









-
Grafting, an agricultural practice with a 4,000-year history in China, involves attaching a branch or bud (scion) of one plant to the rootstock of another plant[1]. This horticultural strategy is widely applied to horticultural crops such as vegetables[2], and fruit trees[3]. In fruit tree cultivation, dwarfing interstocks represent a significant technological approach. By selecting or cultivating interstocks with dwarfing growth characteristics, tree height and canopy size can be effectively controlled, enhancing fruit yield and quality[4]. The rootstock and scion interact, with the rootstock enhancing the growth and yield of the scion, improving the plant's response to biotic or abiotic stresses[5], increasing the content of functional components such as sugars, vitamins, and polyphenols[6], and also influencing the expression of the scion's transcriptome and miRNAs[7].
Apple is one of the most important temperate fruit trees in China and globally[8], and the apple industry holds a significant position in China's agricultural sector[9]. In recent years, the application of dwarfing interstocks in fruit tree cultivation has become increasingly widespread, particularly in apple cultivation, where they have emerged as one of the primary means to achieve dwarfing and dense planting[10]. In apple cultivation, grafting is a key agricultural technique for achieving dwarfing and dense planting, which can promote early flowering, increase yield, and improve fruit quality. Dwarfing and dense planting involve two types of rootstocks: dwarfing self-rooted rootstocks and dwarfing interstocks[11]. Due to the poor cold tolerance of dwarfing self-rooted rootstocks, which makes them unsuitable for apple production in cold northern regions, dwarfing interstocks are commonly used in apple production areas such as Xinjiang, Liaoning, and Shandong[12].
Total phenols, a class of compounds with diverse biological activities, are widely present in fruits and play crucial roles in determining fruit flavor, color, and health benefits[13]. In apple fruits, total phenols and their components, such as chlorogenic acid, catechin, and epicatechin, not only participate in the fruit's antioxidant and defense mechanisms but are also closely related to flavor, browning, and health-promoting properties[14,15]. Understanding the effects of different dwarfing interstocks on total phenolic content and polyphenol composition in apple fruits is crucial for optimizing orchard management practices and enhancing fruit quality. Nevertheless, the mechanisms by which dwarfing interstocks influence fruit quality, particularly their impact on phenolic content and composition, remain poorly understood, highlighting the need for further investigation.
Despite the extensive research on the effects of dwarfing interstocks on overall tree performance and fruit yield, studies specifically focusing on their impact on the polyphenol content and composition of apple cores remain scarce[16]. This is particularly noteworthy given that apple cores, often discarded as waste in processing and consumption, represent a significant portion of the fruit and may harbor valuable bioactive compounds[17]. The polyphenol-rich core could potentially be harnessed for various applications, including the development of functional foods, natural preservatives, or even pharmaceutical products[18]. Fruit cores, often viewed as waste in fruit processing, have been discovered to harbor polyphenol levels that exceed those in the peel but are lower than those in the pulp. The polyphenols found in fruit cores exhibit diverse biological activities, rendering them valuable for use in food, pharmaceuticals, and cosmetics. Historically, fruit cores were discarded, resulting in considerable wastage of these beneficial compounds. Identifying and quantifying the specific polyphenols in fruit cores is crucial for understanding their composition and potential health benefits. Grafting onto dwarfing interstocks can elevate the content of polyphenol components in the fruit core. This study investigates how different dwarfing interstocks influence the polyphenol profile of 'Huahong' apple cores, aiming to uncover novel insights for the sustainable and valuable utilization of this often-overlooked fruit component. By understanding the ways in which dwarfing interstocks affect polyphenol content and composition in apple cores, this research seeks to inspire innovative approaches to apple breeding and cultivation, enhancing the health-promoting properties of the entire fruit. Additionally, the findings may support the development of advanced processing techniques specifically designed to extract and utilize polyphenols from apple cores, thereby minimizing waste and increasing the overall value of apple production.
'Huahong' apple, as an important apple variety, is influenced by various factors, including rootstock selection and grafting methods, in terms of fruit quality[19]. This study investigates the mechanisms through which dwarfing interstocks influence apple fruit quality by examining their effects on the total phenolic content and polyphenol composition in the core of 'Huahong' apples. Using Malus as the base rootstock, six interstocks ('CG24', 'MD001', 'SH3', 'SH38', 'Mac9', and 'CX5') were utilized to graft 'Huahong' apples for experimental analysis. Polyphenol components and their concentrations were determined using ultra-high-performance liquid chromatography coupled with mass spectrometry (UHPLC-MS), facilitating an evaluation of the impact of different dwarfing interstocks on total phenolics and polyphenol composition in apple fruits. The core plays a pivotal role in fruit development and post-harvest processing. Investigating the dynamic changes in polyphenol composition within the core not only enhances our understanding of the biochemical mechanisms governing fruit quality attributes but also establishes a scientific framework for refining apple processing methodologies and facilitating the development of value-added products. These advancements drive both technological innovation and industrial growth within the field. Furthermore, the findings offer a robust scientific basis for the strategic selection and implementation of dwarfing interstocks to improve apple quality and production efficiency, while simultaneously providing theoretical foundations for optimizing orchard management practices and enhancing overall fruit quality parameters.
-
Grafting experiments were conducted at the apple dwarfing interstock demonstration garden of Hanjiagou Fruit Farm in Huludao City, Liaoning Province, China between August 2010 and August 2011. Subsequent laboratory analyses were performed at the Research Institute of Pomology, Chinese Academy of Agricultural Sciences (CAAS), in Xingcheng, Liaoning Province, China, from June 2023 to November 2023.
Experimental materials and instrumentation
Experimental materials
-
The experimental materials were sourced from the National Pear and Apple Germplasm Repository in Xingcheng, Liaoning Province (China). Six dwarfing interstocks (Table 1) were utilized in this study. In August 2010, these interstocks were grafted onto Malus rootstock using the bud grafting technique. Subsequently, in August 2011, the apple cultivar 'Huahong' was grafted onto the six dwarfing interstocks following the same method. For the control group, 'Huahong' was grafted directly onto Malus rootstock. All plants were maintained under standard field management practices.
Table 1. Dwarfing interstocks and breeding units.
No. Dwarfing interstocks Breeding units 1 'CG24' Cornell University and New York State Agricultural Experiment Station, Geneva 2 'SH38' Pomology Institute, Shanxi Academy of Agricultural Sciences 3 'SH3' Pomology Institute, Shanxi Academy of Agricultural Sciences 4 'MD001' Mudanjiang Branch of Heilongjiang Academy of Agricultural Sciences 5 'Mac9' Michigan State University (USA) 6 'CX5' Research Institute of Pomology, Chinese Academy of Agricultural Sciences 7 None, rootstock
was Malus (CK)/ During the fruit growth and development period in 2023, samples were collected every 20 d from June 9th to October 8th, totaling seven samplings. Both the different dwarfing interstock combinations and the control group of 'Huahong' apples were managed with the same bagging practice. For each treatment and the control, three trees with consistent growth were selected, and 10 fruits with uniform growth and development were collected from the outer and middle parts of the trees. After harvest, the fruit cores were extracted following the method described by Li et al.[20], 5 g of the apple core was taken and homogenized. The homogenate was then rinsed into a 100 mL volumetric flask with 80 mL of distilled water. The flask was then placed in a water bath at 100 °C and kept warm for 30 min. The flask was then removed, allowed to cool, and then the volume made up to the mark. The solution was then filtered and the filtrate retained for subsequent analysis.
Experimental apparatus
-
The following instruments were used in the experiments: a UV-Vis spectrophotometer (TU-1800SPC, Beijing Purkinje General Instrument Co., Ltd., Beijing, China); an ultra-performance liquid chromatography system coupled with a quadrupole mass spectrometer (UPLC Xevo/TQ) (Waters, USA), equipped with a photodiode array detector (Acquity PDAeλ); a Waters ACQUITY UPLC® HSS T3 column (2.1 mm × 150 mm, 1.8 μm); a frozen grinder (6870, SPEX, USA); a high-speed refrigerated centrifuge (CF16RXII, Hitachi, Japan); a solid-phase extraction (SPE) system (Preplinc platform, J2 Scientific, USA); SPE cartridges (Waters Oasis® HLB, 200 mg/6CC); a rotary evaporator (R-210, BUCHI, Switzerland); syringe filters (0.22 μm); an ultra-low temperature freezer (DW-HL388, Zhongke Meiling); and an ultrapure water system (Milli-Q Direct 8, MilliPore, USA).
