









-
Lysozyme, a ubiquitous bacteriolytic enzyme found across a wide range of organisms[1], facilitates the hydrolysis of the β-(1,4)-glycosidic bond linking N-acetylglucosamine (NAG) and N-acetylmuramic acid (NAM) in peptidoglycan (PGN), leading to the lysis of bacterial cells[2,3]. Lysozymes are divided into six distinct categories based on variations in their structural, catalytic, and immunological properties. Within vertebrate animals, two types of lysozymes are recognized: chicken-type (c-type) and goose-type (g-type)[2]. These two types vary in amino acid sequence and enzymatic characteristics[2]. Typically, g-type lysozymes are larger (ranging from 20 to 22 kDa) compared to c-type lysozymes (which weigh about 11 to 15 kDa) and lack a signal peptide[2].
G-type lysozyme first recognized as an antimicrobial agent in the egg whites of various bird species has since been discovered across a broad range of vertebrates, spanning mammals, reptiles, and fish[4]. The initial identification and study of g-type lysozyme in fish occurred in Paralichthys olivaceus[5]. Later, this type of lysozyme has been cloned in other teleosts, including Gadus morhua L.[6], Ctenopharyngodon idellus[7], Scophthalmus maximus[8], Ictalurus punctatus[9], Dicentrarchus labrax L.[10], Microptenus salmoides[11], Labeo rohita[12], Totoaba macdonaldi[13], and Trachidermus fasciatus[14]. In fish, g-type lysozyme is regarded as a crucial part of the immune system, demonstrating potent activity against both Gram-positive and Gram-negative bacteria, such as Micrococcus luteus[8], Streptococcus iniae[15], Vibrio harveyi[16], Vibrio alginolyticus[17], Photobacterium damselae[10], and Aeromonas hydrophila[11]. Apart from their antimicrobial activities, g-type lysozymes could also perform many other functions, including against viral particles and parasites[17], immunomodulatory[18], anti-inflammatory[14], and regulation of early development[3]. However, as of now, there is limited information available concerning the molecular characteristics and functions of g-type lysozymes in pufferfish.
Obscure puffer Takifugu obscurus is a marine species found in the East China Sea, Yellow Sea, and Bohai Sea[19]. Recently, T. obscurus has gained significance as a highly valuable aquaculture species. However, its breeding has been hindered by a variety of infectious diseases, resulting in substantial economic losses. Consequently, research into the molecular mechanisms underpinning the host's immune defense against pathogens has garnered increasing attention. Our previous work cloned and identified a c-type lysozyme (ToLyzC) in T. obscurus, providing compelling evidence for its pivotal role in the antimicrobial immunity of this species[20]. Building on this foundation, the current research aimed to clone a novel g-type lysozyme of T. obscurus (named ToLyzG), and to analyze its expression and antibacterial activities.
-
V. harveyi was cultivated in tryptic soy broth (TSB) and incubated overnight at 28 °C. A. hydrophila was inoculated into Luria-Bertani (LB) broth and cultured at 28 °C. Vibrio parahaemolyticus was cultured in an LB medium containing 3% NaCl and grown at 37 °C. Staphylococcus aureus, Bacillus subtilis, and Bacillus thuringiensis were all cultivated in LB medium at 37 °C. Two hundred T. obscurus (body size of ~12 cm) were obtained from a pufferfish breeding base in Nantong, China, and kept at 23 °C in freshwater. The pufferfish were adapted to lab conditions for10 d before experimental interventions.
Cloning of ToLyzG
-
A coding region sequence homologous to lysozyme was retrieved from a high-throughput transcriptome data of T. obscurus and named ToLyzG. Two primers (ToLyzG-F and ToLyzG-R, Table 1) were designed to verify the nucleotide sequence accuracy. The PCR products were subcloned into the pEasy-T3 vector and underwent sequence analysis at Nanjing Springen, China.
Table 1. Primers used in this study.
Primers Nucleotide sequence (5'-3') Tm value
(°C)ToLyzG-F CAAGCCACCGAAATCCTGGTTGAGTT 70.5 ToLyzG-R GACATTTTTCTCACCCTTATTGTAGGCAGC 69.0 ToLyzG-qF CACCGAAATCCTGGTTGAGT 57.2 ToLyzG-qR CACCCTTATTGTAGGCAGCTAA 57.2 β-actin-qF GACAGCTATGTGGGAGATGAAG 56.5 β-actin-qR GGAGCTCGTTATAGAAGGTGTG 56.3 ToLyzG-exF GCCATGGCTGATATCGGATCCATGCCTTACGGAAAGATAGAGGATATA 84.6 ToLyzG-exR ACGGAGCTCGAATTCGGATCCAAACCCCATGTTGCTTTTGTACCACTG 89.5 Sequence analysis
-
The ToLyzG sequence was analyzed utilizing the BLAST program (
http://blast.ncbi.nlm.nih.gov/Blast.cgi )[21] and the ExPASy platform (https://web.expasy.org/translate/ )[22]. A domain search was performed with SMART (http://smart.embl-heidelberg.de/ )[23] and visualized using PyMOL 2.5. Three-dimensional structural models were constructed using the SWISS-MODEL (https://swissmodel.expasy.org/ )[24]. The molecular weight (Mw), isoelectric point (pI), and instability index were calculated with the ProtParam (https://web.expasy.org/protparam/ )[22]. Multiple sequence alignment was generated using the Clustal Omega (www.ebi.ac.uk/Tools/msa/clustalo/ )[25]and the GENEDOC software. Based on this alignment, a phylogenetic tree was developed using the neighbor-joining (NJ) algorithm in MEGA 7.0 software, and 1000 bootstraps were applied to assess reliability[26].Bacterial infections and sample collection
-
To investigate the expression pattern of ToLyzG, three untreated pufferfish were dissected, and the liver, spleen, kidney, gills, heart, and intestine were harvested. These three untreated pufferfish were of similar age and weight, ensuring that any observed differences in the study outcomes could not be attributed to variations in these factors. In the immune challenge experiment, 90 T. obscurus were randomly assigned to three groups. In two challenge groups, pufferfish were injected with 100 μL of V. harveyi or S. aureus (about 3.0 × 107 CFU/mL). In the control group, fish were injected with 100 μL of sterile PBS. At 0, 12, 24, 36, 48, and 72 h post-injection, the liver, spleen, kidney, and intestine tissues were sampled from three pufferfish. All isolated tissue samples were snap-frozen in liquid nitrogen and kept at −80 °C for future use.
RNA extraction and cDNA synthesis
-
Total RNA was extracted from the above samples utilizing TRIzol Reagent. To ensure the absence of DNA contamination, DNase I (Promega, USA) was incorporated. The integrity and purity of RNA were assessed through electrophoresis on a 1.5% agarose gel, while its concentration was measured spectrophotometrically at 260 nm. Then, the first-strand cDNA was synthesized using 2 μg of RNA with the PrimeScriptTM RT reagent kit (TaKaRa, Japan). Following synthesis, the cDNA was diluted fivefold with ddH2O and stored at −20 °C for future use.
Expression pattern analysis
-
To quantify the expression of ToLyzG across various pufferfish tissues, quantitative real-time PCR (RT-qPCR) was conducted, utilizing β-actin as the housekeeping gene for normalization. Gene-specific primers (ToLyzG-qF and ToLyzG-qR, β-actin-qF and β-actin-qR) were designed and are listed in Table 1. The RT-qPCR reactions were set up using the SYBR Premix Ex Taq™ II Kit (TaKaRa, Japan). Each 10 μL reaction mixture contained 1 μL of diluted cDNA template, 5 μL of 2× Ex Taq™ SYBR premix, 0.4 μL of each primer, and 3.2 μL ddH2O. The relative expression of ToLyzG mRNA was calculated using the 2−ΔΔCᴛ method[27].
Recombinant expression and purification
-
The coding sequence of ToLyzG was amplified using two primers ToLyzG-exF and ToLyzG-exR (Table 1) with the restriction enzyme site BamH I. The resulting PCR product was purified, digested, and subcloned into the expression plasmid (pET32a, Novagen, Germany). The ligation product was then transformed into Escherichia coli DH5α (TransGen, China), and the positive clone was identified through PCR screening and confirmed by sequencing. Subsequently, the pET32a-ToLyzG plasmid was transformed into E. coli BL21 (DE3) (TransGen, China). Recombinant expression was induced at 37 °C for 5 h by adding isopropyl β-D-1-thiogalactopyranoside (IPTG, 0.5 mM). As a negative control, pET-32a without an insert fragment was used, which expresses a thioredoxin-His-tagged protein (TRX). The recombinant ToLyzG and TRX proteins were detected by 12% SDS-PAGE and purified by His Bind resin chromatography (Novagen, Germany).
