








-
Muyocopronaceae was validly introduced by Hyde et al.[1], with a type genus Muyocopron Speg. The family currently comprises nine genera (viz. Arxiella Papendorf, Leptodiscella Papendorf, Mycoleptodiscus Ostaz., Muyocopron Speg., Neocochlearomyces Pinruan, Sommai, Suetrong, J.Z. Groenew. & Crous, Neomycoleptodiscus Hern.-Restr., J.D.P. Bezerra & Crous, Paramycoleptodiscus Crous & M.J. Wingf., Pseudopalawania Mapook & K.D. Hyde, and Setoapiospora Mapook & K.D. Hyde), based on molecular phylogeny and morphology[2−7]. Interestingly, members of Muyocopronaceae have been reported to have the potential for antimicrobial and cytotoxic activities[7−10]. Pseudopalawania siamensis polyketide-derived secondary metabolite produced with the potential of antimicrobial and cytotoxic activities. This comprised a new heterodimeric bistetrahydroxanthone, namely pseudopalawanone[7]. An endophytic fungus, Muyocopron laterale (ECN279), which was isolated from a healthy leaf of Canavalia lineata (Fabaceae), produced two new azaphilones, namely muyocopronones A and B with weak antibacterial activity[10]. An endophytic species, Mycoleptodiscus indicus, which is currently named Muyocopron sahnii[3], produced a new triterpenoid[9] and three new azaphilones, namely mycoleptones A, B, and C with four known polyketides. All compounds were weakly active when tested in antileishmanial and cytotoxicity assays[8]. However, the study of secondary metabolite production from Muyocopronaceae is still in the initial stages.
Muyocopron is a diverse genus which can be found in tropical and temperate regions, occurring as saprobes and pathogens on various plant parts such as dead aerial twigs, branches, stems and leaves and can also be an opportunistic pathogen on humans and animals[3, 11−13]. A mycoleptodiscus-like asexual morph has been described for the genus with several additional species based on molecular analyses[3]. The asexual morphs are characterized by irregular sporodochium-like conidiomata, globose or broadly ellipsoidal to ampulliform with enteroblastic, monophialidic conidiogenous cells, and fusiform or fusoid-ellipsoid, curved, hyaline, aseptate or septate conidia with terminal and/or lateral appendages, with or without dark brown appressoria[3]. Presently, 68 species epithets are listed in Index Fungorum[14] with 56 probable species listed in Species Fungorum[15].
In this study, we provide morphological descriptions and illustrations of a new collection of Mu. dipterocarpi from Zanthoxylum fagara, in northern Thailand. The identification was confirmed using both multigene analyses and morphological comparisons. The results of preliminary screening for antimicrobial activity is also provided. The lifestyle and function with the potential for secondary metabolites production are also discussed. In addition, a neotype is designated here for Mu. dipterocarpi due to the original material no longer existing.
-
Fresh material was collected from Phrae Province (India) in September 2016. The micromorphology was examined following the methodology as described by Mapook et al.[6]. Single spore isolation and culture morphology were obtained following the methods of Senanayake et al.[16]. Germinated spores were observed on MEA media (malt extract agar: 33.6 g/l sterile distilled water, Difco malt extract) within 24 h and transferred to new MEA media and incubated at room temperature (25 °C) in the dark. Pure cultures were used for molecular study and pre-screening tests for antimicrobial activity. The specimens with collection details and living cultures are deposited in the Herbarium of Mae Fah Luang University (Herb. MFLU) and Culture collection Mae Fah Luang University (MFLUCC), Chiang Rai, Thailand.
Preliminary screening of antimicrobial activity
-
Preliminary screening of antimicrobial activity was carried out following Mapook et al.[6]. Antibacterial discs of penicillin and ciprofloxacin, with antifungal discs of nystatin were used as positive control for screening[17]. Gram-positive bacteria (Bacillus subtilis, DSM10), Gram-negative bacteria (Escherichia coli, DSM498), and filamentous fungus (Mucor plumbeus, MUCL 49355) were determined by the zone of inhibition using agar plug diffusion method, compared with positive control[18].
