









-
Plants require a significant array of TFs to effectively regulate numerous processes related to growth and development, such as cellular morphogenesis and the transmission of signals in reaction to various environmental stresses[1]. TFs influence the transcriptional activity of their target genes through their binding to particular cis-regulatory elements, either enhancing or inhibiting this process[2,3]. As a result, the roles and structures of TFs have garnered considerable attention from researchers and are currently prominent topics within the realm of plant molecular biology. Given their crucial roles in gene expression regulation and evolutionary mechanisms in plants, the examination of TFs is of significant interest. TFs are categorized into various families based on their conserved domains, such as NAC, bZIP, WRKY, bHLH, AP2/ERF, TCP, Cys2-His2 zinc finger (ZF), among others[3]. Various kinds of TFs have evolved to control the expression of specific genes or signals unique to plants. The DOF gene family is one such group of plant-specific TFs commonly found in higher plants. Since the initial isolation of the first DOF gene (Zmdof1) from maize[4], many DOF genes have been investigated in various other plant species[1,5−8].
DOF TF proteins are composed of 200 to 400 amino acids and feature a conserved DOF domain at the N-terminal region, which includes a ZF structure made up of 50 to 52 amino acid residues, alongside a transcriptional regulatory domain at the C-terminal region[9]. In contrast to other zinc finger proteins, DOF TFs possess a single Cys2/Cys2 (C2/C2) zinc finger, which specifically interacts with the upstream cis-elements (T/A)/AAAG of their target genes[9,10]. Previous studies in Arabidopsis have discovered 37 DOF members, with one being designated as a pseudogene[7], and 30 DOF members have been reported in rice[11]. To date, a growing number of DOF gene members have been discovered in different species[3], including 18 in maize[12], 41 in poplar[13], 60 in apple[6], 26 DOF genes in birch[14], and 24 in rose[15]. Although the variation among species, the roles of most DOF TFs in the monocot model organism remain unclear. In addition, the debate over whether there are functional differences or redundancy among DOF proteins is particularly contentious. Some researchers argue that the roles of DOFs exhibit a considerable degree of redundancy[16,17], while others contend that there are significant distinctions among them[18].
DOF family members play a crucial role in overseeing the growth and developmental phases of various plant tissues, including root development, elongation of the hypocotyl, plant shape formation, leaf growth, and the development of floral organs[16,19]. DOF family genes also play a role in various biological processes in plants, particularly in how they respond to both biotic and abiotic stresses. For instance, DOF genes contribute to viral resistance in tobacco and pepper, and they also enhance resistance to fungi in cucumber[20−22]. Additionally, DOF genes are engaged in responses to several abiotic factors like photoperiod and temperature and are crucial during different developmental stages, including flowering, seed development, and leaf senescence[21,23,24]. These response mechanisms are typically linked to the regulation of phytohormones, indicating that DOF genes may also be regulated by these hormones[25]. Furthermore, DOF genes are also involved in processes such as dormancy and seed germination, fruit ripening, and plant metabolism[3].
Citrus fruits rank among the most significant agricultural products globally, supplying essential nutrients to humans, including various vitamins, citric acid, and carotene[26]. Among citrus varieties, the sweet orange (Citrus sinensis) is extensively grown due to its appealing appearance and delightful flavor. Nonetheless, the quality of citrus fruits can be significantly influenced by various cultivation methods and environmental factors, such as cold, drought, and salt stress[27−29]. Previous studies have demonstrated that DOF TFs are involved in the stress response mechanisms of various plant species[30−32]. However, comprehensive studies investigating the specific physiological functions of DOF genes in citrus are lacking. Given that DOF genes are vital for numerous developmental processes and responses to diverse environmental challenges, it is essential to conduct an in-depth investigation of the DOF gene family in citrus. This study aims to explore the chromosomal distribution and expression patterns of CsDOF genes under drought conditions, conduct a phylogenetic analysis comparing these genes with the DOF TFs of Arabidopsis, and evaluate their potential roles in development through sequence similarity, evolutionary relationships, gene expression analysis, and conserved motif examination. In addition, we further investigated the functional characteristics of CsDOF10 following its overexpression in Arabidopsis. The results of this study will provide an essential foundation for the functional analysis of unexplored DOFs in citrus and will inform future research on the response of citrus to drought stress.
-
Citrus sinensis trees were cultivated at the Citrus Breeding Center located at Huazhong Agriculture University in Wuhan, China. During the flowering and fruiting seasons, samples were collected from ten-year-old trees, including mature leaves, tender roots, fully open flowers, tender stems, and fruit that was 25 d post-flowering. After being collected, all samples were immediately frozen in liquid nitrogen and subsequently kept in an ultra-low temperature freezer for future use. Additionally, wild-type Arabidopsis (Columbia), tobacco (N. tabacum), and transgenic plants with overexpression traits were grown in a greenhouse maintained at 24 °C, maintaining a relative humidity of 60% along with a cycle of 16 h of light and 8 h of darkness.
For the drought treatment, two-year-old potted seedlings of sweet orange grafted onto trifoliate orange rootstock were utilized in this study. The entire treatment process was conducted in a controlled temperature plant growth chamber maintained at 24 °C. The sweet orange trees were divided into two groups: a treatment group and a control group, with three trees in each group (three biological replicates). The control group received regular watering at consistent intervals, while the experimental group was not watered. After two weeks, the sweet orange trees in the experimental group began to exhibit curled leaves, at which point leaf samples were collected for further experimentation. The experimental design for treating Arabidopsis is analogous to that for sweet orange. After one week of treatment, wild-type Arabidopsis seedlings begin to display observable phenotypes. Leaf samples are then promptly collected, and physiological indicators are assessed.
Identification of DOF TFs in Citrus sinensis
-
A total of 36 DOF TFs from Arabidopsis were obtained from the TAIR database, along with 24 citrus CsDOF TFs sourced from the PlantTFDB v4.0 (
http://planttfdb.cbi.pku.edu.cn/ )[33]. To confirm the presence of a conserved Dof domain in the identified proteins, we utilized the SMART and Pfam databases[34]. The conserved motifs of CsDOF proteins were subsequently examined using the Multiple Expectation Maximization for Motif Elicitation (MEME) version 5.4 (https://meme-suite.org/meme/tools/meme ) with default settings, identifying motifs ranging from 6 to 50 residues in width[35]. For the bioinformatics analysis of CsDOF protein characteristics, including molecular weight (MW), length of protein and number of intron/exon, theoretical isoelectric point (pI), instability and aliphatic index, scaffold location, grand average of hydropathicity (GRAVY), the ProtParam online tool was utilized as previously described[36].Scaffold location and phylogenetic analysis of CsDOFs
-
The positions of CsDOFs were sourced from the JOINT GENOME INSTITUTE (JGI) database (
https://phytozome-next.jgi.doe.gov/ ), and visualized using MapInspect software. The amino acid sequences of CsDOFs were aligned using ClustalX software, as detailed in a prior study[37]. Following this, Bayesian and Neighbor-Joining (N-J) phylogenetic trees were generated with 1,000 bootstrap replications according to the methodology described by Tamura et al.[38]. Additionally, maximum likelihood and minimum evolution methods were utilized for tree construction to corroborate the findings from the N-J method in MEGA 6.0.Sequence characteristics analysis of CsDOF genes
-
To analyze the promoter sequence characteristics of CsDOF genes, the PlantCARE tool was employed to investigate the cis-acting elements located in the 2,000 bp promoter region upstream of the CsDOFs. The exon-intron arrangements of CsDOFs were examined by comparing the coding sequences (CDS) with the genomic sequences using the Gene Structure Display Server v2.0[39].
Vector construction and plant genetic transformation
-
The CDS of CsDOF10 was amplified using the EX Taq DNA Polymerase (TaKaRa). The PCR protocol consisted of an initial denaturation, denaturation, annealing, and elongation steps, with specific parameters established in accordance with our previous description[37]. The complete CDS was then ligated into the pBI121 vector and subsequently transformed into A. tumefaciens via the heat shock method to facilitate plant transformation. The primers utilized for cloning the CsDOF10 are listed in Supplementary Table S1. For the transformation of Arabidopsis plants, the floral dip method was employed according to the description of a previous study[40].
