








-
The Lagerstroemia genus, in the Lythraceae family and colloquially referred to as crape myrtle, is esteemed for its large, visually striking flowers[1,2]. A substantial number of breeding programs have been implemented, focusing on diversifying and improving floral forms, colors, leaf pigmentation, plant habits, and disease resistance. Over 20 Lagerstroemia cultivars, displaying significant ornamental characteristics and powdery mildew resistance were developed through the hybridization of L. indica with L. fauriei[3−5]. These cultivars are distinguished by their large flowers and compact, dwarf growth habits. Knight & Mclaurin bred cultivars featuring dark foliage, followed by Pounders et al. who generated a range of cultivars with dark purple leaves through hybridization[6,7]. Pooler successfully bred two cultivars displaying vibrant red flowers through crosses involving L. indica, L. fauriei, and L. limii[8].
To date, 55 species in the genus Lagerstroemia and at least 500 named crape myrtle cultivars are available across the US, Europe, and Asia[9]. Flower color serves as a critical ornamental characteristic of crape myrtle. Naturally, Lagerstroemia species exhibit flower colors such as purple, red, pink, and white, yet lack yellow-flowered cultivars[10]. Researchers have introduced new traits in Lagerstroemia by hybridization, such as purple leaves, vibrant red flowers, and a compact, dwarf growth habit[3−5,8,11]. Despite efforts, breeding Lagerstroemia cultivars with yellow flowers remains an unrealized goal. Heimia species with yellow flowers range naturally from Mexico to Argentina and are valued for their medicinal significance, belong to the Lythraceae family, and are closely allied with Lagerstroemia[12,13]. There exists potential for breeding novel yellow-flowered Lagerstroemia cultivars through distant hybridization with Heimia.
Distant hybridization has the potential to overcome reproductive barriers, facilitating the transfer and recombination of superior traits that have evolved over long periods, thereby acquiring elite characteristics inaccessible through close hybridization. This approach aims to achieve targeted plant improvement and develop novel plant types with distinct specificity and unprecedented traits[14,15]. Currently, intergeneric hybridization has been effectively utilized in plant breeding programs of Brassicaceae[16,17], Ericaceae[18], and Sapindaceae[19]. In the late 19th century, French breeders hybridized Rosa 'Antoine Ducher' with R. foetida 'Persian Yellow', resulting in yellow-flowered R. 'Soleil d'Or', which is regarded as the progenitor of all known yellow and orange modern rose cultivars[20]. Using R. rugosa 'Hunchun' with pink flowers as the female and R. xanthina with yellow flowers as the male parent, six progeny lines with a range of dark to light yellow petals were obtained[21]. Furthermore, a cross between Camellia nitidissima with single petals and yellow flowers and C. japonica with double petals and pink flowers resulted in offspring with double petals and light-yellow flowers[22].
Distant hybridization is challenging due to differences in chromosome homology and complex genetic backgrounds among species[23,24]. Reproductive isolation is categorized into two primary types: pre-fertilization and post-fertilization barriers[25]. The reasonable selection of hybrid parents, utilization of mentor pollen, and repeated pollination, along with the application of plant hormones or chemical reagents to the stigma after pollination, can effectively mitigate pre-fertilization disorders[18,24]. For instance, the application of IAA and GA can significantly enhance embryonic cell growth and development, these hormones facilitate protein synthesis and supply additional energy to the embryo[26]. In grape production, GA3 enhances yields by stimulating cluster growth through cell elongation and division[27]. Additionally, spraying different concentrations of gibberellin, boric acid, and 2,4-D at the flowering stage of Ziziphus jujuba could increase the fruit setting rate to different degrees[28]. Immature embryo rescue is recognized as the most efficacious method for overcoming post-fertilization barriers[29,30], and it is imperative to select embryos with higher maturity for rescue procedures[31].
We aimed to breed Lagerstroemia cultivars with yellow flowers through intergeneric hybridization between Lagerstroemia and Heimia. The occurrence sites of fertilization disorder between the two genera were observed. Fruiting efficiency was improved through pollen guidance and repeated pollination, and the development time of hybrid fruits was prolonged with the application of hormones. Hybrid offspring were obtained through in vitro culture of hybrid embryos. The findings of the present study established a novel foundation for broadening the color spectrum of Lagerstroemia species via distant hybridization.
-
Seven cultivars of L. indica ('Dallas Red', 'Sarah's Favorite', 'Catawba', 'Centennial Spirit', 'Near East', 'Pocomoke', and 'Qianceng Feixue'), L. fauriei, H. myrtifolia, and H. salicifolia were used as hybridizing parents (Fig. 1). All materials were planted in the nursery of the China National Engineering Research Center for Floriculture (CNERCF) (Beijing, China) (40°02' N, 115°50' E).

Figure 1.
Parental materials of Lagerstroemia and Heimia species and cultivars.
Pollen collection and viability determination
-
Pollen collection was performed following the protocol described by Ju et al.[11]. The collected pollen grains were subsequently dried and maintained at 4 °C. Before pollination, Lagerstroemia pollen grains were incubated in a medium containing 150 g·L−1 sucrose, 20 mg·L−1 H3BO3, 20 mg·L−1 CaCl2, and 100 g·L−1 PEG4000[32]. Meanwhile, Heimia pollen were cultivated in a medium containing 500 mg·L−1 boric acid and 15% sucrose[33]. Both incubations were carried out at 25 °C for 4 h. The viability of the pollen grains was subsequently assessed using an optical microscope (EX20, SDPTOP). The mean pollen viability was determined using the formula: Pollen viability (%) = (Number of germinated pollen grains / Total number of pollen grains) × 100%.
Pollination combinations
-
Eight Lagerstroemia species or cultivars were reciprocally hybridized with H. salicifolia and H. myrtifolia respectively, and a total of 26 pollination combinations and three control groups were set (Table 1). Artificial pollinations were carried out according to the method described by Ju et al.[11].
Table 1. Pollination combinations of Lagerstroemia and Heimia.