Influence of pH and temperature on the lytic activity of rToLyzG
-
To determine the optimal conditions for the lytic activity of rToLyzG, a turbidimetric assay was conducted using Micrococcus lysodeikticus (Sigma, USA) as the substrate. A bacterial suspension with OD600 of 0.3 was prepared in 0.1 M phosphate buffer ranging from pH 5.0 to 8.0, with increments of 0.5. Subsequently, 150 μL aliquots of these suspensions were mixed with 50 μL of rToLyzG (500 μg/mL). The initial OD600 reading denoted as A0 was immediately recorded using a spectrophotometer. The mixtures were then incubated at 37 °C for 30 min, and subsequently cooled on ice to terminate the reaction. The OD600 was measured again, denoted as A. The lytic activity (UL) of the enzyme was calculated using the formula: UL = (A0 − A)/A.
For the determination of the optimal temperature, a mixture of M. lysodeikticus and rToLyzG, adjusted to the optimal pH of 7.0, was incubated in a water bath at temperatures varying from 15 to 50 °C, with increments of 5 °C, for 30 min. The OD600 values were recorded before and after incubation, denoted as A0 and A, respectively. All measurements were performed in triplicate to ensure accuracy and reproducibility.
Bacteriolytic activity of rToLyzG
-
The bacteriolytic spectrum of rToLyzG was evaluated under optimal pH and temperature conditions using a methodology similar to the previous one. Six bacterial strains were selected as test substrates: V. harveyi, V. parahaemolyticus, A. hydrophila, S. aureus, B. subtilis, and B. thuringiensis. The suspension of each bacterium was prepared in 0.1 M phosphate buffer (adjusted to pH 7.0) to achieve an OD at 600 nm of 0.3. One hundred and fifty μL of each suspension was combined with 50 μL of rToLyzG (500 μg/mL) and incubated at 37 °C for 30 min. Absorbance measurements were taken before (A0) and after (A) the incubation period. The lytic activity of the enzyme (UL) was calculated.
Polysaccharide binding assay
-
To evaluate the binding affinity of rToLyzG to lipopolysaccharide (LPS) or peptidoglycan (PGN) sourced from Sigma–Aldrich, USA, an enzyme-linked immunosorbent assay (ELISA) was employed. LPS and PGN were dissolved in ddH2O to 80 μg/mL and then coated onto the wells of a microplate with 50 μL per well. The microplate was placed at 37 °C overnight and incubated at 60 °C for 30 min. Then each well was blocked with 200 μL of 3% bovine serum albumin (BSA) at 37 °C for 1 h. Subsequently, 100 μL of purified rToLyzG or rTRX (0, 3, 5, 8, 15, 25, and 50 μg/mL) were added to each well and incubated for 3 h. One hundred μL of rabbit anti-His antibody, diluted 1:2000 (Abbkine, USA), was then added and incubated for 2 h. One hundred μL of peroxidase-conjugated goat anti-rabbit IgG (1:5000 diluted) was added and incubated for 1 h. After each incubation step, the wells were washed four times with PBS containing 0.5% Tween-20 to remove unbound reagents. To initiate the colorimetric reaction, 0.01% 3,3ʹ,5,5ʹ-tetramethylbenzidine was added to each well. The reaction was subsequently stopped by the addition of 2 M H2SO4, followed by the measurement of absorbance at a wavelength of 450 nm.
Bacterial binding activity of rToLyzG
-
Six bacteria (V. harveyi, V. parahaemolyticus, A. hydrophila, S. aureus, B. subtilis, and B. thuringiensis) were used for the binding assay. These bacteria were harvested, thoroughly washed, and resuspended in PBS to 3.0 × 107 CFU/mL. Subsequently, 500 μL of each suspension was incubated with 500 μL of rToLyzG (500 μg/mL). The mixtures were gently rotated for 45 min to facilitate binding. Following incubation, the samples were centrifuged, and the cells were washed three times with PBS. To elute the bound proteins, a 5% SDS solution was employed. rTRX protein was used as a control. The eluted proteins and the final bacterial pellets were analyzed by Western blot using anti-His antibody.
The interaction between bacteria and proteins was further evaluated using ELISA. One hundred μL of each bacterial suspension was dispensed into a microtiter plate and incubated overnight at 4 °C. Following this, the bacterial cells were fixed using 50 μL of 0.05% glutaraldehyde and subsequently blocked with 100 μL of 3% BSA. Afterward, 100 μL of rToLyzG (100 μg/mL) was added and incubated for 3 h. The wells added with rTRX at the same concentration were employed as control. Following this incubation period, the ELISA protocol continued with subsequent steps.
Bacterial growth inhibition assay
-
To investigate the inhibitory effect of rToLyzG on bacterial growth, V. harveyi, V. parahaemolyticus, A. hydrophila, S. aureus, B. subtilis, and B. thuringiensis were cultivated in TSB or LB medium as previously specified until they reached OD600 of 0.1. Subsequently, rToLyzG was added to these cultures at 100 μg/mL. Under identical conditions, rTRX was utilized as a control. The growth was continued for 8 h, and OD600 was measured every hour with a spectrophotometer.
Statistical analysis
-
All experiments were independently replicated at least three times. Statistical analysis was carried out using GraphPad Prism 9.0 software. The results were reported as the mean ± SD from triplicate experiments. An unpaired t-test was utilized to evaluate differences between the challenged and control groups. Statistical significance was determined when p values were less than 0.05.
-
The cDNA encoding ToLyzG was retrieved from the previously constructed cDNA library and deposited in NCBI GenBank with the accession number PQ633364. The complete coding region sequence of ToLyzG was 573 bp long encoding a 190-amino-acid protein with Mw of 20.88 kDa and theoretical pI of 5.96 (Fig. 1a). In the ToLyzG protein sequence, there were 22 positively charged residues and 25 negatively charged residues. The computed instability index, which was 30.13, classified the protein as stable. Sequence analysis predicted that ToLyzG contained an SLT domain (residues 48–174) with a GLMQ motif in the deduced amino acid sequence. However, no signal peptide was detected. Analysis of the 3D structure of ToLyzG using a g-type lysozyme from Takifugu rubripes (SMTL ID: P61133.1.A) as a template structure showed that the predicted 3D structure of ToLyzG contained eight α-helices (Fig. 1b). Notably, no β-sheet regions were identified within the protein structure.
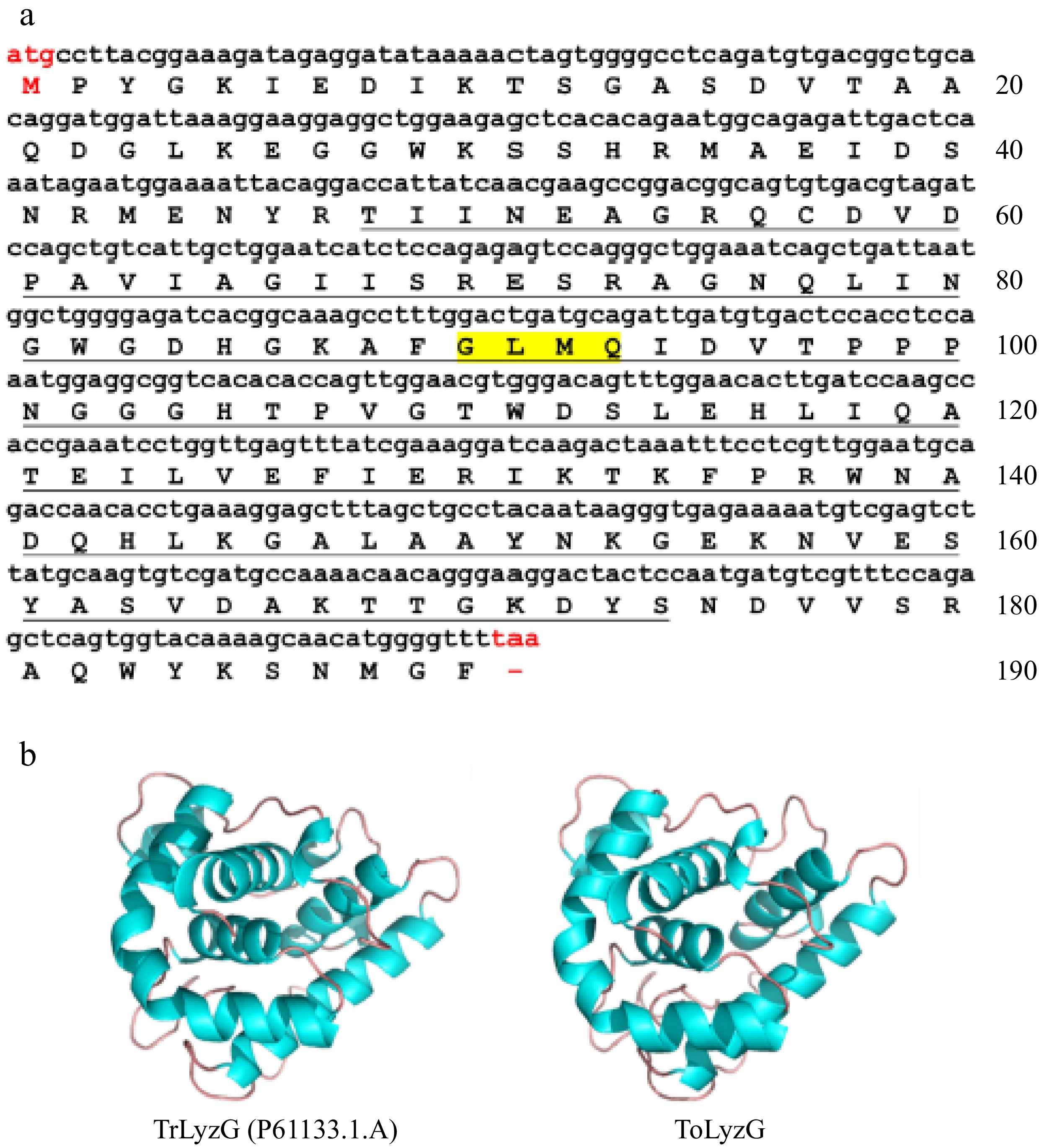
Figure 1.