RNA extraction of citrus tissue and quantitative real-time PCR (qRT-PCR)
-
Total RNA extraction from citrus plants was carried out using the RNeasy Plant Mini kit (Qiagen, Hilden, Germany). Following the quality assessment, total RNA was utilized for reverse transcription experiments, employing the PrimScript™ kit (TaKaRa, Otsu, Japan) for the RNA reverse transcription process. Primer design, qRT-PCR protocols, and data analysis were conducted in accordance with our earlier research[41]. The PCR products ranged from 100 to 300 bp, and their specificity was verified through PCR following primer synthesis. The analysis of qRT-PCR was conducted using the ABI PRISM 7000 system from Applied Biosystems, and each sample included three biological replicates. The citrus Actin gene served as the reference gene. The gene expression levels were assessed based on their expression changes relative to the reference gene. The relative expression of the CsDOF genes was calculated using the 2−ΔΔCᴛ method[42].
Histochemical GUS analysis
-
The promoter region of CsDOF10, spanning from −1 to −1,500 bp relative to the start codon, was subcloned into the pBI101 vector containing the GUS tag, resulting in the pCsDOF10:GUS construct. This fusion plasmid, pCsDOF10:GUS, was then introduced into A. tumefaciens GV3101. Subsequently, pCsDOF10:GUS was transformed into Arabidopsis, and T3 generation seedlings were utilized to assess GUS expression. The analysis of GUS activity in the transgenic plants was conducted using a histochemical staining method that has been outlined in a prior study[37].
Subcellular localization and transcription activity analysis of CsDOF10
-
To examine the subcellular localization of CsDOF10, we eliminated the stop codon from CsDOF10 and integrated it into the pBI121 vector, which contains the CaMV35S promoter and features a GFP tag, resulting in the construction of the 35S:CsDOF10-GFP vector. The positive control was provided by 35S:GFP, while VirD2NLS-mCherry functioned as a marker for the nucleus. The fusion vector plasmids 35S:CsDOF10-GFP were then electroporated into A. tumefaciens GV3101, followed by transient expression in the epidermal cells of tobacco leaves. Green fluorescence signals were detected after 48 h of growth through the use of a confocal fluorescence microscope. This procedure is in accordance with previously reported methods[43].
To analyze transcriptional activity, the sequence encoding CsDOF10 protein was cloned into the pGBKT7 vector. Subsequently, the pGBKT7-CsDOF10 fusion vector was introduced into yeast AH109 and selected on SD/-Trp medium. The transcriptional activity of CsDOF10 was evaluated by analyzing yeast growth on SD/-Trp/-His medium that was enhanced with X-α-galactosidase, as outlined in a previous study[44].
Yeast one-hybrid (Y1H) and dual-luciferase (LUC) assays
-
The Y1H assay was conducted following the method described by Zeng et al.[45]. The CDS of CsKNOX9 was amplified and cloned into the pGADT7 vector to create the effector construct AD-CsKNOX9, while the sequence of the CsDOF10 promoter was cloned into the pAbAi vector, resulting in the creation of the reporter construct pAbAi-pCsDOF10. Both the effector and reporter constructs were subsequently cotransformed into the YIHGold yeast strain, and the yeast cells were screened on SD/-Leu medium and also on SD/-Leu medium that was enriched with 100 ng/mL Aureobasidin A (AbA). The combination of pGADT7 and pAbAi-pCsDOF10 acted as the negative control.
In the LUC assay, The CDS of CsKNOX9 was incorporated into the pGreenII 62-SK vector to function as the effector. Meanwhile, the promoter region of CsDOF10 was cloned into the pGreenII 0800-LUC vector to act as the reporter. The pGreenII 62-SK vector lacking any insert, along with reporters containing the CsDOF10 promoter, served as a negative control. All constructs were introduced into A. tumefaciens GV3101 (pSoup-p19) via electroporation. An infiltration buffer composed of 10 mM MES, 10 mM MgCl2, and 150 mM acetosyringone at pH 5.6 was utilized for the injection. For the dual-luciferase assays, N. benthamiana leaf epidermal cells were employed as previously described[46], and the ratio of LUC to REN was assessed using the dual-luciferase reporter assay system provided by Promega. Details of the primers used for vector construction can be found in Supplementary Table S1.
Statistical analysis
-
All statistical analyses were conducted using DPS software. The error bars represent the standard deviation from various replicates. For graphing, GraphPad Prism 8 software was utilized, and the t-test was employed to identify significant differences (* p < 0.05; ** p < 0.01).
-
Initially, 28 CsDOF protein sequences from sweet orange were retrieved from the JOINT GENOME INSTITUTE database (
https://phytozome-next.jgi.doe.gov/ ). Subsequently, the presence of conserved Dof domains in the candidate sequences was verified using the SMART and Pfam websites. After eliminating some duplicate sequences and pseudogenes, 24 members of the CsDOF TF family were discovered in Citrus sinensis. These genes are located across 19 scaffolds (Fig. 1a). For the sake of convenience, we named these genes as CsDOF1 to CsDOF24 according to their order of appearance on scaffold.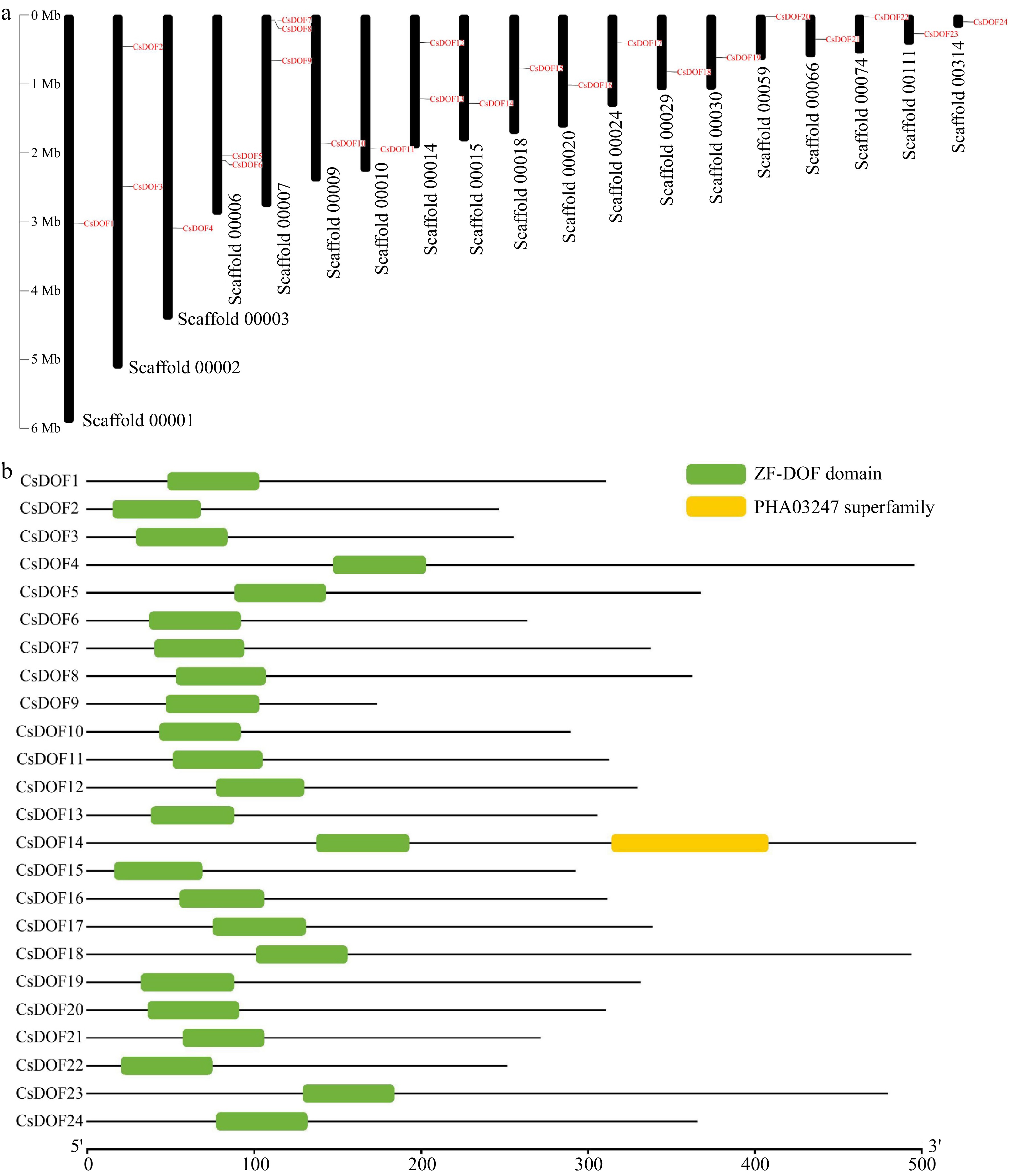
Figure 1.