Determination of total phenolic content in apples
-
The total phenolic content was determined using the Folin-Ciocalteu method[21], with gallic acid serving as the standard for the calibration curve. The fruit cores from seven combinations and seven different stages were chopped and homogenized using a tissue homogenizer. A 5 g sample of the homogenate was washed with 80 mL of distilled water into a 100 mL volumetric flask. The mixture was then subjected to a boiling water bath at 100 °C for 30 min, cooled, and diluted to 100 mL with distilled water. After filtration, 1.0 mL of the filtrate was taken, and 1.0 mL of Folin-Ciocalteu reagent solution was added. The mixture was diluted to 10 mL with distilled water, mixed well, and allowed to stand at room temperature for color development. The absorbance was measured at 765 nm. By referencing the standard curve and considering the dilution factor, the total phenolic content of the samples was calculated.
Extraction, purification, and determination of phenolic compounds in apples
-
The method for determining phenolic compounds in 'Huahong' apples was based on the approach outlined by Li et al., with minor modifications[20].
Extraction of phenolics
-
Ten grams of frozen and ground fruit core samples were taken, and phenolic compounds were extracted twice using 20 mL of 80% methanol each time with ultrasonic assistance. The combined extracts were diluted to 50 mL. Then, 10 mL of the extract was evaporated under reduced pressure at 40 °C using a rotary evaporator to remove methanol. The solid-phase extraction (SPE) column was activated with 5 mL of methanol, and the sample solution was loaded onto the column. The column was then rinsed twice with 5 mL of distilled water, which was discarded. Subsequently, the column was rinsed twice with 4 mL of methanol to collect the elution, which was then diluted to 5 mL with methanol. One milliliter of this solution was evaporated under reduced pressure at 40 °C to remove methanol, and the residue was redissolved in 5 mL of methanol. The solution was filtered through a 0.22 μm organic phase filter before analysis.
Mass spectrometry conditions
-
The ion source temperature was set at 150 °C, with a desolvation temperature of 400 °C. The desolvation gas flow rate was 800 L/h, and the cone gas flow rate was 50 L/h. Ultra-performance liquid chromatography (UPLC) conditions were as follows: column temperature at 40 °C, flow rate of 0.3 mL/min, injection volume of 2.0 μL. The mobile phase consisted of acetonitrile (A) and 0.5% formic acid solution (B), with a gradient elution program: A = 0% (0 min) → 10% (1 min) → 25% (16 min) → 40% (18 min) → 100% (19 min), returning to the initial condition at 20 min and equilibrating for 5 min. The PDA scanning wavelength was set to 210−600 nm, with channels at 280, 320, and 360 nm.
Qualitative and quantitative analysis
-
The phenolic compounds were identified and confirmed based on their relative retention times from chromatographic separation, characteristic spectral data from PDA, and various mass spectrometry modes including parent ion scan, daughter ion scan, and neutral loss scan from the quadrupole mass spectrometer. Quantitative analysis was then performed for the identified compounds.
Statistical analysis
-
Data processing in this study was conducted using Microsoft Excel 2016 software. Statistical analyses were performed using SPSS 26. The significance of differences between treatments was compared using the t-test (p < 0.05). Graphs were generated using online platforms such as OmicShare (
www.omicshare.com ) and hiplot (https://hiplot.cn/ ). -
Figure 1a and Supplementary Table S1 illustrate the dynamic changes in total phenolic content in fruit cores under different interstocks across seven developmental stages. The results showed that the total phenolic content in the fruit cores decreased progressively throughout fruit development. There was no significant difference in the total phenol of each combination. Specifically, the total phenolic content declined significantly during the early stages, stabilized during the mid-to-late stages, and reached its lowest level at fruit maturity (October 8th). Importantly, no significant differences were observed in the influence of interstocks on polyphenol content across the developmental stages. On June 9th (Fig. 1b), the total phenol content of different dwarfing interstocks followed the order: CK > CG24 > Mac9 > SH38 > SH3 > MD001 > CX5. Among them, the CK combination had the highest total phenol content which was 4,196.04 mg/kg, while the CX5 combination had the lowest, at 3,518.79 mg/kg. On October 8th (Fig. 1c), the total phenol content of different dwarfing interstocks was in the order: SH38 > CG24 > CK > CX5 > MD001 > Mac9 > SH3. The SH3 combination had the lowest total phenol content, which was 434.45 mg/kg, whereas the SH38 combination had the highest, at 514.39 mg/kg.
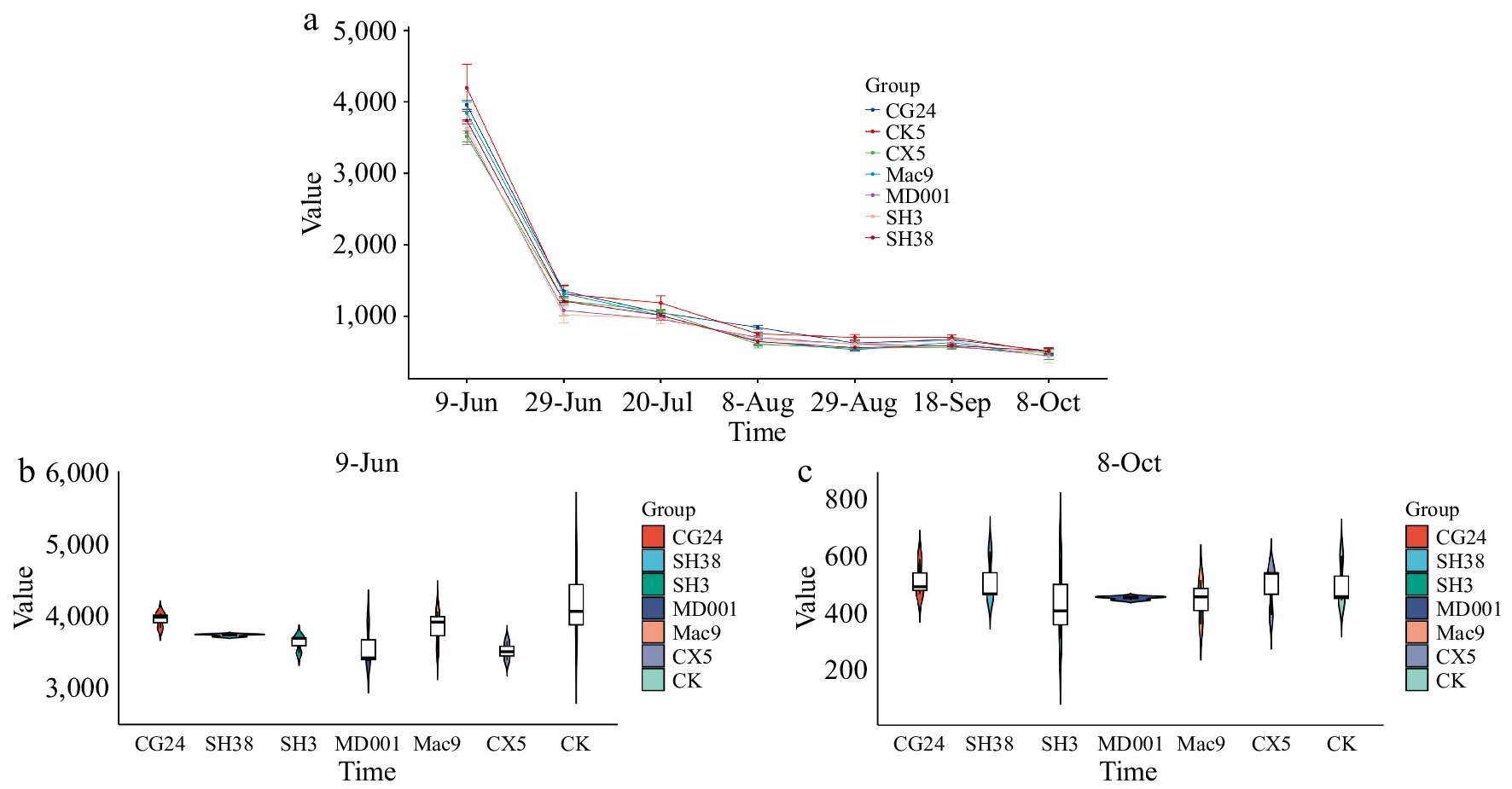
Figure 1.
Dynamic changes of total phenolics in the core of 'Huahong' Apple. (a) Dynamic changes of total phenols in Huahong apples with different dwarfing interstocks at seven stages. (b) Total phenol content in Huahong apples with different dwarfing interstocks on June 9th. (c) Total phenol content in Huahong apples with different dwarfing interstocks on October 8th.
Dynamic effects of different dwarfing interstocks on polyphenol component and contents in the cores of 'Huahong' apples
Changes in polyphenol component and contents in the core of 'Huahong' fruit at different stages
-
An analysis of the polyphenols in the core of 'Huahong' fruit was conducted. In this study, a total of 16 polyphenol components were detected, which were respectively Quercetin3-glucoside, Qercetin3-rhamnoside, Chlorogenic acid, Cryptochlorogenic acid, 5-O-p-coumaroyl quinic acid, 4-O-p-coumaroyl quinic acid, Procyanidin B2, Procyanidin C1, Epicatechin, Phloridzin-unknown 1, Phloridzin-unknown 2, Phloridzin-unknown 3, Phloridzin-unknown 4, Phloridzin-unknown 5, Phloridzin-unknown 6, and Phloridzin.