Bioinformatics analysis of ToLyzG. (a) Nucleotide and amino acid sequence of ToLyzG from T. obscurus. The numbers on the right indicate the total number of amino acids in each line, with the start codon (ATG) and stop codon (TAA) highlighted in red. The predicted SLT domain is underlined, and the GLMQ motif is shaded in yellow. (b) 3D structure prediction of ToLyzG. The blue spirals depict the helical regions, while the brown lines signify the coiled regions.
Bioinformatics analysis of ToLyzG
-
The analysis by BLASTP search indicated that compared to g-type lysozymes of other fish species, ToLyzG was more identical with g-type lysozymes from other puffer species such as T. rubripes (99.47%) and Takifugu flavidus (97.37%). It also shared sequence homology with g-type lysozymes of Tetraodon nigroviridis (69.78%) and Takifugu bimaculatus (69.23%). Multiple sequence alignments further revealed that g-type lysozymes from different puffer species were highly conserved, especially in the SLT domain (Fig. 2). The phylogenetic tree depicted in Fig. 3 was constructed using g-type lysozymes from several known fish species. This tree revealed the presence of two primary branches: one branch contained seven g-type lysozymes from five species of pufferfish, and the other branch comprised of g-type lysozymes from the remaining fish. ToLyzG exhibited the closest genetic relationship to g-type lysozyme derived from T. rubripes.

Figure 2.
Comparison of the amino acid sequences of ToLyzG with known g-type lysozymes in pufferfish. The red boxes represent the SLT domains.
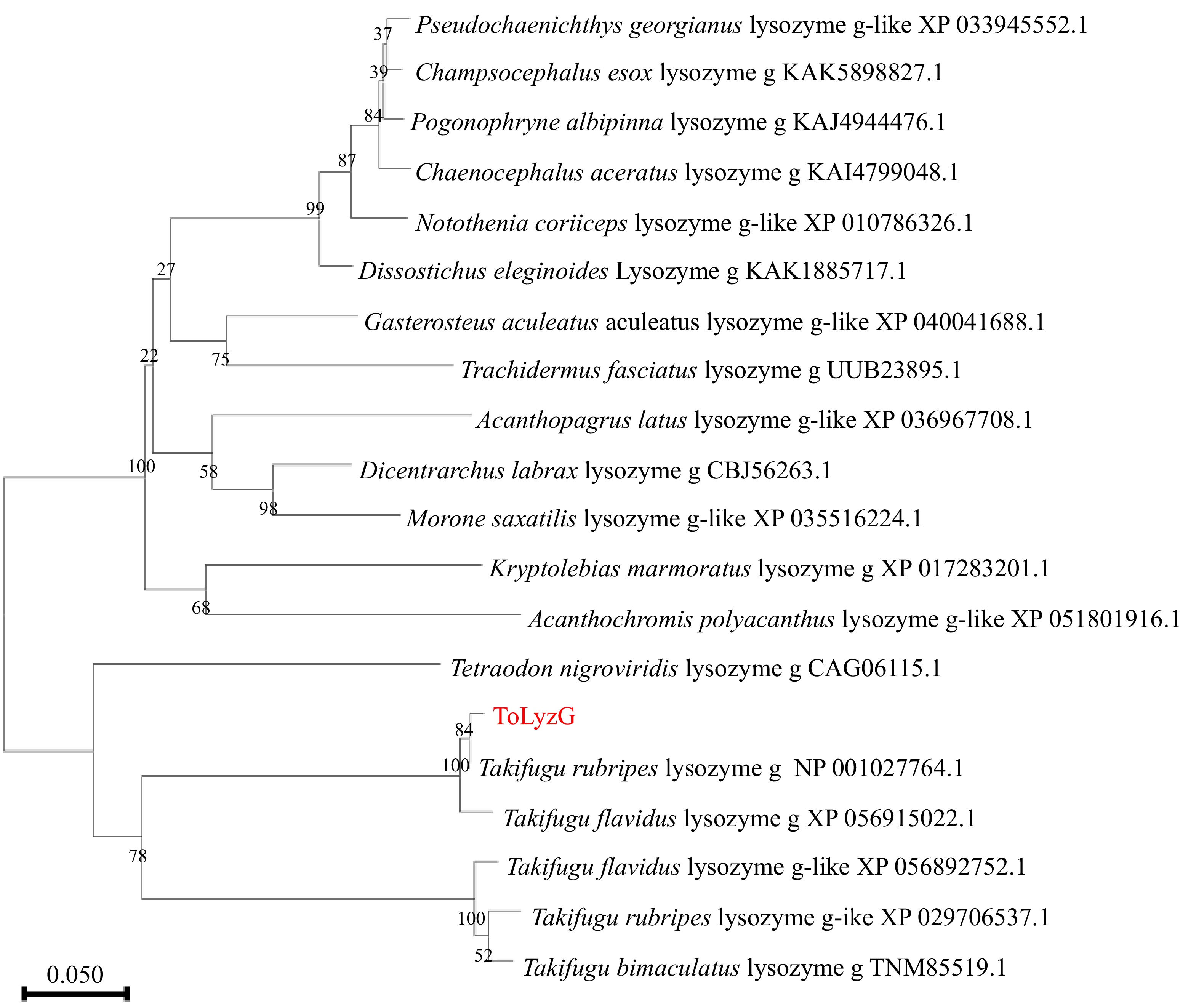
Figure 3.
Phylogenetic tree of g-type lysozymes from different organisms. The evolutionary history of the analyzed taxa was represented by a bootstrap consensus tree inferred from 1000 replicates. The bootstrap values are indicated at the branch points (forks) of the tree. Follow the branches, the species names, and their corresponding accession numbers are provided.
Tissue distribution of ToLyzG
-
Results from RT-qPCR revealed the expression pattern of ToLyzG mRNA across various normal tissues. Notably, the highest levels of ToLyzG transcripts were detected in the liver, followed by the kidney, intestine, spleen, and gills. Conversely, the lowest expression levels were detected in the muscle and heart (Fig. 4).
