellis, a well-developed layer consisting of incrusted repent hyphae withbrown pigments in the wall, 5.4–6.3 µm broad. Clamp connections present in all the tissue.
Habit, habitat, phenology and distribution - Gregarious on leaf litter of bamboo (Dendrocalamus sp.) and rotten wood chips in Kalimpong District, West Bengal India. May – September. Distribution reported from New Zealand, Australia[28] and Korea[3].
Specimen examined - India, West Bengal State, Kalimpong District, 16th mile, Algarah, elevation 1780 m, 27 May 2018, J. Tamang (Coll.), JT-58/2018 (CUH AM763).
-
Pholiota multicingulata, is distinguished by brown sienna to brick red pileus that is darker at the disc and has concentric rings of appressed scales on the surface, multizonate stipe, pleurocystidia and cheilocystidia with yellow-brown colloidal content, amorphous cap over apex, and basidiospores measuring 6.3–8.5 × 4.8–5.9 µm in diameter and having a mean Q value of 1.42.
The macro- and micro-morphological characteristics of our Indian sample show similarity with the type specimen description[28], with the exception for slight variation in spore size (6.3– 8.5 × 4.8–5.9 µm vs 6.5–8 × 4.5–5 µm) and pileus size (17−34 mm vs 55 mm). The specimen reported from Korea differs by having a bit variation in basidiospores size (6.3–8.5 × 4.8–5.9 µm vs 7.3–8.2 × 4.7–5.4 µm) and pileus color (burnt sienna to brick red vs cream to cocoa cream). This variation may occur due to climatic and geographical variations.
The present taxon has been previously reported from New Zealand, Australia[28], and Korea[3]. However, it has not been reported from India prior this study. Among morphologically related species, the Indian collection shares certain gross morphological and microscopic features with P. scamba[3]. However, it differs in having ventricose-shaped pleurocystidia and cheilocystidia .
-
The authors declare that they have no conflict of interest.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Tamang J, Thapa A, Acharya K. 2023. New record of Pholiota multicingulata (Strophariaceae) from India based on morphological data and phylogenetic analyses. Studies in Fungi 8:4 doi: 10.48130/SIF-2023-0004 

New record of Pholiota multicingulata (Strophariaceae) from India based on morphological data and phylogenetic analyses
- Received: 22 October 2022
- Accepted: 11 January 2023
- Published online: 31 January 2023
Abstract: Pholiota multicingulata is reported for the first time from West Bengal, India. Detailed morphological descriptions, color photographs, and phylogenetic trees to show its position within species of Pholiota and comparisons with morphologically similar species are provided.
-
Key words:
- New record /
- nrITS sequence /
- phylogeny /
- taxonomy