Analysis of the position and structure of sweet orange DOF family members. (a) The positional information of CsDOF TFs on the chromosomes (scaffolds) of the sweet orange genome. (b) Conserved domain of CsDOF TFs.
We conducted a further analysis of the structures of these 24 DOF proteins using the TBTOOLS software, revealing that the conserved DOF domains are predominantly situated near the proteins' N-terminus. Notably, DOF14 also possesses an additional PHA03247 superfamily domain (Fig. 1b). The amino acid counts for the CsDOF-encoding proteins ranged from 172 for CsDOF9 to 496 for CsDOF14, with pI values spanning from 4.86 to 9.35, and MWs between 19.36 and 54.78 kDa (Supplementary Table S2). An analysis of the DNA sequence structure indicated that the DNA is relatively compact, measuring less than 4,000 bp in length and containing a maximum of 3 exons (Supplementary Fig. S1). Detailed information, including coding gene ID, intron and exon counts, instability index, and aliphatic index, is provided in Supplementary Table S2. These findings suggest that the variation in amino acid sequence lengths among CsDOFs may reflect adaptations to differing functional demands and physical or chemical characteristics.
Characterization analysis of CsDOFs in sweet orange
-
To uncover conserved motifs, we analyzed the CsDOF protein sequences using the MEME suite[35]. The first motif (Motif 1) is the Dof domain, which is notably conserved (Supplementary Fig. S2a). Each CsDOF protein exhibits a specific distribution of this motif, as illustrated by the consensus sequence logo, representing a conserved Dof domain motif consisting of 50 amino acids (Supplementary Fig. S2b). The remaining amino acid sequences within the CsDOF proteins are largely divergent. Besides the highly conserved Dof domain, the MEME analysis identified several common conserved motifs across CsDOF proteins (Supplementary Fig. S2a−b). However, these motifs are neither common nor unique to members of the CsDOF family, a search in the PROSITE database for known motifs indicated that these protein motifs do not have any identified functions[47]. Subsequently, we performed a sequence alignment analysis of the Dof domain structure in CsDOF proteins, which revealed that their Dof domain sequences exhibit significant similarity and strong conservation (Supplementary Fig. S3).
To investigate the homologous relationship between CsDOFs in sweet orange and the DOF family in Arabidopsis, we conducted a phylogenetic analysis of 24 CsDOF proteins from sweet orange and 36 DOF family members from Arabidopsis. The results indicated that these 60 DOFs can be divided into six categories: A, B, C, D, E, and F (Fig. 2). Among the 24 proteins identified in sweet orange, six CsDOF proteins belong to group A, five to group B, two to group C, eight to group D, and an additional three to group A. Notably, there are no CsDOF proteins in group E (Fig. 2).
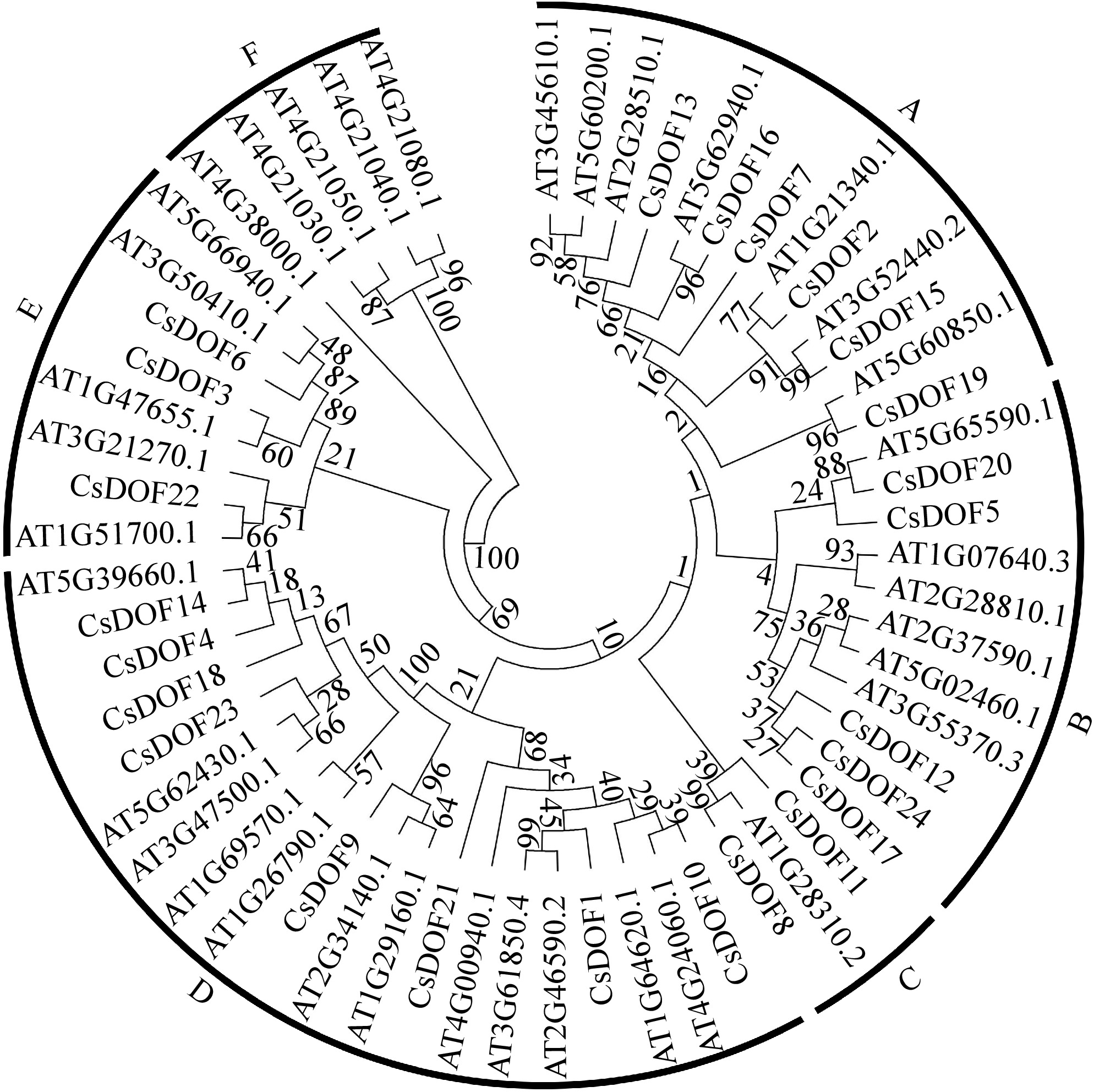
Figure 2.
Phylogenetic tree of CsDOFs from sweet orange and Arabidopsis. The complete amino acid sequences were aligned using ClustalX version 1.83, and a phylogenetic tree was generated with MEGA 7.0 employing the neighbor-joining method, supported by 1,000 bootstrap replicates.
Promoters analysis and expression profiling of CsDOF genes under drought stress conditions
-
Cis-elements are crucial for regulating gene transcription throughout plant development and in response to environmental stressors. To explore the mechanisms of transcriptional regulation, we identified the cis-elements within the 2,000 bp promoter regions of CsDOF genes using the PlantCARE database. Alongside well-known cis-acting elements such as the CAAT-box and TATA-box, numerous other cis-elements were detected within the promoter regions of 24 CsDOFs (Supplementary Fig. S4). Notably, light-responsive elements were the most abundant and were found in all CsDOF promoters. Furthermore, other less common cis-elements associated with transcription regulation were identified, relating to functions such as circadian control, protein binding, and responsiveness to hormones or other elicitors. This indicates that the transcription of CsDOFs can be influenced by various environmental and developmental factors, indicating their role in important physiological functions and developmental processes.
