

-
Seeds are the primary means of safely conserving crop germplasm resources[1]. To ensure that these resources maintain high viability, seed viability must be monitored both before and during conservation in Genebanks. Currently, the traditional germination test is the primary method for assessing seed viability[2]. However, during germination, some seeds may become infected by fungi, which can hinder seed germination and affect the development of the radicle and cotyledon. This reduces the germination percentage and index, complicating accurate viability assessment and severely impacting the safe conservation of germplasm resources.
Some studies have reported that seeds of faba bean (Vicia faba L.), pea (Pisum sativum L.), soybeans (Glycine max L.), maize (Zea mays L.), sunflower (Helianthus annuus L.), peanut (Arachis hypogaea L.), and sorghum (Sorghum bicolor L.) are often infected by large quantities of fungi during the germination process[3−9]. The rapid proliferation of fungi competes for nutrients originally intended for the growth of the radicle or cotyledon. Additionally, the fungal mycelium spreads and envelops the radicle and cotyledon, inhibiting their growth. Fungal proliferation requires an abundant supply of nutrients such as amino acids, lipids, sugars, and others[10]. These nutrients primarily come from the breakdown of seed storage materials during germination, such as the conversion of starch into simple sugar, proteins into peptides and amino acids, and fats into fatty acids and glycerol[11−13]. The growth of fungi reduces the availability of carbon, nitrogen, and energy for embryo development, thus inhibiting the growth of the radicle and plumule. Fungi also produce toxins, such as cladosporin and deoxynivalenol, which inhibit the growth of host cells. However, plants have an immune system that is specifically tailored to combat fungal contamination. The strength of a plant's immune system is directly proportional to its survival rate[14]. The structure of the seed coat also plays a critical role in the carriage of fungi. Some seeds have wrinkled or indented seed coats, which provide niches for microbes to lodge. For example, the outer seed coat of sorghum seeds has grooves, making thorough disinfection difficult, while the seed coats of peanuts are brittle and thin, with low moisture content, making them prone to damage[15].
By monitoring the viability of seeds in Genebanks, faba bean, pea, soybean, maize, sunflower, peanut, and sorghum are particularly susceptible to fungal contamination, highlighting the need for optimized germination protocols for these crops. In this study, the types and abundances of fungi in these seeds were identified using high-throughput sequencing and Sanger sequencing. The goal was to further optimize disinfection conditions and germination substrates to mitigate the effects of fungal contamination and to develop tailored germination protocols for these crops, providing technical support for seed viability monitoring.
-
Seeds from 11 varieties of each crop such as faba bean, pea, soybean, maize, sunflower, peanut, and sorghum are listed in Supplementary Table S1. Seed germination was carried out according to the International rules for seed testing (ISTA) protocol[16]. The seed of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species'), and sorghum ('CP21') were selected as a representative for subsequent experiments. The factors of agar concentration, NaClO concentration, and disinfection time were evaluated at three levels in an orthogonal experiment for optimize germination protocol. The test varied NaClO solution concentrations (6%, 8%, and 10%), disinfection times (15 min, 25 min, and 35 min), and agar concentrations in the culture medium (1.75 g/L, 3.5 g/L, and 7 g/L), with a pH range of 5.8−6.0.