DNA extraction, PCR amplification and sequencing
-
DNA extraction, PCR amplification and sequencing were carried out following the methodology as described by Mapook et al.[6]. The partial large subunit nuclear rDNA (LSU) was amplified with primer pairs LROR and LR5[19]. The partial small subunit nuclear rDNA (SSU) was amplified with primer pairs NS1 and NS4[20]. The internal transcribed spacer (ITS) was amplified by using primer pairs ITS5 and ITS4[20]. The translation elongation factor 1-α (TEF1) was amplified by using primers EF1-983F and EF1-2218R[21]. The PCR products were sent for sequencing at SeqLab GmbH (Microsynth AG), Göttingen, Germany.
Phylogenetic analysis
-
Phylogenetic analysis was carried out following Mapook et al.[6] and Dissanayake et al.[22]. The closest relative strains were selected following Mapook et al.[6,7]. The combined aligned dataset was analyzed based on maximum likelihood (ML) and Bayesian inference (BI) via the CIPRES Science Gateway platform (V. 3.3,
www.phylo.org )[23]. ML analysis was performed by RAxML-HPC2 on XSEDE (8.2.12) tool[24,25] using the GTR+I+G model of nucleotide substitution with 1,000 rapid bootstrap replicates. BI analysis was performed by using MrBayes on XSEDE (3.2.7a) tool with the GTR+I+G model. Six simultaneous Markov chains were performed for 5,000,000 generations, and trees were sampled every 1000th generation. Phylogenetic trees were drawn using FigTree 1.4.0[26] and edited by Microsoft Office PowerPoint 365 and Adobe Photoshop CS6. The sequences used for analyses with accession numbers are given in Table 1.Table 1. Taxa used in this study and their GenBank accession numbers. New sequences are in bold.
Taxa Strain no.1 GenBank accession numbers2 LSU SSU ITS TEF1 Acrospermum adeanum M133 EU940104 EU940031 EU940180 − Acrospermum compressum M151 EU940084 EU940012 EU940161 − Acrospermum gramineum M152 EU940085 EU940013 EU940162 − Arxiella dolichandrae CBS 138853T KP004477 − KP004449 − Arxiella terrestris CBS 268.65T MH870201 − MH858565 − Dyfrolomyces phetchaburiensis MFLUCC 15-0951T MF615402 MF615403 − − Dyfrolomyces rhizophorae BCC15481 − KF160009 − − Dyfrolomyces rhizophorae JK 5456A GU479799 − − GU479860 Dyfrolomyces thailandica MFLU 16-1173T KX611366 KX611367 − − Dyfrolomyces thamplaensis MFLUCC 15-0635T KX925435 KX925436 − KY814763 Dyfrolomyces tiomanensis NTOU3636T KC692156 KC692155 − KC692157 Leptodiscella africana CBS 400.65T MH870275 − MH858635 − Leptodiscella brevicatenata FMR 10885T FR821311 − FR821312 − Leptodiscella chlamydospora MUCL 28859 FN869567 − FR745398 − Leptodiscella rintelii CBS 144927T LR025181 − LR025180 − Lophium mytilinum AFTOL-ID 1609 DQ678081 DQ678030 − DQ677926 Melomastia maolanensis GZCC 16-0102T KY111905 KY111906 − KY814762 Muyocopron alcornii BRIP 43897T MK487708 − MK487735 MK495956 Muyocopron atromaculans MUCL 34983T MK487709 − MK487736 MK495957 Muyocopron castanopsis MFLUCC 10-0042 − JQ036225 − − Muyocopron castanopsis MFLUCC 14-1108T KU726965 KU726968 MT137784 MT136753 Muyocopron chromolaenae MFLUCC 17-1513T MT137876 