L. indica 'Dallas Red' × H. myrtifolia H. myrtifolia × L. indica 'Dallas Red' L. indica 'Dallas Red' × H. salicifolia H. salicifolia × L. indica 'Dallas Red' L. indica 'Sarah's Favorite' ×
H. myrtifoliaH. myrtifolia × L. indica 'Sarah's Favorite' L. indica 'Sarah's Favorite' ×
H. salicifoliaH. salicifolia × L. indica 'Sarah's Favorite' L. indica 'Catawba' × H. myrtifolia H. myrtifolia × L. indica 'Catawba' L. indica 'Catawba' × H. salicifolia H. salicifolia × L. indica 'Catawba' L. indica 'Qianceng Feixue' ×
H. myrtifoliaH. myrtifolia × L. indica 'Qianceng Feixue' L. indica 'Qianceng Feixue' ×
H. salicifoliaH. salicifolia × L. indica 'Qianceng Feixue' L. fauriei × H. myrtifolia H. myrtifolia × L. fauriei L. fauriei × H. salicifolia H. salicifolia × L. fauriei L. indica 'Near East' × H. myrtifolia H. myrtifolia × L. indica 'Near East' L. indica 'Pocomoke' × H. myrtifolia H. myrtifolia × L. indica 'Pocomoke' L. indica 'Centennial Spirit' ×
H. myrtifoliaH. myrtifolia × L. indica 'Centennial Spirit' L. indica 'Qianceng Feixue' ×
'Sarah's Favorite'*H. myrtifolia × H. myrtifolia* H. salicifolia × H. salicifolia * Pollen mentor
-
When Heimia species were the female parents, the Heimia pollen was used as mentor pollen, mixed with Lagerstroemia pollen in a 1:1 volume ratio. Conversely, when Lagerstroemia species and cultivars were the female parents, mentor pollen was collected from 'Sarah's Favorite' and 'Qianceng Feixue', and mixed with Heimia pollen in a 1:1:2 volume ratio. The mentor pollen was stored at room temperature for 4 to 5 d, and pollen inactivation was validated using the method described in Pollen collection and viability determination.
Stigma treatment with hormone after pollination
-
To extend the development period of hybrid fruits on the plants, the stigmas were sprayed with different concentrations of GA3 and 2,4-D at different time points after pollination. Specifically, 100 mg·L−1 and 200 mg·L−1 GA3, as well as 20 mg·L−1 and 50 mg·L−1 2,4-D, were applied at 2 h, 1, 2, 4, and 6 d after pollination. The fruiting rate was evaluated according to the method described by Cai et al.[32].
Observation of pollen germination and pollen tube elongation
-
Styles were collected at 2, 4, 6, 8, 12, 24, 48, and 72 h after pollination and fixed in FAA (75% alcohol : acetic acid : 38% formaldehyde = 18:1:1, v:v:v) for at least 24 h at 4 °C. The fixed styles were then pressed and stained according to the method described by Kho & Baër[34], and then observed under a fluorescence microscope (ZEISS Axio Scope TYPE A1). Each treatment involved five styles and was repeated three times.
Observation of hybrid embryo development
-
Hybrid embryos were sampled at 2, 3, 4, 5, 10, 15, 20, 25, 30, 40, 50, and 60 d after pollination and fixed in FAA for at least 24 h at 4 °C. Paraffin sections were prepared following the protocol detailed by Wang et al.[35]. The sections were examined using an optical microscope (EX20, SDPTOP).
Embryo rescuing
-
Hybrid fruits were collected at 10, 15, 20, 25, and 30 d after pollination and rinsed under running water for a minimum of 2 h. The fruits were surface-sterilized on a clean bench through sequential treatments: first with 75% ethanol for 30 s, followed by immersion in 2% sodium hypochlorite solution for 10 min. The conical flask was agitated during sterilization to ensure complete coverage. The fruits were subsequently rinsed with sterile water three to four times. Seeds were removed from the fruits and placed onto the medium, with 10−12 seeds per Petri dish. At each developmental stage, a sample of 5 to 10 fruits was selected, with each treatment repeated three times. The medium included agar (7 g·L−1), sucrose (30 g·L−1), and various hormone ratios (Table 2), with a pH range of 5.8−6.0.
Table 2. Establishment media used in embryo rescuing.
Medium ID Medium Hormone combination (mg·L−1) 6-BA NAA GA3 CK MS 0 0 0 1 MS 0.5 0 1 2 1/2 MS 0.5 0.5 1 3 MS 0.5 0 0.5 4 MS 0.5 0.5 1 5 MS 0.5 0.5 1.5 6 MS 0.5 0 0 7 MS 0.5 0.5 0 Identification of hybrid progenies using simple sequence repeats (SSR) markers
-
A total of 761 SSR markers, developed from the transcriptome of Lagerstroemia species[36], were selected. Leaf samples were obtained from both parental plants and their hybrid progeny. DNA was extracted using the TIANGEN DNAsecure Plant Kit (TIANGEN Biotech, Beijing, China) and used as templates for PCR amplification. SSR marker primers that demonstrated stable amplification in the parents and displayed complementary specific loci for both were selected.
A TP-M13 fluorescent adapter (5'-TGTAAAACGACGGCCAGT-3') was added to the 5' end of the upstream primers. The primers and adapters were synthesized by RiboBio Technology (Beijing) Co., Ltd. PCR amplification was conducted according to the protocol described by Zhang et al.[37]. Data analysis was conducted using Genemarker version 2.2.0 software.
-
The pollen viability of Lagerstroemia species and cultivars, as well as Heimia species, ranged from 55% to 92% (Table 3, Fig. 2), which met the criteria necessary for hybridization.
Table 3. Pollen germination rates of eight Lagerstroemia and two Heimia species and cultivars.