Isolation and identification of fungi using Sanger sequencing
-
Seven species of crop seeds were treated under optimal disinfection conditions. From each crop, four seeds were selected, and four tissue sections were cut from each seed and placed on potato dextrose agar (PDA) medium (potato 200 g, D-glucose 20 g, agar 20 g, distilled water
1,000 mL). The plates were subsequently incubated in the dark at 28 ± 2 °C, and fungal growth was monitored daily. Ampicillin and chloramphenicol were added to the PDA medium to a final concentration of 10 μg/mL, based on the modification of the concentration used by Fang[17], to inhibit bacterial growth. Once fungal colonies appeared, the fungi were purified by transferring a small amount of mycelia from the edge of each colony onto new PDA plates. Each pure strain was preserved in two centrifuge tubes on PDA plates at room temperature. Sanger sequencing was performed using a T5 direct PCR kit (TSE011; TsingKe, Beijing, China) and the V9G primer (5'-TTAAGTCCCTGCCCTTTGTA-3') according to the methods described by Cheng et al.[18]. ITS sequence-based identification was carried out using BLAST against NCBI GenBank database, with hits showing more than 97% similarity to published sequences and their corresponding accession numbers. Additionally, fungi grown from sorghum seeds during germination according to the ISTA protocol were also selected for Sanger sequencing.DNA extraction and high-throughput sequencing
-
DNA extraction was performed from 10 seeds using the Magnetic Soil and Stool DNA Kit (TIANGEN, China). PCR amplification was conducted using fungal primers targeting the ITS1 region, with the primer pair ITS1F (5'-CTTGGTCATTAGAGGAAGTAA-3') and ITS2R (5'-GCTGCGTTCTTCATCGATGC-3')[19]. Each PCR reaction contained 15 μL of Phusion® High-Fidelity PCR Master Mix (New England Biolabs, USA), 0.2 μM of each primer, and 10 ng of genomic DNA template. The PCR cycling conditions included an initial denaturation at 98 °C for 1 min, followed by 30 cycles consisting of denaturation at 98 °C for 10 s, annealing at 50 °C for 30 s, and extension at 72 °C for 30 s, with a final extension at 72 °C for 5 min. The sequencing data were subsequently analyzed on the MiSeq platform at Novogene (Beijing, China).
Processing and analysis of sequencing data
-
Using the methods described by Edgar et al.[20], Cutadapt (version 3.3), FLASH (version 1.2.11,