MT137881 MT137777 MT136756 Muyocopron chromolaenicola MFLUCC 17-1470T MT137877 MT137882 MT137778 MT136757 Muyocopron coloratum CBS 720.95T MK487710 − NR_160197 MK495958 Muyocopron dipterocarpi MFLU 18-2582 − MW079363 MW063196 − Muyocopron dipterocarpi MFLUCC 14-1103T KU726966 KU726969 MT137785 MT136754 Muyocopron dipterocarpi MFLUCC 17-0075 MH986833 MH986829 MH986837 − Muyocopron dipterocarpi MFLUCC 17-0354 MH986834 MH986830 MH986838 − Muyocopron dipterocarpi MFLUCC 17-0356 MH986835 MH986831 MH986839 − Muyocopron dipterocarpi MFLUCC 17-1464NT OQ861270 OQ861267 OQ832759 OQ856779 Muyocopron dipterocarpi MFLUCC 18-0470 MK348001 MK347890 MK347783 − Muyocopron garethjonesii MFLU 16-2664T KY070274 KY070275 − − Muyocopron geniculatum CBS 721.95T MK487711 − MK487737 MK495959 Muyocopron heveae MFLUCC 17-0066T MH986832 MH986828 MH986836 − Muyocopron laterale CBS 127677 MK487718 − MK487744 MK495965 Muyocopron laterale CBS 141029T MK487712 − MK487738 MK495960 Muyocopron laterale CBS 141033 MK487715 − MK487741 MK495963 Muyocopron laterale CBS 145309 MK487722 − MK487748 MK495969 Muyocopron laterale CBS 145310 MK487719 − MK487745 MK495966 Muyocopron laterale CBS 145311 MK487724 − MK487750 − Muyocopron laterale CBS 145312 MK487725 − MK487751 MK495971 Muyocopron laterale CBS 145313 MK487721 − MK487747 MK495968 Muyocopron laterale CBS 145314 MK487723 − MK487749 MK495970 Muyocopron laterale CBS 145315 MK487720 − MK487746 MK495967 Muyocopron laterale CBS 145316 MK487726 − MK487752 MK495972 Muyocopron laterale CBS 719.95 MK487714 − MK487740 MK495962 Muyocopron laterale FMR 13797 MK874616 − MK874615 MK875803 Muyocopron laterale IMI 324533 MK487713 − MK487739 MK495961 Muyocopron laterale URM 7801 MK487717 − MK487743 − Muyocopron laterale URM 7802 MK487716 − MK487742 MK495964 Muyocopron lithocarpi − MK447738 MK447740 − − Muyocopron lithocarpi MFLU 18-2087 MK347930 MK347821 MK347716 − Muyocopron lithocarpi MFLU 18-2088 MK347931 MK347822 MK347717 − Muyocopron lithocarpi MFLUCC 10-0041 JQ036230 JQ036226 − − Muyocopron lithocarpi MFLUCC 14-1106T KU726967 KU726970 MT137786 MT136755 Muyocopron lithocarpi MFLUCC 16-0962 MK348034 MK347923 − − Muyocopron lithocarpi MFLUCC 17-1465 MT137878 MT137883 MT137779 MT136758 Muyocopron lithocarpi MFLUCC 17-1466 MT137879 MT137884 MT137780 MT136759 Muyocopron lithocarpi MFLUCC 17-1500 MT137880 MT137885 MT137781 MT136760 Muyocopron zamiae CBS 203.71T MK487727 − − MK495973 Mycoleptodiscus endophytica MFLUCC 17-0545T MG646946 MG646978 MG646961 MG646985 Mycoleptodiscus suttonii CBS 141030 MK487729 − − MK495975 Mycoleptodiscus suttonii CBS 276.72T MK487728 − MK487753 MK495974 Mycoleptodiscus terrestris CBS 231.53T MK487730 − MK487754 MK495976 Mycoleptodiscus terrestris IMI 159038 MK487731 − MK487755 MK495977 Mytilinidion rhenanum CBS 135.34 FJ161175 FJ161136 − FJ161092 Neocochlearomyces chromolaenae BCC 68250T MK047514 MK047552 MK047464 MK047573 Neocochlearomyces chromolaenae BCC 68251 MK047515 MK047553 MK047465 MK047574 Neocochlearomyces chromolaenae BCC 68252 MK047516 MK047554 MK047466 MK047575 