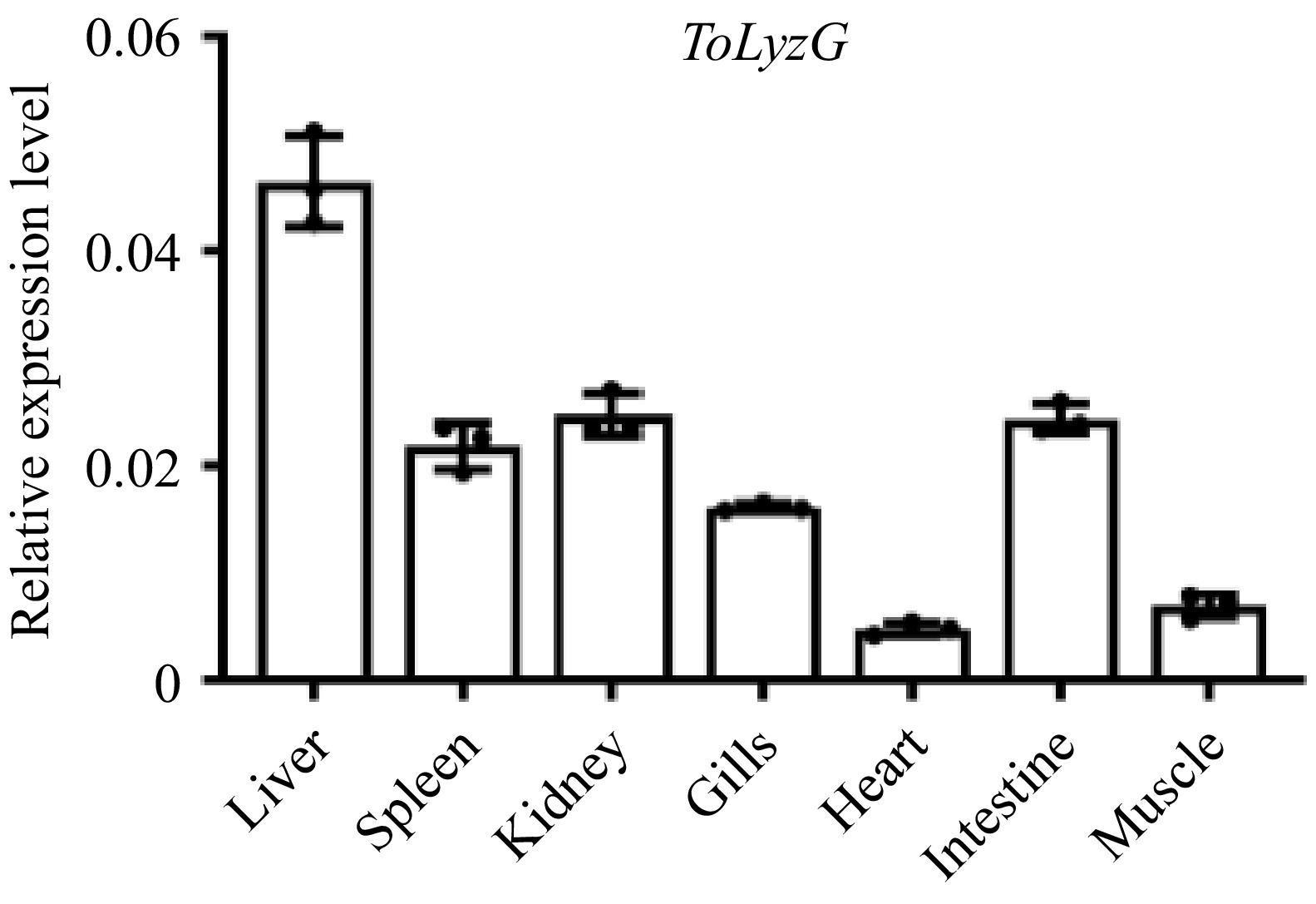
Figure 4.
Tissue distribution of ToLyzG. The expression of ToLyzG gene in six tissue samples of T. obscurus was quantified using RT-qPCR. β-actin served as the internal reference gene for normalization. Data are presented as the mean ± SD (n = 3).
Expression profiles of ToLyzG upon bacterial infections
-
The expression patterns of ToLyzG in four immune-related tissues after V. harveyi or S. aureus challenge were investigated by qPCR. After infection with V. harveyi, ToLyzG in the liver showed significant upregulation, as compared to control, with a peak (6.35-fold, p < 0.01) at 36 h post-infection (hpi) (Fig. 5a). The expression of ToLyzG in the spleen and intestine showed consistent and heightened upregulation from 12 to 48 hpi, reaching a peak at 48 hpi (9.28- and 9.11-fold, p < 0.001, Fig. 5b & d), followed by a marked decrease at 72 hpi. In the kidney, ToLyzG expression increased rapidly at 12 hpi (5.96-fold, p < 0.001, Fig. 5c), followed by a decline at later time points. Upon S. aureus stimulation, ToLyzG mRNA in the liver and intestine was upregulated only at 12 hpi (1.34- and 1.18-fold, p < 0.05, Figs. 6a & d). No obvious changes in ToLyzG expression were observed in the spleen and kidney, only slightly upregulated at 36 hpi (1.54-fold, p < 0.05, Fig. 6b) and 48 hpi (1.70-fold, p < 0.05, Fig. 6c), respectively. There were no notable alterations in ToLyzG expression observed in the PBS-injected group over the period from 0 to 72 h.
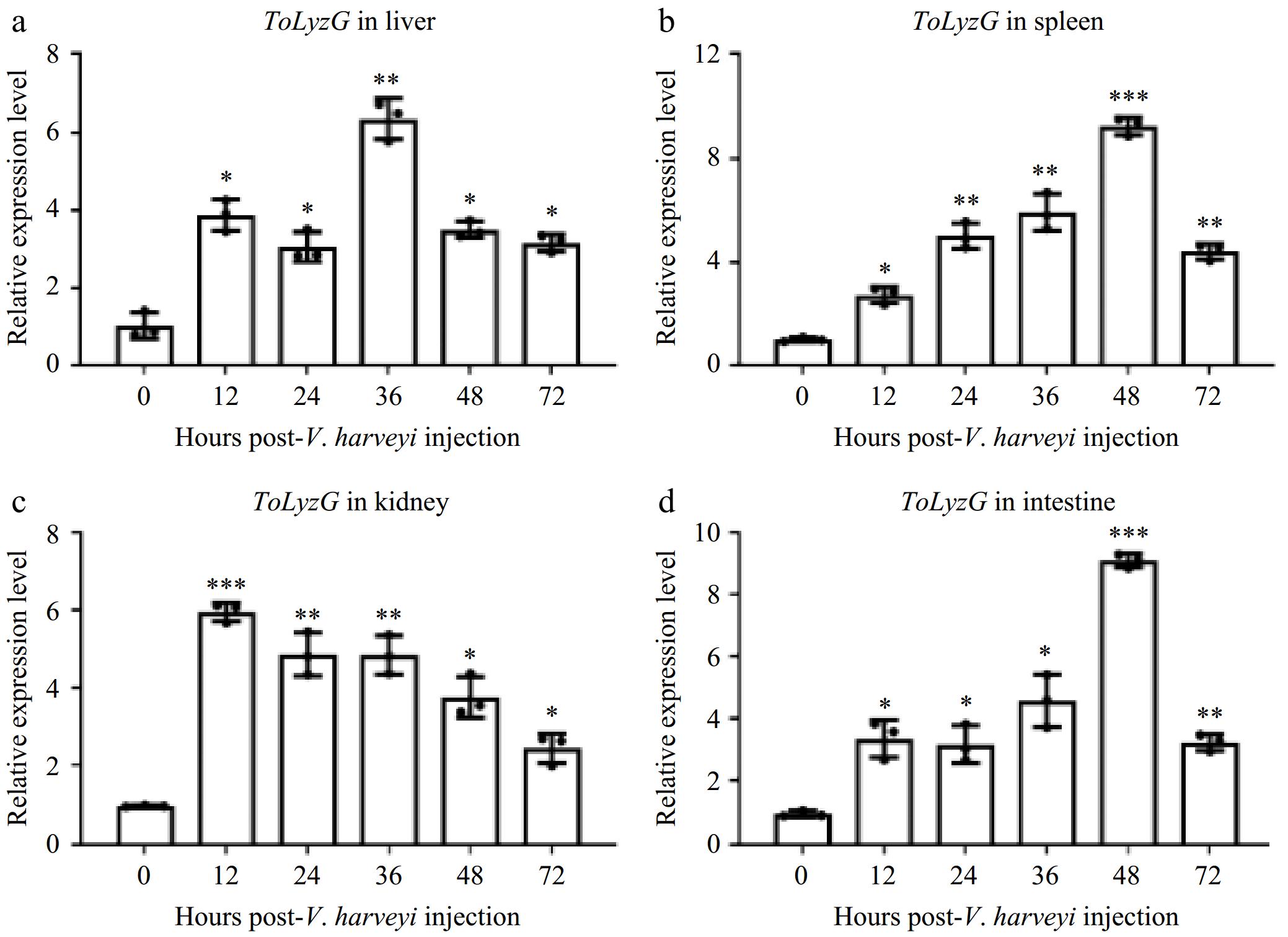
Figure 5.
ToLyzG mRNA expression after V. harveyi challenge. Different time courses of ToLyzG expression in the (a) liver, (b) spleen, (c) kidney, and (d) intestine at 0, 12, 24, 36, 48, and 72 h post-injection with V. harveyi was analyzed by RT-qPCR. Data are shown as an average of three independent replicates ± SD. Significant differences are indicated by asterisks, * p < 0.05, ** p < 0.01, *** p < 0.001.