http://ccb.jhu.edu/software/FLASH/ ), and fastp (version 0.23.1) were applied to filter, merge, quality control, and remove chimera from all the raw sequences of the samples. Denoising was subsequently carried out using the DADA2 module in QIIME2 software (version QIIME2-202202) to generate amplicon sequence variants (ASVs)[21].Normalized ASV abundance data was used for α-diversity analysis. Community richness was assessed using the Chao1 index, while diversity was evaluated with the Shannon and Simpson indices. Coverage was assessed using QIIME2. Samples with the highest relative abundance at the genus level were selected to generate relative abundance distribution histograms using the SVG function in Perl. Unique information among different taxa was visualized using Venn diagrams in R and SVG functions in Perl.
Statistical analysis
-
The germination rate, radicle length, and plumule length data were analyzed using one-way ANOVA in SPSS 24.0 (IBM Inc., Armonk, NY, USA). A p-value less than 0.05 was considered as statistically significant.
-
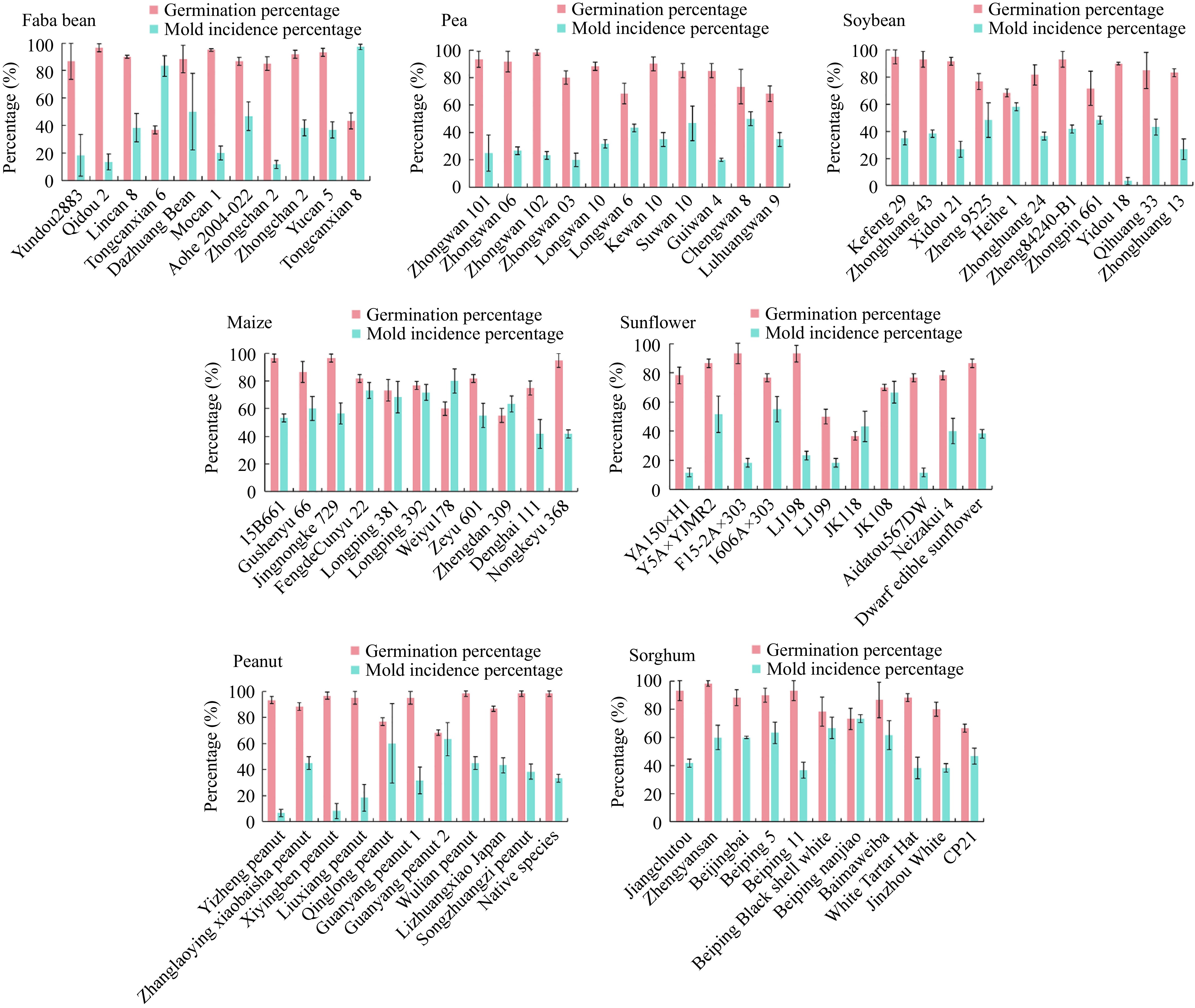
To better analyze the impact of fungi on the seed viability monitoring, 11 varieties of seeds of faba beans, peas, soybeans, maize, sunflowers, peanuts, and sorghum were selected from the Genebank for germination experiments according to the ISTA protocol. The results showed that fungal contamination was a common occurrence during germination of these seven crops (Fig. 1). The fungal mold incidence varied across different crops and seed viability levels. Notably, a positive correlation was observed between germination percentage and mold incidence: seeds with lower germination percentages tended to exhibit higher mold incidence (Fig. 1). Fungi with rapid growth rates are more likely to negatively affect seed germination and development. Those results indicated that the seed was affected by fungi during germination, which inhibited seedling development, leading to a reduced germination percentage and vigor indexes. During the conservation in Genebanks, the decline in seed viability is inevitable, requiring the monitoring of germplasm viability to ensure high vitality is maintained. However, as viability decreases, the likelihood of fungal infection and its impact on seed germination intensifies, which can affect the accurate assessment of seed viability (data not shown).