Neomycoleptodiscus venezuelense CBS 100519T MK487732 − MK487756 MK495978 Palawania thailandensis MFLU 16-1871 KY086494 − MT137788 − Palawania thailandensis MFLUCC 14-1121T KY086493 KY086495 MT137787 − Paramycoleptodiscus albizziae CBS 141320 KX228330 − KX228279 MK495979 Paramycoleptodiscus albizziae CPC 27552T MH878220 − − − Pseudopalawania siamensis MFLUCC 17-1476aT − MT137789 MT137782 MT136752 Pseudopalawania siamensis MFLUCC 17-1476b − MT137790 MT137783 − Setoapiospora thailandica MFLUCC 17-1426T MN638847 MN638851 MN638862 MN648731 1 AFTOL-ID: Assembling the Fungal Tree of Life; BCC: BIOTEC Culture Collection; BRIP: Biosecurity Queensland Plant Pathology Herbarium, Brisbane, Australia; CBS: Westerdijk Fungal Biodiversity Institute, Utrecht, The Netherlands; CPC: Culture collection of Pedro Crous, the Netherlands; FMR: Facultad de Medicina, Reus, Tarragona, Spain; GZCC: Guizhou Culture Collection; IMI: The International Mycological Institute Culture Collections; JK: J. Kohlmeyer; MFLU: the Herbarium of Mae Fah Luang University; MFLUCC: Mae Fah Luang University Culture Collection, Chiang Rai, Thailand; MUCL: Belgian Coordinated Collections of Microorganisms; URM: Universidade Federal de Pernambuco; T: ex-type isolates; NT: Neotype.
2 LSU: 28S large subunit of the nrRNA gene; SSU: 18S small subunit of the nrRNA gene; ITS: internal transcribed spacer regions 1 and 2 including 5.8S nrRNA gene; TEF1: partial translation elongation factor 1-α gene. -
Seventy-seven strains are included in the combined dataset of LSU, SSU, ITS, and TEF1 sequence data, including our strain (MFLUCC 17-1464), which comprise 3969 characters with gaps. Tree topology of the ML analysis was similar to the BYPP. A best scoring RAxML tree with a final likelihood value of −25,539.739638 is presented in Fig. 1. The matrix had 1738 distinct alignment patterns, with 49.64% of undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.237294, C = 0.253374, G = 0.287766, T = 0.221566; substitution rates: AC = 1.187676, AG = 2.326266, AT = 1.446900, CG = 1.082572, CT = 5.256850, GT = 1.000000; gamma distribution shape parameter α = 0.309869. The phylogram generated from ML analysis (Fig. 1) shows that our strain grouped within Muyocopron dipterocarpi clade. In a BLASTn search of NCBI GenBank, the closest match of the LSU, ITS, and TEF1 sequence of MFLUCC 17-1464 is Mu. dipterocarpi with 100% similarity to the strain MFLUCC 14-1103 (ex-holotype), while the closest match of the SSU sequence was identical with 99.90% similarity to Mu. dipterocarpi (strain MFLUCC 14-1103).

Figure 1.
Phylogram generated from maximum likelihood analysis based on combined dataset of LSU, SSU, ITS, and TEF1 sequence data. Bootstrap support values for maximum likelihood (ML) equal to or greater than 60% and Bayesian posterior probabilities (BYPP) equal to or greater than 0.97 are given above the nodes. Newly generated sequences are in dark red bold and ex-type isolates are in bold. Lophium mytilinum (AFTOL-ID 1609) and Mytilinidion rhenanum (CBS 135.45) are used as outgroup taxa.