To further confirm the expression profile of CsDOF genes in citrus subjected to drought stress, the expression levels of 24 CsDOF genes in sweet orange leaves were examined through qRT-PCR analysis. The results allowed us to classify these 24 CsDOF genes into three distinct categories according to their expression profiles: up-regulated, down-regulated, and no significant change. Specifically, 11 CsDOF genes were identified as upregulated after drought treatment, including CsDOF2, CsDOF4, CsDOF5, CsDOF7, CsDOF10, CsDOF13, CsDOF17, CsDOF18, CsDOF21, CsDOF23, and CsDOF24. In contrast, four CsDOF genes were found to be downregulated, namely CsDOF1, CsDOF14, CsDOF15, and CsDOF22, while nine CsDOF genes showed no notable alterations in their expression levels (Fig. 3). Notably, the expression of the CsDOF10 gene significantly rose in reaction to drought stress (Fig. 3). These results imply that members of the CsDOF TF may have regulatory functions in the citrus response to drought stress.
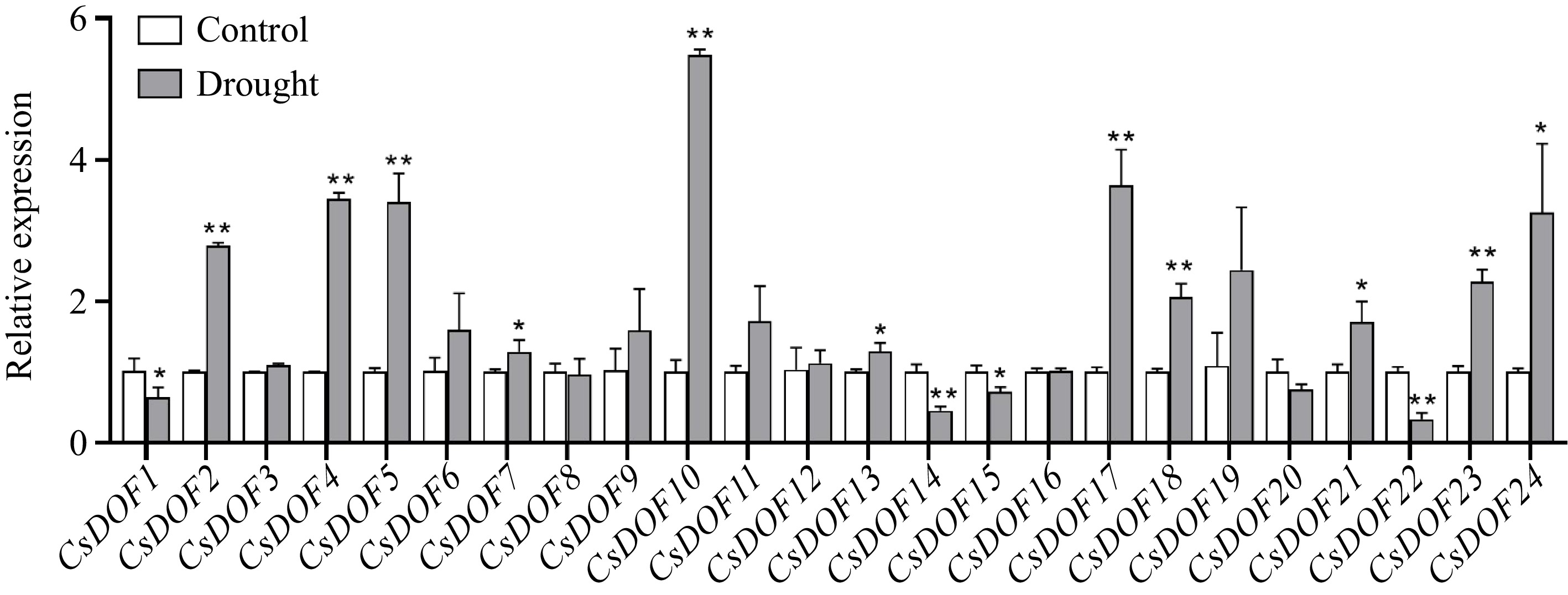
Figure 3.
Expression of CsDOFs in response to drought treatment. Fold changes of CsDOFs expression are shown. Error bars represent the standard deviations of means (n = 3). * and ** represent significant difference at p < 0.05 and p < 0.01, Students t-test (n = 3).
Overexpression of CsDOF10 regulates root development and enhances drought resistance in Arabidopsis
-
The analysis of homologous proteins revealed that CsDOF10 shares the closest homologous relationship with the Arabidopsis gene ATG04060. Protein sequence alignment between CsDOF10 and ATG04060 demonstrated that their Dof domain sequences are highly similar, differing by only a single amino acid (Fig. 4a). To investigate the regulatory function of CsDOF10 on drought stress in citrus development, we transformed the CsDOF10 gene in Arabidopsis using the overexpression vector pBI121, which contains the 35S promoter. Through screening and identification with Kanamycin, we obtained a total of eight transgenic strains (Fig. 4b). To conduct a more detailed examination of the role of CsDOF10, three independent transgenic lines, designated OE1, OE2, and OE3, were chosen from the third generation for phenotypic analysis. The 35S:CsDOF10 transgenic lines exhibit a phenotype of longer root compare to the wild-type plants (Fig. 4c). Subsequently, we performed a statistical analysis of the germination rate (Fig. 4d), plant height (Fig. 4e), and root length (Fig. 4f) of transgenic seedlings compared to the controls to assess whether CsDOF10 exerts a regulatory effect on other physiological processes. The results indicated that there were no notable differences in the rates of seed germination or plant height of 35S:CsDOF10 transgenic plants compared to the control (Fig. 4d, e); however, the root length of transgenic plants was significantly greater than that of the controls (Fig. 4f).
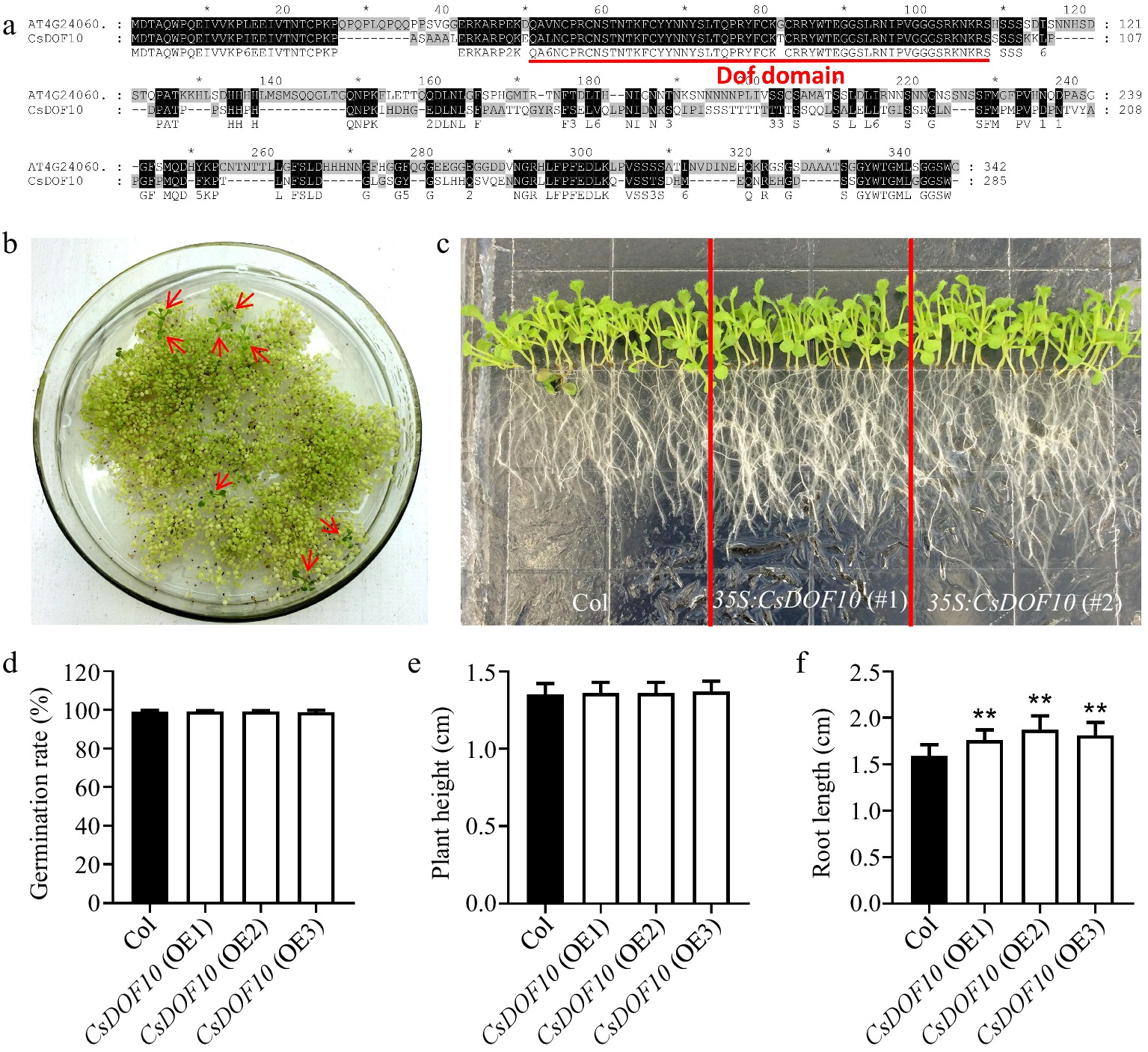
Figure 4.
