









-
In angiosperms, the reproduction process starts once the pollen grain lands on the stigma. Before pollen tubes grow into stigma for fertilization, complicated communications between pollen and stigma papillary cells occur. The stigma papillae have to distinguish compatible pollen from foreign pollen, incompatible pollen and fungal spores. In general, flowering plants could be classified into dry stigma type and wet stigma type. In dry stigma plants such as Brassicaceae, stigma provides water for pollen hydration and germination after stringent pollen-stigma interactions. Encompassing diverged kinds of pollens including self-compatible (SC), self-incompatible (SI), unilaterally compatible (UC) and unilaterally incompatible (UI), the Brassicaceae represents an ideal model family for extensive study of the molecular mechanism of pollen-stigma interactions.
Pollen-stigma communications via receptor-ligand pairs -
It has been long known that high levels of reactive oxygen species (ROS) present in the stigma across all representative angiosperm species[1]. Recent studies in SC plant Arabidopsis thaliana discovered that CrRLK1L receptor kinases FERONIA and ANJEA (FER/ANJ) sense the cysteine-rich peptides RAPID ALKALINIZATION FACTOR (RALF) 23/33 to activate downstream signaling for ROS production[2]. The pollen specific cysteine-rich peptides POLLEN COAT PROTEIN B-class peptides (PCP-Bs) have been characterized regulating pollen hydration and germination[3]. Upon pollination, the PCP-B peptides compete the RALF23/33 peptides for binding to FER/ANJ receptors, reducing stigmatic ROS and promoting pollen hydration[2]. With regard to the ubiquitous presence of CrRLK1Ls, RALFs, PCP-Bs and all the signaling components in flowering plants, the antagonistic control of ROS levels by pollen PCP-Bs and stigma RALFs via CrRLK1Ls might represent a fundamental mechanism underlying SC responses.
In SI plants, the SI response of pollen and stigma within species is regulated by the pollen determinant S-locus cysteine-rich protein/S-locus protein 11 (SCR/SP11) and the receptor S-locus receptor kinase (SRK) on the surface of stigma papilla cells. The SRK could only recognize the SCR/SP11 from the same haplotype but not from the different haplotype, leading to the activation of the SI responses[4]. It is also found that FER and its downstream ROS production signaling pathway are required for the SI responses in Brassica rapa[5].
In more complicated cases in interspecific hybridization, pollen shows UI on SI pistils, that is the SI stigma rejects the pollen, but UC on SC pistils, that is the SC stigma accepts pollen. The results from multiset hybridization experiments showed that the presence of the SI system is important for the outcome of intraspecific and interspecific hybridization, suggesting the involvement of the S-locus in UI responses[6]. However, the mechanism underlying SI, UI, SC, and UC and their interconnections remain elusive. In the latest issue of Nature, Huang et al.[7] now report that SCR/SP11 peptides from SI pollen and UI pollination activate the SRK receptor kinase, which subsequently recruit and activate the FER signaling pathway for incompatible pollen rejection; in compatible response, signals from SC and UC pollen activate nitric oxide (NO) to nitrosate FER, reduce ROS accumulation and promote pollen hydration and germination. This work leads to the implication that the SRK-mediated SI signaling pathway and the FER-mediated SC signaling pathway dominate the intraspecific and interspecific barriers.
SRK and FER function in intra- and interspecific pollen recognition -
Using Brassica rapa stigma and pollen from intraspecific B. rapa and interspecific Brassica oleracea, the more distant intergeneric plants Barbarea vulgaris and A. thaliana, Huang et al. showed that both the stable and the antisense-oligo deoxyribonucleotide (AS-ODN) SRK mutation disturb the intraspecific and interspecific pollen rejection responses including ROS level alteration and pollen germination[7]. Moreover, interspecific pollination of B. rapa with AS-ODN-FER and transgenic SI A. thaliana harboring SCR/SP11, SRK and ARC1 from A. halleri S13 haplotype in the fer-4 mutant background show that FER mutation breaks both the SI and the UI responses and promotes pollen hydration and germination. In addition, inhibition of the SI pathway components downstream of the SRK such as M-locus protein kinase (MLPK) and ARM-repeat containing1 (ARC1) E3 ubiquitin ligase result in similar responses compared with SRK mutation. Further biochemistry assays manifest the interaction between FER and SRK, which could be enhanced by the SI pollen extract and the SCR peptides from the same haplotype, as well as the UI pollen extract[7].
