









-
Almond (Prunus dulcis) is a traditional crop in the Mediterranean Basin, and an important nut crop with high economic significance due to its ecological, organoleptic, and nutritional characteristics, as well as its flavour and culinary versatility[1]. Almond production is significant in countries such as the USA, Spain, Australia, Türkiye, and Morocco[2]. The recent expansion in almond cultivation has been accomplished by using self-fertile cultivars, intensification of production systems, and installation of irrigation, creating favourable conditions for the resurgence and spread of fungal diseases[3,4].
Among the most significant threats to almond production is anthracnose, caused by fungi in the genus Colletotrichum, namely C. acutatum, C. fioriniae, C. godetiae, C. nymphaeae, and C. simmondsii (members of the acutatum species complex)[3,5−7]. Colletotrichum godetiae is the predominant species in the Mediterranean Basin, according to data from Portugal, Spain, and Israel, although C. acutatum prevails in Australia[8,9]. On almond, Colletotrichum godetiae behaves as a hemibiotrophic pathogen that can infect flowers, young leaves, branches, and especially young fruits, causing circular, depressed orange lesions with white mycelium, gummosis, and orange spore masses[4]. The affected fruits mummify and remain on the tree during autumn and winter, becoming the main source of inoculum. Some adventitious plants in the orchard are also capable of inoculum dissemination, via sporulation upon tissue colonisation, or via epiphytic growth and secondary conidiation[10]. The disease is particularly severe during wet springs, with temperatures between 20 and 25 °C, often resulting in significant yield losses[3]. Currently, management practices are based on the preventive application of synthetic fungicides during the critical phenological stages of flowering and early fruit development. However, these chemical-based strategies raise concerns about environmental sustainability, the development of fungicide-resistant pathogen populations, and potential negative impacts on beneficial microbiota[11].
The aerial parts of plants create a habitat for communities of microorganisms known as the phyllosphere[12]. The microbial communities of the phyllosphere include endophytes that inhabit the inside of plant tissues, and epiphytes that colonize the external surface[13,14] without causing visible damage. The composition of microbial communities in the phyllosphere is influenced by both abiotic and biotic factors, such as the employment of different agricultural practices[15]. In this sense, the epiphytic community is more exposed to environmental stress (temperature, humidity, and solar radiation), as well as to the morphological and physiological characteristics of the plant[16]. In contrast, endophytes are exposed to different selection pressures within plant tissues, where they have to tolerate host defence factors[17]. Endophytes may occur inter- and intra-cellularly, systemically, or locally in the hosts without causing infection or disease. Asymptomatic endophyte colonisation is explained by the 'balanced antagonism' hypothesis, whereby the host-endophyte relationship is maintained by inactivation of the host defence, and activation of the fungal resistance mechanisms to toxic plant metabolites[18]. However, this relationship may be dynamic and unstable, as the boundary between mutualism and parasitism often blurs, and some endophytic species may switch between endophytic, pathogenic and saprophytic lifestyles[19] depending on the environmental conditions, availability of nutrients, and the physiological state of the host[20,21]. An example is Colletotrichum magnum, whose ability to cause symptoms is inhibited by disrupting the genetic loci that induce anthracnose in Cucurbitaceae, and it therefore becomes an endophyte[22].
Biological control has emerged as an integrated strategy for pest and disease control, offering a sustainable alternative to conventional chemical control[14,23], thus promoting more sustainable and resilient farming. This approach is based on the use of antagonistic microorganisms to inhibit phytopathogenic activity by multiple mechanisms, such as competition for space and nutrients, mycoparasitism, volatile compounds production, inhibition of biofilms, induced resistance, and modulation of the immune responses of host plants[24−26]. Endophytic fungi are distinguished as biocontrol agents because they tend to colonise plant tissues without causing symptoms and attack pathogens in the same ecological niche[27]. Endophytic agents may also contribute to increased tolerance to pathogens, growth and biomass production, and prevention of abiotic stress[28]. They can also produce toxins that inhibit the growth of other fungi[23].
Endophytic fungi have been shown to have biocontrol potential against diseases caused by Colletotrichum spp. Recent studies have shown that the genera Trichoderma, Chaetomium, Aureobasidium, and Epicoccum are effective in controlling Colletotrichum species[14,29]. In vitro and field trials with strains of Trichoderma harzianum have shown efficacy in inhibiting the growth of Colletotrichum acutatum, C. gloeosporioides, and C. truncatum in strawberry, pepper, lemon, banana, and mango plantations by mycoparasitism and antibiosis mechanisms[30−34]. Aureobasidium pullulans is another fungal species that acts as an antagonist of Colletotrichum acutatum in peppers, strawberries, olives, and apples after harvest[33,35−37]. These examples highlight the diversity of fungal antagonists that are effective against different species of Colletotrichum and reinforce their potential for the development of biological control strategies in different crops.
In plum trees (Prunus domestica), the endophytic fungus Phaeosphaeria nodorum was found to produce volatile compounds that play a significant role in reducing the spread of Monilinia fructicola in fruits[38]. The transition from chemical to biological control has clear environmental benefits, as reducing reliance on synthetic fungicides helps to maintain beneficial microbial communities in plants, minimises the accumulation of chemical residues in agroecosystems and delays the emergence of resistant strains of fungicides, which is an increasingly recognised threat to sustainable crop protection[39]. However, the use of antagonistic fungi for the control of phytopathogens requires specific knowledge about the structure of endophytic and epiphytic microbial communities, which are influenced by genetic variations between cultivars[14], and the state of development and organ of the host plant[40], as well as knowledge on the persistence of such microbial agents on the final product and putative downstream implications.
Despite studies aimed at characterising the diversity of fungal communities naturally associated with Prunus dulcis[41,42], the biocontrol potential of these fungi is still unknown. No study to date has evaluated the antagonistic activity of epiphytic and endophytic fungi against C. godetiae, which causes anthracnose in almond trees.
The objective of this study is to identify the fungal communities (endophytes and epiphytes) associated with the almond crop by isolating fungi from plant material with or without surface disinfection. The aim is to assess the inhibitory activity of several fungal isolates against C. godetiae in vitro to identify and select potential biological control agents against Colletotrichum species responsible for the outbreak of almond anthracnose.
-
The plant materials were collected from almond trees in winter and spring 2023, covering the phenological stages of flowering and fruit development, allowing the collection of flowers, leaves, branches, and fruits. The areas under study were selected for their importance in the current context of almond production in Portugal. This selection was mainly due to the recent establishment and onset of production of new intensive almond orchards in the regions of Alentejo, Beira Baixa, and Trás-os-Montes. The three regions have different landscapes, territorial conditions, soil, and climate characteristics. In Alentejo, 11 farms were visited, located in the municipalities of Aljustrel, Alvito, Beja, Évora, Ferreira do Alentejo, and Portel. Nine almond plantations were under an intensive system, and two were under super-intensive production systems. An almond orchard in Idanha-a-Nova, with both an intensive and super-intensive irrigated system, was visited in Beira Baixa. In Trás-os-Montes, four almond orchards were visited in the localities of Alfândega da Fé, Macedo de Cavaleiros, Mirandela, and Torre de Moncorvo. All four orchards were under intensive systems, two of which were not irrigated. Considering the three regions, the almond cultivars evaluated were 'Belona', 'Carmel', 'Constantine', 'Ferragnes', 'Francoli', 'Guara', 'Lauranne', 'Marinada', 'Marta', 'Monterey', 'Nonpareil', 'Penta', 'Shasta', 'Soleta', 'Vairo', and 'Wood Colony'. On each farm, 12 almond trees were selected at each sampling point, considering the cultivars and year of installation of the plot. Flowers, leaves, branches, and healthy and symptomatic (anthracnose) almonds were collected from each tree, between the phenological stages E and J[43]. Plant materials were transported in plastic bags and stored in a refrigerator (4 °C) until fungal isolation.
Isolation of fungi
-
Plant material was cut into fragments measuring approximately 0.50 cm2 (for fruits and branches, only the superficial part of the respective organs were used). Plant material was surface-disinfected in 1% sodium hypochlorite (NaClO) for 1 min, followed by rinsing in distilled water for 30 s, and then drying on sterile absorbent paper for 30, for the isolation of endophytic fungi. The second approach consisted of incubating the plant material without surface disinfection to promote the development of endophytic and epiphytic fungi. For both approaches, three Petri dishes containing Potato dextrose agar (PDA, Difco, Sparks, MD, USA) culture medium with bacterial growth inhibitor potassium thiocyanate (KSCN, 5 g/L) (Sigma-Aldrich, Darmstadt, Germany) were inoculated with five segments of plant material, representing each sampling point and type of plant material. Dishes were incubated at 25 °C in the dark.