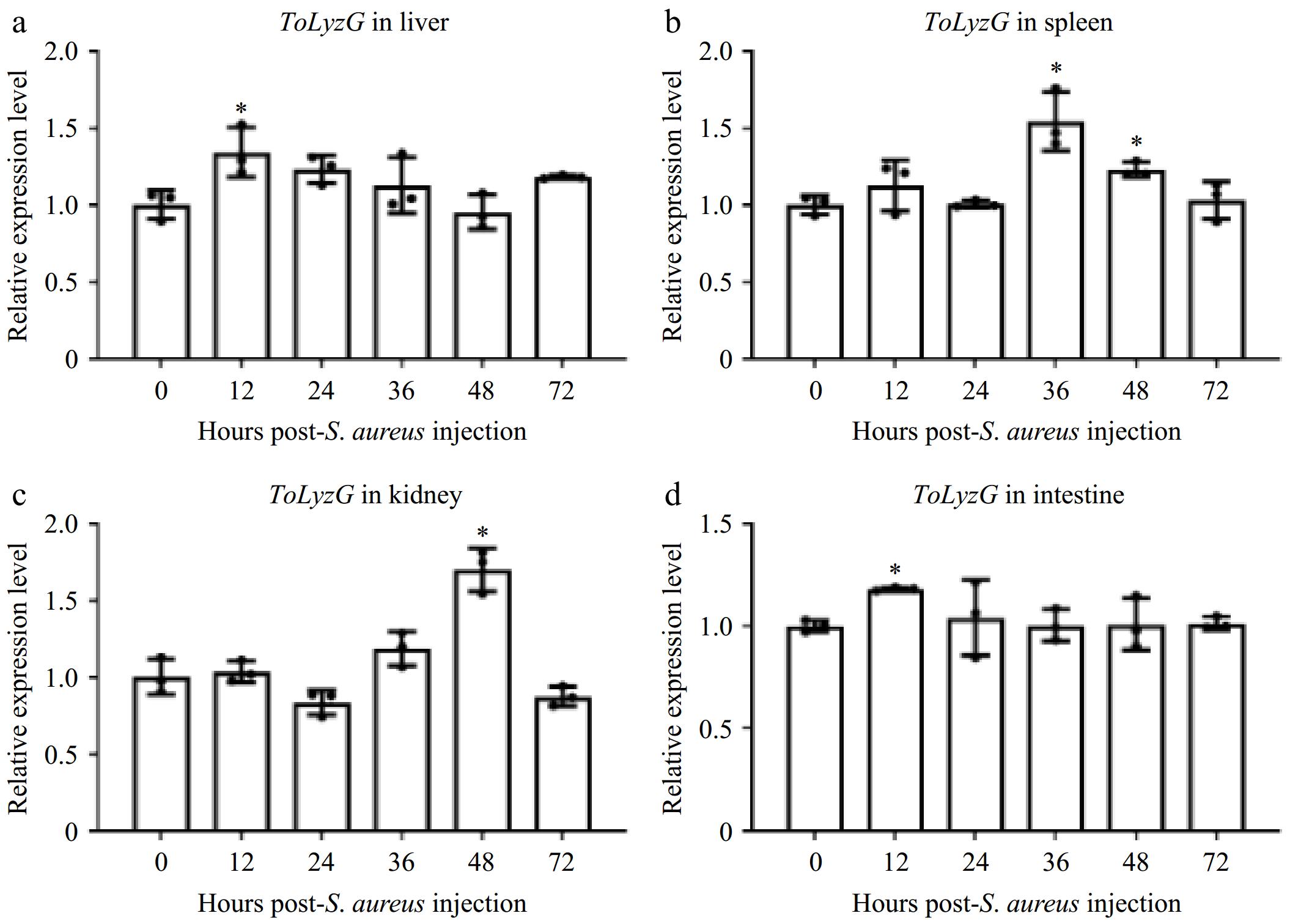
Figure 6.
ToLyzG mRNA expression after S. aureus challenge. Different time course of ToLyzG expression in the (a) liver, (b) spleen, (c) kidney, and (d) intestine at 0, 12, 24, 36, 48, and 72 h post-injection with S. aureus was analyzed by RT-qPCR. Data are represented as the mean ± SD (n = 3). Significant differences are indicated by asterisks, * p < 0.05.
Expression and purification of rToLyzG
-
The recombinant ToLyzG protein was successfully expressed in E. coli after being induced with IPTG. Upon analysis by SDS-PAGE, an expected protein band of about 41 kDa, corresponding to rToLyzG, was detected in the supernatant of the protein extract (Fig. 7a). The rToLyzG fusion protein was then purified using a Ni-NTA column, resulting in the isolation of a single protein band. The concentrations were measured as 614 μg/mL for the rToLyzG protein and 586 μg/mL for the rTRX protein, respectively.
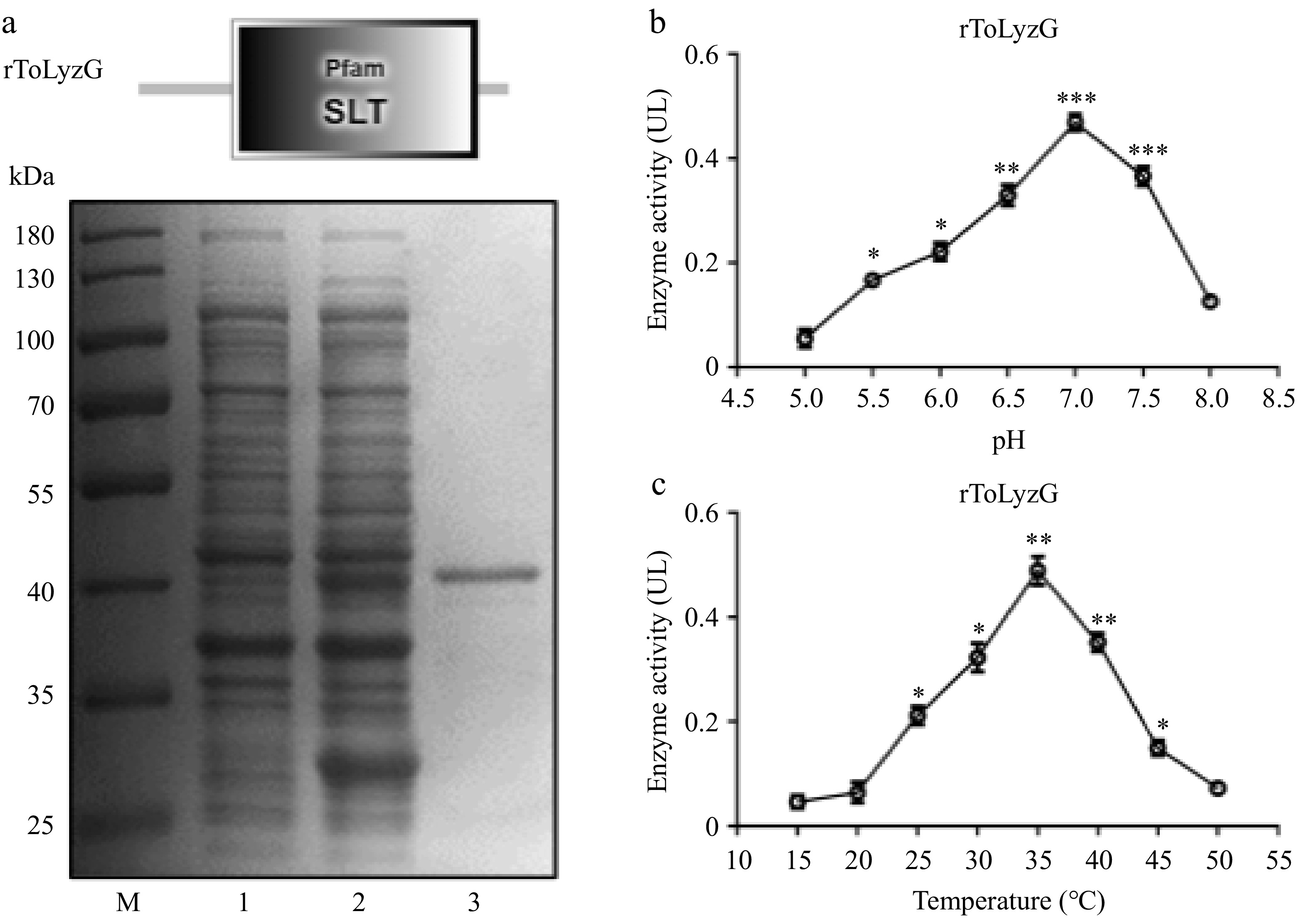
Figure 7.
Prokaryotic recombinant expression and purification of ToLyzG. (a) SDS-PAGE analysis of rToLyzG in E. coli BL21 (DE3). Lane M: protein molecular standard; lane 1: pET-32a-ToLyzG in BL21 (DE3), not induced with IPTG; lane 2: pET-32a-ToLyzG in BL21 (DE3), IPTG-induced for 5 h; lane 3: purified rToLyzG. Lytic activity of rToLyzG at various (b) pH and (c) temperatures measured by turbidimetric assay. Lytic activities are expressed as mean UL. Data are presented as the mean ± SD (n = 3). Significant differences are shown with asterisks, * p < 0.05, ** p < 0.01, *** p < 0.001.
Influence of pH and temperature on the lytic activity of rToLyzG
-
The peak activity of lysozymes from each fish species varies at different temperatures and pH values. Therefore, the lytic capacity of rToLyzG was assessed through a turbidimetric test, utilizing M. lysodeikticus as the test substrate. The results indicated an optimal pH range of 5.5 to 8.0, with a peak performance at pH 7.0 (UL = 0.47, Fig. 7b). Furthermore, rToLyzG exhibited robust lytic activity between 25 °C and 45 °C, achieving its maximum activity at 35 °C (UL = 0.49, Fig. 7c).