Species and cultivars Germination percentage L. indica 'Dallas Red' 85% L. indica 'Sarah's Favorite' 92% L. indica 'Qianceng Feixue' 85% L. fauriei 55% L. indica 'Catawba' 87% L. indica 'Centennial Spirit' 78% L. indica 'Near East' 75% L. indica 'Pocomoke' 80% H. myrtifolia 55% H. salicifolia 65% 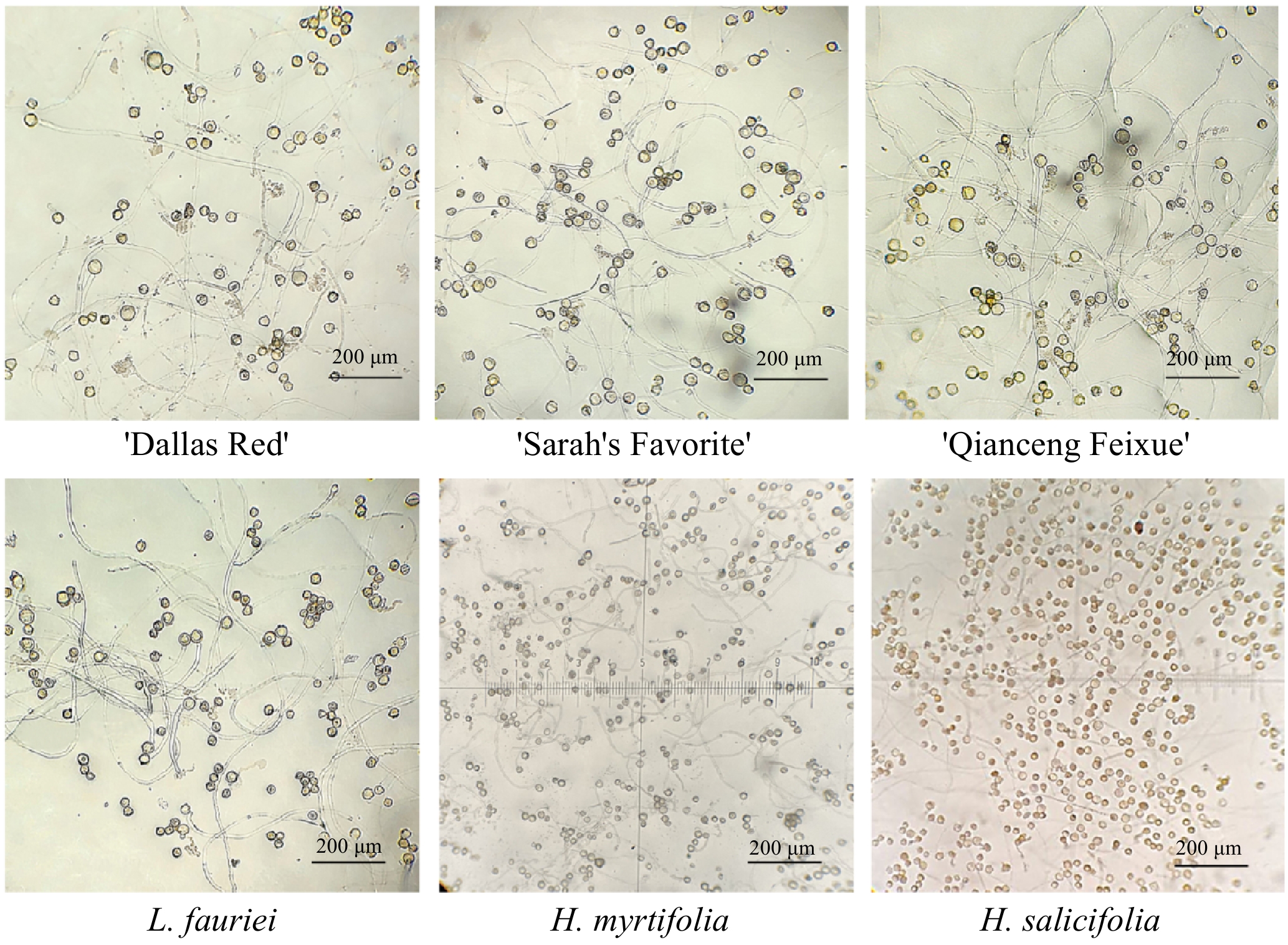
Figure 2.
Pollen viability of Lagerstroemia and Heimia species and cultivars.
Fruiting rate of distant hybridization
-
The fruiting rate of L. indica 'Qianceng Feixue' × L. indica 'Sarah's Favorite' reached 79.8%, while those of H. myrtifolia × H. myrtifolia and H. salicifolia × H. salicifolia were 74.1% and 81.3%, respectively. When Lagerstroemia species and cultivars were the maternal parents, several fruits fell off within 3−5 d after pollination, with complete fruit shedding occurring within 5 d. Conversely, when Heimia species were the maternal parents, no fruit abscission was observed, however, all fruits wilted within 10 to 15 d after pollination. After applying 20 mg·L−1 2,4-D on the stigma surfaces of pollinated flowers, fruits were successfully obtained in six pollination combinations: 'Sarah's Favorite' × H. myrtifolia, 'Sarah's Favorite' × H. salicifolia, H. salicifolia × 'Dallas Red', H. salicifolia × 'Sarah's Favorite', H. myrtifolia × 'Sarah's Favorite', and H. myrtifolia × L. fauriei. The fruit set percentage crosses ranged from 0.7% to 6.0%. All other pollination combinations failed to produce fruit (Table 4), highlighting substantial reproductive barriers between Lagerstroemia and Heimia.
Table 4. Fruiting rates of pollination combinations between Lagerstroemia and Heimia after spraying 20 mg·L−1 2,4-D.
Female parents (♀) Male parents (♂) No. of pollinated flowers Fruiting
rate (%)L. indica 'Dallas Red' H. myrtifolia 282 0.7 H. salicifolia 275 0.0 L. indica 'Sarah's Favorite' H. myrtifolia 545 0.0 H. salicifolia 330 1.1 L. indica 'Catawba' H. myrtifolia 224 0.0 H. salicifolia 268 0.0 L. indica 'Qiancengfeixue' H. myrtifolia 850 0.0 H. salicifolia 509 0.0 L. fauriei H. myrtifolia 200 0.0 H. salicifolia 350 0.0 L. indica 'Centennial Spirit' H. myrtifolia 126 0.0 L. indica 'Near East' 199 0.0 L. indica 'Pocomoke' 215 0.0 H. salicifolia L. indica 'Dallas Red' 284 5.4 L. indica 'Sarah's Favorite' 361 4.1 L. indica 'Catawba' 98 0.0 L. indica 'Qiancengfeixue' 127 0.0 L. fauriei 144 0.0 H. myrtifolia L. indica 'Dallas Red' 292 0.0 L. indica 'Sarah's Favorite' 1,650 6.0 L. indica 'Catawba' 326 0.0 L. indica 'Qiancengfeixue' 63 0.0 L. indica 'Centennial Spirit' 306 0.0 L. indica 'Near East' 242 0.0 L. indica 'Pocomoke' 308 0.0 L. fauriei 1,489 3.7 *L. indica 'Qiancengfeixue' L. indica 'Sarah's Favorite' 193 79.8 *H. myrtifolia H. myrtifolia 85 74.1 *H. salicifolia H. salicifolia 75 81.3 * indicates the control group. In seven pollination combinations where Lagerstroemia served as the female parent (Table 5), we applied a range of GA3 and 2,4-D concentrations to the stigmas after mentor pollination. The fruit set rates after the application of 2,4-D were higher than those treated with GA3 within 3 to 5 d after pollination. However, fruits treated with 2,4-D abscised rapidly in 5 to 10 d after pollination, and all hybrid fruits dropped within 20 to 25 d. The fruit set rates following GA3 treatment decreased gradually in the first 10 d after pollination, with slight changes after 25 d. Application of 200 mg·L−1 GA3 yielded higher fruit set rates compared to 100 mg·L−1 GA3 (Fig. 3). The seeds from distant crosses were predominantly either non-plump or devoid of endosperm (Fig. 4a).
Table 5. Fruiting rates of pollination combinations between Lagerstroemia and Heimia following treatments with varying concentrations of 2,4-D and GA3.