Identification and functional study of CsDOF10 transgenic Arabidopsis. (a) Sequence alignment of CsDOF10 and Arabidopsis homologous protein AT4G24060 using ClustalX version 1.83. (b) Screening of transgenic positive seedlings using kanamycin. (c) Phenotypic analysis of root length in CsDOF10 transgenic Arabidopsis. (d) Statistical evaluation of seed germination rates in transgenic seedlings compared to wild-type controls. (e) Measurement of plant height in transgenic seedlings and controls. (f) Comparison of root lengths between transgenic seedlings and controls. Error bars represent the standard deviations of means (n = 10). ** represent significant difference at p < 0.01, Students t-test (n = 10).
Moreover, additional studies indicated that the formation of lateral roots in 35S:CsDOF10 transgenic Arabidopsis was notably suppressed (Fig. 5a, b). To assess drought resistance linked to CsDOF10, two distinct transgenic lines were chosen for drought evaluation. Before drought treatment, there were no significant phenotypic variations between the 35S:CsDOF10 transgenic lines and the controls (Fig. 5c). However, under drought conditions, marked differences emerged between the transgenic plants and the controls (Fig. 5c). The leaf rolling in 35S:CsDOF10 transgenic Arabidopsis occurred significantly earlier than in the control plants (Fig. 5c). Furthermore, key physiological parameters were assessed under drought conditions. Following drought exposure, the Peroxidase (POD) levels were considerably higher in the 35S:CsDOF10 transgenic plants compared to the controls (Fig. 5d), while the electrical conductivity in the transgenic lines was significantly lower than that in the controls (Fig. 5e), indicating that the 35S:CsDOF10 transgenic Arabidopsis exhibits enhanced drought resistance compared to the control.
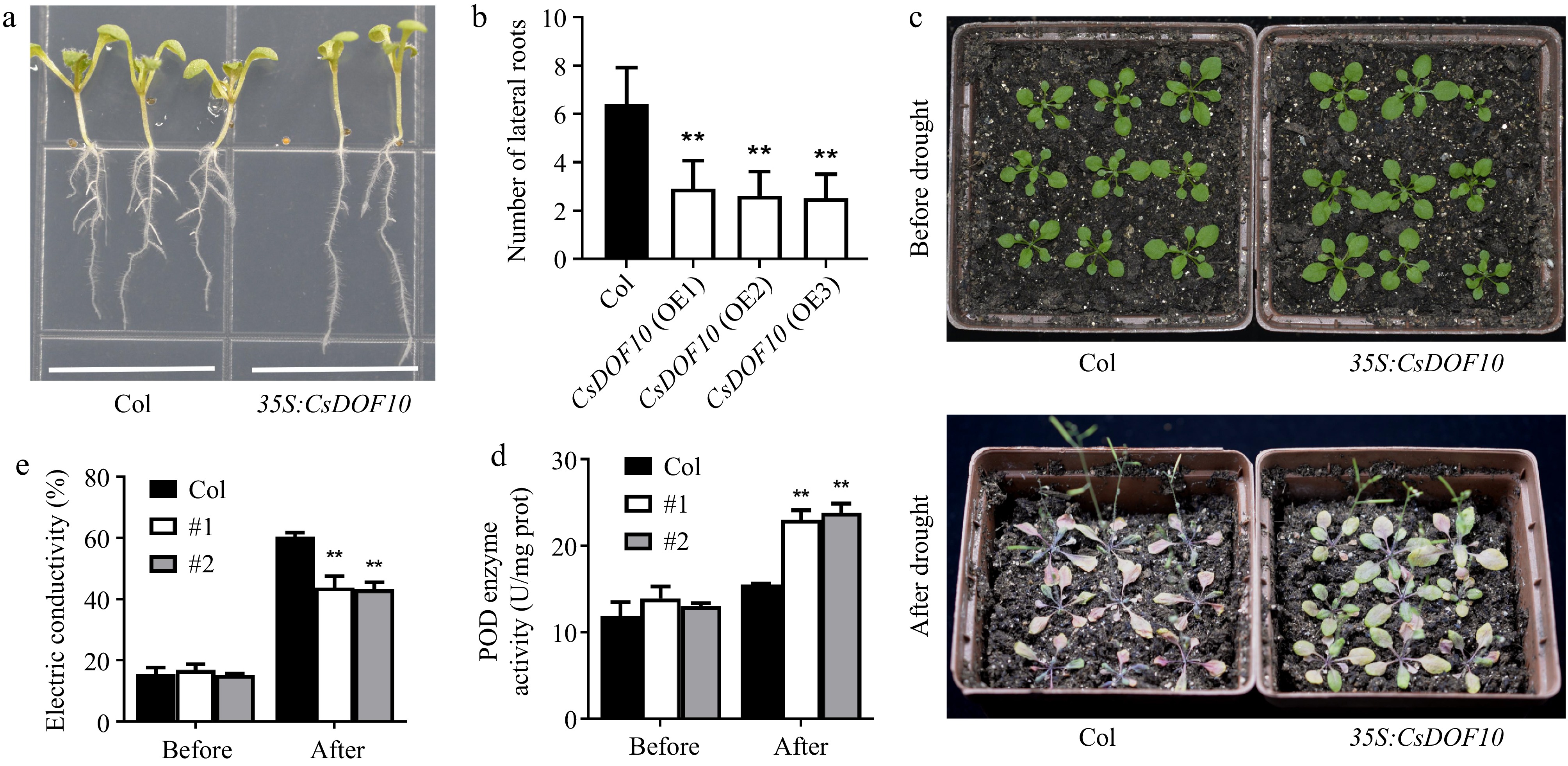
Figure 5.
CsDOF10 transgenic seedlings exhibit strong resistance to drought stress. (a) Root developmental phenotypes of CsDOF10 transgenic Arabidopsis compared to the control. (b) The quantity of lateral roots in CsDOF10 transgenic Arabidopsis vs the control. (c) Phenotypes of CsDOF10 transgenic Arabidopsis and control plants before and after drought treatment. (d) Variations in POD activity, and (e) electrical conductivity of CsDOF10 transgenic seedlings and controls before and after drought treatment. ** represent significant difference at p < 0.01, Students t-test (n = 10).
CsDOF10 is located in the nucleus and has transcriptional activation activity
-
The various software programs predicted that all 24 CsDOF10 proteins were situated in the nucleus (Supplementary Table S2). To examine the expression of CsDOF protein, we employed N. benthamiana leaves to analyze the cellular localization of CsDOF10. The findings revealed that the CsDOF10-GFP fusion protein exhibited intense green fluorescent signals within the nucleus (Fig. 6a), thereby confirming the nuclear localization of this protein. These observations align with the characteristics of TFs and the predictions made by the software.