Lytic activity of rToLyzG against bacteria
-
The lytic activity of rToLyzG against six species of bacteria was determined at optimum pH and temperature (Table 2). rToLyzG protein exhibited stronger bacteriolytic capacity against Gram-negative bacteria V. harveyi (UL = 0.865), V. parahaemolyticus (UL = 0.785), followed by A. hydrophila (UL = 0. 658). The bacteriolytic capacity was relatively low against Gram-positive bacteria S. aureus (UL = 0. 516), B. subtilis (UL = 0. 483), and B. thuringiensis (UL = 0. 466), respectively.
Table 2. Lytic activity of rToLyzG against bacteria.
Bacteria UL Vibrio harveyi 0.865 Vibrio parahaemolyticus 0.785 Aeromonas hydrophila 0.658 Staphylococcus aureus 0.516 Bacillus subtilis 0.483 Bacillus thuringiensis 0.466 Binding of rToLyzG to polysaccharide
-
The direct binding of rToLyzG to polysaccharides was determined by ELISA. Figure 8 illustrated that rToLyzG could bind to LPS and PGN with different binding affinities. The saturation of rToLyzG binding to polysaccharides was observed, with the binding profile aligning with a logarithmic pattern. Notably, LPS demonstrated a stronger binding affinity for rToLyzG compared to PGN.
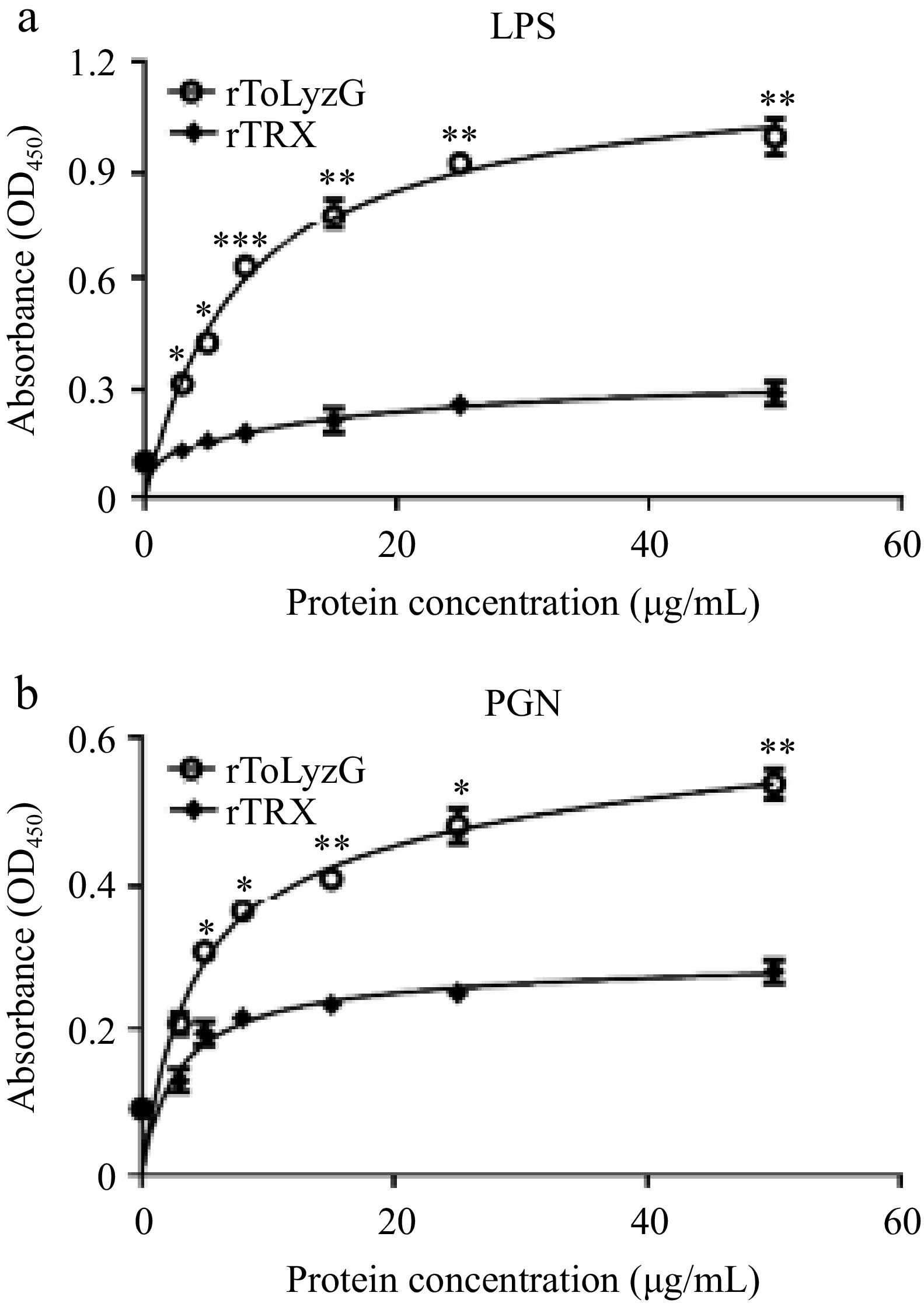
Figure 8.
Carbohydrate binding ability of rToLyzG. The binding of different concentrations of rToLyzG to (a) LPS and (b) PGN was assessed by ELISA, with rTRX serving as a control. Data are presented as the mean ± SD (n = 3). Significant differences are shown with asterisks, * p < 0.05, ** p < 0.01, *** p < 0.001.
Binding assay of rToLyzG to bacteria
-
The binding capacity of rToLyzG to bacteria was assessed through Western blot and ELISA analyses. Western blot results showed distinct reaction bands (about 41 kDa) with high specificity in the elution solution, indicating that rToLyzG could bind to V. harveyi, V. parahaemolyticus, A. hydrophila, S. aureus, B. subtilis, and B. thuringiensis (Fig. 9a). In ELISA, the P/N values of rToLyzG for six bacteria tested were all greater than 2.1 (Fig. 9b). Furthermore, compared to Gram-positive bacteria, rToLyzG exhibited a stronger binding affinity towards Gram-negative bacteria.
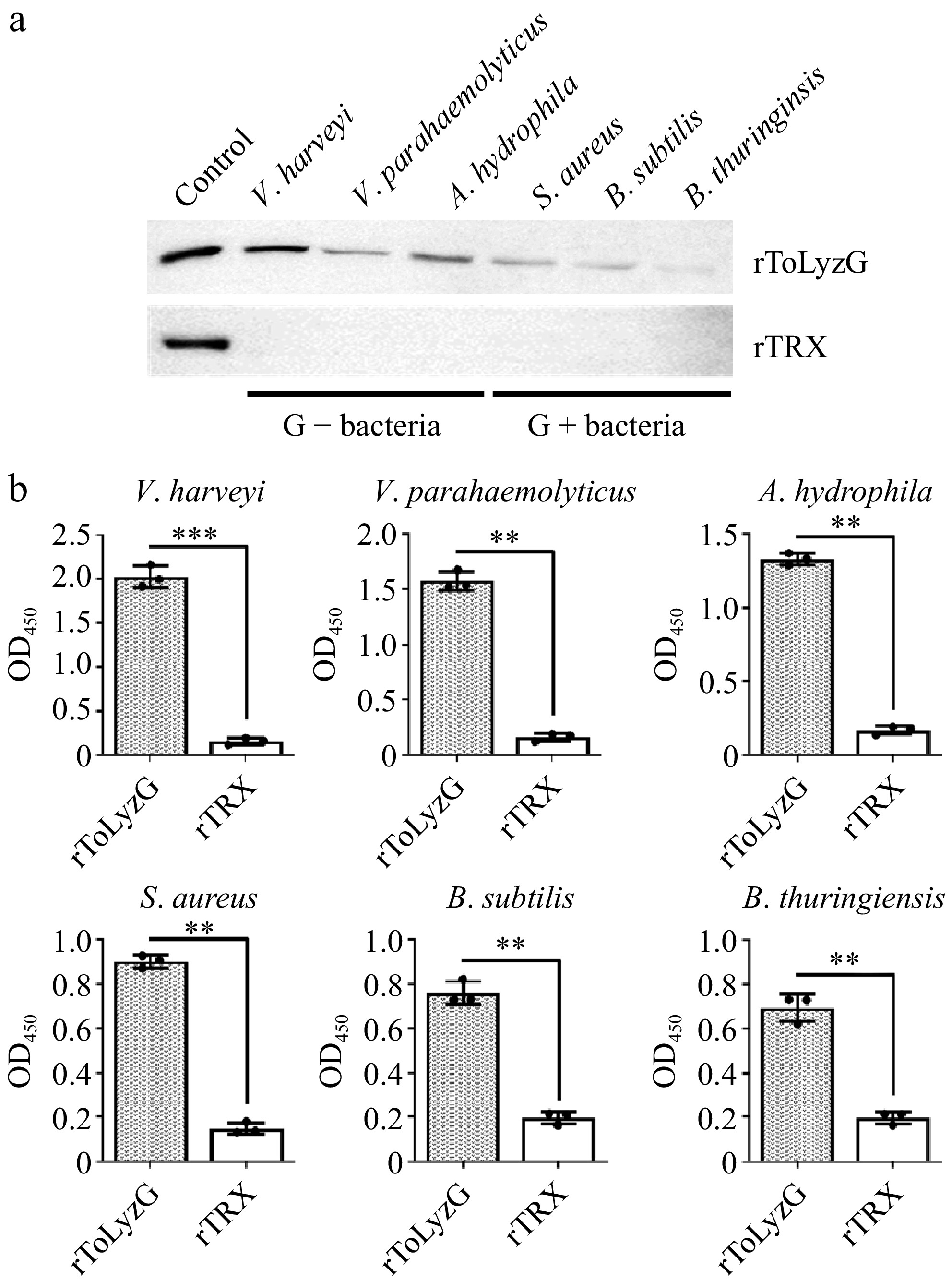
Figure 9.