To investigate the impact of different dwarfing interstocks on the polyphenols in the fruit core, Fig. 2 presents river plots illustrating the content changes of various dwarfing interstocks at different time points. Figure 2a depicts the river plot of polyphenol content percentages, while Fig. 2b shows the river plot of polyphenol concentrations. Among the 16 compounds detected in this study, Chlorogenic acid, Epicatechin, Phloridzin, Procyanidin B1, and Procyanidin C1 were the major polyphenols in the fruit core. Except for a decrease in the proportion of Procyanidin B1 and Procyanidin C1 on June 29th, they remained relatively stable at other times.
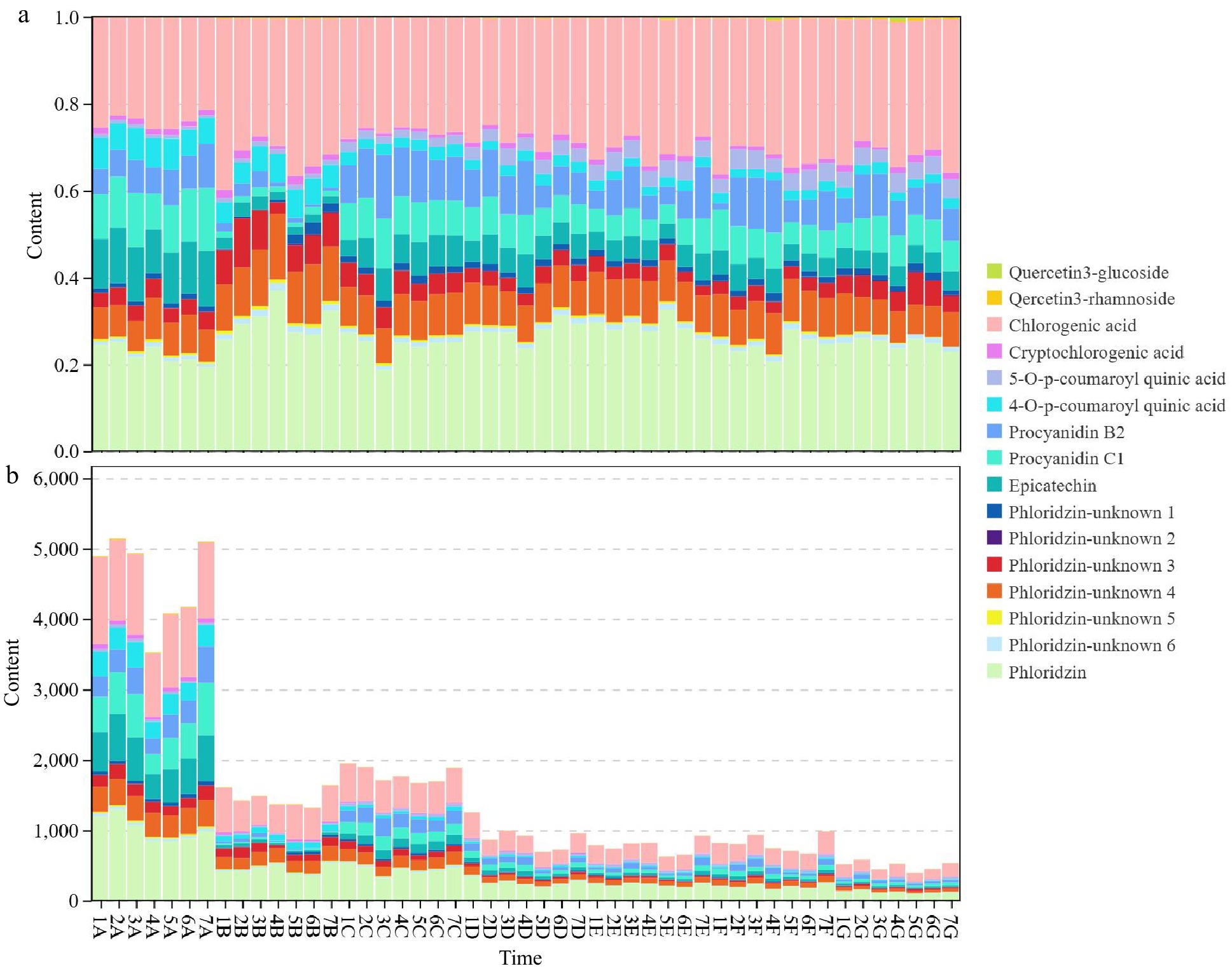
Figure 2.
Dynamic changes in polyphenol components of different dwarfing interstocks represented as a river plot. (a) River plot of percentage composition of polyphenols. (b) River plot of polyphenol component contents. The horizontal axis represents time, spanning seven-time points from 9-Jun to 8-Oct; the vertical axis represents compound concentrations, ranging from 0 to 6,000. 1 was 'CG24', 2 was 'SH38', 3 was 'SH3', 4 was 'MD001', 5 was 'Mac9', 6 was 'CX5', and 7 was CK; A was 9-Jun, B was 29-Jun, C was 20-Jul, D was 8-Aug, E was 29-Aug, F was 18-Sep, and G was 8-Oct, respectively.
An analysis of polyphenol components in different interstocks was conducted. Quercetin 3-glucoside reached its peak content during the early stages of fruit development and subsequently decreased gradually. Quercetin 3-rhamnoside also exhibited high levels in the early stages but showed an overall decreasing trend. Both Chlorogenic acid and Cryptochlorogenic acid had high initial contents, followed by a gradual decline. The contents of 5-O-p-coumaroyl quinic acid and 4-O-p-coumaroyl quinic acid remained relatively stable but showed an overall decreasing trend. Procyanidin B2 and Procyanidin C1 had high initial contents, which then gradually decreased. Epicatechin also showed high levels in the early stages and subsequently declined. The concentrations of Phloridzin-unknown 1−6 remained low and evenly distributed across all time points, with some polyphenols, such as Phloridzin-unknown 3 and Phloridzin-unknown 4, showing significant increases in content during the mid-stages of fruit development. Phloridzin gradually decreased and stabilized as the fruit developed.
In summary, Quercetin 3-glucoside, Quercetin 3-rhamnoside, Chlorogenic acid, Cryptochlorogenic acid, procyanidins, and Epicatechin exhibited high levels in the early stages of fruit development and then gradually decreased.
Correlation analysis of polyphenol components in the core of 'Huahong' fruit
-
Figure 3 presents a correlation heatmap of polyphenol components in the core of 'Huahong' fruit, intuitively reflecting the correlation coefficients between various components through a color gradient, and all the polyphenol data presented are the averages across seven periods. In the heatmap, multiple regions of highly correlated polyphenol components could be clearly observed. For instance, Quercetin-3-glucoside exhibited significant positive correlations with components such as 5-O-p-coumaroyl quinic acid and Chlorogenic acid, suggesting potential strong interactions or shared functional characteristics among them. Concurrently, some regions, such as those with Phloridzin-unknown 4 and Phloridzin-unknown 5, exhibited weak correlations, suggesting distinct chemical properties or biological functions.