Taxonomy
-
Muyocopron dipterocarpi Mapook, Doilom, Boonmee & K.D. Hyde, Phytotaxa 265(3): 232 (2016)
Index Fungorum number: IF551617, Facesoffungi number: FoF 01889; Fig. 2
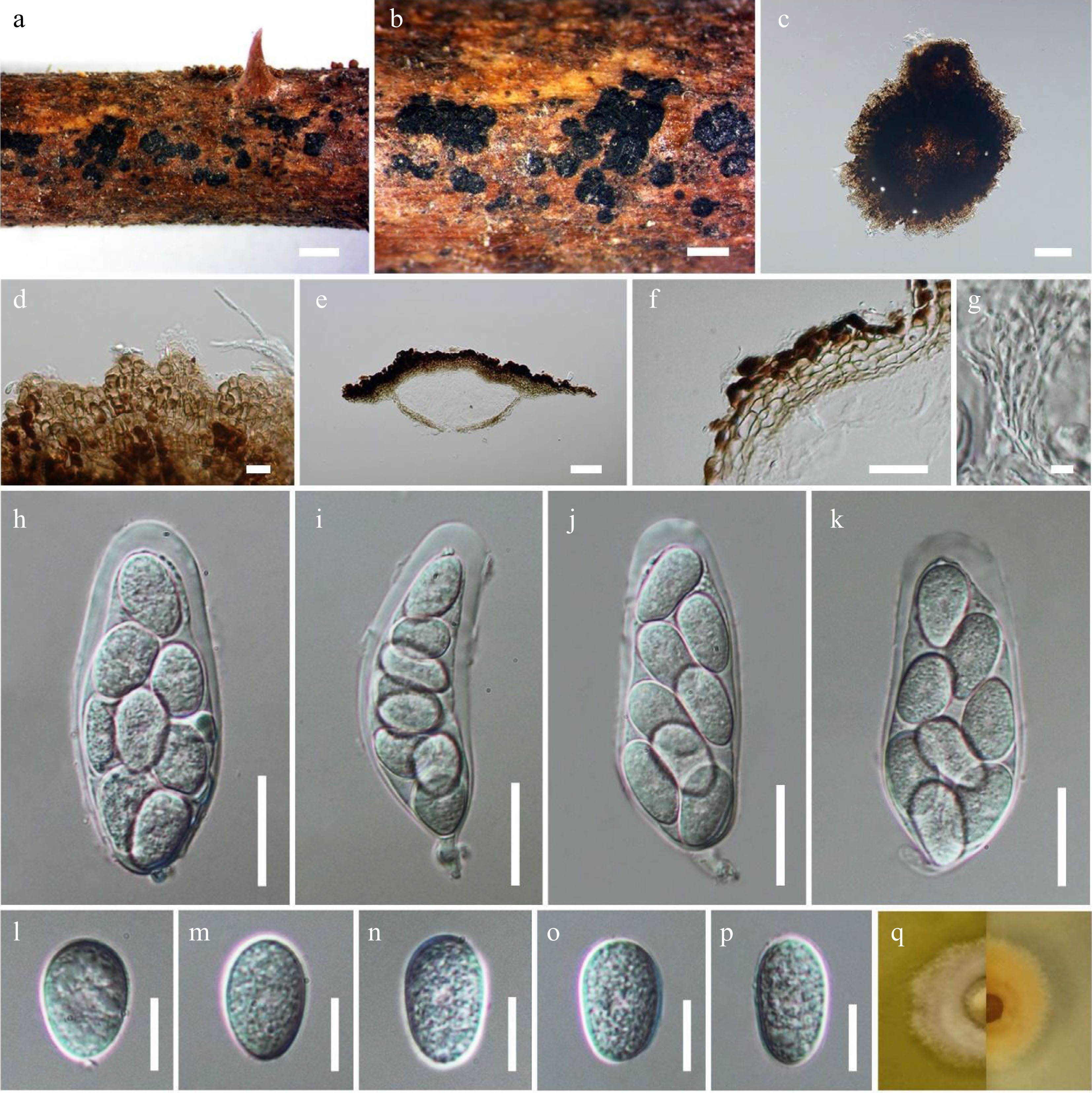
Figure 2.