Female
parents
(♀)Male
parents
(♂)Hormone Concentration (mg·L−1) No. of pollinated flowers Fruiting rate (%) L. fauriei H. salicifolia GA3 100 126 11.9 200 193 16.1 2,4-D 20 350 0 50 290 0 L. indica
'Sarah's
Favorite'H. salicifolia GA3 100 230 3.5 200 222 6.3 2,4-D 20 330 1.1 50 173 1.2 L. indica
'Qianceng Feixue'H.myrtifolia GA3 100 93 8.6 200 83 14.5 2,4-D 20 850 0 50 163 0 H. salicifolia L. indica
'Dallas Red'GA3 100 107 0.9 200 170 1.2 2,4-D 20 284 5.4 50 43 7 L. indica
'Sarah's
Favorite'GA3 100 119 0 200 27 0 2,4-D 20 361 4.1 50 90 8.9 L. indica
'Qianceng Feixue'GA3 100 52 0 200 53 0 2,4-D 20 127 0 50 43 0 L. fauriei GA3 100 79 0 200 71 0 2,4-D 20 144 0 50 40 0 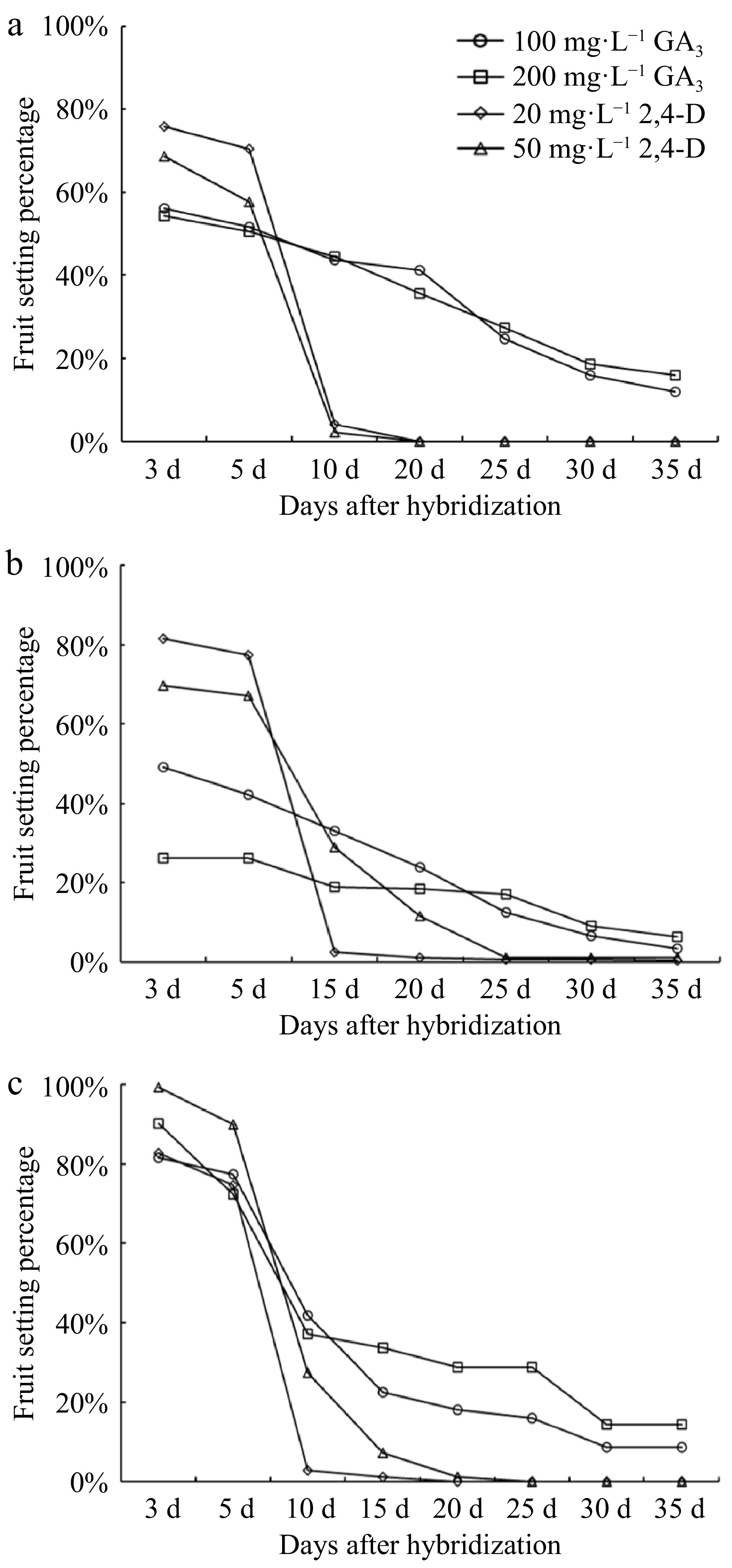
Figure 3.
Fruiting rate of three pollination combinations with Lagerstroemia as female parent treated with GA3 and 2,4-D. (a) L. fauriei × H. salicifolia; (b) 'Sarah's Favorit' × H. salicifolia; (c) 'Qianceng Feixue' × H. myrtifolia.

Figure 4.
Fruit morphology after distant hybridization. (a) Fruit and seeds of 'Qianceng Feixue' × H. myrtifolia (left) and 'Qianceng Feixue' × 'Sarah's Favorite' (right) 35 d after pollination. (b) Hybrid fruits of H. salicifolia × 'Qianceng Feixue' (left), and H. salicifolia × H. salicifolia (right) 20 d after pollination.
In pollination combinations with Heimia was the female parent, the fruits turned brown and withered approximately 15 to 20 d after pollination following treatment with GA3. In contrast, fruits treated with 2,4-D did not desiccate until 20 to 25 d after pollination (Fig. 4b), and the fruit retention rate treated with 50 mg·L−1 2,4-D was higher. In summary, when Lagerstroemia and Heimia serve as the maternal parents, treatments with GA3 and 2,4-D can extend the developmental period of hybrid fruits respectively. When Heimia was employed as the maternal parent, 2,4-D treatment yields more fruits. Conversely, GA3 treatment was more effective when Lagerstroemia served as the maternal parent.
Pollen tube behavior during distant hybridization
-
Mentor pollen was found to facilitate the elongation of pollen tubes (Fig. 5). No matter whether the female parent was Lagerstroemia or Heimia, pollen grains could germinate on the stigma surface within 1 to 2 h after pollination. However, callose accumulation on the stigma surface or during pollen tube elongation impeded further elongation of the pollen tube.

Figure 5.