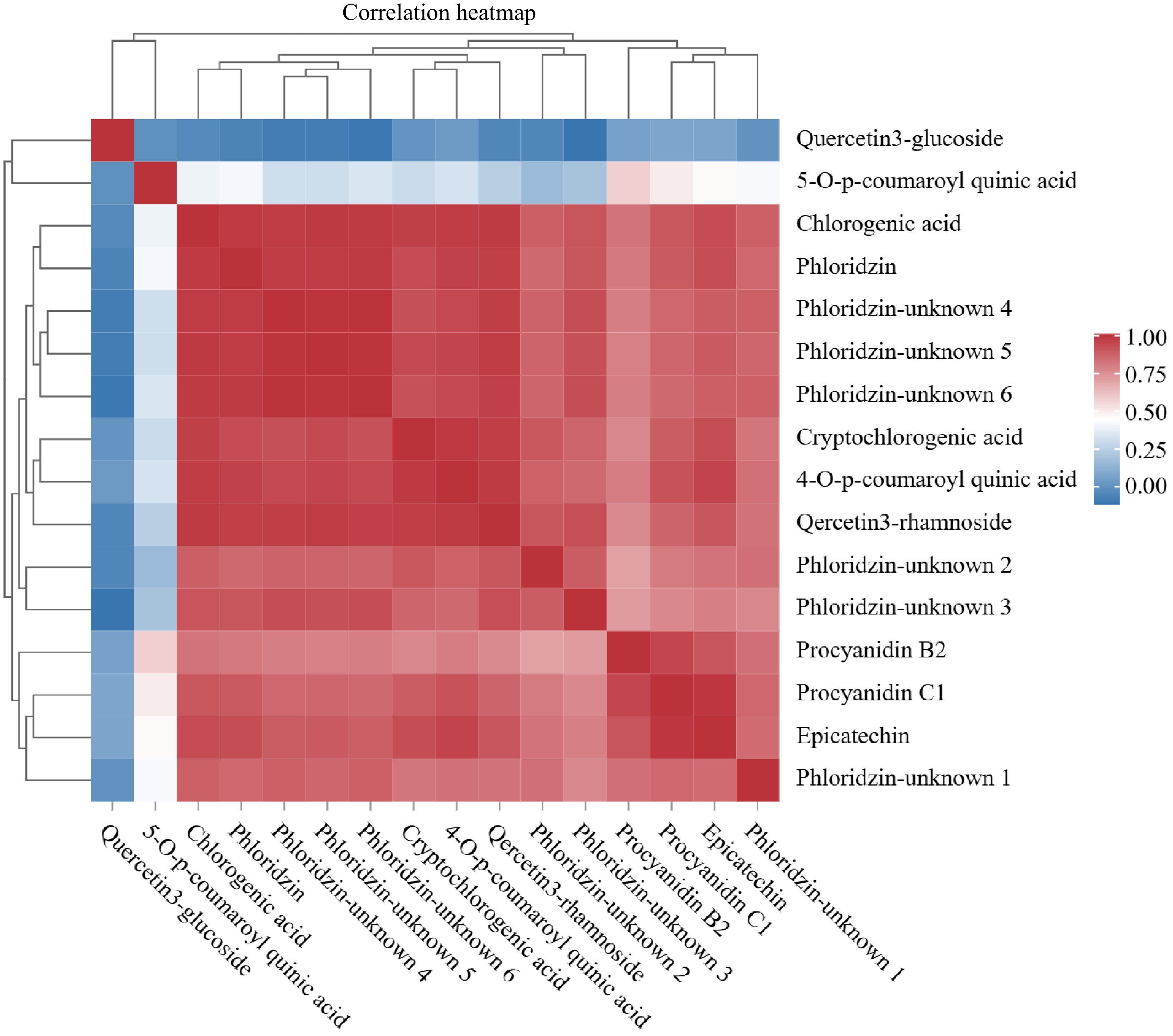
Figure 3.
Heat map of correlation of polyphenol components in cores of 'Huahong'.
The dendrogram on the left side of the heatmap grouped similar components together, revealing potential associations between them. For example, Phloridzin and its unknown components were tightly adjacent in the dendrogram, implying possible chemical or biological connections between them. Specific polyphenol components, like Quercetin-3-glucoside and 5-O-p-coumaroyl quinic acid, demonstrated extensive strong correlations in the heatmap, potentially indicating their significant roles in the 'Huahong' fruit core or their synergistic interactions with other components, jointly influencing the fruit's quality and biological activity.
Trend analysis of dynamic changes in polyphenol components of 'Huahong' cores from different dwarfing interstocks
-
To comprehensively analyze the dynamic changes in polyphenol components during fruit development, this study generated trend charts depicting core polyphenol variations at different developmental stages influenced by diverse dwarfing interstocks. Figure 4 presents the dynamic trends in polyphenol composition within the cores of 'Huahong' apples, categorized into four clusters (Cluster 1 to Cluster 4). Each subplot corresponded to a specific cluster, with the horizontal axis representing the sampling period (June to October) and the vertical axis showing the logarithmic values of polyphenol expression changes.
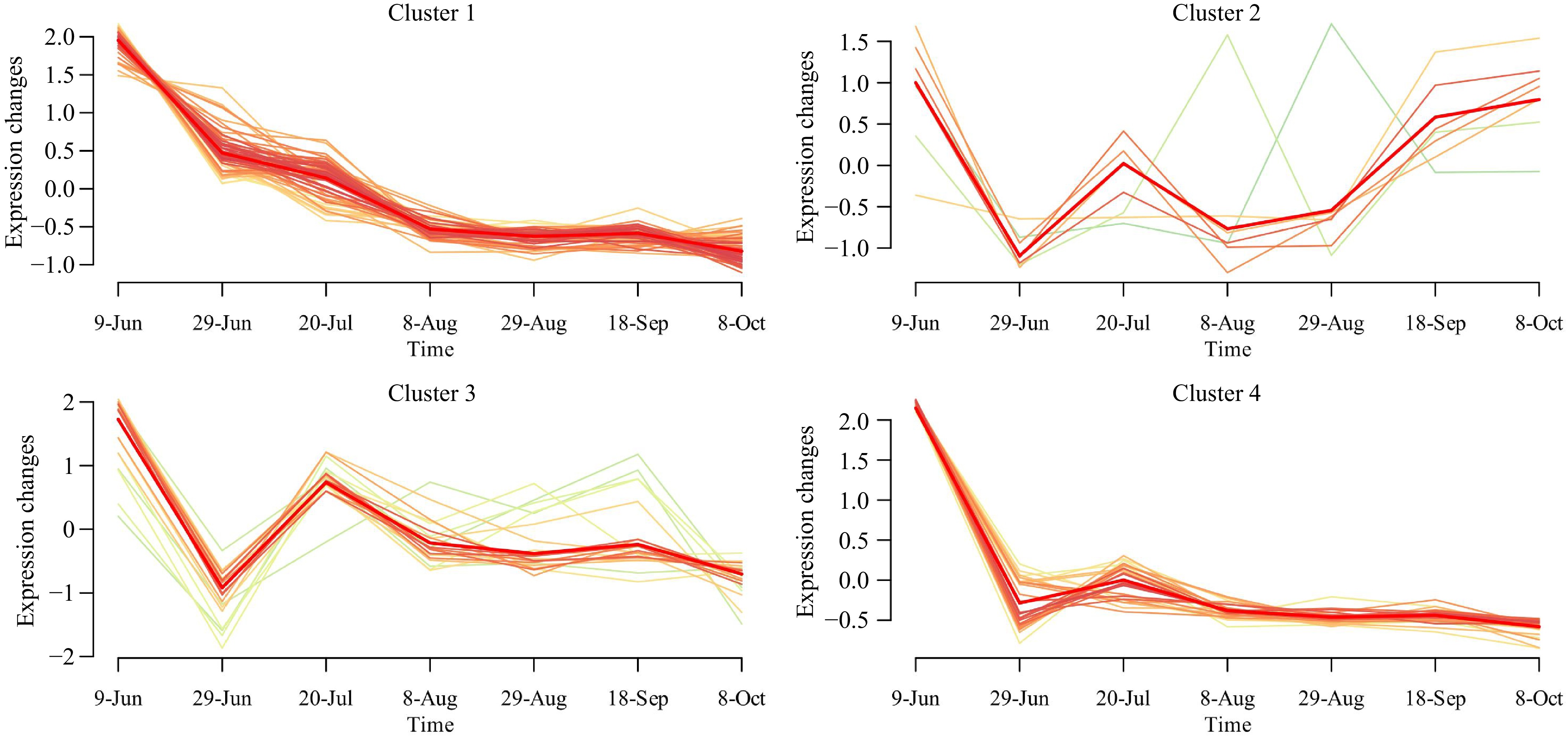
Figure 4.
Trend analysis of dynamic changes in polyphenol components of 'Huahong' cores. The x-axis represents time, and the y-axis represents changes in expression.
The analysis identified four distinct clusters of polyphenol expression patterns during fruit development. In Cluster 1, polyphenol expression gradually decreased from June to August and then stabilized. This trend suggested that polyphenol levels were high in the early stages of fruit development and gradually declined as the fruit matured. Cluster 2 demonstrated a more complex fluctuation in polyphenol expression. From June to July, expression levels significantly decreased, followed by fluctuations between mid-August and early September, before ultimately stabilizing. The polyphenol expression pattern in Cluster 3 demonstrated a marked decline between June and July, followed by a distinct peak from late July to early August, after which levels gradually stabilized. This peak may have represented a significant accumulation of polyphenols during the mid-stage of fruit development. In contrast, Cluster 4 exhibited a simpler pattern of polyphenol expression change. Starting in June, expression levels rapidly decreased and remained low throughout the observation period, with only a slight fluctuation occurring between late August and early September. This indicated that in this cluster, polyphenols rapidly declined in the early stages of fruit development and maintained a stable low level in the later stages.