Binding of rToLyzG to bacteria. The binding activity of rToLyzG to different bacteria, including V. harveyi, V. parahaemolyticus, A. hydrophila, S. aureus, B. subtilis, and B. thuringiensis was determined by (a) Western blot and (b) ELISA. rTRX was used as a negative control. Significant differences are represented with asterisks, ** p < 0.01, *** p < 0.001.
Antimicrobial activity of rToLyzG
-
The impact of the rToLyzG protein on the growth of the above bacteria was further studied. During this experiment, rToLyzG was incubated with bacteria samples, and the growth of bacteria was monitored at defined intervals. Compared with the rTRX protein treatment group, rToLyzG significantly suppressed the growth of V. harveyi (Fig. 10a), V. parahaemolyticus (Fig. 10b), and A. hydrophila (Fig. 10c). Conversely, the inhibitory effect of rToLyzG against S. aureus (Fig. 10d), B. subtilis (Fig. 10e), and B. thuringiensis (Fig. 10f) was relatively low.
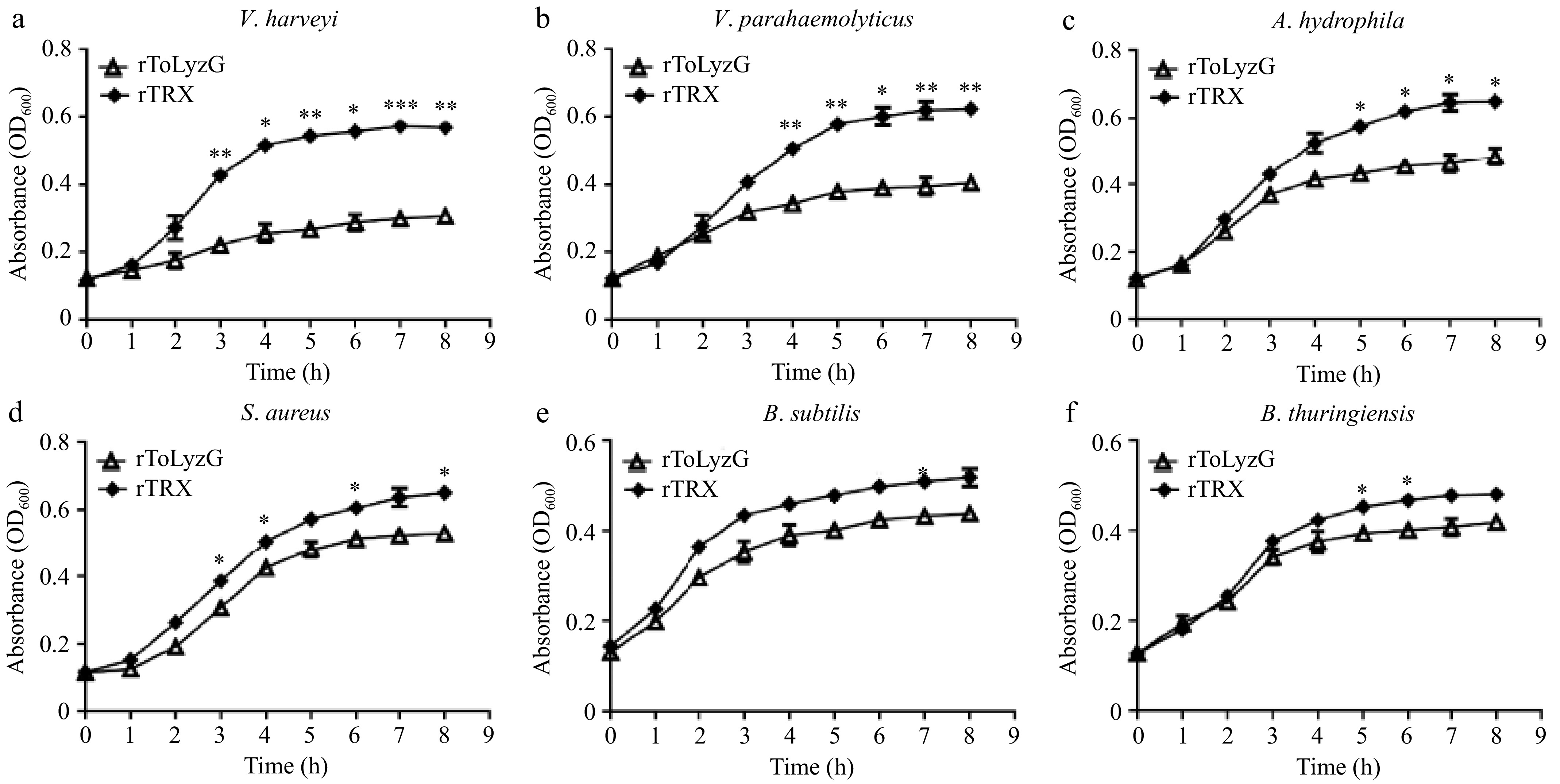
Figure 10.
Antimicrobial activity of rToLyzG against different microbes. (a) V. harveyi, (b) V. parahaemolyticus, (c) A. hydrophila, (d) S. aureus, (e) B. subtilis, or (f) B. thuringiensis was inoculated with rToLyzG protein. Bacterial growth was evaluated by measuring the absorbance at 600 nm every 1 h. rTRX was used as the control. Significant differences are represented with asterisks, * p < 0.05, ** p < 0.01, *** p < 0.001.
-
Lysozymes, owing to their lytic activity, are crucial immune molecules found across vertebrates and invertebrates. These enzymes exhibit varying amino acid sequences and enzymatic characteristics[28,29]. In various fish species, several g-type lysozymes have been previously cloned and are known for their conserved antimicrobial properties, whereas the structural and functional roles they play in pufferfish remain uncharacterized.
In the current research, a g-type lysozyme, named ToLyzG, was identified from T. obscurus, and its lytic characteristics further investigated. The cDNA sequence of ToLyzG encoded a polypeptide consisting of 190 amino acids, with an estimated Mw of 21 kDa. The amino acid number of ToLyzG was comparable to that reported in most fish species such as Channa striatus[30], Euryglossa orientalis[31], L. rohita[12], and T. fasciatus[14]. The majority of fish g-type lysozymes, including the ToLyzG studied here, lack a signal peptide at the N-terminus, indicating that these enzymes may not be secreted via cellular or tissue pathways. The absence of cysteine residues is a prevalent trait among many fish g-type lysozymes[32]. Notably, ToLyzG lacked the four highly conserved cysteine residues found in birds and mammals, aligning with the theory that ToLyzG may function as an intracellular protein. Sequence comparison of ToLyzG with lysozymes from diverse species unveiled the presence of the GLMQ motif, which is postulated to enhance binding affinity for NAG-NAM. Furthermore, the GLMQ motif has been implicated in enzymatic activities or the modulation of specific lytic transglycosylases[33]. Phylogenetic analysis demonstrated that ToLyzG was primarily clustered with other pufferfish g-type lysozymes and, to a lesser extent, with those from other teleost fish. Consistent with traditional taxonomic classifications, these species belong to the same family, and the clustering of their g-type lysozymes offer molecular support for the study of biological evolution.