Identification of fungal isolates
-
Fungal colonies were observed 10 d after inoculation for identification and counting of the fungal genus of each colony. Morphological analysis was first performed through the characteristics of the mycelium, hyphae, and reproductive structures, which were observed under a magnifying glass (Leica MZ125, Wetzlar, Germany)[44]. For molecular identification, a representative isolate of each morphotype was selected, and for this purpose, the target colonies of the fungus were transferred to PDA and incubated under the conditions described above, resulting in the formation of pure cultures by means of the serial dilution method[45]. DNA from pure culture isolates was extracted[46] for amplification of the internal transcribed spacer region (ITS1, 5.8S, ITS2) of the rDNA using the primers ITS4[47] and V9G[48] by the polymerase chain reaction (PCR). For isolates that could not be identified using rDNA-ITS, additional loci were sequenced to improve the taxonomic resolution of the analysis. The β-tubulin 2 (TUB2) and DNA-directed RNA polymerase II core subunit RPB2 (RPB2) genes were sequenced using T1[49]/Bt2B[50] and RPB2-5F2[51]/fRPB2-7cR[52] primer pairs, respectively. This multilocus approach allowed ITS to provide a tentative taxonomic location, while TUB2 and RPB2 resolved ambiguities between closely related taxa, thus increasing the reliability of the identification of species[49,51,53]. The PCR reactions were performed in a volume of 20 µL, containing 40 ng of DNA, 0.4 μM of each primer, and 10 µL of Taq DNA polymerase + dNTPs (NZYTaq II 2× Green Master Mix; Nzytech, Lisboa, Portugal). The thermocycler was programmed for: 1 cycle of 5 min at 95 °C, 30 cycles of 30 s at 95 °C, 30 s at 53 °C (RPB2) or 56 °C (ITS, TUB2), and 60 s at 72 °C, ending with 1 cycle of 10 min at 72 °C. The sequencing of the samples was performed by STABvida (Caparica, Portugal) in forward and reverse directions, so the sequence of each isolate was assembled and edited using Geneious Prime software (Boston, USA). For preliminary identification of fungal species, a search for homologous sequences was performed using BLAST software[54] in the database 'Internal transcribed spacer region (ITS) from fungi type and reference material' provided by NCBI (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ). The set of sequences (rDNA-ITS, TUB2, RPB2) was aligned using MAFFT v7.490[55] and represented by a phylogenetic tree constructed using the maximum likelihood method using FastTree2 v2.1.10[56]. Pure cultures of each isolate were preserved and deposited in the fungal plant pathogens collection of the Instituto Superior de Agronomia (Universidade de Lisboa).Assessment of fungi diversity
-
The occurrence of endophytes and of the combination of endophytes and epiphytes was measured by calculating the colonisation frequency and relative abundance. The colonisation frequency (CF) was determined by considering the total number of plant material fragments colonised by each endophyte, divided by the total number of plant material fragments[14]. Relative abundance was determined by the total number of isolates of a given genus, divided by the total number of fungal isolates[14]. The diversity of endophytes and of the set of endophytes and epiphytes was assessed by calculating the Shannon-Wiener species diversity index (H), which determines the abundance and uniformity of each taxon[57].
Antagonism tests
-
The selection of fungal isolates to be tested for their antagonistic activity against C. godetiae was made according to the following criteria: ease of cultivation on PDA; frequency (abundant species in the study were preferred to infrequent species); and previous knowledge on the putative biocontrol potential of species from the same genus. According to these criteria, isolates from 24 species were selected. The biocontrol potential of each isolate was evaluated by dual culture assay. The fungi were cultivated on PDA for 7 d at 25 °C in the dark. After this, mycelium discs (diameter 4 mm) from the actively growing margins of each test fungus and C. godetiae (isolate MR007) were removed by aseptic cutting, and placed at a distance of 3 cm from each other on Petri dishes (diameter 9 cm) containing 13 mL of PDA medium. For Aspergillus sp. and Penicillium expansum, due to their high sporulation rate, the inoculum used was 10 μL of spore suspension (1 × 106 spores/mL, in 0.025% Tween 80). The negative control consisted of PDA plates containing two inocula of C. godetiae. Each treatment was incubated at 25 ± 2 °C in the dark for 5 d, and each treatment was repeated five times. During the test, the plates were photographed daily until the tested fungus reached the edge of the plate. The area of C. godetiae colonies was determined using ImageJ v.2 software (National Institutes of Health, Boston, MA, USA). The average inhibition percentage and the inhibition percentage on the last day of the test were calculated using the adapted formula by Frari et al.[58]: Percentage of inhibition = [(CC − DC)/CC] × 100, where CC is the growth of the control colony area (cm2), and DC is the growth of the C. godetiae area (cm2).
After the end of the test, the inhibition of C. godetiae sporulation was determined by using three replicates of each fungus under test. For this purpose, a spore suspension was prepared by flooding the colonies with sterile distilled water and filtering the mycelium using a 30 μm mesh. The spore concentration (spores/mL) was determined with a Neubauer counting chamber. This value was subsequently multiplied by the total volume of suspension to obtain the absolute number of spores produced per colony. To normalize this parameter between isolates, the absolute number of spores was divided by the area of the colony to obtain the production of conidia expressed in spores/cm2. The ability of the tested fungi to reduce the sporulation of C. godetiae was evaluated by calculating the percentage of inhibition in comparison to the control. The percentage inhibition was calculated as the difference between the control and the tested colony, divided by the control value, and multiplied by 100.
Macroscopic analysis of dual-culture interactions
-
Macroscopic analysis of the dual culture interactions and morphological evolution of the C. godetiae colonies was performed through daily observation of the cultures. The interaction between the isolates and the pathogen was also described according to the following criteria[59]: (1) contact inhibition–growth of both fungi stops at the contact line (no clear zone formed); (2) distance inhibition–neither fungus enters the area inhabited by the other (clear zone formed); (3) excessive growth of one mycelium in relation to the other; (4) replacement–the mycelium of one fungus is replaced by its opponent; and (5) mutual replacement–both fungi gained part of the other's territory.
Statistical analysis
-
The inhibition coefficient and sporulation percentage of the tested fungi were analysed using analysis of variance (ANOVA). For the parameters for which replicated measurements were available per isolate, one-way ANOVA followed by Tukey-HSD test (p < 0.05) was used to assess significant differences between isolates. This included the six parameters, namely: average colony inhibition, colony inhibition last day, conidia production, conidia production inhibition per unit area, conidia production (total conidia), and conidia production inhibition per total. For the antagonism average colony inhibition, each plate measurement on each day after inoculation (dpi) was treated as an independent observation. Therefore, colony inhibition over time was analysed using a two-factor ANOVA with isolate and dpi as fixed factors. The statistical analysis was performed using R v.4.5.1 software.
-
In total, 19,802 isolates of endophytic and epiphytic fungi were obtained from flowers, leaves, branches, and fruits of 16 almond cultivars. For endophytic fungi (obtained from disinfected samples), a total of 12,211 isolates were obtained, while the combination of endophytes and epiphytes (samples without disinfection) resulted in a total of 7,591 isolates. The higher number of isolates obtained from disinfected samples may reflect the greater effectiveness of tissue-fragment isolation methods in recovering fungal colonies. Within the endophytic group, most isolates were found in the cultivars 'Lauranne' (2,701 isolates), 'Soleta' (2,195), and 'Belona' (1,119). The same cultivars also had a high Shannon-Wiener diversity index (H = 2.34, 2.25, and 2.14, respectively) (Table 1).