ROS and NO regulate compatible responses -
A FER-regulated signaling pathway for ROS production was characterized in A. thaliana roots and root hairs. FER and its co-receptor LORELEI (LRE)/LRE-LIKE GLYCOSYLPHOSPHATIDYLINOSITOL-ANCHORED PROTEIN 1 (LLG1) form a receptor-coreceptor complex to activate guanine nucleotide exchange factor (RopGEFs) and Rho-like GTPases (RAC/ROPs), which then activates NADPH oxidases for ROS production[8,9]. Similarly, in A. thaliana stigma papillae perceiving the autocrine RALF23/33 peptides the FER/ANJ‒LLG1 complex activates the RopGEF‒ROP‒RBOHD signaling pathway to promote ROS generation. Upon compatible pollination, the RALF23/33‒FER/ANJ-controlled ROS generation pathway would be repressed by pollen PCP-Bs peptides via antagonistic peptide competition, resulting in reduced ROS levels[2]. Compared to the wild type, the fer-4 A. thaliana stigma shows less reduction of ROS levels, faster pollen hydration and longer pollen tubes upon interspecific pollination than A. thaliana pollination. Pull-down results also show stronger binding affinity of AtFER extracellular domain to the intraspecific AtPCP-Bγ rather than the interspecific BrPCP-Bγ, suggesting that FER acts as an important regulator of the intraspecific pollination and therefore a barrier of the interspecific pollination[7].
NO and ROS are closely associated reactive molecules[10]. In A. thaliana, NO levels increased 2 to 5 min after pollination followed by a sharp decrease and the NO inhibitor cPTIO treatment of stigma failed to induce the stigmatic ROS reduction. The NO-deficient and NO-overaccumulating mutants also support the function of NO in SC pollination. Furthermore, the Cys730 and Cys752 in the FER kinase domain are nitrosylated upon SC pollination, which interferes with the interaction between FER and downstream ROP and in turn the production of ROS. Nitrosylation of downstream regulator RBOHs could also be detected. Moreover, in B. rapa, stigmatic NO is stimulated by SC pollination but not by SI or UI pollination[7]. These results together confirm the function of NO in compatible pollination.
Trying to break the interspecific barriers, the ROS scavenger Na-SA, NO donor GSNO, and the AS-ODN disrupting the BrSRK, BrFER and BrRBOH are applied to the B. rapa stigma followed with intra and interspecific pollination. Plenty of enlarged ovules are detected upon B. rapa SI pollination as well as in B. oleracea and B. vulgaris UI pollination with less numbers[7] (Fig. 1).