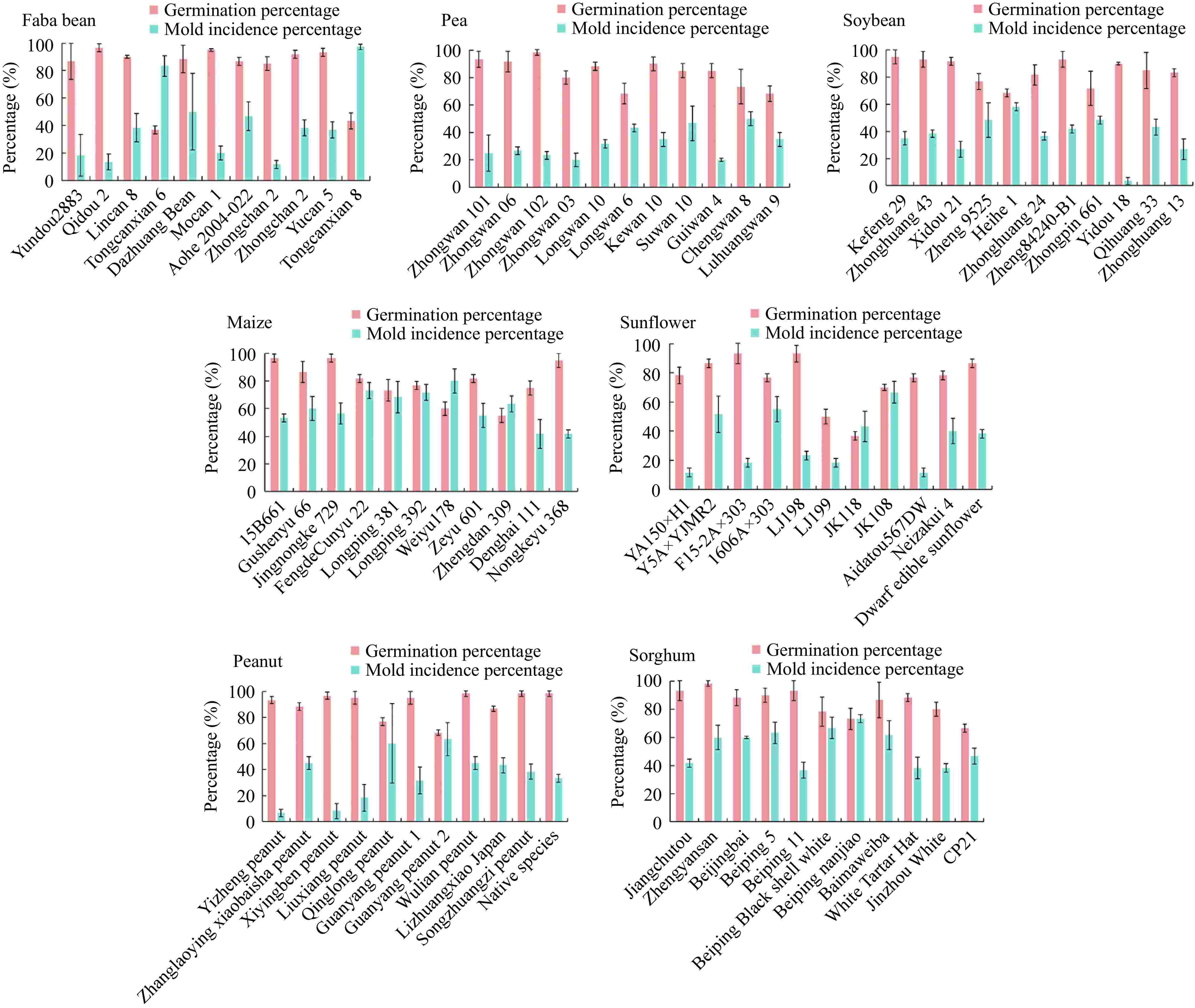
Figure 1.
Detection germination percentage and mold incidence percentage in seeds from 11 varieties of each faba bean, pea, soybean, maize, sunflower, peanut, and sorghum, germinated according to the ISTA protocol. Values are presented as means ± SD (n = 3).