In conjunction with Supplementary Table S2, the majority of polyphenols (such as Quercetin 3-rhamnoside, Chlorogenic acid, and Phloridzin) were classified into Cluster 1. The Quercetin 3-glucoside components from different dwarfing interstocks were consistently categorized into Cluster 2, 5-O-p-coumaroyl quinic acid into Cluster 3, and Epicatechin into Cluster 4. This indicated that dwarfing interstocks had a minimal impact on the dynamic changes of these components within the fruit core. Conversely, different dwarfing interstocks exerted a significant influence on the variations of Chlorogenic acid, Cryptochlorogenic acid, 4-O-p-coumaroyl quinic acid, Procyanidin B2, and Procyanidin C1. Procyanidin B2 was primarily classified into Cluster 3, whereas Procyanidin C1 and 4-O-p-coumaroyl quinic acid were predominantly found in Cluster 4. The contents of Procyanidin B2, Procyanidin C1, and Cryptochlorogenic acid in 'MD001', as well as the Cryptochlorogenic acid content in 'SH3', exhibited a gradual stabilization trend as the fruit developed, which differed from the CK.
The influence of different dwarfing interstocks on the trend of polyphenol composition in the core of 'Huahong' apples was significant, as evidenced by the following observations in the past: Most polyphenols, such as quercetin-3-rhamnoside, chlorogenic acid, and phloridzin, exhibited high expression levels in the early stages of fruit development, followed by a gradual decline and stabilization (Cluster 1). Quercetin-3-glucoside consistently showed an initial significant decrease, subsequent fluctuations, and eventual stabilization across different dwarfing interstocks (Cluster 2). 5-O-p-coumaroylquinic acid demonstrated a distinct peak during the mid-stage of development before stabilizing (Cluster 3), while epicatechin exhibited a rapid decline in the early stages and remained at low levels thereafter (Cluster 4). Furthermore, dwarfing interstocks had a pronounced effect on the dynamic changes of chlorogenic acid, cryptochlorogenic acid, 4-O-p-coumaroylquinic acid, procyanidin B2, and procyanidin C1. Notably, some polyphenol components in 'MD001' and 'SH3' showed a tendency towards stabilization in the later stages of fruit development, differing from the control (CK).
Principal Component Analysis (PCA) in polyphenol components of 'Huahong' cores
PCA analysis of polyphenol components in 'Huahong' rock core with different dwarfing interstocks
-
A PCA analysis was conducted on 16 polyphenol components in the fruit cores of different dwarfing interstocks. The horizontal axis represented PC1, which accounted for 95.4% of the total variation and was the primary source of data variability. The vertical axis represented PC2, accounting for 2.4% of the total variation and serving as a secondary source of variability. On the PC1 axis, the distribution of cryptochlorogenic acid, 5-O-p-coumaroylquinic acid, and 4-O-p-coumaroylquinic acid was particularly prominent. On the PC2 axis, certain compounds such as quercetin 3-glucoside, quercetin 3-rutinoside, and phloridzin also exhibited some distribution. Additionally, it could be observed from Fig. 5 that the compounds could be roughly divided into two groups. One group was located near the origin, including quercetin 3-glucoside, quercetin 3-rutinoside, chlorogenic acid, etc. The other group was distributed on the positive half of the PC1 axis, primarily consisting of cryptochlorogenic acid, 5-O-p-coumaroylquinic acid, 4-O-p-coumaroylquinic acid, and other compounds.
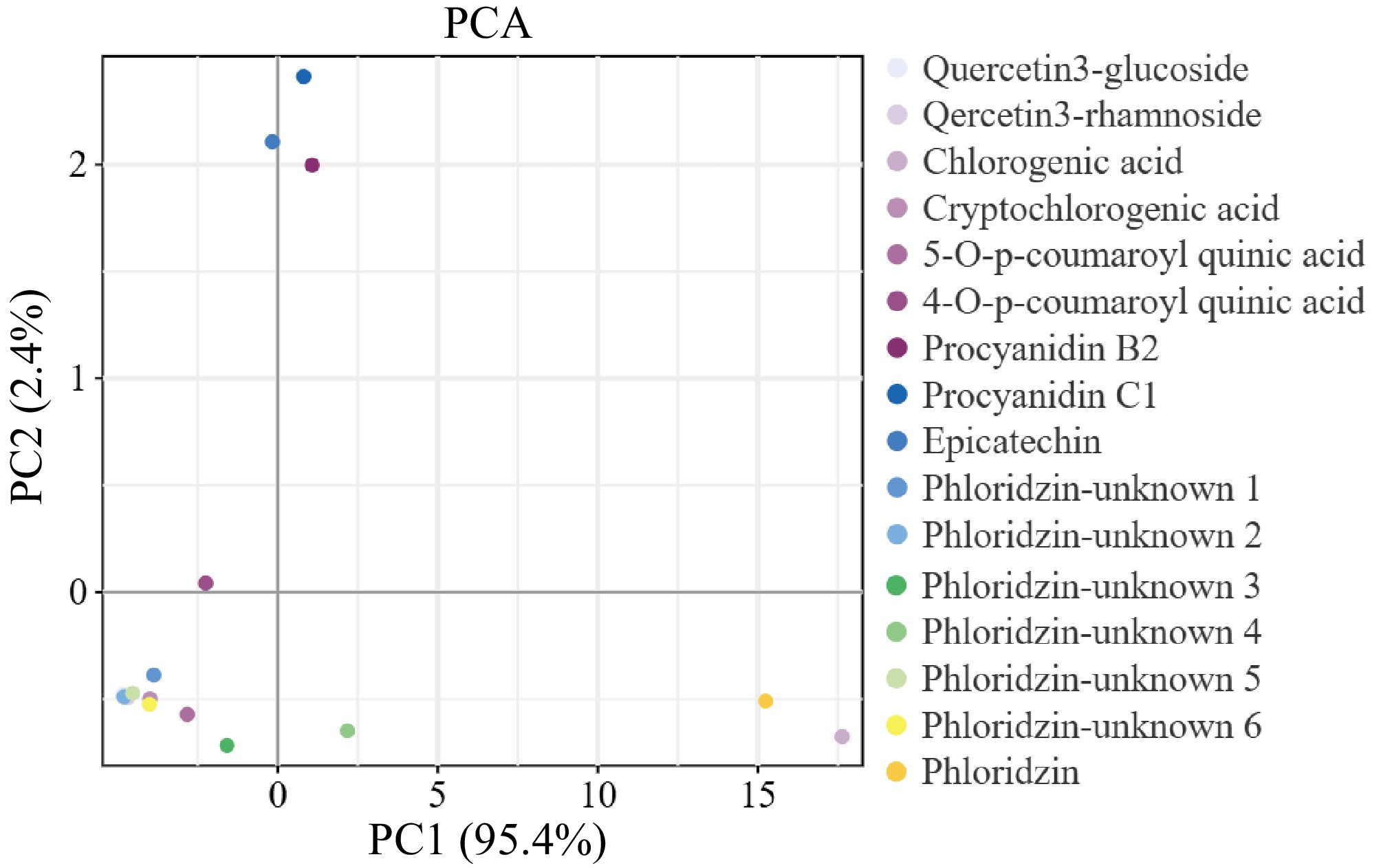
Figure 5.
PCA of polyphenol components in 'Huahong' core with different dwarfing interstocks. This PCA plot illustrates the distribution of various polyphenol compounds in the core of 'Huahong' fruit, influenced by different dwarfing interstocks. The first principal component (PC1) accounts for 95.4% of the variance, while the second (PC2) accounts for 2.4%, indicating that most of the variability in polyphenol composition can be explained by PC1. Different compounds are represented by distinct colors and shapes, showing how they cluster based on their PCA scores.
PCA of polyphenols in 'Huahong' core at different stages with various dwarfing interstocks
-
As shown in Fig. 6, the horizontal axis represented the first principal component (PC1), which accounted for 79.8% of the data variation, while the vertical axis represented the second principal component (PC2), explaining 7.7% of the data variation. The polyphenol components of different dwarfing interstocks were relatively distinct during the early stage (from June 9th to 29th) and mid-stage (July 20th) of fruit development, but became more concentrated during the mid-to-late stages. There were overlapping areas in the two-dimensional space, indicating a certain degree of similarity in polyphenol components among different dwarfing interstocks during the mid-to-late stages of fruit development. In contrast, the samples collected on June 9th and October 8th were more dispersed, suggesting significant differences in the principal components among different dwarfing interstocks.