Considerable investigative efforts have been devoted to exploring the tissue distribution of g-type lysozymes across diverse fish species, consistently demonstrating their widespread occurrence in all tissue categories. In S. maximus, a high-level expression of SmLysG was detected in the kidney, gills, brain, heart, and spleen[8]. Abundant expression of Ec-lysG was observed in the head kidney, stomach, gills, and intestine of Epinephelus coioides[17]. D. labrax L. g-type lysozyme was highest expressed in the gills and head kidney[10]. Lr-lysG was mainly expressed in the skin, anterior kidney, brain, liver, and spleen of L. rohita[12]. The current study found ToLyzG to be expressed in six tissues, particularly with heightened expression in the liver, kidney, intestine, spleen, and gills. Lysozyme, a well-documented antimicrobial protein is known for its broad-spectrum inducibility against various pathogens, particularly bacteria. However, following bacterial challenges, the expression patterns of g-type lysozymes in fish immune organs showed variability. For instance, C. idellus challenged with A. hydrophila exhibited upregulated g-type lysozyme in the liver, head kidney, spleen, and gills[7]. In S. maximus, Lyg2 expression was upregulated in mucosal tissues after challenge with V. anguillarum and S. iniae[34]. The expression of EuOr LysG was upregulated in the gills, kidney, liver, spleen, and muscle of E. orientalis injected with V. parahaemolyticus[31]. V. harveyi can cause luminescent vibriosis in aquatic animals, leading to high mortality rates and significant economic losses in aquaculture[35]. S. aureus can contaminate feed and water, posing a risk to the health of aquatic animals[36]. Infection of T. obscurus with V. harveyi and S. aureus resulted in increased levels of ToLyzG mRNA, hinting at a potential antimicrobial function of ToLyzG in the host's immune defenses. While most lysozymes are known to be effective against Gram-positive bacteria, with limited reports on their activity against Gram-negative bacteria[37], the present study observed a significantly greater upregulation of ToLyzG in response to Gram-negative bacteria.
Studies suggest that g-type lysozymes in fish exhibit varying pH and temperature dependence patterns. G-type lysozymes of P. olivaceus[5], Siniperca chuatsi[38], C. idellus[7], and E. orientalis[31] exhibited the highest activities at pH 6.0 to 7.0, whereas gLYS in G. morhua L. possessed peak activity at pH ≤ 4.8[6]. The optimal temperatures of g-type lysozymes in S. maximus[8], E. coioides[17], and seahorse Hippocampus abdominalis[39] were 20, 30, and 35 °C, respectively. In contrast, the g-type lysozyme of C. idellus showed maximum activity at 60 °C[7]. The present research revealed that the optimal conditions for rToLyzG activity were pH 7.0 and 35 °C. rToLyzG exhibited high activity within a broad pH range of 5.5 to 8.0 and a temperature range spanning from 25 to 45 °C. These findings are in accordance with the hypothesis that ToLyzG functions as a highly active enzyme under the physiological conditions typically encountered in its natural environment. Further testing of rToLyzG's lytic activity against a diverse array of bacteria revealed that, at optimal conditions of pH 7.0 and 35 °C, it exhibited potent lytic activity against Gram-negative bacteria V. harveyi, V. parahaemolyticus, and A. hydrophila. In contrast, its activity against Gram-positive bacteria was relatively lower. V. parahaemolyticus severely threatens fish health, causing diseases that can lead to significant mortality in aquaculture[40]. A. hydrophila is a pathogenic bacterium that can cause septicemia, furunculosis, and other diseases in fish, seriously affecting their health and survival rates[41]. It was also reported that rLr-lysG in L. rohitahad had lytic activity against A. hydrophila and Edwardsiella tarda[12]. G-type lysozymes of E. coioides and S. chuatsi possessed lytic activities against V. alginolyticus and A. hydrophila[38,42]. In E. orientalis, rEuOr LysG showed strong lytic activities against bacteria Bacillus cereus and A. hydrophila[31].
A fundamental role of lysozyme is to function as an antimicrobial agent, which often involves direct interaction with bacteria. The present study demonstrated that rToLyzG could bind directly to LPS and PGN, the primary constituents of the cell walls in Gram-negative and Gram-positive bacteria, with a notably higher binding affinity towards LPS than PGN. rToLyzG could also bind to V. harveyi, V. parahaemolyticus, A. hydrophila, S. aureus, B. subtilis, and B. thuringiensis, with a stronger binding ability to Gram-negative bacteria. The binding captivity of rToLyzG to bacteria helps to prevent these pathogens from invading cells, instead keeping them confined to the extracellular surface or matrix. To investigate the possible antibacterial captivity of rToLyzG, growth inhibition experiments were performed at pH 7.0. The findings indicated that rToLyzG exhibited a stronger inhibitory effect against Gram-negative bacteria. V. harveyi exhibited a more pronounced growth reduction in response to rToLyzG than V. parahaemolyticus and A. hydrophila, hinting at potential differences in cell wall structures that make V. harveyi more susceptible to rToLyzG. Contrary to our results, rSmLysC showed stronger growth inhibition against M. luteus, S. aureus, and S. iniae, but had weaker effects against E. tarda, Pseudomonas fluorescens, and V. anguillarum[43]. These results further suggest that ToLyzG plays a significant but distinct role in the defense against Gram-negative and Gram-positive bacterial infections in T. obscurus.
-
In the current study, a g-type lysozyme of T. obscurus (ToLyzG) was first identified. ToLyzG was expressed in six tissues of T. obscurus. ToLyzG expression was upregulated after V. harveyi challenge in the liver, spleen, kidney, and intestine. The recombinant ToLyzG had lytic activity against various bacteria. Furthermore, rToLyzG could not only bind to LPS, PGN, and six bacteria but also inhibit the growth of these bacteria. Notably, rToLyzG exhibited stronger inhibitory effects against Gram-negative bacteria compared to its effects on Gram-positive bacteria. The combined results suggest a crucial role of ToLyzG in the immunity of T. obscurus against bacterial challenge.
The current study was supported by the Open project of Key Laboratory of Marine Biogenetic Resources, Third Institute of Oceanography, Ministry of Natural Resources (HY202402), the Guangdong Provincial Key Laboratory of Applied Marine Biology (LAMB20221006), the Project for Seed Industry Vitalization of Jiangsu Province (JBGS[2021]133), and the Research Funds of the Jiangsu Marine Bioresources Innovation Center (2022YHTZZZ04).
-
The sampling protocol was approved by the Ethics Committee of Experimental Animals at Hohai University (identification number: hhuhy-23-012, approval date: 2023/5/25) and adhered to the Animal Care Guidelines issued by the Ministry of Science and Technology (China). No wild animals were used in this study.
-
The authors confirm contribution to the paper as follows: methodology: Huang Y, Shen R, Cui LF, Shi Y, Song XR; software: Huang Y, Shen R, Cui LF, Shi Y; supervision, writing - original draft: Huang Y; funding acquisition: Huang Y, Shi Y, Zhao Z; writing - review & editing: Huang Y, Zhao Z; data curation: Shen R; formal analysis, validation: Shen R, Cui LF, Song XR; resources, conceptualization: Zhao Z. All authors reviewed the results and approved the final version of the manuscript.
-
Data will be made available upon reasonable request to the corresponding author.
-
No potential conflicts of interest were disclosed. The manuscript is approved by all authors for publication. Zhe Zhao is the Editorial Board member of Animal Advances who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Huang Y, Shen R, Cui LF, Shi Y, Song XR, et al. 2025. Involvement of a goose-type lysozyme from Takifugu obscurus in immune response against Gram-negative bacteria. Animal Advances 2: e001 doi: 10.48130/animadv-0024-0009 

Involvement of a goose-type lysozyme from Takifugu obscurus in immune response against Gram-negative bacteria
- Received: 24 October 2024
- Revised: 24 November 2024
- Accepted: 12 December 2024
- Published online: 21 January 2025
Abstract: Lysozyme plays a crucial role in the immune response to bacterial infections. In this research, a goose-type lysozyme (designed as ToLyzG) was identified from Takifugu obscurus. ToLyzG cDNA encoded a 190-amino-acid protein. The phylogenetic tree showed that ToLyzG was primarily grouped with five g-type lysozymes from other pufferfish. ToLyzG was expressed in all examined tissues of untreated T. obscurus. The expression of ToLyzG in the liver, spleen, kidney, and intestine could be induced after Vibrio harveyi infection. To understand its biological activity, the coding region of ToLyzG was expressed and purified. The optimal conditions for recombinant ToLyzG were pH 7.0 and 35 °C. rToLyzG had lytic activity against a variety of bacteria with the highest activity against V. harveyi, Vibrio parahaemolyticus, and Aeromonas hydrophila. rToLyzG protein could not only bind to lipopolysaccharide, peptidoglycan, and six species of bacteria, but also inhibit the growth of these bacteria. Remarkably, rToLyzG demonstrated more potent suppressive activity against Gram-negative bacteria than Gram-positive bacteria. The present research enhances the comprehension of the biochemical features of g-type lysozyme from T. obscurus, potentially shedding light on its significant function in the host's defense strategy against bacterial pathogens.
-
Key words:
- G-type lysozyme /
- Expression analysis /
- Lytic activity /
- Innate immunity /
- Takifugu obscurus