Table 1. Number of isolates, number of genera, colonisation frequency and Shannon-Wiener diversity index (H) of fungi obtained from 16 cultivars and their plant material of Prunus dulcis
Cultivar/Organ Endophytes Endophytes and epiphytes No. of isolates No. of genera FC (%) H No. of isolates No. of genera H Cultivar Belona 1,119 21 75.9 2.14 355 17 1.64 Carmel 588 23 74.4 2.31 202 15 2.10 Casa Nova 67 8 44.7 1.65 83 12 1.97 Constantine 216 15 62.4 2.07 130 14 1.76 Ferragnes 31 6 32.3 1.72 65 10 1.79 Ferralise 32 8 26.7 1.56 88 9 1.46 Francoli 31 5 47.0 1.11 35 11 2.12 Guara 684 23 50.1 2.05 680 18 1.85 Lauranne 2,701 26 54.0 2.34 2,303 22 2.19 Marinada 813 20 64.4 2.23 449 15 1.96 Marta 274 16 61.0 2.18 175 10 1.82 Monterey 741 22 64.0 2.41 416 15 2.14 Nonpareil 700 21 68.6 2.42 321 17 2.28 Penta 399 16 67.4 2.13 193 12 1.90 Shasta 314 12 74.4 2.13 108 6 1.27 Soleta 2,195 28 67.1 2.25 1,078 22 1.96 Vairo 614 17 49.4 2.23 630 18 2.09 Wood Colony 693 22 70.9 2.42 284 11 1.98 Organ Fruits 3,576 28 92.5 2.15 290 16 2.21 Leaves 2,504 27 63.1 2.25 1,463 19 2.21 Flowers 2,605 26 43.7 2.13 3,354 28 2.09 Branches 3,527 33 58.6 2.18 2,488 27 2.20 The results are presented separately for endophytic communities and for the combined endophytes and epiphytes communities. FC: Frequency of colonization (%); H: Shannon-Wiener index. The highest colonisation frequency was recorded in almonds (92.8%), suggesting a high rate of endophytic fungi colonisation compared to the other organs. This trend was consistent across all cultivars evaluated (Supplementary Table S1). Fungal diversity of endophytic fungi was highest in leaves (H = 2.25) and branches (H = 2.18), followed by fruits (H = 2.15) and flowers (H = 2.13). The combined epiphytic and endophytic isolates revealed that the greatest diversity was observed in fruits and leaves (H = 2.21). The highest number of combined epiphytic and endophytic isolates was found in the cultivars 'Lauranne' (2303), 'Soleta' (1078), and 'Guara' (680) (Table 1). The highest number of different genera was found, in descending order, in the cultivars 'Soleta', 'Lauranne', and 'Guara', in both modes of plant material disinfection. Alternaria was the most prevalent fungal genus in both disinfection approaches. In plant material subjected to surface disinfection, a total of 12,211 isolates were obtained, of which 3,134 (25.7%) corresponded to the genus Alternaria. In plant material without disinfection, of the total 7,591 isolates obtained, 1,810 (23.8%) corresponded to Alternaria (Fig. 1). The genera Cladosporium, Rhizopus, Penicillium, and Trichoderma showed a high prevalence in both disinfection modalities, accounting for 16.0%, 11.8%, 7.6%, and 9.0% of isolates with 1% NaClO disinfection, and 26.7%, 8.8%, 12.6%, and 1.4% of isolates without disinfection, respectively (percentages ranked in order of the genera presented). The genera Acremonium, Clonostachys, Meyerozyma, Neopestalotiopsis, and Talaromyces were recorded less frequently, appearing only in disinfected plant material (Fig. 1), while Lambertella and Stemphylium were exclusive to the non-disinfected modality.
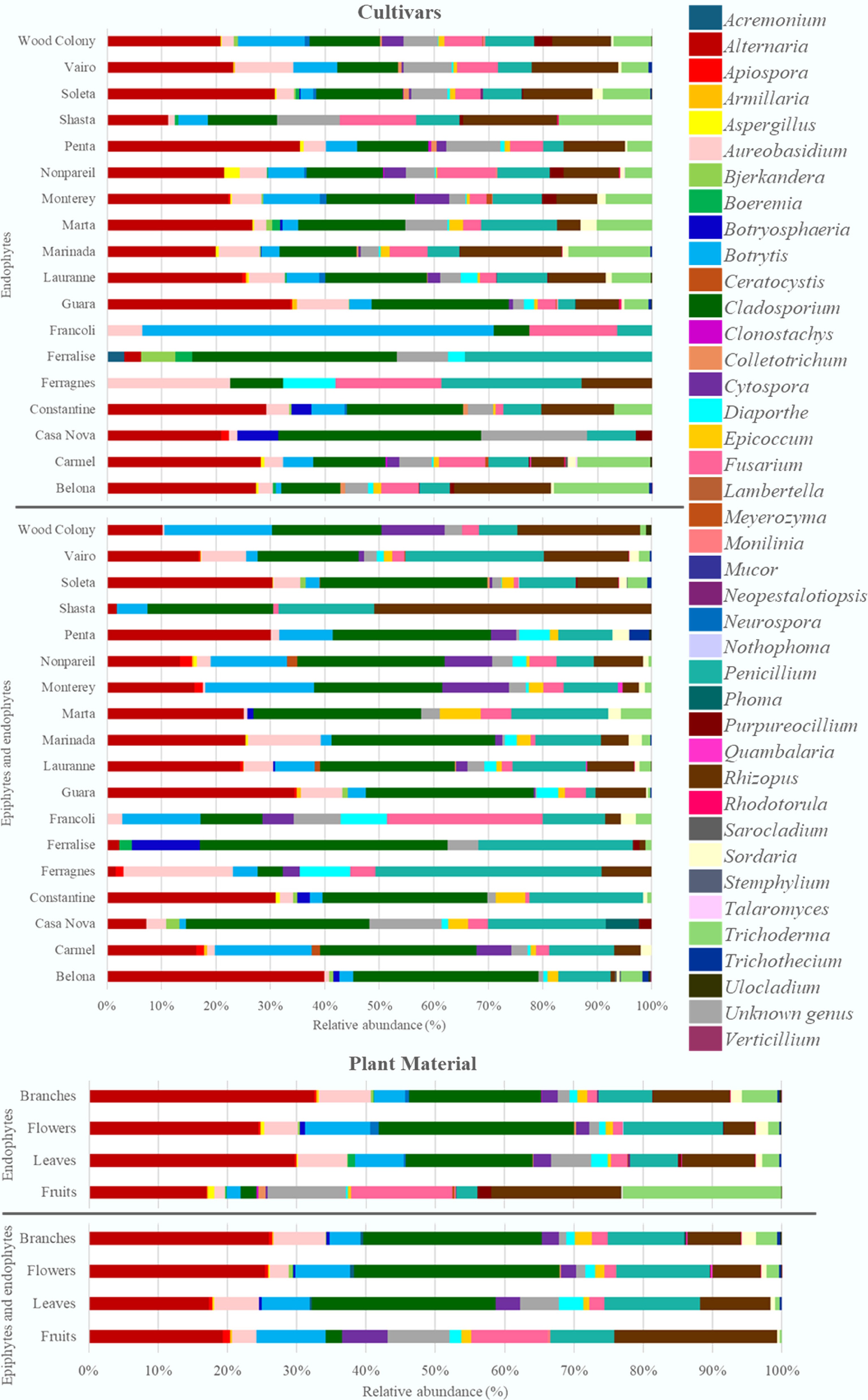
Figure 1.
Relative abundance of genera of endophytic fungi and the set of epiphytic and endophytic fungi associated with the plant material from the 16 almond cultivars analysed.
The branches presented the highest number of isolates (6,015 fungal colonies), with 58.6% of isolates coming from disinfected material (Table 1), thus reflecting a slight predominance of endophytic species in this plant tissue. The most predominant genera in the branches were Alternaria (32.5%), Cladosporium (19.0%), Rhizopus (11.1%), Penicillium (7.8%), and Aureobasidium (7.5%) (Fig. 1). The flowers, with a total of 5,959 isolates, showed a distinct trend, with 56.3% (3,354 isolates) from non-disinfected petals and sepals, indicating greater epiphytic colonisation. Cladosporium (28.2%), Alternaria (24.5%), Penicillium (14.4%), and Botrytis (9.4%) were the dominant genera. A total of 3,966 isolates were counted on leaves, of which 2,504 isolates were collected from disinfected leaves, which represents a 63.1% colonisation frequency, indicating that the number of endophytic fungi present on the leaf surface is higher than that of epiphytic fungi (Table 1). From the incubation of leaves, Alternaria, Cladosporium, Rhizopus, Penicillium, and Aureobasidium stand out in both disinfection methods for this tissue (Fig. 1). The predominant genera obtained from almonds were Trichoderma, Fusarium, and Epicoccum, with consistent isolation of Trichoderma in this organ in different cultivars.