Muyocopron dipterocarpi (neotype). (a), (b) Superficial ascomata on substrate. (c), (d) Squash mounts showing ascomata walls. (e) Section of ascoma. (f) Peridium. (g) Pseudoparaphyses. (h)−(k). Asci. (l)−(p) Unicellular ascospores. (q) Culture characteristic on MEA. Scale bars: a = 1,000 µm, b = 500 µm, c = 100 µm, e = 50 µm, f, h−k = 20 µm, d, l−p = 10 µm, g = 5 µm.
Saprobic on dead twigs of Zanthoxylum fagara (L.) Sarg. Sexual morph: Ascomata (65–)80–110 µm high × (230–)340–395 µm diam. (
$\overline{\chi} $ $\overline {\chi} $ $\overline {\chi} $ Culture characteristics: Ascospores germinating on MEA within 24 hr at room temperature and germ tubes produced from the ends of the ascospore. Colonies on MEA irregular, initially aerial mycelium white, slightly raised, filiform, becoming light brown from the center, flattened on surface, pale brown to light brown in reverse from the center of the colony with white margin.
Pre-screening for antimicrobial activity: Muyocopron dipterocarpi (MFLUCC 14-1103, ex-holotype) showed antimicrobial activity against Bacillus subtilis and Escherichia coli (11 and 10 mm inhibition zone, respectively), observable as partial inhibition, when compared with the positive control (26 and 9 mm, respectively), but no inhibition of Mucor plumbeus; Mu. dipterocarpi (MFLUCC 17-1464) showed antimicrobial activity against M. plumbeus, B. subtilis and E. coli (11, 8, and 10 mm inhibition zone, respectively), observable as partial inhibition, when compared with the positive control (17, 26 , and 9 mm, respectively).
Known hosts and distribution: On dried twigs of Dipterocarpus tuberculatus (Dipterocarpaceae) in Chiang Rai Province, Thailand[12]; on dried twig of Hevea brasiliensis (Euphorbiaceae) in Phayao Province, Thailand[27]; on dead twigs of Mangifera indica (Anacardiaceae) in Sukhothai Province, Thailand[28]; on decaying pod septum of Delonix regia (Fabaceae) in Phrae Province, Thailand[29]; on dead leaves and decaying twig of Celtis formosana (Cannabaceae) in Taiwan[30, 31].
Material examined: THAILAND, Phrae Province, on dead twigs of Zanthoxylum fagara (Rutaceae), 22 September 2016, A. Mapook, (DPKP1, MFLU 23-0072, neotype designated here), ex-neotype culture MFLUCC 17-1464.
Note: We identify our isolate (MFLUCC 17-1464) as Mu. dipterocarpi, based on phylogenetic analyses, together with morphological comparison. However, our isolate was found on different host families as compared with other previously reported strains. Therefore, the isolate is introduced here as a new host record on Zanthoxylum fagara from Thailand.