The behavior of pollen tubes in different cross combinations. (a) Pollen tubes elongated downward 12 h after pollination in 'Sarah's Favorite' × H. salicifolia. (b) Few pollen tubes extend into the ovary 48 h after pollination in 'Sarah's Favorite' × H. salicifolia. (c) Pollen tubes were mostly confined to about 1/4 of the stigma in L. fauriei × H. salicifolia. (d) Few pollen tubes reached the base of pistil in L. fauriei × H. salicifolia. (e) Pollen tubes stretched 2 h after pollination in H. salicifolia × 'Dallas Red'. (f) A few pollen tubes entered the ovary 48 h after pollination in H. salicifolia × L. fauriei. (g) The main obstacle occurred at the base of the pistil 72 h after pollination in H. myrtifolia × L. fauriei. (h) Callose were produced in the style 12 h after pollination in 'Sarah's Favorite' × H. salicifolia. (i) A few pollens germinated 4 h after pollination in H. salicifolia × 'Sarah's Favorite' without mentor pollens. (j) Pollen failed to germinate in 4 h after pollination in 'Sarah's Favorite' × H. salicifolia without mentor pollens. Pt: Pollen tube. Pg: Pollen grain. Ca: Callose. Ou: Ovule.
When Lagerstroemia species served as the female parent, pollen tubes elongated downward in clusters between 4 to 12 h after pollination (Fig. 5a), and most pollen tubes reached one-third to one-fourth of the length of the stigma within 24 to 48 h after pollination (Fig. 5c, d), a few pollen tubes continued to elongate and entered the ovary between 48 to 72 h (Fig. 5b). When Heimia species served as the female parent, pollen grains germinated on the stigma surface within 1 to 2 h after pollination (Fig. 5e) and reached the base of the stigma within 24 h. However, most pollen tubes failed to penetrate the base of the stigma (Fig. 5g), and only a few entered the ovaries within 72 h (Fig. 5f). Pollen tubes were observed to extend into the ovary in only eight cross combinations including 'Sarah's Favorite' × H. salicifolia, 'Dallas Red' × H. myrtifolia, 'Sarah's Favorite' × H. myrtifolia, H. myrtifolia × 'Dallas Red', H. myrtifolia × 'Sarah's Favorite', H. myrtifolia × L. fauriei, H. salicifolia × 'Dallas Red', and H. salicifolia × 'Sarah's Favorite'.
Development of hybrid embryos between Lagerstroemia and Heimia
-
Embryo development was observed in only two cross combinations (H. salicifolia × 'Sarah's Favorite' and H. myrtifolia × L. fauriei). The seeds of self-pollinated Heimia fruits developed into heart-shaped embryos within 15 d after pollination, and from heart-shaped embryos to torpedo embryos between 15 to 30 d (Fig. 6a). The seeds of 'Qianceng Feixue' × 'Sarah's Favorite' formed heart-shaped embryos approximately 25 d after pollination (Fig. 6b). Only two intergeneric crosses (H. salicifolia × 'Sarah's Favorite', H. myrtifolia × L. fauriei) were observed to develop into heart-shaped embryos, while the embryos of other cross combinations stagnated at the endosperm development stage. The results showed that there was a post-fertilization disorder between Lagerstroemia and Heimia.
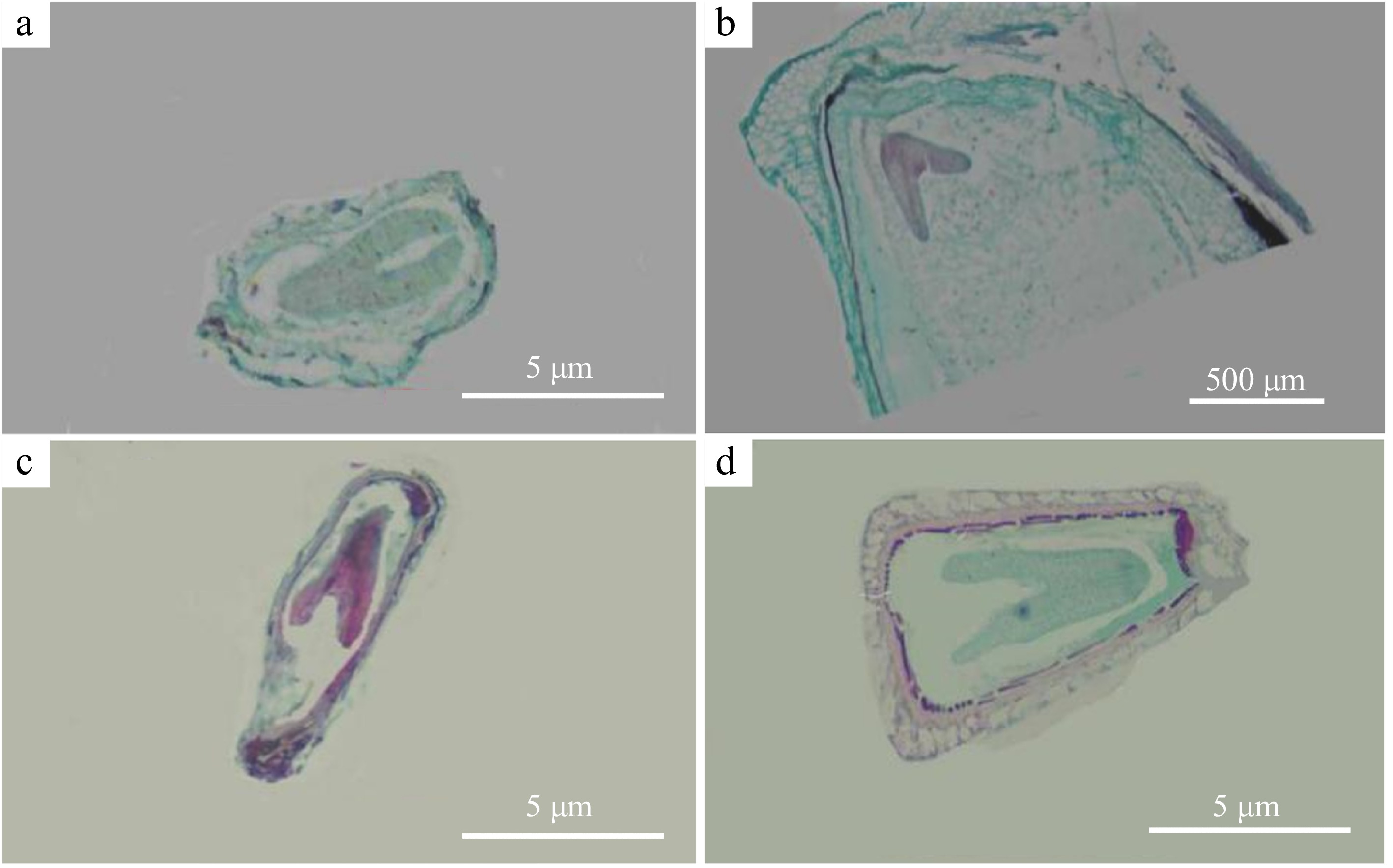
Figure 6.