The cultivars with the highest number of isolates were 'Lauranne', 'Soleta', 'Belona', 'Guara', and 'Nonpareil' (Table 1). The cultivars 'Lauranne', 'Soleta', and 'Belona' showed a higher predominance of endophytic isolates, including higher proportions of the fungal genera Alternaria, Rhizopus, Trichoderma, Cladosporium, and Penicillium (Fig. 1). The cultivars 'Guara' and 'Vairo' presented more balanced profiles between the two isolation methods, with an equitable distribution of endophytes and combination of epiphytes and endophytes: 'Guara' with 684 (50.1%) isolates with disinfection and 680 (49.9%) isolates without disinfection, and 'Vairo' with 614 (49.4%) and 630 (50.6%), respectively (Table 1). In both cultivars, the most prevalent genera were Alternaria, Rhizopus, Cladosporium, and Aureobasidium as endophytes and epiphytes (Fig. 1). The cultivars 'Shasta', 'Francoli', and 'Ferralise', although with a lower absolute number of isolates, also showed distinct colonisation patterns. 'Shasta', for example, had 72.5% endophytes, while 'Francoli' and 'Ferralise' showed a contrast, with greater diversity in the epiphytic and endophytic set detected in the non-disinfected modality.
Based on morphological analysis, the isolates were grouped into 39 different fungal genera belonging to 14 families divided into three phyla: Ascomycota (33 genera), Basidiomycota (four genera), and Mucoromycota (two genera) (Fig. 1). Of these genera, 19 representative morphotypes were subsequently identified by molecular analysis at the species level (Table 2; Supplementary Fig. S1). Of the isolates obtained, 4.2% could not be identified due to the absence of distinct morphological characteristics or because cultures became non-viable before molecular identification.
Table 2. List of 24 isolates used in this study (and the almond anthracnose pathogen), according to their provenance and identification based on a match of the ITS sequence using a phylogenetic tree constructed using the maximum likelihood method
Isolate code ITS GenBank Accession No. Origin of isolates Family Species Location, cultivar and plant material Treatment1 Dothideomycetes-Cladosporiales MR101 PX249825 Ferreira do Alentejo, 'Marinada'
FlowerD Cladosporiaceae Cladosporium sp. Dothideomycetes-Dothideales MR110 PX250994 Aljustrel, 'Belona', Flower ND Saccotheciaceae Aureobasidium pullulans Dothideomycetes-Pleosporales MR130 PX251240 Alfândega da Fé, 'Casa Nova', Flower ND Didymellaceae Epicoccum italicum MR090 PX249718 Ferreira do Alentejo, 'Francoli', Branch D Epicoccum layuense MR106 PX250333 Évora, 'Soleta', Branch ND Nothophoma spiraeae MR141 PX251299 Alvito, 'Nonpareil', Leaf D Pleosporaceae Alternaria sp. MR104 PX249852 Aljustrel, 'Soleta', Leaf D Alternaria vitis MR098 PX249723 Aljustrel, 'Soleta', Flower ND Stemphylium vesicarium Eurotiomycetes-Eurotiales MR117 PX251002 Beja, 'Soleta', Flower D Aspergillaceae Aspergillus ochraceus MR108 PX250988 Ferreira do Alentejo, 'Marinada', Flower ND Aspergillus tubingensis MR092 PX249719 Mirandela, 'Ferralise', Leaf ND Penicillium expansum MR149 PX251741 Aljustrel, 'Nonpareil', Almond D Trichocomaceae Talaromyces amestolkiae Sordariomycetes-Diaporthales MR099 PX249724 Macedo de Cavaleiros, 'Ferragnes', Flower ND Diaporthaceae Diaporthe foeniculina MR105 PX249856 Ferreira do Alentejo, 'Marinada', Flower ND Cytosporaceae Cytospora italica Sordariomycetes-Hypocreales MR153 PX251777 Beja, 'Penta', Almond D Bionectriaceae Clonostachys chloroleuca MR142 PX395649 Aljustrel, 'Soleta', Branch D Hypocreaceae Trichoderma citrinoviride MR093 PX249720 Ferreira do Alentejo, 'Soleta', Branch ND Trichoderma viridescens MR089 PX249569 Aljustrel, 'Soleta', Almond D Nectriaceae Fusarium chlamydosporum species complex MR127 PX251237 Aljustrel, 'Nonpareil', Branch ND Fusarium tricinctum species complex MR134 PX251287 Idanha-a-Nova, 'Guara', Almond D Fusarium annulatum MR094 PX249721 Portel, 'Lauranne', Branch ND Fusarium sambucinum species complex Sordariomycetes-Sordariales MR126 PX251117 Mirandela, 'Vairo', Leaf ND Sordariaceae Neurospora intermedia Sordariomycetes-Amphisphaeriales MR096 PX249722 Beja, 'Soleta', Flower D Apiosporaceae Apiospora kogelbergensis MR137 PX251290 Beja, 'Soleta', Leaf D Sporocadaceae Neopestalotiopsis elaeagni MR007 PX251788 Beja, 'Belona', Almond ND Glomerellaceae Colletotrichum godetiae 1Treatment of plant material: D–Plant material disinfected with 1% NaClO; ND–Plant material not disinfected. The antagonistic activity of the obtained fungal isolates against Colletotrichum godetiae
-
In the dual culture assay, 24 fungal isolates were tested for their inhibitory capacity against C. godetiae (Table 3). The isolates MR093 (Trichoderma viridescens), MR126 (Neurospora intermedia), and MR142 (Trichoderma citrinoviride) showed the highest percentages of average mycelial inhibition (83.69%, 81.81%, and 77.21%, respectively), as well as maximum inhibition on the last day of the test (91.61%, 88.53%, and 73.30%, respectively). These isolates were also notable for their ability to significantly reduce sporulation of C. godetiae, with the most effective strain being MR093 (Trichoderma viridescens), with a total conidia production inhibition of 99.31%. This fungus presented a type 3 mycelial inhibition (rapid growth of the antagonist over the pathogen). In contrast, isolates such as MR106 (Nothophoma spiraeae), MR101 (Cladosporium sp.), MR089, MR094, MR127, and MR134 (Fusarium sp.) were not effective, with negative inhibition percentages and high sporulation levels, comparable to or higher than the control.