Identification and classification of fungi associated with seeds
-
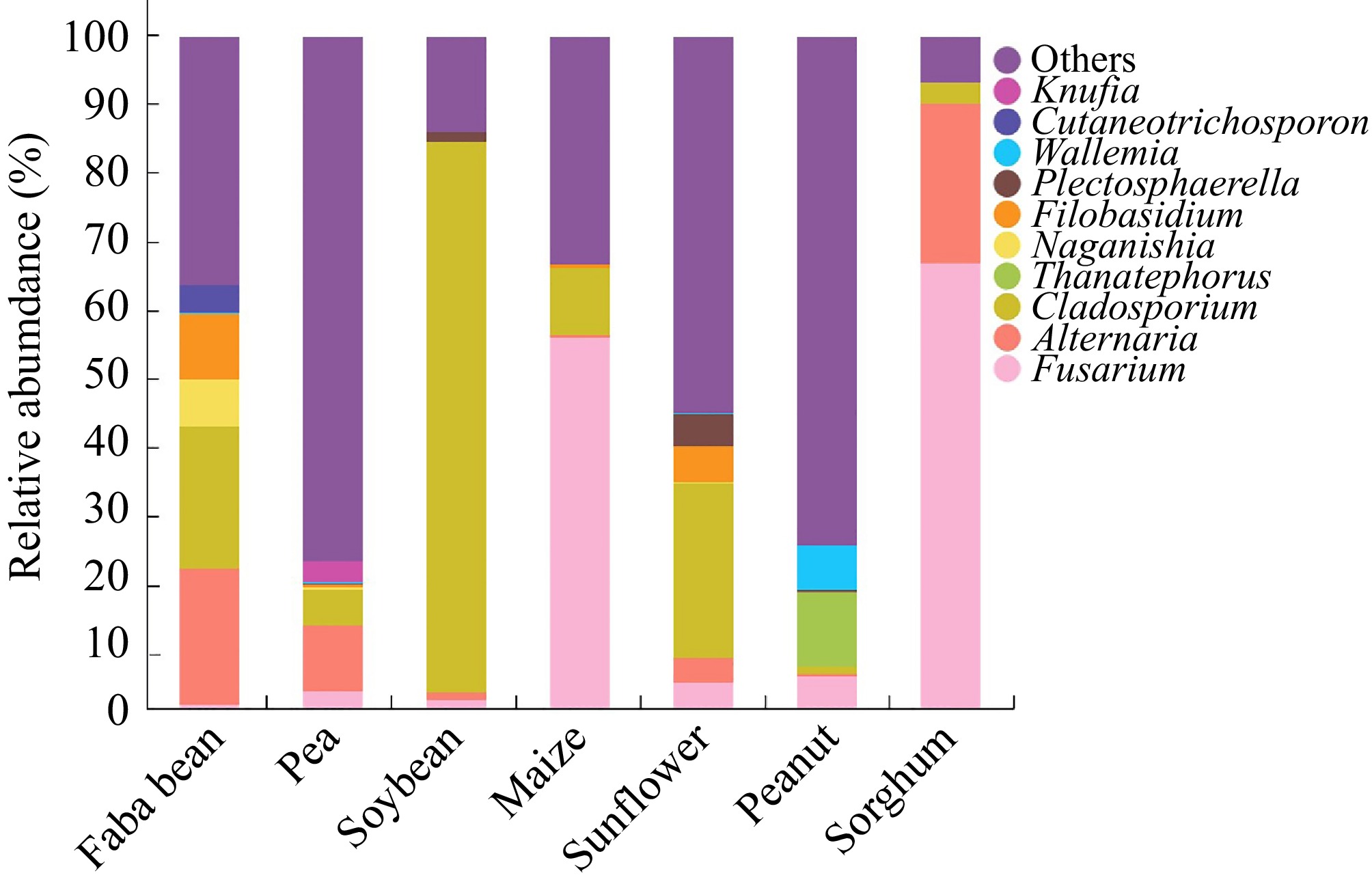
Further analysis of fungal presence on the seeds was conducted by selecting one sample from each of the seven crops for high-throughput sequencing analysis. The results revealed that Alternaria, Cladosporium, and Fusarium were present and distributed across all seven crop species seeds, with relatively high abundances (Fig. 2). Fusarium is a diverse and widely distributed pathogen that infects many crops. It commonly affects the roots, stems, and ears of plants, causing rot diseases that inhibit plant growth[22]. Certain Fusarium variants produce toxic metabolites, such as deoxynivalenol, a mycotoxin that can damage cell structure[23]. Numerous studies have shown that deoxynivalenol disrupts the physiological process during seed development in crops like sorghum and maize[9,24−27]. The relative abundance of Fusarium in sorghum ('CP21') and maize ('Nongkeyu 368') seeds was 66.6% and 55.4%, respectively, which may contribute to the low vigor index observed in these seeds. Alternaria, a fungal genus of significant concern in plant diseases, produces toxic metabolites like Alternaria mycotoxins, which cause plant wilting, lesions, and rotting. Previous research has identified these fungi in various crops[28−31]. However, some studies suggest that certain Alternaria species are relatively weak opportunistic pathogens[32]. In sorghum ('CP21'), faba bean ('Tongcanxian 6'), and pea ('Zhongwan 06') seeds, Alternaria accounted for 23.6%, 20.3%, and 9.8%, respectively, of the relative abundance. Cladosporium can produce mycotoxins, such as cladosporin, which damage the cell structure and function, leading to tissue necrosis and decay[33−35]. In faba bean ('Tongcanxian 6'), soybean ('Zhonghuang 13'), and sunflower ('Dwarf edible sunflower') seeds, Cladosporium accounted for 21.1%, 18.8%, and 25.9%, respectively, of the relative abundance of fungi. Some studies also indicated that certain Cladosporium species cause minimal damage and may be considered weak pathogens[36].