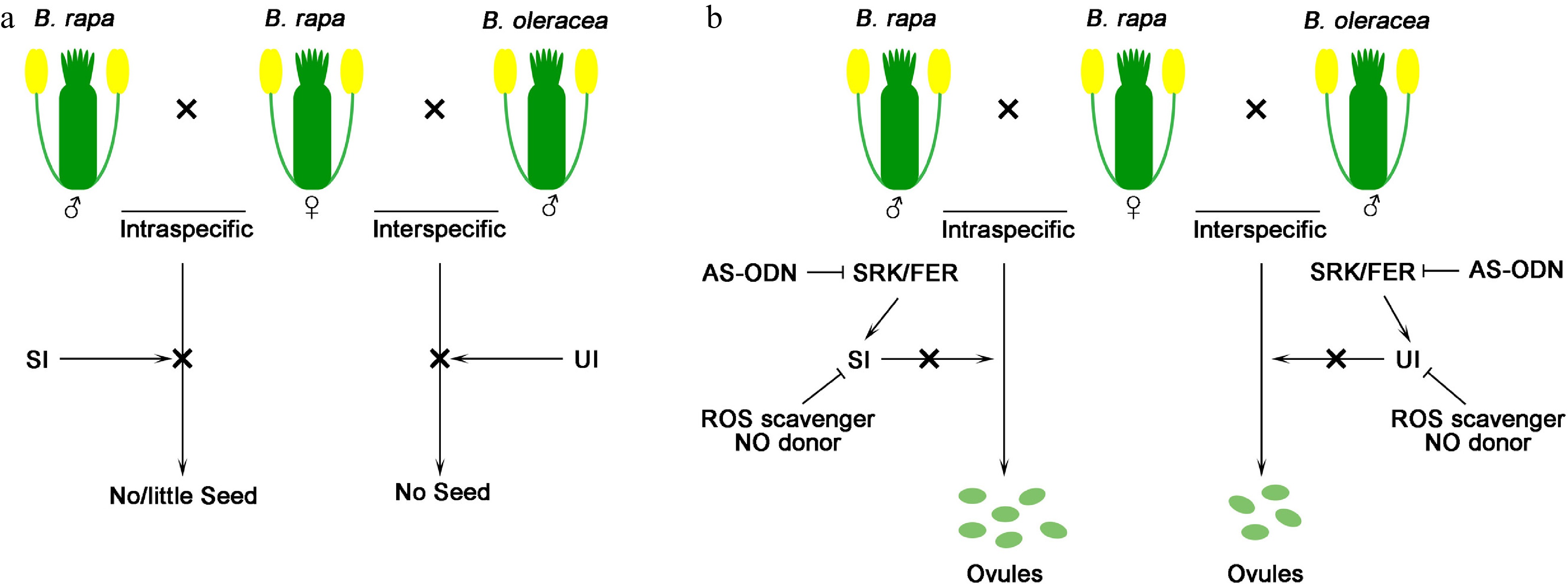
Figure 1.
Schematic illustration of intra- and interspecific hybridization by interference of self-incompatible (SI) and unilaterally incompatible (UI) responses. (a) Intraspecific and interspecific pollination could not produce ovules due to SI or UI barriers. (b) Intraspecific and interspecific hybridization by interfering with SI and UI responses. Knocking down SRK and FER signaling by AS-ODN and application of ROS scavenger and NO donor both enable the ovule development in interspecific or intraspecific hybridization.
Concluding remarks and future perspectives -
So far, studies have discovered the interplay between the two crucial receptor kinases SRK and FER in intraspecific and interspecific pollen-stigma recognition. However, many interesting questions await further study. How does compatible pollination induce the generation of the stigmatic NO? How is the NO level alteration regulated? The mechanism of the redox regulation of the stigma papillae would deepen our understanding of the pollen-stigma interaction processes. Besides, further exploration of the signal from UI pollen that activates the SRK and FER signaling should provide valuable clues for the UI responses. In addition, the Brassicaceae family include many important vegetable and oil crops, manipulation of the ROS, NO, and/or SRK‒FER signaling components could be useful approaches for distant hybridization.
The authors are supported by grants from the National Natural Science Foundation of China (32230009 and 31972863).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
Acknowledgments Conflict of interest Rights and permissions (1) References(10) - About this article
Cite this articleCheng L, Li C. 2023. Stigma receptors function as barriers between intraspecies and interspecies in Brassicaceae. Seed Biology 2:3 doi: 10.48130/SeedBio-2023-0003 

-
Stigma receptors function as barriers between intraspecies and interspecies in Brassicaceae
- Received: 30 January 2023
- Accepted: 31 January 2023
- Published online: 09 February 2023
-
Key words:
- Receptors /
- Intraspecies /
- Interspecies /
- Stigma