Embryo development in different cross combinations. (a) Embryos 25 d after open pollination in H. salicifolia. (b) Embryos 25 d after pollination in 'Qianceng Feixue' × 'Sarah's Favorite'. (c) Embryos 15 d after pollination in H. salicifolia × 'Sarah's Favorite'. (d) Embryos 15 d after pollination in H. myrtifolia × L. fauriei.
Obtaining hybrid seedlings through immature embryo rescuing
-
In crosses with Heimia as the female parent, a total of 576 embryos at each developmental stage were inoculated, with 72 seeds per medium combination. For crosses with Lagerstroemia as the female parent, a total of 480 embryos at each developmental stage were inoculated, with 60 seeds per medium combination.
A total of 42 seeds germinated from immature embryo rescue (Table 6). The highest seed germination rate, with H. myrtifolia as the female parent, occurred within 15 d after pollination, yielding 14 seedlings from H. myrtifolia × 'Sarah's Favorite'. Similarly, with H. salicifolia as the female parent, the highest seed germination rate was noted within 25 d after pollination, yielding 10 seedlings from H. salicifolia × 'Sarah's Favorite'. When Lagerstroemia species and cultivars served as female parents, only two crosses produced seedlings: two of 'Sarah's Favorite' × H. salicifolia, and one of 'Dallas Red' × H. salicifolia.
Table 6. Number of seedlings obtained in different media and different times after pollination.
Days after pollination 10 d 15 d 20 d 25 d 30 d No. of medium ck 1 2 3 4 5 6 7 ck 1 2 3 4 5 6 7 ck 1 2 3 4 5 6 7 ck 1 2 3 4 5 6 7 ck 1 2 3 4 5 6 7 Hybrid combinations L.indica 'Sarah's Favorite' 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 0 0 0 0 L.indica 'Dallas Red' × H.salicifolia 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 H.salicifolia × L.indica 'Dallas Red' 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 H.salicifolia × L.indica 'Sarah's Favorite' 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6 3 1 1 0 0 0 0 0 0 0 0 0 0 0 H.myrtifolia × L.fauriei 0 0 0 0 0 0 0 0 0 0 0 0 5 0 2 0 0 0 0 0 0 4 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 H.myrtifolia × L.indica 'Sarah's Favorite' 0 0 0 0 0 0 0 0 0 1 0 0 4 0 6 3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 The hormone combinations corresponding to the medium IDs are presented in Table 2. The germinated seeds experienced three different processes: 1) Immature seeds developed callus or vitrification and subsequently died. 2) Immature seeds developed only radicles without germination, then gradually dried up and died. 3) Immature seeds exhibited normal germination, with both germ and radicle development (Fig. 7). Ultimately, only four seedlings were successfully obtained through embryo rescue, including one of H. salicifolia × 'Dallas Red', one from H. salicifolia × 'Sarah's Favorite', and two of 'Sarah's Favorite' × H. salicifolia.

Figure 7.
Immature embryo rescuing of H. salicifolia × 'Sarah's Favorite'. (a) Seedlings 7 d after sowing. (b) Transferred seedlings 20 d after germination. (c) Rooted hybrid seedlings. (d) Rooted hybrid seedlings planted in a pot.
Using SSR analysis, 63 out of 771 primer pairs were amplified in both Lagerstroemia and Heimia. One SSR markers (YYJ951) confirmed that the offspring from the cross between H. salicifolia and 'Sarah's Favorite' as true hybrid, which contained specific markers from both the maternal and paternal parents (Table 7, Fig. 8). The other three offspring showed no male-specific markers. The hybrids appearance and leaf size were intermediate, more closely resembling H. salicifolia (Fig. 9). The hybrids leaf size was 5.83 cm × 1.15 cm, compared to H. salicifolia with 3.48 cm × 0.61 cm and 'Sarah's Favorite' with 6.51 cm × 4.42 cm. The leaf morphology and floral traits of the hybrid closely resembled those of H. salicifolia (Fig. 9).
Table 7. SSR loci used in hybrid identification.
Locus Repeat motif Forward primer sequence (5'-3') Reverse primer sequence (5'-3') YYJ-951 (GAA)6 5'-GTTCCTGGAGACTTGGGCTC-3' 5'-TCCGAACTTCCCGACGAATG-3' 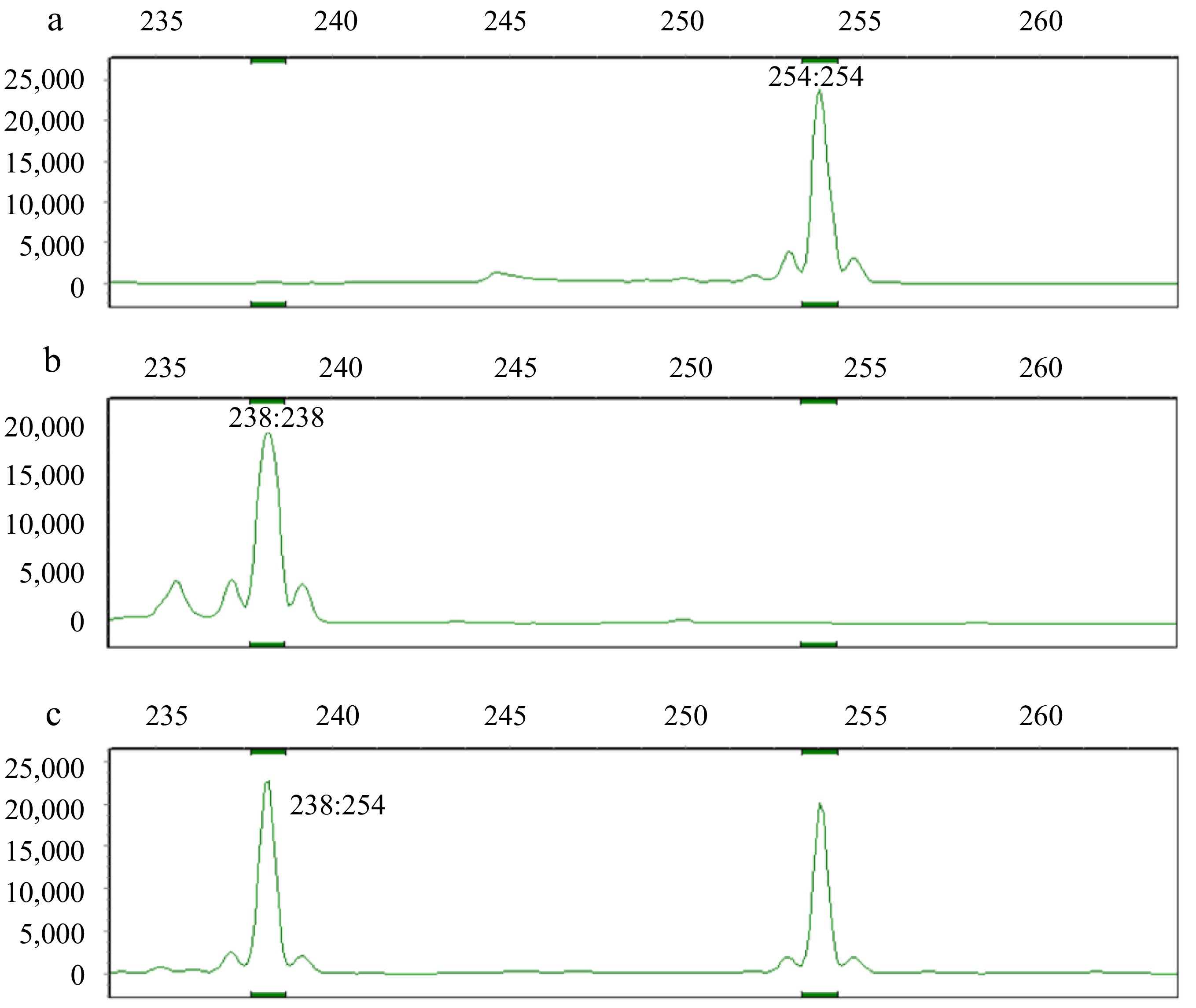
Figure 8.