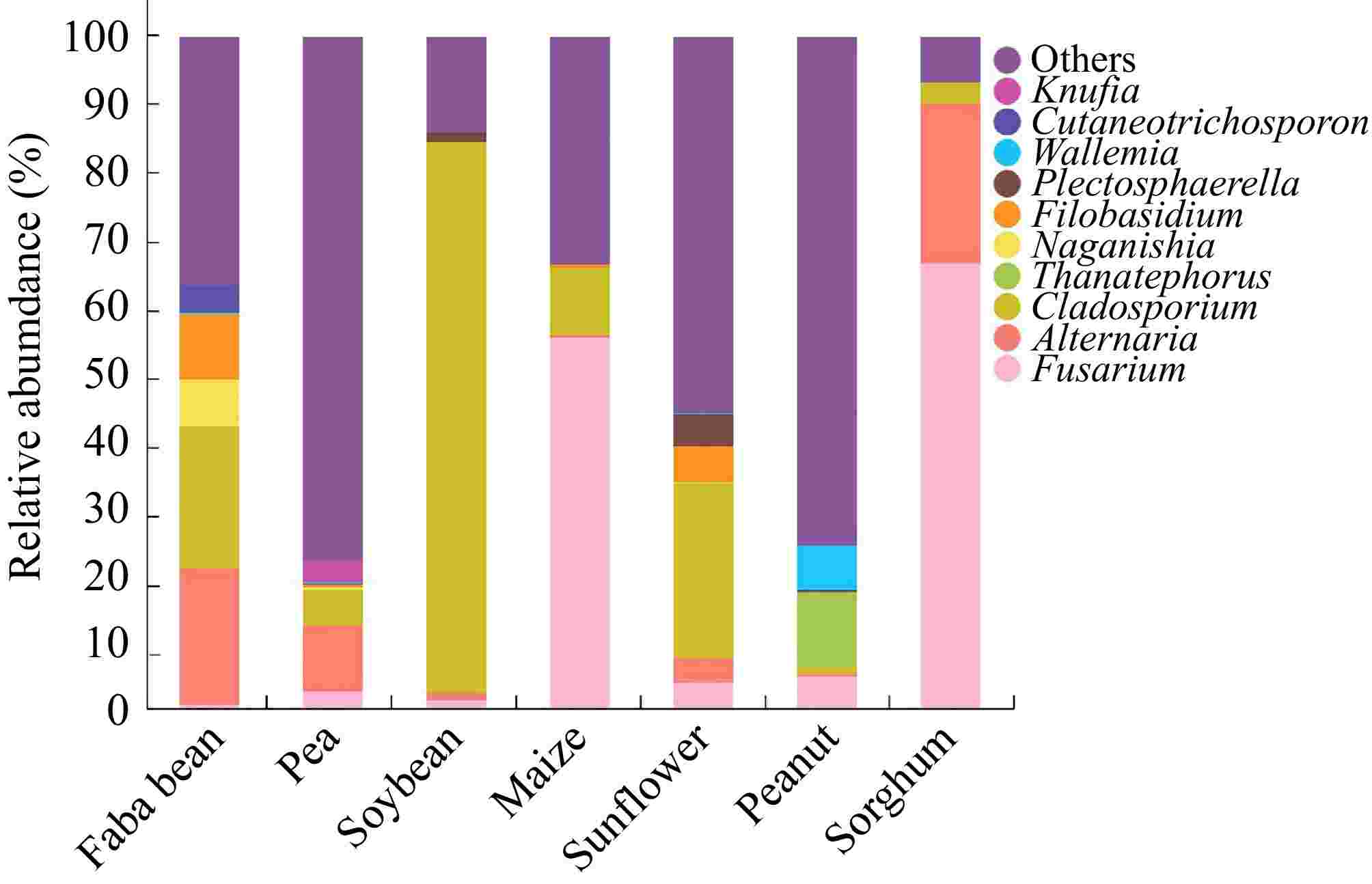
Figure 2.