During this study, we tried to examine the original collection of this species. Unfortunately, the type material of Mu. dipterocarpi could not be located in the MFLU fungarium, where the holotype specimen was deposited with a code number MFLU 15–1132[12]. A neotype for Mu. dipterocarpi is, therefore, needed to facilitate subsequent taxonomic work in stabilizing the application of name, and to serve as a foundation for further applied research of this species. Although the species have been reported from various hosts and is mostly distributed in Asia, especially in Thailand[6,12,27−29], there are no collections from the same locality as indicated in the protologue. Neotypes from different locations may also be considered as long as the author are confident that they are conspecific[32]. Therefore, a neotype specimen is designated here for Mu. dipterocarpi due to the original material no longer existing, and in accordance with Art. 9.16[33]. This specimen is recent and well-preserved, and has typical morphology suitable to the description given in the protologue, completed with illustrations, molecular data and additional evidence of a potential for antimicrobial activity based on a preliminary screening.
-
Muyocopron species can be saprobic, endophytic, or pathogenic on various hosts with an ability to produce specialized infection structures such as appressoria from germinating ascospores and therefore probably has an endophytic lifestyle[3,27,34]. Muyocopron dipterocarpi is mostly reported from northern Thailand and probably not specific to the host, due to the species have been reported on a variety of plant families such as Anacardiaceae, Dipterocarpaceae, Euphorbiaceae, and Fabaceae[6,12,27−29]. The species have not been reported to form any specialized infection structures from the other hosts, as well as our strain in this study except the first isolate from a dried twig of Hevea brasiliensis (Euphorbiaceae), which was assumed to have endophytic lifestyle[12,27−31,34,35]. This suggests that Muyocopron species may have the ability to be endophytic or pathogenic and probably not specific to any hosts due to the genus reported on various hosts. Although fungi have the ability to change their lifestyle which can be exhibited in more than one lifestyle in a different host, the mechanism of appressoria production and the ecological lifestyle of Muyocopron is not well understood. In addition, Mu. dipterocarpi also has potential for antimicrobial activity against the tested organism based on a preliminary screening in this study.
Ausana Mapook would like to thank the Mae Fah Luang University Fund (Grant No. 651A16029), entitled 'Taxonomy, phylogeny, risk assessment, and potential impact of fungi on Siam weed in northern Thailand'. Kevin D. Hyde thanks the Basic Research Fund 2022 (Grant No. 652A01001), entitled 'Studies of fungi associated with Asteraceae and the discovery of biological properties', Princess Srinagarindra's Centenary Celebrations Foundation (Grant No. 64316001), and National Research Council of Thailand (NRCT) grant, entitled 'Total fungal diversity in a given forest area with implications towards species numbers, chemical diversity and biotechnology' (Grant No. N42A650547).
-
Kevin D. Hyde is the Editorial Board members of Journal Studies in Fungi. He was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and his research groups.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Mapook A, Hyde KD. 2023. Neotypification of Muyocopron dipterocarpi, a new host record on Zanthoxylum fagara (Rutaceae) and the potential for secondary metabolite production in Muyocopronaceae. Studies in Fungi 8:9 doi: 10.48130/SIF-2023-0009 

Neotypification of Muyocopron dipterocarpi, a new host record on Zanthoxylum fagara (Rutaceae) and the potential for secondary metabolite production in Muyocopronaceae
- Received: 26 January 2023
- Accepted: 09 April 2023
- Published online: 27 April 2023
Abstract: Muyocopron is a genus with a diverse lifestyle, occurring in tropical and temperate regions and can be found on various hosts and habitats. The present study confirmed a new host record of Muyocopron dipterocarpi from dead twigs of Zanthoxylum fagara in northern Thailand, based on both morphological comparisons with multigene analyses of LSU, SSU, ITS, and TEF1 sequence data. A preliminary screening test also showed that M. dipterocarpi has a potential for antimicrobial activity, observable as partial inhibition, when compared with a positive control. In addition, a neotype is designated here for Mu. dipterocarpi due to the original material no longer existing. This will facilitate subsequent taxonomic work in stabilizing the application of a name, and to serve as a foundation for further applied research of this species.
-
Key words:
- Dothideomycetes /
- Northern Thailand /
- Phylogeny /
- Preliminary screening /
- Taxonomy