Table 3. Antagonistic effect of 24 fungal isolates on the mycelial growth and sporulation of Colletotrichum godetiae
Family Species Isolate code Macroscopic analysis1 Average
colony
inhibition (%)Colony inhibition
last day (%)Conidia production (conidia/cm2) Conidia production inhibition (%)
per unit areaConidia production
(total of conidia)Conidia production inhibition (%) per total Cladosporiaceae Cladosporium sp. MR101 1 −1.84 ± 2.2 gh2 −12.54 ± 4.1 g2 1.27 × 106 ± 7.8 × 104g2 −91.53 ± 11.8 b2 3.89 × 107 ± 1.3 × 106 h2 −455.69 ± 19.0 c2 Saccotheciaceae Aureobasidium pullulans MR110 1 4.05 ± 2.2 fg 21.07 ± 4.1 eh 4.38 × 105 ± 7.8 × 104 bcdef 33.78 ± 11.8 a 1.26 × 107 ± 1.3 × 106 g −79.79 ± 19.0 a Didymellaceae Epicoccum italicum MR130 2, 3 26.83 ± 2.3 cd 32.53 ± 5.3 def 1.78 × 105 ± 5.5 × 104 abc 73.11 ± 8.3 a 1.81 × 106 ± 9.4 × 105 abc 74.12 ± 13.5 a Epicoccum layuense MR090 2, 3 19.06 ± 2.2 de 24.28 ± 4.1 ef 5.55 × 105 ± 7.8 × 104 def 16.11 ± 11.8 a 1.05 × 107 ± 1.3 × 106 fg −49.68 ± 19.0 a Nothophoma spiraeae MR106 2 −8.54 ± 2.0 h 0.21 ± 4.1 gh 3.83 × 105 ± 7.8 × 104 abcdef 42.17 ± 11.8 a 9.14 × 106 ± 1.3 × 106 dfg −30.44 ± 19.0 a Pleosporaceae Alternaria sp. MR141 1 35.34 ± 3.1 bc 53.88 ± 9.3 bcde 3.80 × 105 ± 7.8 × 104 abcdef 42.60 ± 11.8 a 2.62 × 106 ± 1.3 × 106 abcd 62.56 ± 19.0 a Alternaria vitis MR104 2 4.30 ± 1.9 fg 10.06 ± 4.1 fgh 3.32 × 105 ± 7.8 × 104 abcdef 49.83 ± 11.8 a 7.67 × 106 ± 1.3 × 106 cdfg −9.56 ± 19.0 a Stemphylium vesicarium MR098 2 22.87 ± 2.2 cde 29.38 ± 4.1 ef 2.56 × 104 ± 2.1 × 105 a 96.13 ± 31.2 a 5.35 × 105 ± 3.5 × 106 ab 92.37 ± 50.3 a Aspergillaceae Aspergillus ochraceus MR117 1 2.71 ± 2.8 fgh 8.32 ± 4.6 fgh 4.75 × 105 ± 7.8 × 104 cdef 28.27 ± 11.8 a 6.93 × 106 ± 1.3 × 106 bcdfg 1.02 ± 19.0 a Aspergillus tubingensis MR108 2 24.50 ± 1.6 cd 37.05 ± 4.1 cde 5.95 × 105 ± 5.5 × 104 def 10.16 ± 8.3 a 8.98 × 106 ± 9.4 × 105 dfg −28.12 ± 13.5 a Penicillium expansum MR092 2 32.30 ± 2.2 c 39.29 ± 4.1 cde 1.61 × 106 ± 7.8 × 104 g −143.12 ± 11.8 b 2.86 × 107 ± 1.3 × 106 e −307.76 ± 19.0 b Trichocomaceae Talaromyces amestolkiae MR149 1 12.32 ± 2.5 ef −2.70 ± 4.1 g 3.85 × 104 ± 5.5 × 104 a 94.18 ± 8.3 a 4.27 × 105 ± 9.4 × 105 a 93.90 ± 13.5 a Diaporthaceae Diaporthe foeniculina MR099 1, 3 21.82 ± 2.2 cde 31.96 ± 4.1 ef 1.40 × 106 ± 7.8 × 104 g −111.29 ± 11.8 b 2.62 × 107 ± 1.3 × 106 e −274.41 ± 19.0 b Cytosporaceae Cytospora italica MR105 1, 3 24.59 ± 2.2 cd 39.12 ± 4.1 cde 6.55 × 105 ± 7.8 × 104 ef 0.98 ± 11.8 a 1.05 × 107 ± 1.3 × 106 fg −49.39 ± 19.0 a Bionectriaceae Clonostachys chloroleuca MR153 4 9.75 ± 4.2 efg 15.85 ± 9.3 efgh 5.03 × 104 ± 7.8 × 104 ab 92.40 ± 11.8 a 6.34 × 105 ± 1.3 × 106 ab 90.95 ± 19.0 a Hypocreaceae Trichoderma citrinoviride MR142 3 77.21 ± 2.5 a 73.30 ± 4.1 ab 1.60 × 105 ± 7.8 × 104 abc 75.87 ± 11.8 a 5.52 × 105 ± 1.3 × 106 ab 92.12 ± 19.0 a Trichoderma viridescens MR093 3 83.69 ± 2.2 a 91.61 ± 4.1 a 2.84 × 104 ± 2.1 × 105 a 95.71 ±31.2 a 4.83 × 104 ± 3.5 × 106 a 99.31 ± 50.3 a Nectriaceae Fusarium chlamydosporum SC MR089 1, 3 27.83 ± 2.2 cd 42.34 ± 4.1 cde 2.76 × 105 ± 1.2 × 105 abcde 58.31 ± 18.0 a 3.88 × 106 ± 2.0 × 106 abcdf 44.62 ± 29.1 a Fusarium tricinctum SC MR127 1 −1.30 ± 3.2 fgh 4.21 ± 5.3 fgh 2.46 × 105 ± 7.8 × 104 abcd 62.91 ± 11.8 a 3.88 × 106 ± 1.3 × 106 abcdf 44.60 ± 19.0 a Fusarium annulatum MR134 1, 3 31.27 ± 2.5 c 26.95 ± 4.6 ef 2.88 × 105 ± 7.8 × 104 abcde 56.52 ± 11.8 a 2.48 × 106 ± 1.3 × 106 abcd 64.61 ± 19.0 a Fusarium
sambucinum SCMR094 1, 3 47.52 ± 2.2 b 58.71 ± 4.1 bc 1.34 × 105 ± 7.8 × 104 abc 79.79 ± 11.8 a 1.47 × 106 ± 1.3 × 106 abc 79.02 ± 19.0 a Sordariaceae Neurospora intermedia MR126 3 81.81 ± 2.5 a 88.53 ± 4.1 a 1.64 × 105 ± 7.8 × 104 abc 75.15 ± 11.8 a 2.60 × 105 ± 1.3 × 106 a 96.30 ± 19.0 a Apiosporaceae Apiospora kogelbergensis MR096 1, 3 24.17 ± 2.2 cde 42.29 ± 4.1 cde 9.92 × 104 ± 9.2 × 104 abc 85.01 ± 13.9 a 1.46 × 106 ± 1.6 × 106 abc 79.12 ± 22.5 a Sporocadaceae Neopestalotiopsis elaeagni MR137 1, 3 49.95 ± 2.8 b 58.46 ± 4.6 bcd 2.64 × 105 ± 7.8 × 104 abcde 60.06 ± 11.8 a 1.51 × 106 ± 1.3 × 106 abc 78.47 ± 19.0 a Glomerellaceae Colletotrichum godetiae MR007 − − − 6.62 × 105 ± 5.6 × 104 f − 7.00 × 106 ± 9.5 × 105 cdfg − Each column shows the mean values ± SE. Average colony inhibition represents the mean inhibition across all assay days and is provided as a descriptive summary of the overall performance of each isolate. 1Macroscopic analysis: (1) The growth of the mycelium of both fungal colonies stops at the contact line; (2) Neither colony grows in the area occupied by the other; (3) Rapid growth of the mycelium of the fungus antagonistic to C. godetiae is observed; (4) Replacement of the mycelium of C. godetiae by the antagonistic fungus. 2In each column, the mean values followed by the same letter are not significantly different according to Tukey test (p < 0.05), which was conducted following the verification of significant differences in the ANOVA for each parameter: Average colony inhibition (%) – F(23,1099) = 116, p-value = 2 × 10−16; Colony inhibition last day (%) – F(23,81) = 40.27, p-value = 2 × 10−16; Conidia production (conidia/cm2) – F(26,612) = 27.26, p-value = 2 × 10−16; Conidia production inhibition (%) per unit area – F(27,610) = 27.20, p-value = 2 × 10−16; Conidia production (total of conidia) – F(26,612) = 31.34, p-value = 2 × 10−16; Conidia production inhibition (%) total – F(27,610) = 38.10, p-value = 2 × 10−16. For the parameter 'Average colony inhibition (%)', Tukey comparisons were applied to the time-averaged values for descriptive purposes, complementing the two-factor ANOVA (Isolate × dpi) presented in the Results section: F(23,1099) = 116, p-value = 2 × 10−16. The significant differences between the isolates were also marked in relation to the total average inhibition rate, average inhibition on the last day, and total conidia production. Average percentage inhibition of C. godetiae colonies ranged from −8.53% (MR106) to 83.69% (MR093), and the last day inhibition percentage ranged from −12.54% (MR101) to 91.61% (MR093). Isolate MR106 (Nothophoma spiraeae) showed 0.21% inhibition on the last day of the test, while isolates MR101 (Cladosporium sp.) and MR149 (Talaromyces amestolkiae) showed negative inhibition values (−12.54% and −2.70%, respectively) on the last day, being the least effective in inhibiting the growth area of the C. godetiae colony. The fungi Trichoderma viridescens, Stemphylium vesicarium, Talaromyces amestolkiae, and Clonostachys chloroleuca are notable in terms of their ability to inhibit conidial production of C. godetiae by more than 90%, both in absolute values and per colony area (Table 3). The two-way ANOVA revealed that the fungal isolate and the number of dpi had a significant effect on the inhibition of C. godetiae (F(23,889) = 190.56, p-value < 2 × 10−16; F(14,889) = 17.43, p-value < 2 × 10−16, respectively). A significant interaction between isolate and dpi was also detected (F(196,889) = 3.43, p-value < 2 × 10−16), indicating that the inhibition dynamics varied over time depending on the antagonist tested. These results indicate that isolates differ significantly not only in their overall inhibitory potential but also in the time course of inhibition during the test, and that the effect is highly dependent on the specific interaction between isolate and pathogen.