Identification and classification of fungi associated with seeds of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species'), and sorghum ('CP21') as determined by high-throughput sequencing.
Additionally, Table 1 shows the highest abundance of the top 10 fungal genera in the seven crop seeds. Plant pathogens have been reported in most of these genera. Aspergillus is an opportunistic fungal pathogen of oilseed crops, contaminating them with mycotoxins. Bipolaris is another pathogen that causes leaf blight disease[37,38]. Rhizopus and Thanatephorus can secrete enzymes such as keratinase and cellulase to degrade the cuticle and cell walls, enabling them to invade and reproduce within cells, thus disrupting cell metabolic function[39−41]. Talaromyces has been identified as a plant pathogen responsible for plant rot and discoloration[42]. Other genera, such as Didymella and Plectosphaerella, produce toxins that might inhibit the growth of host cells and were also detected in seeds[43,44]. To further investigate the presence of pathogenic fungi in the seeds of the seven crop species, Sanger sequencing was used to identify fungi infecting the seedlings germinated under the ISTA protocol, as well as fungi growing on seed tissue blocks in PDA plates. The results showed that fungi strains isolated from both sources included Alternaria, Cladosporium, and Fusarium, which is consistent with the high-throughput sequencing results (Table 2).
Table 1. The fungal genera of the top 10 most abundant detected in seeds of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species'), and sorghum ('CP21') as identified through high-throughput sequencing.
Faba bean Pea Soybean Maize Sunflower Peanut Sorghum Cladosporium Alternaria Cladosporium Fusarium Cladosporium Thanatephorus Fusarium Alternaria Cladosporium Humicola Cladosporium Filobasidium Wallemia Alternaria Filobasidium Didymella Fusarium Sarocladium Plectosphaerella Fusarium Cladosporium Naganishia Knufia Plectosphaerella Filobasidium Fusarium Nigrospora Ramichloridium Cutaneotrichosporon Fusarium Hannaella Trichoderma Alternaria Cladosporium Edenia Talaromyces Vacuiphoma Alternaria Alternaria Golovinomyces Neocosmospora Ophiosphaerella Vishniacozyma Botrytis Aspergillus Papiliotrema Botrytis Paramyrothecium Phaeosphaeria Aspergillus Celosporium Rhodosporidiobolus Rhizopus Penicillium Talaromyces Hannaella Fusarium Nigrospora Acremonium Aspergillus Rhizopus Malassezia Moesziomyces Didymella Dactylonectria Malassezia Vishniacozyma Hanseniaspora Plectosphaerella Bipolaris Table 2. The occurrence of Alternaria, Cladosporium, and Fusarium identified in seedlings and tissue blocks through Sanger sequencing, and in entire seeds through high-throughput sequencing of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species'), and sorghum ('CP21').