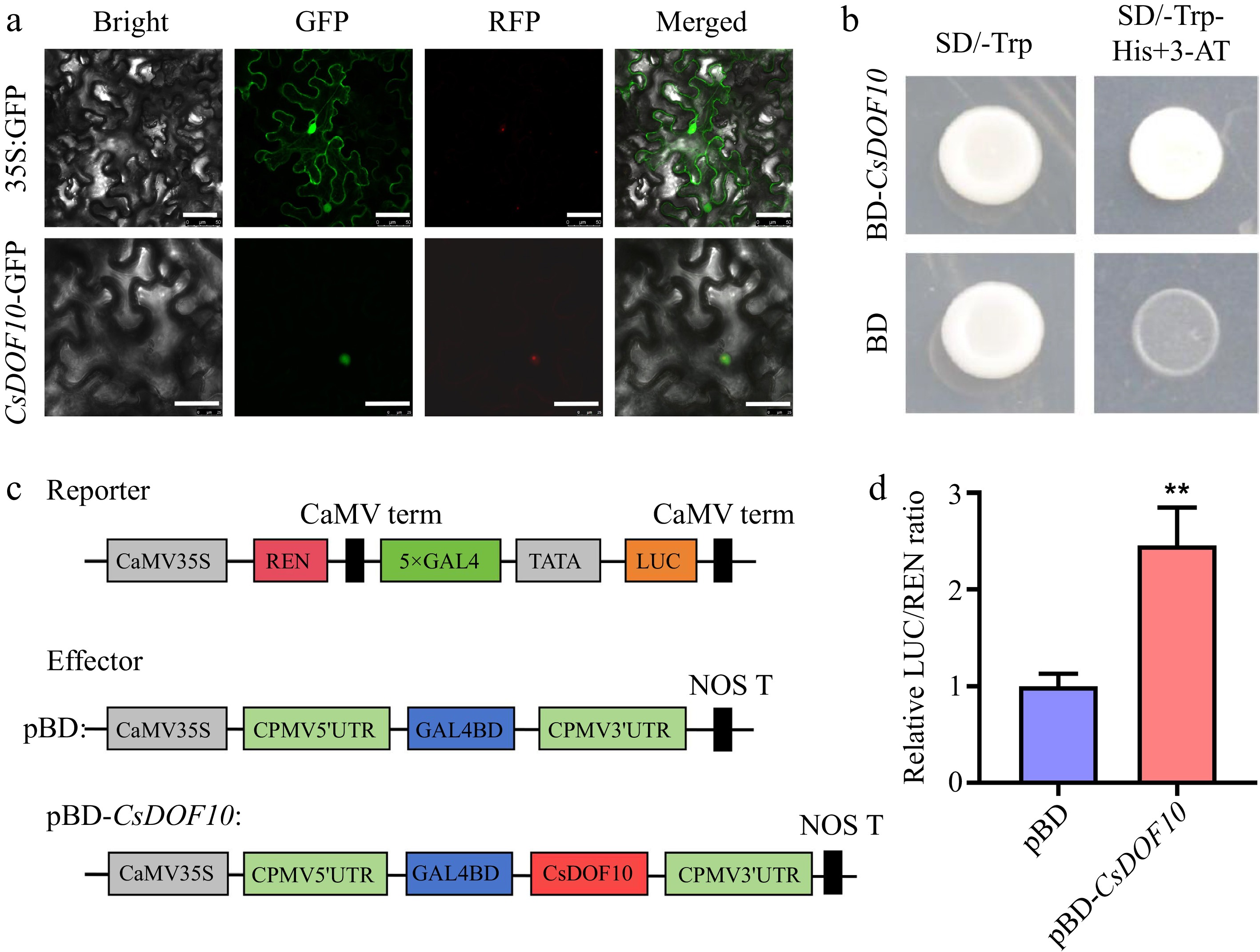
Figure 6.
Subcellular localization and transcriptional activity of CsDOF10. (a) Instantaneous transformation of tobacco leaves displayed that CsDOF10 is located in the nucleus of the cell. Red color represents the fluorescence of nuclear marker VirD2NLS-mCherry. Bars = 50 μm. (b) Yeast experiments have shown that CsDOF10 has transcriptional activation activity. (c) LUC experiments showed that CsDOF10 has transcriptional activation activity. ** represent significant difference at p < 0.01, Students t-test (n = 3).
To investigate the transcriptional activity of CsDOF10, the complete CDS of CsDOF10 was cloned into the pGBKT7 vector (BD), which contains the GAL4 DNA-binding domain, and subsequently introduced into the AH109 yeast (Fig. 6b). All yeast cells exhibited normal growth on the SD/-Trp medium. However, only those yeast cells containing the pGBKT7-CsDOF10 (BD-CsDOF10) construct thrived on the selective SD/-Trp/-His medium supplemented with 3-AT (Fig. 6b). In contrast, yeast cells transformed with the empty pGBKT7 vector failed to survive on the selective medium. To further verify if CsDOF10 acts as a transcriptional activator, a LUC reporter assay was conducted in N. benthamiana (Fig. 6c). The results indicated that CsDOF10 significantly enhanced LUC activity in comparison to the control, with the relative LUC/REN ratio of pBD-CsDOF10 being markedly higher than that of the control pBD (Fig. 6c), suggesting that CsDOF10 TF primarily operates as a transcriptional activator.
Spatiotemporal expression pattern analysis of CsDOF10
-
To investigate the expression pattern of the CsDOF10 gene in various organs of sweet orange, we measured its expression levels in the roots, stems, leaves, flowers, and fruits. The results indicated that the CsDOF10 was expressed in all examined organs, with notably higher expression levels in the flowers (Fig. 7a). To further explore its expression pattern, we cloned the promoter of the CsDOF10 and ligated it to the GUS reporter gene, subsequently transforming it into Arabidopsis plants. GUS histochemical staining demonstrated that the expression of the CsDOF10 promoter was observed in all plant tissues, particularly in flowers, with a notably intense degree of staining (Fig. 7b). The quantification of GUS is conducted to assess the intensity of staining, yielding results that are consistent with those obtained from GUS staining (Fig. 7c).
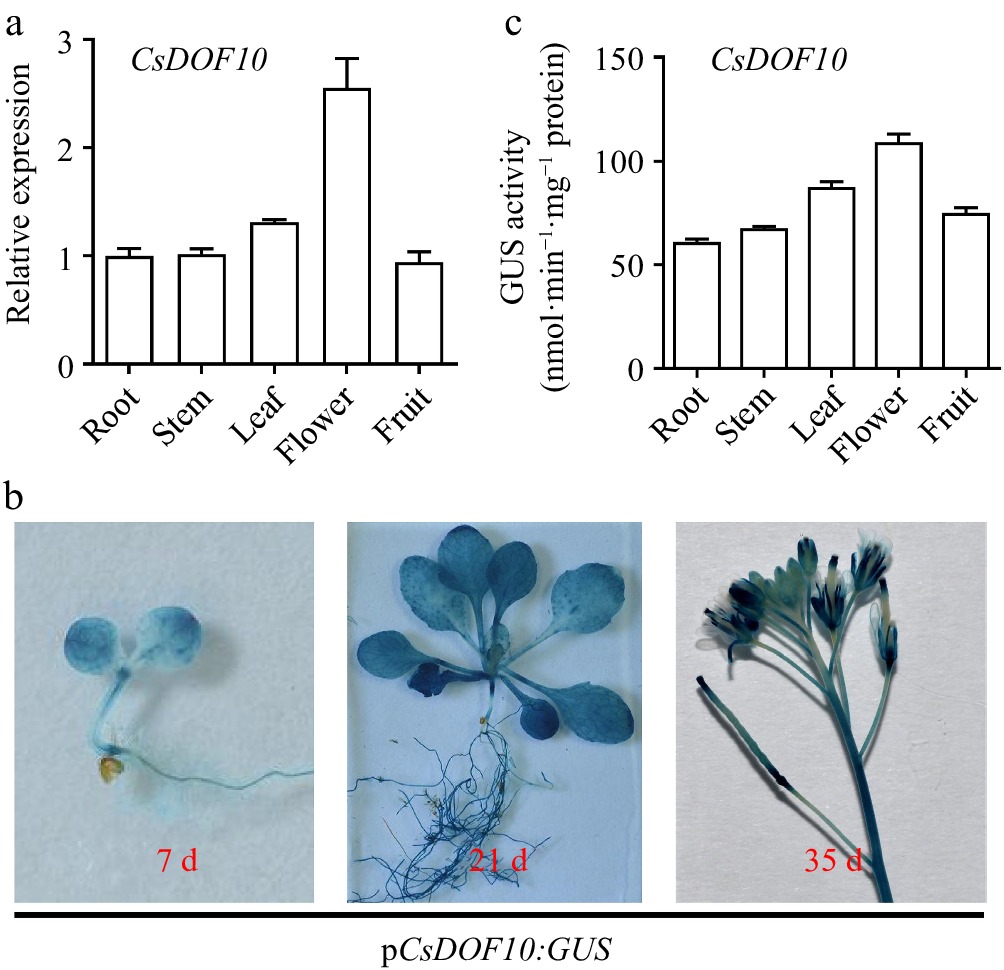
Figure 7.