Intermediate inhibition was observed in Fusarium spp. (MR089, MR094, MR127, and MR134), Epicoccum italicum, Neopestalotiopsis elaeagni, Trichoderma citrinoviride, and Neurospora intermedia, with an inhibitory value of between 80% and 55%, when referred to conidia/cm2. However, the absolute number of conidia was lower in the interactions with MR126 of Neurospora intermedia (2.6 × 105 spores) and MR142 of Trichoderma citrinoviride (5.5 × 105 spores), suggesting that although inhibition per unit area is modest, the restriction of colony growth has led to a significant reduction in the number of spores of the pathogen. Penicillium expansum (MR092), Diaporthe foeniculina (MR099), and Cladosporium sp. (MR101) resulted in higher total conidia production per colony than in the control group (MR007), ranging from 2.6 × 107 to 3.9 × 107 conidia per colony (Table 3), with a relative inhibition of −274.4% to −455.7%. In these cases, both the production of conidia per unit area and the total number of conidia were higher than in the controls, and sporulation of C. godetiae was stimulated when in contact with these isolates. The results show that the most effective antagonists are predominantly concentrated in the Sordariomycetes class, particularly in the Hypocreaceae (Trichoderma spp.), Bionectriaceae (Clonostachys chloroleuca), and Sordariaceae (Neurospora intermedia). Fungi in these families showed a high rate of inhibition of C. godetiae mycelial growth, as well as a decrease in sporulation. Conversely, several species from the classes Eurotiomycetes and Dothideomycetes showed less promising results of inhibition.
Concerning inhibition of conidia production, isolates of MR098 (Stemphylium vesicarium), MR149 (Talaromyces amestolkiae), and MR153 (Clonostachys chloroleuca) decreased C. godetiae sporulation by 96.13%, 94.18%, and 92.40%, respectively. Interestingly, this effect occurred despite the low efficacy of the three fungi in inhibiting mycelial growth (22.87%, 11.32%, and 9.74%, respectively). Thus, spore production capacity was significantly reduced even during colony growth, and reached a level of inhibition similar to that obtained by Trichoderma viridescens, which combined high mycelial inhibition with a profound reduction of sporulation (Table 3). The species Cladosporium sp., Diaporthe foeniculina, and Penicillium expansum showed weak mycelial inhibition (less than 33%), and no reduction of sporulation, indicating negative inhibition of sporulation by C. godetiae.
Macroscopic analysis of colony inhibition throughout the dual culture assay enabled grouping the tested isolates according to the morphological changes observed in the Colletotrichum colonies (Table 3). A gradual darkening of the pathogen colonies was observed in the area near contact for six isolates, namely MR092, MR094, MR096, MR106, MR108, and MR137 (Fig. 2b). The pathogen and isolate MR149 produced a reddish colouration in the area near contact in both colonies (Fig. 2e). These contrasted with the negative control, which showed a uniform beige colouration at the end of the test. The colouration of the remaining isolates was similar throughout the test. Regarding colony growth, several antagonists, predominantly in the Sordariomycetes, were found to have a more rapid development than C. godetiae, thereby limiting the growth of the pathogen colony. This behaviour was also observed in Fusarium spp. (MR089, MR094, and MR134), Trichoderma spp. (MR093 and MR142), Diaporthe foeniculina, Cytospora italica, and Neopestalotiopsis elaeagni (Fig. 2a & b). Among these, Trichoderma viridescens stood out because it grew faster than C. godetiae and occupied the entire plate at the end of the experiment. The remaining isolates showed similar growth, with the two fungi accompanying each other and occupying similar portions of the plate. Considering Boddy[59] evaluation criteria, it was possible to observe four different interactions between the pathogen and the species being tested, with distance inhibition (2), and excessive growth (3) being the most recurrent, followed by contact inhibition (1), and mycelium replacement (4) (Table 3, Fig. 2).
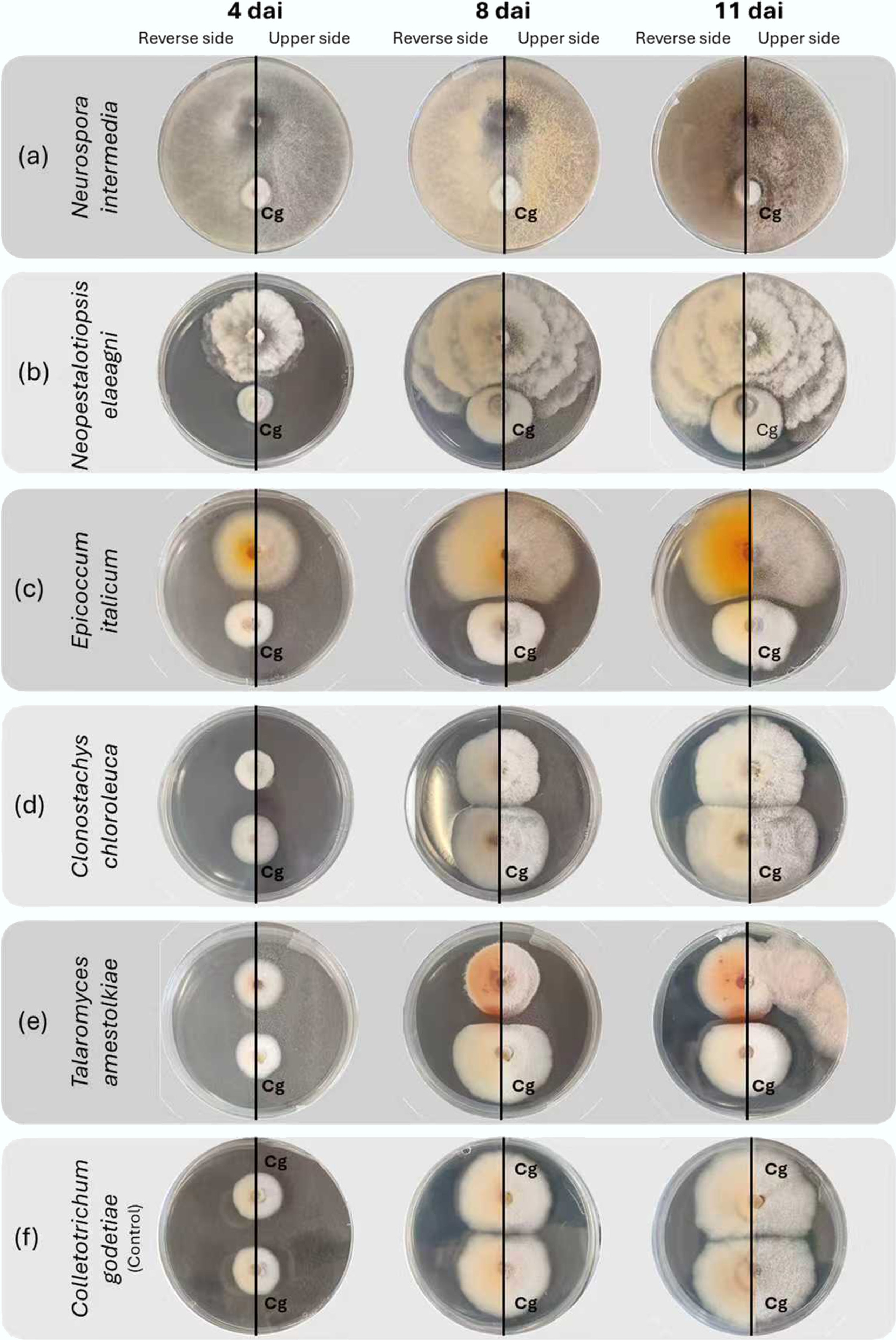
Figure 2.