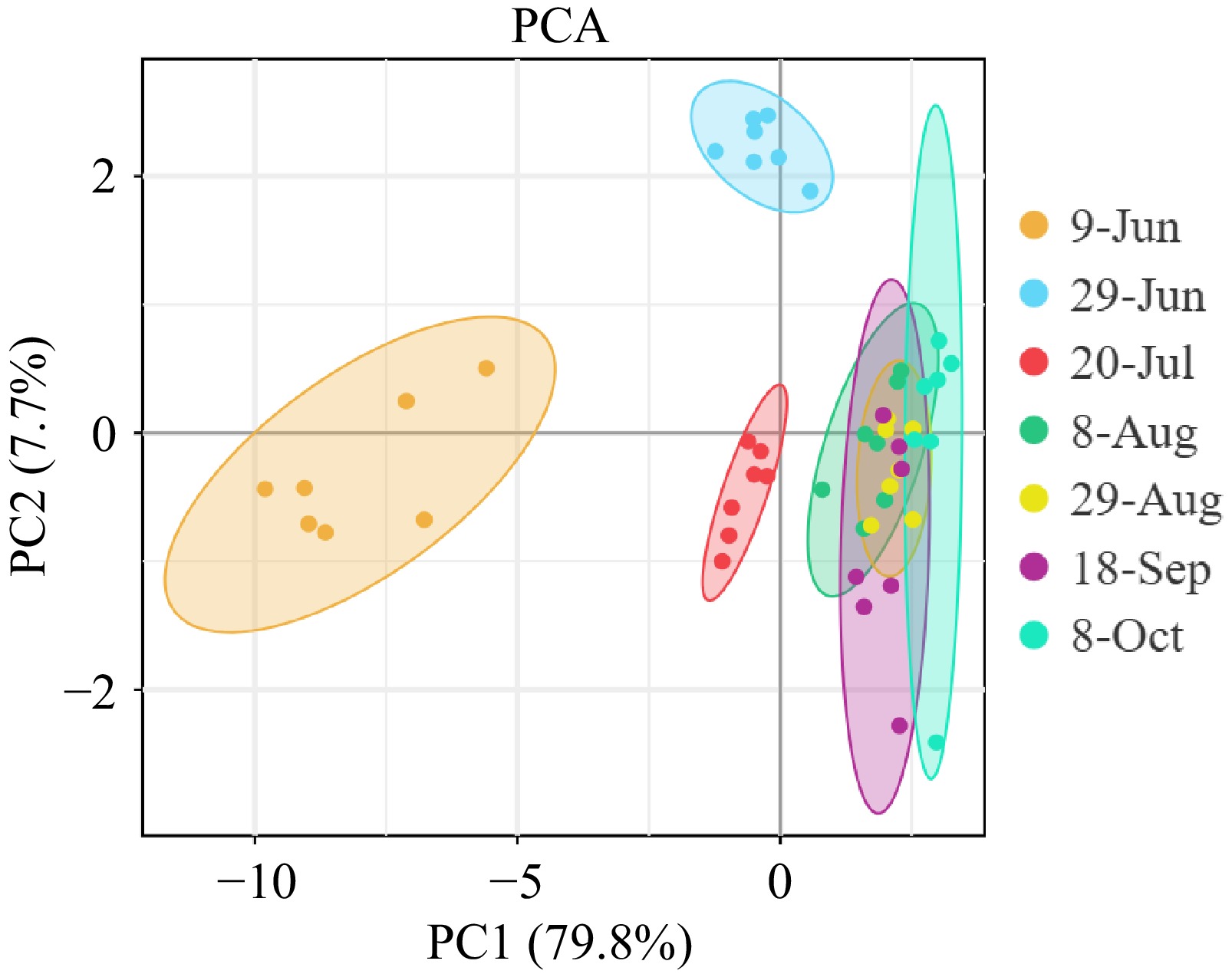
Figure 6.
PCA of polyphenols in 'Huahong' rock core at different stages with various dwarfing interstocks.
Effects of different dwarfing interstocks on polyphenol component contents in 'Huahong' apples at different developmental stages
-
Based on the trend analysis presented in Fig. 4, it was observed that there were significant variations in the polyphenol components between June 29th (early-stage), August 8th (mid-stage), and October 8th (late-stage). In conjunction with the PCA analysis outlined in effects of different dwarfing interstocks on polyphenol component contents in the core of 'Huahong' apples during late-stage, it was further revealed that these variations were influenced by the different dwarfing interstocks used.
Effects of different dwarfing interstocks on polyphenol component contents in the core of 'Huahong' apples during early-stage
-
As shown in Supplementary Fig. S1, during the early stage of fruit development, different dwarfing interstocks influenced the contents of epicatechin, cryptochlorogenic acid, procyanidin B2, and phloridzin. Specifically, 'MD001' significantly increased the content of phloridzin compared to other interstocks. 'SH38' notably elevated the levels of phloridzin-unknown2, procyanidin B2, and epicatechin. 'SH3' significantly increased the contents of procyanidin B2 and procyanidin C1.
Effects of different dwarfing interstocks on polyphenol component contents in the core of 'Huahong' apples during mid-stage
-
As shown in Supplementary Fig. S2, during the mid-stage of fruit development, 'CG24' significantly increased the contents of chlorogenic acid, 4-O-p-coumaroyl quinic acid, procyanidin B2, procyanidin C1, phloridzin, phloridzin-unknown 4, and phloridzin in the fruit core. 'MD001' significantly elevated the levels of procyanidin B2, procyanidin C1, and epicatechin. 'SH3' notably increased the content of procyanidin C1. However, compared to the control, all intermediate rootstocks significantly reduced the content of phloridzin-unknown 5, with statistically significant differences.
Effects of different dwarfing interstocks on polyphenol component contents in the core of 'Huahong' apples during late-stage
-
During the late stage of fruit development, as shown in Supplementary Fig. S3, 'MD001' significantly increased the contents of quercetin-3-glucoside and 5-O-p-coumaroyl quinic acid. Both 'CG24' and 'SH38' elevated the level of phloridzin, although the differences compared to the control were not statistically significant.
Comparative analysis of polyphenol content in 'Huahong' apples across different stages
-
As shown in Fig. 7, ternary plots were constructed for the polyphenol composition of 'Huahong' fruit cores at three stages: June 29th (early stage), August 8th (mid-stage), and October 8th (late stage). The aim was to reveal the relative content changes of polyphenol components across these stages. From the figure, it can be observed that the relative levels of 5-O-p-coumaroyl quinic acid, Epicatechin, Procyanidin B2, and Procyanidin C1 gradually decreased as the fruit developed. Concurrently, the concentrations of Quercetin-3-rhamnoside, 4-O-p-coumaroyl quinic acid, Chlorogenic acid, and Phloridzin progressively increased with fruit development. The concentration of Quercetin-3-glucoside exhibited a different trend; it initially increased as the fruit developed and then gradually decreased during the fruit ripening stage.
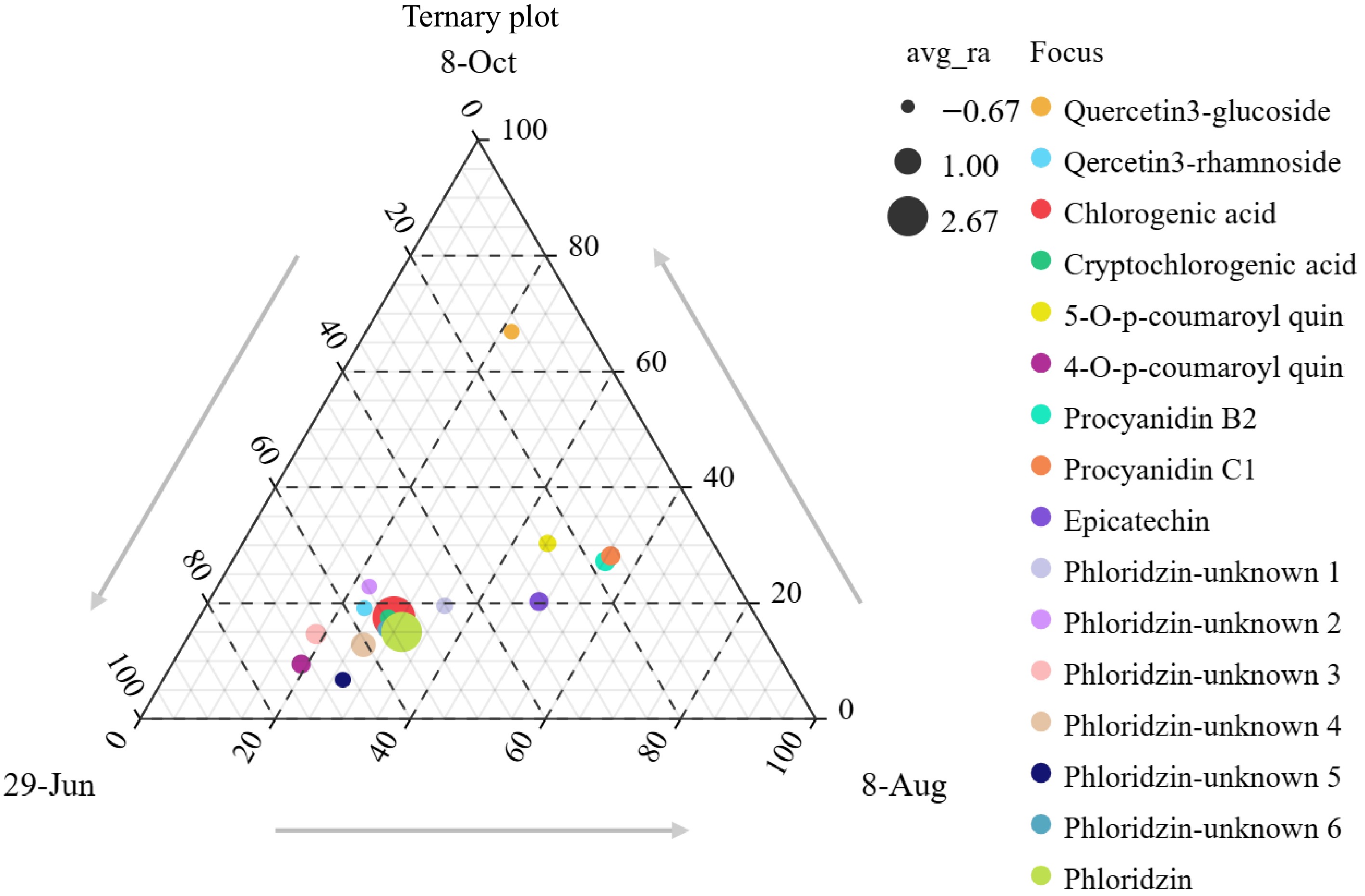
Figure 7.