Profiles obtained using SSR marker (YYJ951). (a) female (H. salicifolia) profile. (b) paternal ('Sarah's Favorite') profile. (c) hybrid profile. The allele sizes of male and female parents, and hybrid were marked.

Figure 9.
Branches and flowers of hybrid (H. salicifolia × 'Sarah's Favorite') and its parents. From left to right: H. salicifolia, hybrid, and 'Sarah's Favorite'.
-
Cross-incompatibility barriers prevailed in intergeneric hybridization[19]. After artificial pollination, the hybrid fruits of Lagerstroemia and Heimia could not develop naturally, which indicated that there was serious reproductive isolation between Lagerstroemia and Heimia. Pre-fertilization barriers are chiefly attributed to the abnormal swelling, deformation, or complete halt in the growth of the pollen tube, preventing it from reaching the ovule[38]. While pollen viability correlates with seed set rates, there was no inevitable causality[39,40]. In our study, parental pollen viability exceeded 50%, meeting the standards required for successful cross-pollination[11]. Researchers suggested that normal pollen tube elongation requires the absence of callose deposition at the tube apex within the stigma, along with continuous exocytosis and endocytosis[41]. The male pollen from 26 cross-combinations could germinate on the stigma surface of the female parent. However, aberrant callose accumulation post-pollination triggered callose plug formation, inducing pollen tube growth arrest and failed ovary penetration. This phenomenon may represent the most significant challenge contributing to the low seed-setting rate observed.
Post-fertilization barriers are evidenced by the halt in ovule development, the inability of seeds to germinate, and the emergence of albino seedlings[42]. In our study, untreated intergeneric hybrid combinations with Lagerstroemia as the maternal parent exhibited complete fruit abscission within 5 d after pollination, while those with Heimia as the maternal parent resulted in fruit shriveling and blackening within 15 d. We successfully prolonged the fruit development period by applying mentor pollen and exogenous hormones. Hybrid fruits with Lagerstroemia as the maternal parent exhibited extended development beyond 35 d after pollination, whereas those with Heimia as the maternal parent showed signs of wilting after 25 d, which likely resulted from embryo abortion occurring after the development of the embryos to the heart-shaped stage[43]. Paraffin sectioning results indicated that only a small proportion of fruits reached the heart-shaped embryo stage, suggesting ovule abortion or incomplete seed maturation. Additionally, there are also symptoms of post-fertilization barriers, including low seedling germination rates, premature senescence, and stunted growth during the embryo rescue process[42]. The results demonstrated significant pre-fertilization and post-fertilization barriers between Lagerstroemia and Heimia.
Overcoming the barrier of distant hybridization
-
The location of double fertilization barriers could occur in three stages: pollen-stigma interaction, pollen tube growth in the style, and gametes fusion[25]. In our study, pre-fertilization barriers predominantly occurred between the pollen tube and the style. Compatibility in distant hybridization can be enhanced by employing various pollination techniques, including mentor pollination, repeated pollination, heated pollen method, cut-style pollination, and delayed pollination method[18,44]. Self-incompatibility in Brassicaceae refers to the mechanism by which stigma mastoid cells detect the S-locus cysteine-rich protein (SCR) signals presented by self-pollen through the S-locus receptor kinase (SRK), inhibiting pollen germination. Pollen carried small peptides (PCP-Bs) that interact with FERONIA receptor kinase to overcome this inhibition[16]. Stigma-mediated rejection of incompatible pollen is essential to prevent outcrossing and this barrier can be mitigated with the application of mentor pollen[17]. In our study, mentor pollen reduced callose production and enhanced both pollen germination and pollen tube elongation to an extent.
Endogenous hormonal abnormalities and imbalances that occur during embryogenesis frequently lead to embryonic abortion, especially in the case of hybridizations between distantly related species[26,45]. In distant hybridization, applying exogenous hormones to the stigma of the female parent can help maintain fruit viability, enhance fruit set, and promote fruit growth[1]. Numerous studies have indicated that auxin regulates fruit growth and development by interacting with other hormones[27,46]. Measuring endogenous hormones during fruit development reveals variations in hormone levels across plant species and developmental stages[18]. In distant hybridization of peonies, aborted embryos exhibited elevated levels of ABA, IAA, and GA[26]. The absence of ABA and BR in stigmas contributed to inter-subgeneric hybridization barriers in water lilies[47]. GA3 treatment played a certain role in maintaining the fruiting rate in the middle and late stages after pollination[48]. In this study, the stigma was treated with exogenous hormonal treatment after pollination, which alleviated the fruit shedding to a certain extent. Utilizing Lagerstroemia as the maternal parent, GA3 achieved higher fruit set rates than 2,4-D. In contrast, when Heimia species served as the maternal parent, hybrid fruits treated with 2,4-D retained their green color for a more extended period compared to those treated with GA3. Although post-fertilization barriers were not completely surmounted, the hormonal treatment succeeded in prolonging the development of the hybrid fruits.