Evolution of interactions on potato dextrose agar between Colletotrichum godetiae and fungal antagonists obtained from almond trees, 4, 8, and 11 days after inoculation (dai). The fungal isolates to be tested are located at the top of the Petri dishes, while Colletotrichum godetiae (Cg) is located at the bottom. Excessive growth of (a) Neurospora intermedia mycelium on C. godetiae is observed; excessive growth of (b) Neopestalotiopsis elaeagni on C. godetiae by contact inhibition, without the formation of a visible inhibition zone; overgrowth of (c) Epicoccum italicum mycelium on C. godetiae, creating a dividing line without contact between the colonies; C. godetiae mycelium is replaced by (d) Clonostachys chloroleuca; the growth of (e) Talaromyces amestolkiae and C. godetiae stops at the contact line of the mycelium of both colonies, causing a reddish discolouration at the contact edge of C. godetiae; (f) Colletotrichum godetiae in the control culture.
-
This research is an effort to identify and characterise the diversity of endophytic and epiphytic fungi associated with Prunus dulcis, and to evaluate their biocontrol potential against C. godetiae, the main pathogen of almond anthracnose. The results obtained demonstrate a high diversity of fungal genera associated with various plant materials and cultivars. Dual culture assays allowed the identification of fungal genera with a significant inhibitory effect on C. godetiae development and sporulation.
The survey of the genetic diversity of fungi in almond trees revealed that branches and fruits were the plant tissues with the greatest richness and diversity of endophytic fungi, characterized by higher Shannon-Wiener indices, indicating greater structural and ecological stability. These findings indicate that perennial tissues, such as branches, provide better protection against environmental changes, allowing for the establishment of long-lived communities of fungi. Flowers and leaves, on the other hand, represented a more superficial community of fungi, subject to more abiotic variation and containing a higher proportion of endophytic and epiphytic fungi, such as Cladosporium and Alternaria. Similar patterns have been observed in other woody hosts, such as olive trees, where the anthracnose pathogen persists more stably in the branches during the low disease season, while inoculum levels on leaves, although higher initially, decrease more rapidly during the off-season[60].
The high frequency of colonisation recorded in almonds by endophytic fungi (92.8%) suggests that the fruits constitute a distinct reservoir, possibly influenced by physical and chemical barriers, such as the presence of pubescent exocarp and the production of volatile terpene compounds with antifungal properties[61,62]. These traits may limit epiphytic colonisation but favour internally adapted endophytes. It is important to further investigate the relationship between the diversity of fungi and the organs of plants, as each organ represents a micro-environment with different physiological and chemical characteristics. This pattern has already been observed in other woody species, such as olive and oak trees, where it has been shown that the plant organ significantly affects the composition of the endophytic fungal community[63]. Understanding these ecological differences at the organ level has practical implications for the management of orchards. If fruit and branches are key reservoirs for endophytes, including potentially beneficial antagonists and latent pathogens, management strategies could focus on these organs for monitoring, selective sampling, or even inoculation with candidate biocontrol agents at times when stable colonisation is most likely to occur.
Fungal diversity varied noticeably among the cultivars studied. The cultivars 'Lauranne', 'Soleta', and 'Belona' showed higher diversity values and total number of fungal colonies than the other cultivars, with a predominance of the genera Trichoderma, Aureobasidium, and Epicoccum. The microbiota composition can be influenced by genetic factors of the host plant, such as the composition of exudates and cuticle thickness, as well as abiotic factors, such as temperature variation, relative humidity, exposure to sunlight, and orchard management, among others[16]. However, when comparing isolates obtained with and without surface disinfection, it was observed that while disinfection favours the recovery of endophytes, most fungal genera were detected in both methods. Of the 39 fungal genera identified, only Acremonium, Clonostachys, Lambertella, Meyerozyma, Mucor, Neopestalotiopsis, Stemphylium, and Talaromyces were exclusive to one of the methods. Thus, 30 genera demonstrated the ability to colonise both the surface and interior of plant tissues. This 'dual habitat' behaviour indicates that some species possess sufficient ecological plasticity to alternate between an epiphytic and an endophytic life-cycle[14], a pattern that should be confirmed in future studies using complementary molecular and morphological approaches (e.g., qPCR or metabarcoding) to identify ecological niches more accurately. Among the 24 isolates tested in vitro against C. godetiae, Trichoderma viridescens, T. citrinoviride, and Neurospora intermedia showed the highest percentages of inhibition, both in terms of mycelial growth and sporulation reduction capacity. These results may be attributed to multiple antagonistic mechanisms. In the case of Trichoderma spp. isolates, the most frequently reported mechanism is mycoparasitism, which involves competition for space and nutrients, contact and overlap of the pathogen's mycelium leading to cell lysis, and the production of volatile antifungal compounds and hydrolytic enzymes (chitinase, cellulase, and xylanase)[64]. In natural conditions, these mechanisms may also act in plant tissues, although their efficacy depends on the ability of the antagonist to colonise the niche occupied by the pathogen. Trichoderma spp. are filamentous fungi commonly found in soil ecosystems and plant tissues, and species such as T. harzianum, T. citrinoviride, T. koningii, T. pseudokoningii, and T. víride have been reported as biocontrol agents[33,34]. Several species of the genus Trichoderma promote increased nutrient absorption, growth, and induction of systemic resistance in plants[65], as well as control of pathogens, such as Fusarium oxysporum, Rhizoctonia solani, Pythium aphanidermatum, Alternaria alternata, Macrophomina phaseolina, Colletotrichum acutatum, and Colletotrichum gloeosporioides[27,64,66]. Currently, over 60% of globally homologated biofungicides are produced from Trichoderma spp. formulations, which are the most effective biofungicides for use in agriculture[65]. Neurospora intermedia is another species with strong antagonistic performance. The genus Neurospora is known as a model organism in fungal genetics and physiology due to its rapid growth on various substrates[67]. Some Neurospora species can synthesize pigments, volatile antimicrobial, aromatic, and bioactive compounds[68,69]. Furthermore, a previous study verified the antagonistic potential of N. sitophila against Botrytis cinerea due to the production of phenolic compounds and volatile organic compounds[70].
Stemphylium vesicarium, Clonostachys chloroleuca, and Talaromyces amestolkiae have a high inhibitory capacity on C. godetiae sporulation but have a limited effect on mycelial inhibition. This discrepancy does not distinguish them from isolates with higher overall efficacy, such as Trichoderma viridescens, which was able to suppress the growth and sporulation of colonies. This separation of growth inhibition and reproductive inhibition is of relevance as it suggests that the mechanisms involved may be acting specifically on conidiogenesis. In Clonostachys spp., it is known that many species secrete a variety of enzymes that degrade the cell wall (e.g., chitinases, β-1,3-glucanases), and produce antifungal metabolites that interfere with the cell-wall repair and regulatory processes of the fungal reproductive system[71]. These metabolites may have a disproportionate influence on the formation of conidia, even without causing a significant reduction in mycelial growth. Similarly, a few species of Talaromyces spp. are recognised for the formation of bioactive compounds, including polyketides, terpenes and unstable metabolites, that may modulate conidiogenesis or interfere with signalling pathways involved in the reproductive differentiation of pathogens[72]. The high reduction of sporulation observed in T. amestolkiae may therefore be due to the secretion of specific metabolites which inhibit the major stages of conidiophore formation, or to intense nutrient-competition mechanisms that penalize reproductive rather than vegetative growth.
Neopestalotiopsis elaeagni, Fusarium annulatum, Alternaria sp. and the isolate belonging to the Fusarium sambucinum species complex show intermediate levels of average mycelial growth inhibition of C. godetiae, with a range of between 47.95% and 31.27%. These results indicate some capacity to limit the expansion of the pathogen. However, unlike Trichoderma viridescens, which drastically reduced the production of conidia (96.1% inhibition), these isolates did not significantly reduce sporulation, with the total number of conidia remaining high. This difference indicates that reduction of mycelial growth does not necessarily result in a reduction of pathogen reproduction, suggesting that different antagonism mechanisms, such as competition for space and nutrients or production of secondary metabolites, may be acting differently on growth and sporulation[73,74]. This separation between mycelial growth inhibition and reproductive inhibition indicates that the efficacy of the antagonist should not be assessed solely on the size of the fungal colony, but also on the impact on the pathogen's ability to reproduce, as reflected in the formation of conidia.