Ternary plot of polyphenol content during the early, mid, and late stages of 'Huahong' fruit core development.
-
The composition and content of polyphenols in apples varied significantly depending on factors such as variety, tissue location, growth environment, and maturity[21]. The polyphenol content in wild apple resources was typically higher than that in cultivated varieties. Apples used for winemaking generally had higher polyphenol content than those for fresh consumption or juice production. Apples with bitter and astringent tastes had higher polyphenol content than sour or sweet apples[22,23], and red-skinned apples had higher polyphenol content than yellow- or green-skinned apples[24]. A study by Lata et al.[25] on the polyphenol composition and content of 19 apple varieties found that variety was a crucial factor influencing polyphenol content. Varieties such as 'Red Rome', 'Idared', 'Fiesta', 'Fuji', 'Gloster', 'McIntosh', and 'Pilot' had higher chlorogenic acid content, while 'Elan', 'Elstar', and 'Jonama' had lower chlorogenic acid content. 'Prima', 'Pilot', and 'Elstar' had high epicatechin and catechin content, while 'Starking Delicious', 'Gloster', 'Golden Delicious', and 'Granny Smith' had higher phloridzin content. Light conditions were also an important factor affecting the polyphenol composition and content in apple fruits. Studies showed that fruits within the canopy, shaded by leaves and branches, generally had lower polyphenol content than those on the canopy periphery[26]. Felicetti & Schrader[27] demonstrated that in Fuji apples, direct sunlight exposure increased the content of anthocyanins, epicatechin, chlorogenic acid, and quercetin glycosides. However, excessive light intensity could lead to solar radiation and high-temperature stress, resulting in a decrease in anthocyanin content and an increase in epicatechin, chlorogenic acid, and quercetin glycoside content. Differences in polyphenol composition and content were also significant among different parts of mature fruits. The total polyphenol content was lowest in the pulp, higher in the core, and highest in the peel, with the peel typically containing 2 to 6 times more polyphenols than the pulp[28]. In this study, the total polyphenol content in the core ranged from 3,518.79 to 4,196.04 mg/kg in the early developmental stage and from 434.45 to 514.39 mg/kg at maturity. Compared to previous studies[29], the total polyphenol content in the peel was 5,254.81 to 7,224.33 mg/kg in the early developmental stage and 1,117.50 to 1,486.44 mg/kg at maturity. The pulp had a total polyphenol content of 3,579.34 to 4,466.904 mg/kg in the early developmental stage and 308.86 to 358.24 mg/kg at maturity. Across different stages of fruit development, the peel had the highest total polyphenol content, followed by the core, and the pulp had the lowest.
Previous studies have indicated the presence of 34 polyphenol species in apples[30]. Nie et al[17]. found marked differences in the content and composition of polyphenols among different fruit parts of 'Changfu 2' apples In an earlier investigation of polyphenols in the peel and pulp of 'Huahong' apples, 17 polyphenol components were identified in the peel, whereas 12 were detected in the pulp[31]. In this study, 16 polyphenol components were identified in the core, highlighting the variations in polyphenol composition across different apple fruit parts. Six phloridzin components identified in this study remain unnamed, suggesting the need for more advanced techniques for their identification. In addition to their potential in the food and pharmaceutical industries, fruit core polyphenols also show promise in pest control. For example, polyphenols extracted from fruit cores, such as those from peaches and pears, could potentially be used as natural protectants against pests like the peach and pear fruit moth (Cydia pomonella). These bio-extracts offer a sustainable alternative to synthetic pesticides, contributing to environmentally friendly pest management practices.
Effect of different developmental stages on polyphenol composition and content in apple fruits
-
Pang[32] revealed that the total phenol content was highest during the early stages of apple growth and development, gradually decreasing as time progressed. In this study, as 'Huahong' apples developed, the total phenolic content in their cores progressively decreased. The decline was rapid during the initial developmental stages, followed by stabilization or a slight decrease as the fruit matured. Notably, the trend in total phenolic content remained consistent across different dwarfing interstocks throughout all developmental stages.
The content of apple polyphenols was related to fruit maturity. The concentrations of flavanols, flavonols, and chlorogenic acid exhibited a decreasing trend as the apples grew and developed. They declined rapidly in the early stages of development, then the rate of decline gradually slowed, and finally stabilized or slightly decreased[33]. In the young fruit stage, flavanols, and flavonols constituted a smaller proportion, while chlorogenic acid was the predominant polyphenol in the fruit pulp. As the fruit developed and enlarged, the proportion of chlorogenic acid gradually decreased, whereas the proportions of flavanols and flavonols gradually increased[34]. In mature apple fruits, flavanols, and procyanidins became the major polyphenols, surpassing chlorogenic acid in proportion[35]. Studies showed that the polyphenol content in both the apple peel and pulp gradually decreased with fruit maturity, stabilizing after the fruit reached maturity[36]. The total polyphenol content of 'Hongxun No. 2' and 'Xinjiang Wild Apple' gradually decreased as the fruit developed, with a reduced decline and stabilization occurring between 95 and 110 d after flowering[37]. However, the total polyphenol content in the peel of 'Xinjiang Wild Apple' increased at 65 d after flowering and then gradually decreased[38]. Kondo et al.[36] conducted a study on the impact of harvest time on the total polyphenol content of apples. Mature fruits from 14 apple varieties were harvested three times, with a two-week interval between each harvest. Analysis revealed no significant differences in the total polyphenol content among the three harvests. All five categories of polyphenol components in apples gradually decreased with fruit development. Renard et al.[39] studied two apple varieties used for winemaking and found that from 35 to 100 d after flowering, the total polyphenol content of the fruits decreased, with rapid reductions in flavanols, dihydrochalcones, and flavonols, and a slower decline in hydroxycinnamic acids. In this study, the flavonol content fluctuated across developmental stages but showed an overall decreasing trend. The contents of quercetin, procyanidins, and epicatechin all gradually decreased. Studies indicated that flavanols were synthesized in large quantities in the early stages of fruit development and then increased slowly during the developmental process. Fruit enlargement led to a reduction in flavanol content, while an increase in total polyphenol content might be attributed to the proportion of the increase in fresh weight being less than the proportion of the increase in flavanol content[40,41]. Lata & Verma[42] reported that total phenol content and antioxidant activity significantly increased during fruit ripening, while flavonoid content remained relatively stable across different varieties and harvest times. In this study, the flavonoid content in apples with different dwarfing interstocks stabilized during the later stages of fruit development.