Expression of CsDOF10 in various tissues and analysis of CsDOF10 promoter cis-elements. (a) The expression pattern of CsDOF10 in different tissues of sweet orange. (b) GUS staining analysis of pCsDOF10:GUS transgenic Arabidopsis plants. (c) Quantitative detection of GUS in different tissues of pCsDOF10:GUS transgenic seedlings.
CsKNOX9 TF inhibits its expression by binding to the promoter of CsDOF10
-
To investigate the upstream regulatory pathway of CsDOF10, we used the CsDOF10 promoter as bait to screen yeast one-hybrid libraries derived from citrus. In total, six genes were identified, of which only one is a TF protein. Annotation reveals that this protein is a KNOTTED1-LIKE HOMEOBOX GENE (KNOX) TF (Supplementary Table S3). Based on previous research findings, we designated this KNOX TF as CsKNOX9[37]. Research has demonstrated that KNOX TFs are capable of specifically identifying and binding to the DNA cis-element known as the TGAC core (TGACTGAC or TGACAGG/CT)[48,49]. We examined the promoter cis-elements of CsDOF10 and identified three binding sites located at −0.391, −2.041, and −2.932 kb from the transcription start site (TSS) (Fig. 8a). These findings indicate that CsKNOX9 may directly interact with the CsDOF10 promoter.
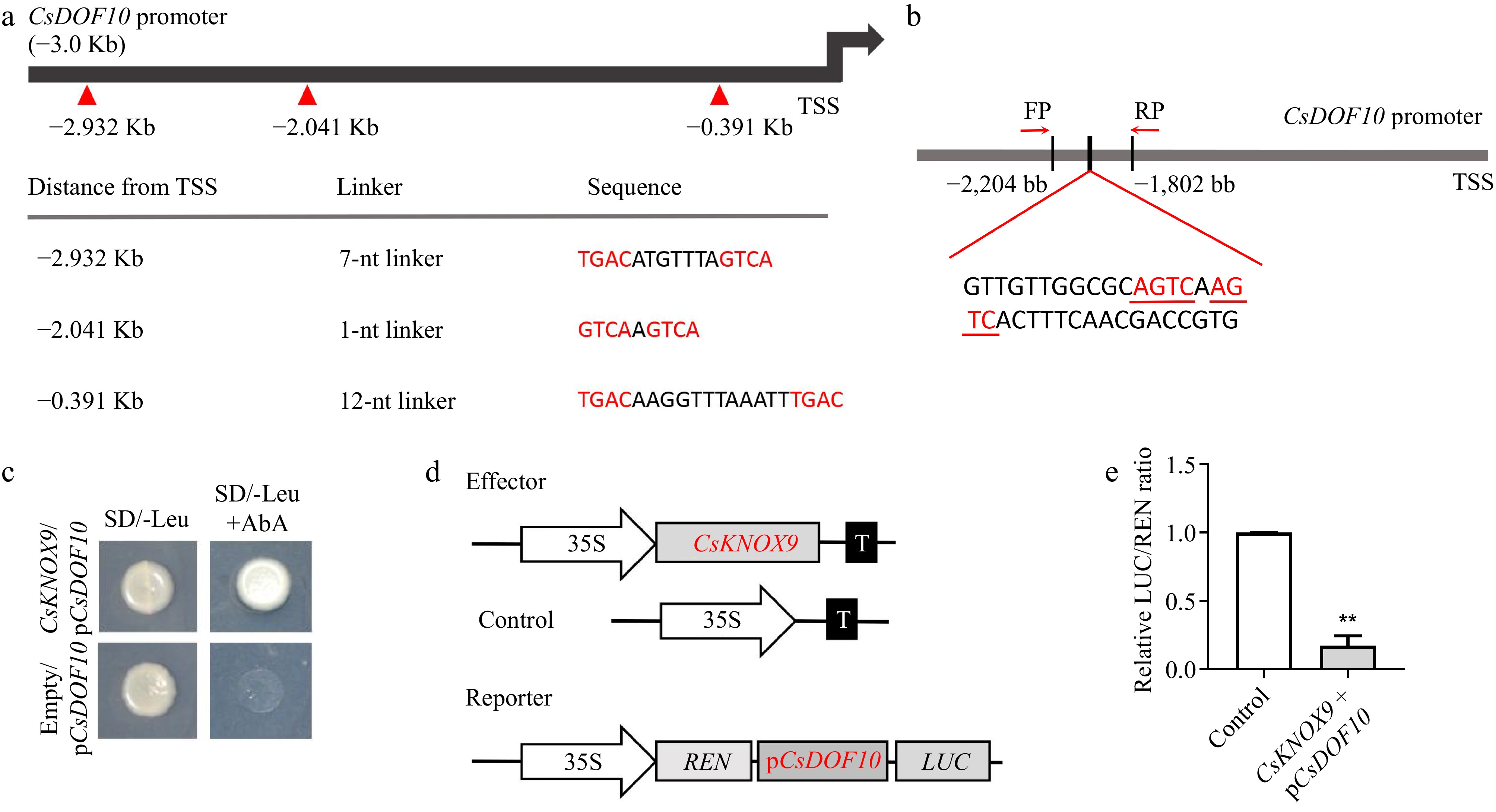
Figure 8.
CsKNOX9 binds to the CsDOF10 promoter and inhibits its expression. (a) Analysis of CsKNOX9 TF binding elements on the CsDOF10 promoter. (b) Schematic diagram illustrating the CsKNOX9 binding element on the CsDOF10 promoter. (c) Yeast one-hybrid assays demonstrate that CsKNOX9 binds to the CsDOF10 promoter. (d) A schematic diagram illustrates the effector and reporter used in the LUC assays. (e) LUC assay results indicate that CsKNOX9 inhibits the expression of CsDOF10. ** represent significant difference at p < 0.01, Students t-test (n = 3).
To verify the interaction of CsKNOX9 TF with the promoter of CsDOF10, we selected a 422 bp fragment containing the GTCAAGTCA site of the CsDOF10 promoter and conducted yeast one-hybrid experiments with CsKNOX9 (Fig. 8b). The results indicated that in the selective medium SD/-Leu, both the experimental and control groups were able to grow normally. However, upon the addition of AbA, the experimental group continued to grow normally, whereas the control group did not, thereby demonstrating that CsKNOX9 interacts with the CsDOF10 promoter (Fig. 8c). To further investigate the transcriptional regulation of CsDOF10 by CsKNOX9, a LUC assay was conducted using a transient expression system in tobacco. The promoter fragment of CsDOF10, which includes the TGAC core, was cloned into the pGreenII 0800 vector, while the CDS for CsKNOX9 was inserted into the pGreenII 62-SK vector (Fig. 8d). The results indicated that the LUC activity was significantly lower when CsKNOX9 was co-transformed with pCsDOF10 compared to the negative control, which involved co-transforming the empty vector pGreenII 62-SK with CsKNOX9, indicating that CsKNOX9 inhibits the expression of CsDOF10 (Fig. 8e).
-
The DOF TF family plays a crucial role in various essential processes in higher plants, with DOF genes uniquely distributed across the plant kingdom. In this research, we performed an extensive computational analysis that led to the identification of a large CsDOF gene family in citrus sweet orange. We provide a detailed overview of this gene family in sweet orange, covering aspects such as gene structures, phylogenetic relationships, chromosomal (Scaffold) locations, and analysis of their cis-regulatory elements on promoters. A total of 24 CsDOF genes were discovered within the sweet orange genome. Our examination of structures indicated that their Dof domains are conserved in the similarity of their sequences. Analysis of protein structures indicates that CsDOFs contain a characteristic conserved region known as the DOF domain, which features a ZF configuration. This structure is vital for their ability to regulate downstream target genes[9,10]. The arrangement of four cysteine residues within the ZF is essential for preserving the stability of the loops, while the tryptophan found in the C-terminal part of the ZF protein is crucial for its ability to bind DNA. In the case of steroid hormone receptors, this specific tryptophan residue appears to be essential for maintaining structural integrity[16]. Furthermore, the phylogenetic comparison of CsDOF proteins with those of Arabidopsis revealed the existence of six major groups.