Penicillium expansum, Diaporthe foeniculina, and Cladosporium sp. not only failed to inhibit C. godetiae mycelial growth and sporulation, but also stimulated the production of conidia, resulting in absolute values significantly higher than control values. Similar phenomena have been described in microbial communities in other crops where certain fungi act as 'pathogen facilitators'[75]. These microorganisms can change the environment of a plant by providing nutrients, altering chemical signals, or suppressing defence mechanisms, leading to more efficient colonisation by pathogenic fungi. For instance, Neofabraea sp. and Neofabraea vagabunda were identified as being positively correlated with the abundance of Colletotrichum in olive trees[75]. The sporulation stimulation of C. godetiae by Cladosporium sp., Diaporthe foeniculina, and Penicillium expansum is therefore consistent with this idea and suggests that these isolates may create conditions favourable to the development of the pathogen rather than compete with it.
It should be noted that the dual culture antagonism tests were performed on PDA, a widely used but artificial in vitro medium[14,26,27,31,37,58]. Although suitable for preliminary screening, PDA does not reproduce the biochemical or structural conditions of plant tissue. Therefore, future studies should include plant-based media, separate tissue, or plant tests to confirm that the observed antagonistic effects can be reproduced under realistic environmental conditions.
The macroscopic observations made were consistent with the situations of interaction between two fungi, as described by Boddy[59]. In contact inhibition, where there is interaction between the hyphae of the two fungi, this can lead to cell death in the interaction zone or to the production of melanin, which is a mechanism for protecting cells against the antagonistic microorganism[76]. These processes may explain the darkening of C. godetiae colonies when in contact with the mycelium of Apiospora kogelbergensis, an isolate of Fusarium sambucinum species complex, Nothophoma spiraeae and Aspergillus sp. colonies. On the other hand, darkening of C. godetiae was also observed because of the inhibition of Penicillium expansum, probably associated with the production of volatile compounds[59]. Species such as Aspergillus niger, Aureobasidium pullulans, and Epicoccum layuense are known for their inhibitory effects on Colletotrichum acutatum[77], Fusarium oxysporum, Macrophomina phaseolina, Pythium aphanidermatum[78], Rhizoctonia solani[79], Phaeomoniella clamydospora, and Phaeoacremonium minimum[58]. However, the mechanisms underlying this stimulation remain unclear, and future work should test metabolite extracts or controlled cultures to determine whether the reduction is due to nutrient leakage, volatile compounds, or altered signalling pathways.
In the characterisation of endophytic and epiphytic species, several species potentially pathogenic to almond trees were detected in plant tissues. Among these are Colletotrichum sp., Botryosphaeria sp., Cytospora sp., Botrytis cinerea, Rhizopus sp., Alternaria sp., Aspergillus sp., Fusarium sp., and Monilinia sp.[80]. Although some of these species have also been tested in the dual culture to assess whether they may have antagonistic activity against C. godetiae, their potential use as biological control agents should be carefully considered. Fungus species such as Fusarium spp., Cytospora italica, Alternaria vitis, and Aspergillus sp. did not inhibit mycelial growth or C. godetiae conidia effectively, but more importantly, their well-documented pathogenicity and toxin-producing potential raise serious biosafety concerns[81,82]. Even if antagonistic metabolites were identified, the development of these taxa as biocontrol agents would be limited by the risk of host plant infection, disruption of the established balance of the resident microbial communities, or unintended environmental effects. Even if the discovery of antifungal compounds from these isolates may contribute to future disease control strategies, such applications would require the purification and biochemical characterisation of the compound rather than the introduction of the organisms themselves[83].
-
This study demonstrated the diversity of the endophytic and epiphytic community of fungi in almond crops, including 39 genera distributed in different plant organs and cultivars. The integrated isolation approach, with and without disinfection, allowed us to distinguish between endophytic fungi and the combination of endophytes and epiphytes, and thus a more complete picture of the fungal community in the almond orchard. The results indicate that the main reservoirs of endophytic fungi are branches and fruits, with particular emphasis on the specificities of the endophytic community of almonds. In the meantime, flowers and leaves have shown more epiphytic diversity. The cultivars 'Lauranne', 'Soleta', and 'Belona' showed a higher fungal diversity, indicating that host genotype influences community structure.
In vitro assays identified Trichoderma viridescens, Neurospora intermedia, and T. citrinoviride as the most effective antagonists of C. godetiae, indicating the action of multiple biocontrol mechanisms, such as mycoparasitism, antibiosis, and competition for resources. Other species, such as Clonostachys chloroleuca and Talaromyces amestolkiae, showed partial antagonistic potential due to inhibition of pathogen sporulation, suggesting indirect modes of action.
These findings provide the basis for future development of native biocontrol agents adapted to almond orchards. However, the transition from laboratory testing to commercial formulations requires further steps, including field validation, environmental performance assessment, and a thorough safety assessment, in particular with regard to toxin formation, effects on non-target microorganisms, and compatibility with current agricultural practices.
Overall, this study has advanced the knowledge of the almond mycobiome, identified fungal isolates with promising biocontrol potential, and contributed to the development of sustainable disease management strategies in line with the principles of sustainable agriculture and food security.
-
It accompanies this paper at: https://doi.org/10.48130/aee-0025-0015.
-
The authors confirm their contributions to the paper as follows: Madalena Ramos: study conception and design; material preparation; data collection and analysis; first draft of the manuscript; response to reviewers; review and approval of the final version of the manuscript. Pedro Talhinhas: study conception and design; material preparation; data collection and analysis; supervision and funding acquisition; response to reviewers; review and approval of the final version of the manuscript. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets used or analyzed during the current study are available from the corresponding author upon reasonable request.
-
This work was funded by FCT – Fundação para a Ciência e a Tecnologia, I.P. (Grant No. 2021.05854.BD), and FCT – Fundação para a Ciência e Tecnologia, I.P. through project UID/04129/2025 (https://doi.org/10.54499/UID/04129/2025) of LEAF-Linking Landscape, Environment, Agriculture, and Food.
-
The authors declare that they have no conflict of interest.
-
Endophytic and epiphytic fungal communities were identified in almond orchards.
Almond tree branches and fruits were the main reservoirs of endophytic fungi.
Trichoderma viridescens strongly inhibited the growth and sporulation of C. godetiae.
Fungal isolates showed potential as biocontrol agents for almond anthracnose.
-
Full list of author information is available at the end of the article.
- The supplementary files can be downloaded from here.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ramos M, Talhinhas P. 2026. Selection of fungi derived from almond orchards for biological control of almond anthracnose caused by Colletotrichum godetiae. Agricultural Ecology and Environment 2: e002 doi: 10.48130/aee-0025-0015 

Selection of fungi derived from almond orchards for biological control of almond anthracnose caused by Colletotrichum godetiae
- Received: 09 November 2025
- Revised: 10 December 2025
- Accepted: 25 December 2025
- Published online: 20 January 2026
Abstract: Almond production in Mediterranean regions has expanded in recent years, accompanied by an increasing incidence of anthracnose, caused mainly by Colletotrichum godetiae. Due to the limitations and environmental impacts of chemical control methods, biological strategies utilizing native microbial communities have emerged as promising alternatives. In this study, the Prunus dulcis fungal community is characterized and the antagonistic potential of selected isolates evaluated against C. godetiae. The plant material was collected from 16 almond cultivars. The fungi were isolated using disinfected and non-disinfected tissues to differentiate endophytic from epiphytic fungi. A total of 39 fungal genera were identified, the most common being Alternaria, Cladosporium, and Trichoderma. Fungal diversity varied across plant organs, with branches and fruits showing the highest rates of endophytic colonisation. Twenty-four isolates, selected among the most abundant species, were tested for antagonistic activity in vitro using dual culture assays. Trichoderma spp. and Neurospora intermedia proved to be efficient in inhibiting mycelial growth (> 75%), and reducing sporulation of C. godetiae. The antagonistic mechanisms observed included overgrowth, contact inhibition, and antibiosis. Other species, such as Talaromyces amestolkiae and Clonostachys chloroleuca, showed low inhibition of the mycelium but effectively reduced conidia production. The effectiveness of the endophytic isolates highlights their adaptation to the host environment and their potential as biological control agents. The results suggest that almond trees have a diverse mycobiota with biocontrol potential, supporting the further development of fungal isolates, especially of Trichoderma viridescens, T. citrinoviride, and Neurospora intermedia, as environmentally sustainable alternatives to chemical fungicides for managing almond anthracnose.
-
Key words:
- Prunus dulcis /
- Endophytes /
- Epiphytes /
- Antagonism tests /
- Fungal communities