Effects of different dwarfing interstocks on the polyphenol components, and content in apples
-
Dwarfing interstocks may influence polyphenol content and composition in apple cores through several physiological and molecular mechanisms. These include altering hormonal balances, which can affect gene expression related to polyphenol biosynthesis. Additionally, changes in resource allocation due to modified rootstock architecture could lead to differences in nutrient availability, impacting the metabolic pathways that produce polyphenols. Environmental stress responses, which can be modulated by rootstocks, might also play a role, as stress can induce the production of certain polyphenols as part of the plant's defense mechanisms. Furthermore, dwarfing interstocks could affect the microRNAs or transcription factors that regulate polyphenol-related genes, thereby influencing the accumulation and types of polyphenols in the apple core. The effects of dwarfing interstocks on apple phenolic compounds are multifaceted. A study by Xie et al.[43] on 'Gala' apples found that grafting onto different rootstocks influences phenolic content, with the highest total phenol content observed when 'Gala' was grafted onto 'A1'. Dwarfing interstocks can generally enhance the functional component content (total sugars, soluble solids, and phenolic compounds) of certain apple varieties. 'Mac9' and 'CG24' grafted 'Honeycrisp' apples exhibited the highest overall evaluation of functional component content[44]. In this study, during the mid-stage of fruit development, 'CG24' significantly increased the concentrations of chlorogenic acid, 4-O-p-coumaroyl quinic acid, procyanidin B2, procyanidin C1, phloridzin, phloridzin-unknown 4, and an additional phloridzin derivative in the apple core. Simultaneously, 'SH38' notably elevated the levels of phloridzin-unknown2, procyanidin B2, and epicatechin. However, in the late-stage of development, while both 'CG24' and 'SH38' resulted in higher phloridzin levels, these differences were not statistically significant compared to the control. Kviklys et al. suggested that rootstocks have a minor impact on the content of quercetin derivatives, epicatechin, and chlorogenic acid in apples[45]. In our experiment, different dwarfing interstocks did not significantly affect the content of epicatechin and chlorogenic acid in the core of 'Huahong' apples but increased the content of quercetin derivatives to varying degrees. 'Gala' apples grafted onto 'M9' rootstock had significantly higher total polyphenol and anthocyanin content than those grafted onto M26 rootstock. Additionally, the 'M9' combination exhibited higher activity of key enzymes in the anthocyanin synthesis pathway, including Phenylalanine Ammonia-Lyase (PAL), Chalcone Synthase (CHS), and Dihydroflavonol 4-Reductase (DFR)[46]. Rootstocks may also affect the stability of apple polyphenols. 'Gala' apples grafted onto 'M26' rootstock had lower anthocyanin content, possibly due to the poor stability of anthocyanins in the fruit, leading to partial degradation during fruit ripening[47].
The total phenol content in the peel and pulp of apples from the dwarfing interstock M26 combination ('Yanfu 6' / 'M26' / Pingyi Sweet Tea) was significantly higher than that of the control ('Yanfu 6' / Pingyi Sweet Tea)[48]. Wang et al.[44] reported that grafting 'Honeycrisp' onto dwarfing interstocks resulted in a significant increase in polyphenol content compared to standard rootstocks. The degree of dwarfing of the rootstock also influences the fruit's phenolic acid content, with rootstocks that induce stronger dwarfing effects leading to higher phenolic acid content in the fruit. Fallahi & Mohan[49] found that different rootstocks can also affect the anthocyanin content of the fruit. During fruit development, different interstocks have varying effects on the polyphenols in the fruit core. In previous studies on the peel and pulp[32], it was found that during fruit ripening, the 'SH3', 'SH38', and 'Mac9' combinations significantly increased the flavonol content in the peel; the 'MD001' combination significantly increased the hydroxycinnamic acid content in the pulp; the 'Mac9' and 'CX5' combinations had lower epicatechin and chlorogenic acid content in the pulp; and the 'Mac9' and 'SH38' combinations significantly increased the anthocyanin content in the peel. In this study, during fruit ripening, 'MD001' significantly increased the contents of quercetin-3-glucoside and 5-O-p-coumaroyl quinic acid, exerting a notable influence on the polyphenol composition. Environmental factors likely interact with dwarfing to influence polyphenol content in apple cores through complex physiological responses, and conducting additional experiments could provide valuable insights into these interactions. The findings on the effects of different dwarfing interstocks on the total phenols and polyphenol compounds in the core of 'Huahong' apples can be applied to practical apple production. Apple producers can select the most suitable dwarfing interstocks based on the desired polyphenol content and fruit quality. By choosing the right interstocks, producers can optimize the polyphenol profile of their apples, leading to higher-quality fruits that meet consumer demands for health-promoting properties. Additionally, understanding the impact of interstocks on polyphenol dynamics during fruit development can guide producers in making informed decisions about harvest timing and post-harvest handling to maximize the retention of these beneficial compounds. By applying these findings to actual production, apple core processing products with higher nutritional value and market competitiveness can be developed, such as juices, jams, or functional food additives. Selecting apple varieties with specific dwarfing interstocks can increase the polyphenol content, thereby enhancing the antioxidant properties of the products and meeting consumers' demand for healthy foods.
-
The fruit core contains a substantial amount of polyphenols, situated between the fruit peel and pulp. By grafting onto dwarfing interstocks, the content of polyphenols and their components in the fruit core can be enhanced, thereby maximizing the value of the fruit core. Furthermore, the potential application of fruit core polyphenols in pest control offers a promising alternative to synthetic agricultural practices. In the cores of 'Huahong' apples, the total phenolic content gradually decreases as the fruit develops. During the fruit ripening stage, the polyphenol compositions in different dwarfing interstock combinations are similar, with Quercetin-3-glucoside, Quercetin-3-rhamnoside, and Phloridzin being the dominant components. The combinations of 'SH3', 'SH38', and 'CG24' significantly increase the polyphenol content in 'Huahong' apples during the mid-stage of fruit development, but the phenolic content becomes lower than that of the control during the later stages of development. The 'MD001' combination slows the decline in polyphenol components during the mid- to late-stages of fruit development, resulting in a substantial accumulation of core polyphenols at maturity. Taken together, the 'MD001' combination is the optimal rootstock for enhancing phenolic compounds in the apple core.
This work was supported by the Agricultural Science and Technology Innovation Program (CAAS-ASTIP-2021-RIP-02) and Fundamental Research Funds for Central Non-profit Scientific Institution (Grant No. 161018201614).
-
The authors confirm contribution to the paper as follows: study conception and design: Wang D, Wang K, Gao Y, Feng J; data collection: Li Q, Li L; analysis and interpretation of results: Li Q, Lu X, Liu Z, Wang L; draft manuscript preparation: Li Q, Sun S. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article, and are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 The total phenolics in core of 'Huahong' Apple.
- Supplementary Table S2 Trend analysis of dynamic changes.
- Supplementary Fig. S1 Effects of different dwarfing interstocks on polyphenol component contents in the core of 'Huahong' Apples during early-stage.
- Supplementary Fig. S2 Effects of different dwarfing interstocks on polyphenol component contents in the core of 'Huahong' Apples during mid-stage.
- Supplementary Fig. S3 Effects of different dwarfing interstocks on polyphenol component contents in the core of 'Huahong' Apples during late-stage.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li Q, Lu X, Liu Z, Feng J, Gao Y, et al. 2025. Effects of different dwarfing interstocks on the total phenols and compounds of polyphenols in the core of 'Huahong' apples. Fruit Research 5: e017 doi: 10.48130/frures-0025-0008 

Effects of different dwarfing interstocks on the total phenols and compounds of polyphenols in the core of 'Huahong' apples
- Received: 30 December 2024
- Revised: 20 February 2025
- Accepted: 05 March 2025
- Published online: 06 May 2025
Abstract: Polyphenols are recognized as a class of compounds possessing a wide range of biological activities that provide numerous health benefits to humans. Apples are abundant in polyphenolic substances. As an integral part of the fruit's internal structure, the polyphenols found in fruit cores cannot be overlooked. This study examines the dynamic effects of six different dwarfing interstocks on the total phenolic content and polyphenol compounds in the core of 'Huahong' apples, aiming to gain a more comprehensive understanding of the phenolic components and their contents in fruit cores, providing novel insights into the comprehensive utilization of fruits. The 'CG24', 'SH38', 'SH3', 'MD001', 'Mac9', and 'CX5' were grafted onto Malus baccata (L.) Borkh. and 'Huahong' was grafted on six interstocks mentioned above, the self-root grafting Malus was used as a control (CK). The components and contents of polyphenols in cores were analyzed at seven periods during 'Huahong' apple growth by using High Efficiency Liquid Chromatography and Mass Spectrometry, and the content of total phenols was determined by the Folin-Ciocalteu method. The polyphenol concentration in the fruit core exceeds that of the peel but is lower than that found in the pulp, and this content, along with its constituent components, exhibits a gradual decline as the fruit undergoes development. At maturity, the polyphenol composition is consistent across dwarfing interstock combinations, primarily comprising Quercetin-3-glucoside, Quercetin-3-rhamnoside, and Phloridzin. The 'SH3', 'SH38', and 'CG24' combinations temporarily increase polyphenol content during mid-development, but these levels drop below the control group in later stages. In contrast, 'MD001' effectively mitigates the reduction of polyphenols from mid- to late development, leading to substantial phenolic accumulation at maturity. Among the interstocks examined, 'MD001' demonstrates its superiority in enhancing phenolic compounds. Interstocks can increase the levels of polyphenolic compounds in the core of apples. The polyphenol content in the fruit core is intermediate between that of peel and flesh, exhibiting significant application value in processing and the extraction of polyphenol components.
-
Key words:
- Interstocks /
- Total phenols /
- Polyphenols /
- Core