Embryo rescue, a technique pivotal in overcoming post-fertilization barriers, is extensively applied in distant hybridization studies, including plum cultivars[49], Anoectochilus[50], Brassica species[43], and other species. The germination rate was also strongly influenced by embryo size, with the optimal embryo developmental stage being a critical factor, as the development of early embryos in vitro conditions is poor, and delayed rescue may induce abortion[49]. The heart-shaped to torpedo-shaped embryo stage is typically optimal for embryo rescue[25,43]. Young embryos need a complex medium, while mature embryos can usually grow in a simple medium. During the post-rescue germination phase, we noted low seedling emergence rates, premature senescence, and inhibited growth in seedling development. These observations may correlate with immature embryo development and late-stage embryo abortion of distant hybridization. We determined that the optimal time for embryo rescue with Heimia as the female parent was 15 to 25 d after pollination, while it was 25 to 30 d after pollination for Lagerstroemia as the female parent. Selecting the optimal period for embryo rescue is crucial for future endeavors in this field.
Prospect of distant hybridization between Lagerstroemia and Heimia
-
Cross compatibility in plant hybridization, particularly in distant hybridization, is closely related to parental chromosome ploidy levels, making the selection of parental materials crucial. It was demonstrated that hybridization is more likely to succeed when the parental ploidy levels are similar, whereas divergent ploidy exacerbates reproductive barriers[24,51]. Variations in the chromosome number or ploidy level between parents exert differential impacts on the hybridization outcomes. In certain plant species, such as Mimulus[24] and Campanula rotundifolia[52], the likelihood of producing triploid offspring is greater when the male parent possesses a higher ploidy level. Conversely, in other species like Brassica oleracea[53] and Festuca[54], the production of triploid offspring is more successful when the female parent has a higher ploidy level. Furthermore, researchers successfully obtained triploid progeny of Centaurea by crossing diploid C. pseudophrygia (female) with a tetraploid C. jacea (male)[55]. Lagerstroemia (2n = 48) and Heimia (2n = 16) are both diploid but with different numbers of chromosomes. In this study, obtaining a normally developed hybrid fruit proved challenging. Nevertheless, the utilization of pollen mentor techniques, hormonal treatments, and embryo rescue strategies have been demonstrated to enhance seed set rates, thereby facilitating the acquisition of hybrid fruits and seedlings. We observed that the ratio of pollen tube elongating into the ovule was higher when Heimia served as the female parent. Future research endeavors should focus on hybridization utilizing Heimia as the maternal parent.
We successfully rescued four embryo seedlings and employed one pair of SSR molecular markers to authenticate the hybrid of H. salicifolia × 'Sarah's Favorite'. SSR markers are widely used in the verification of plant hybrid progenies, where the authenticity of hybrid offspring is commonly ascertained through the detection of 1 to 2 parent-specific markers[11,56]. The phenotype of the hybrid exhibited a stronger maternal resemblance. Matroclinal inheritance, involves the transmission of genes across multiple genetic networks, leading to the traits of hybrid offspring to resemble the female parent[19]. The traits of the hybrid tended to be biased towards the female parent, suggesting the presence of matroclinal inheritance, a phenomenon also noted in studies on Pyrus spp.[57] and intergeneric hybridization of Dimocarpus longan and Litchi chinensis[19].
In subsequent intergeneric hybridization between Lagerstroemia and Heimia, strategies could be employed to enhance the seed-setting rate. Initially, broad parental screening of species or cultivars should be made based on successful hybrid fruit production, followed by the identification of combinations with high cross-compatibility[24,38,52]. Secondly, amphidiploidization of the hybrid plant is essential to restore fertility and facilitate the production of an F2 generation, or backcrossing may be performed for advanced breeding purposes[32,51]. Thirdly, pollination methods should be optimized to overcome pre-fertilization barriers associated with outbreeding incompatibility, increase the success rate of pollen tube penetration into the ovary for fertilization, and simultaneously improve the recovery rate of hybrid embryos at the heart-shaped stage, which is critical for subsequent embryo rescue work[27,40,43,55].
-
We have identified the pre- and post-fertilization barriers in distant hybridization between Lagerstroemia and Heimia. By employing mentor pollen, exogenous hormone spraying, and embryo rescue techniques, we overcame the barriers and successfully obtained one hybrid progeny, and identified the true hybrids at the molecular level. The results will provide a method for breeding yellow-flowered Lagerstroemia cultivars.
This work was supported by the National Key R and D Program of China (2023YFD1200105).
-
The authors confirm contribution to the paper as follows: study conception and design: Pan H; data collection: Lin Q, Chi X, Shen P, Wan Z; analysis and interpretation of results: Lin Q, Wang X, Tao J; draft manuscript preparation: Lin Q, Cai M, Wang J, Zhang Q, Cheng T. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lin Q, Wang X, Wan Z, Tao J, Chi X, et al. 2025. Obtained intergeneric hybrids from distant hybridization between Lagerstroemia and Heimia. Ornamental Plant Research 5: e017 doi: 10.48130/opr-0025-0016 

Obtained intergeneric hybrids from distant hybridization between Lagerstroemia and Heimia
- Received: 07 November 2024
- Revised: 27 February 2025
- Accepted: 03 March 2025
- Published online: 16 April 2025
Abstract: Lagerstroemia comes in a variety of bright petal colors, but it lacks yellow flowers. To breed yellow-flowered cultivars, eight Lagerstroemia and two Heimia species were used for intergeneric hybridization. Mentor pollen was applied to enhance pollination efficiency, and GA3 and 2,4-D were sprayed onto stigmas after pollination to extend hybrid fruit development. Progenies were obtained by embryo rescue and identified as hybrids between Lagerstroemia and Heimia by SSR markers. The results showed that there was strong reproductive isolation between Lagerstroemia and Heimia. When Lagerstroemia was the female parent, the fruits dropped within 5 d after pollination, and when Heimia was the female parent, the fruits did not drop, but dried up within 15 d. After pollination, pollen tubes of Heimia in the styles of Lagerstroemia halted at one-fourth of the style length, with few entering the ovary. While pollen tubes of Lagerstroemia in the style of Heimia reached the stigma base but rarely entered the ovary. Mentor pollen reduced callose production and promoted the growth of pollen tubes. Most hybrid embryos aborted on plants naturally, and applying 200 mg·L−1 GA3 to Lagerstroemia pistils and 20 mg·L−1 2,4-D to Heimia pistils promoted some of the hybrid fruits to the heart-shaped embryo stage. In our experiment, a total of 12,350 flowers were pollinated, and only four hybrid progenies were obtained by embryo rescue, with one hybrid confirmed as a true hybrid. The results provided a foundation for breeding yellow-flowered Lagerstroemia cultivars.
-
Key words:
- Intergeneric hybridization /
- Lagerstroemia /
- Heimia /
- Embryo rescue /
- Reproductive barrier