The characteristics of genes, encompassing their structure, expression patterns, and evolutionary background, are intricately connected to their functions[23,50,51]. This indicates that variations in these attributes may mirror the diversity of gene functions. In terms of structure, the 24 identified CsDOFs exhibited marked differences in several parameters, including protein length, predicted isoelectric point, amino acid count, and molecular weight. Although the exon count remained fairly stable, there was notable variability in exon length. Additionally, both the quantity and composition of conserved motifs showed distinct differences. Furthermore, the cis-acting elements present in the 24 CsDOFs varied significantly in terms of their number, type, and composition. These considerable structural variations are likely to influence functional differences. Our analysis of expression profiles under drought conditions revealed that specific CsDOFs, such as CsDOF1, CsDOF14, CsDOF15, and CsDOF22, exhibited low expression levels after drought treatment, whereas others, including CsDOF2, CsDOF4, CsDOF5, CsDOF7, CsDOF10, CsDOF13, CsDOF17, CsDOF18, CsDOF21, CsDOF23, and CsDOF24, demonstrated high expression levels. It is noteworthy that the expression level of CsDOF10 significantly increases following drought treatment. Consequently, this study further investigates the function of CsDOF10.
Further study indicated that the overexpression of CsDOF10 in Arabidopsis enhances plant’s resistance to drought conditions. The role of DOF TF in drought stress has been reported in studies involving other species. For example, an analysis of cis-elements revealed the potential roles of DOFs in potato development, particularly under multiple abiotic stress conditions, with a specific emphasis on their regulatory functions in drought stress[52]. A study in apple revealed that plants overexpressing MdDof54 exhibited higher survival rates under short-term drought conditions, while those with MdDof54 RNAi showed reduced survival rates[32]. This suggests that the DOF TF plays a crucial regulatory role in the apple's response to drought stress. In addition, studies on tea[53], and birch (Betula platyphylla)[14] also suggest that DOF TFs may play a potential regulatory role in the response to drought. This study also found that the transcriptional expression of CsDOF10 is regulated by the upstream TF CsKNOX9, which belongs to the homeodomain TF family and plays multiple regulatory roles in citrus research[41,45]. A study has also demonstrated that the KNOX TF member STM can regulate drought responses in Arabidopsis, and increased expression of STM is associated with enhanced drought tolerance in Arabidopsis plants[54]. Furthermore, studies have indicated that the homeodomain TF BEL1-like protein (StBEL5) modulates the expression of CYCLING DOF FACTOR1 (StCDF1) in potatoes via tandem TGAC core motifs. These findings imply that CsKNOX9 may influence the drought response mechanisms in citrus by affecting CsDOF10 expression. Nonetheless, the role of CsKNOX9 in this context requires additional investigation. In addition, this study demonstrated that CsDOF10 TF possess transcriptional activation activity, highlighting its regulatory role on downstream target genes as a significant work for future research. Our next step will conduct the genetic transformation of CsDOF10 in citrus, utilizing ChIP-seq and RNA-seq to further explore its downstream target genes.
Root growth and hydraulic conductivity of roots are crucial factors influencing a plant's ability to withstand drought[55,56]. Previous research demonstrated that, under prolonged drought conditions, MdDof54 RNAi plants exhibited reduced root dry weight and decreased root hydraulic conductivity compared to the control GL-3 plants. This suggests that MdDof54 not only contributes to rice's drought tolerance but also plays a beneficial role in the management of root development[32]. In this study, we also observed that CsDOF10 plays a role in root development. Compared to the control, CsDOF10 transgenic Arabidopsis exhibited fewer lateral roots and longer main roots. Therefore, we hypothesize that the regulatory role of CsDOF10 in citrus drought stress is likely closely linked to its capacity to influence root development, however, the underlying mechanisms of this association require further investigation. In addition, it has been reported that the DOF TF OsDOF15 enhances primary root growth in rice by stimulating cell proliferation in the root meristem[57]. In Nicotiana tabacum, the DOF protein NtBBF1 interacts with the rolB promoter in response to auxin, thereby influencing root development[19]. These findings suggest that DOF TF play a significant role in the growth of plant roots. Studies have reported that DOF TFs also play a regulatory role in the germination of plant seeds[3,58,59]. However, we did not find any significant differences in the germination rates of CsDOF10 transgenic seedlings compared to the control in the present study.
Taken together, in this study, we identified 24 candidate CsDOFs distributed across 19 scaffolds of the sweet orange genome. These 24 CsDOFs were classified into six distinct groups. Notably, many CsDOFs exhibited varying expression patterns in response to drought stress, with CsDOF10 showing the most pronounced changes. The overexpression of CsDOF10 in Arabidopsis enhances plant resistance to drought, stimulates the growth of primary roots, and restricts the formation of lateral roots. Additionally, further research has revealed that the expression of CsDOF10 is regulated by the upstream regulatory factor CsKNOX9. These findings provide valuable insights for future research on the adaptation of citrus sweet orange to drought stress.
This work was supported by the National Natural Science Foundation of China (Grant No. 32202408), the Project of the Natural Science Foundation of Guangdong Province, China (2025A1515012694), the Construction Plan of Guangdong Province High-level Universities and the Research Start-up Funds for the High-level Talent Introduction Project of South China Agricultural University (5300-223109), the Modern Agriculture Industry Technology System Innovation Team Construction Project of Guangdong Province (Optimal and Rare Fruit Industry Technology System) (2024CXTD09), and Guangzhou Agricultural Support Fund, 2025 (2503040961).
-
The authors confirm contribution to the paper as follows: conducting the experimental work and data analysis: Hu SF, Zhong ZQ, Qi Y, Sun YH; experiments designing and overall study supervision: Zeng RF; writing manuscript: Hu SF; manuscript revision: Zeng RF; all authors engaged in discussions regarding the collected data, reviewed and provided feedback on the manuscript. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 The primers used in this study.
- Supplementary Table S2 Gene names, genomic information, and predicted protein parameters for the 24 putative CsDOF genes in sweet orange genome.
- Supplementary Table S3 Genes identified through screening of Y1H library.
- Supplementary Fig. S1 Gene structures of CsDOFs in sweet orange. Exons, introns and UTR are shown.
- Supplementary Fig. S2 Conserved motifs analysis of CsDOFs. (a) Conserved motifs of CsDOF proteins identified with the MEME search tool. Motifs are represented by boxes. The numbers (1−10) and different colors in boxes represent motif 1−motif 10, respectively. Box size indicates the length of motifs. (b) Sequence logo of conserved motif of CsDOF proteins in sweet orange.
- Supplementary Fig. S3 Multiple alignments of the Dof domains of the CsDOFs using the ClustalW2 program, and alignments were illustrated in the "ClutalX" color mode.
- Supplementary Fig. S4 Cis-elements in promoter regions of 24 CsDOFs. Elements were identified in the 2,000 bp sequences upstream of the start codon of CsDOFs genes using PlantCARE database. The elements associated with specific functions are indicated by different colors for each gene.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Hu SF, Zhong ZQ, Qi Y, Sun YH, Zeng RF. 2025. Identification of DNA-binding with one finger (DOF) transcription factor family and functional study of CsDOF10 in sweet orange (Citrus sinensis). Fruit Research 5: e019 doi: 10.48130/frures-0025-0011 

Identification of DNA-binding with one finger (DOF) transcription factor family and functional study of CsDOF10 in sweet orange (Citrus sinensis)
- Received: 15 January 2025
- Revised: 10 March 2025
- Accepted: 18 March 2025
- Published online: 13 May 2025
Abstract: DOF transcription factors (TFs), which are specific to plants and contain zinc finger motifs, play a crucial role in various physiological and biochemical processes throughout plant life. Citrus sweet orange, one of the most economically and aesthetically valuable fruit crops globally, has not been the subject of thorough systematic analyses regarding its CsDOF gene members and their functions. In this study, we identified 24 CsDOF genes that are distributed unevenly across 19 scaffolds in sweet orange. We provide a detailed overview of these CsDOF genes, including their scaffold locations, phylogenetic relationships with the Arabidopsis DOF family, and structural characteristics. The phylogenetic analysis grouped the 60 DOF members identified from citrus and Arabidopsis into six main groups. Additionally, we detected distinct expression patterns of CsDOF genes under both normal and drought conditions using qRT-qPCR. Notably, the CsDOF10 gene showed high expression level in response to drought treatment, suggesting a potential adaptation to drought stress. The overexpression of CsDOF10 in Arabidopsis increases the plant's resistance to drought, stimulates the growth of primary roots, and suppresses the formation of lateral roots. CsDOF10 is located in the nucleus and has transcriptional activation activity. Moreover, additional studies have demonstrated that the expression of CsDOF10 is influenced by the upstream regulatory factor CsKNOX9. Our discoveries offer significant insights for future investigations into the ability of citrus sweet orange to cope with drought stress.