Fungi Organism Sequencing Crop species Faba bean Pea Soybean Maize Sunflower Peanut Sorghum Alternaria Seedlings Sanger + Tissue blocks Sanger + + + Seeds High-throughput + + + + + + Cladosporium Seedlings Sanger + + Tissue blocks Sanger + + Seeds High-throughput + + + + + + + Fusarium Seedlings Sanger + + Tissue blocks Sanger + + + Seeds High-throughput + + + + + + + + represents the genus of the fungi detected. To further determine whether those fungi widely infect the seeds of seven crop species during germination according to the ISTA protocol, Sanger sequencing were performed on fungi proliferating during the germination of seeds from 11 varieties of each crop (Supplementary Table S2). The results revealed that Fusarium fungi are commonly present in different varieties of all seven crop species and may contribute to the inhibition of seed germination. Alternaria and Cladosporium were detected at low frequencies across the seven crop species and are considered opportunistic pathogens. Rhizopus and Aspergillus were more frequently observed and may also contribute to the decline in germination indices. Therefore, to accurately assess seed viability, it is essential to optimize and develop germination protocols to eliminate or reduce the inhibitory effects of these genera.
Optimized protocols for crop seed germination
-
To optimize the seed germination protocol that suppresses fungal infection, a three-factor, three-level orthogonal was conducted. The test varied NaClO solution concentrations (6%, 8%, and 10%), disinfection times (15 min, 25 min, and 35 min), and agar concentrations in the culture medium (1.75 g/L, 3.5 g/L, and 7 g/L), with a pH range of 5.8−6.0. The effectiveness of the germination protocols in inhibiting fungi was evaluated based on fungal proliferation, seedling vigor, and germination percentage (Fig. 3, Supplementary Table S3). The results revealed fungal contamination and poor seedling growth in all seven crop seeds germinated in a medium with 1.75 g/L or 3.5 g/L agar after various NaClO disinfection treatments. In contrast, no fungal contamination was observed in cultures of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), or peanut ('Native species') seeds in medium with 7 g/L agar. Only a few soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), and sunflower ('Dwarf edible sunflower') seedlings exhibited slight fungal infection; while sorghum ('CP21') seedlings were infected but showed better viability. These findings suggest that a 7 g/L agar concentration in the culture medium provides relatively optimal germination conditions. We hypothesize that the 7 g/L agar concentration creates a high-osmotic environment that promotes physiological metabolism and enhances immunity, thereby reducing fungal contamination[45,46].

Figure 3.
Assessment of fungal contamination on the germination of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species'), and sorghum ('CP21') seeds treated with NaClO solutions at concentrations of 6%, 8%, and 10%, disinfection times of 15 min, 25 min, and 35 min, and agar concentrations of 1.75 g/L, 3.5 g/L, and 7 g/L in the culture medium, with a pH range of 5.8–6.0.
NaClO has strong oxidizing properties and is commonly used to eliminate fungi in seeds or in vitro-generated plants[47,48]. An appropriate concentration of NaClO and treatment duration can promote healthy seedling growth, while excessive treatment may cause tissue damage and growth inhibition[49,50]. In this study, the most suitable disinfection conditions were found to be 10% NaClO for 35 min for seeds such as faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), sunflower ('Dwarf edible sunflower'), and sorghum ('CP21'); 6% NaClO for 25 min for peanut ('Native species'), and maize ('Nongkeyu 368') seeds; and 8% NaClO for 15 min for soybean ('Zhonghuang 13') seeds (Table 3).
Table 3. The optimal germination protocol for faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species'), and sorghum ('CP21'), including NaClO solution concentrations, disinfection times, and agar concentrations in the culture medium.
Specie NaClO concentration (%) Disinfection time (min) Agar concentration (g/L) Faba bean 10 35 7 Pea 10 35 7 Soybean 8 15 7 Maize 6 25 7 Sunflower 10 35 7 Peanut 6 25 7 Sorghum 10 35 7 The efficacy of the optimized disinfection conditions for fungal elimination from seeds was assessed using PDA medium. The results showed that after culturing on PDA medium, fungal growth was not observed in the tissue blocks from disinfected seeds of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), and peanut ('Native species') . Fungal growth was detected only in the sorghum ('CP21') tissue blocks, suggesting that the germination protocols for sorghum require further optimization.
Impact of optimizing germination protocols on fungal species and seed viability
-
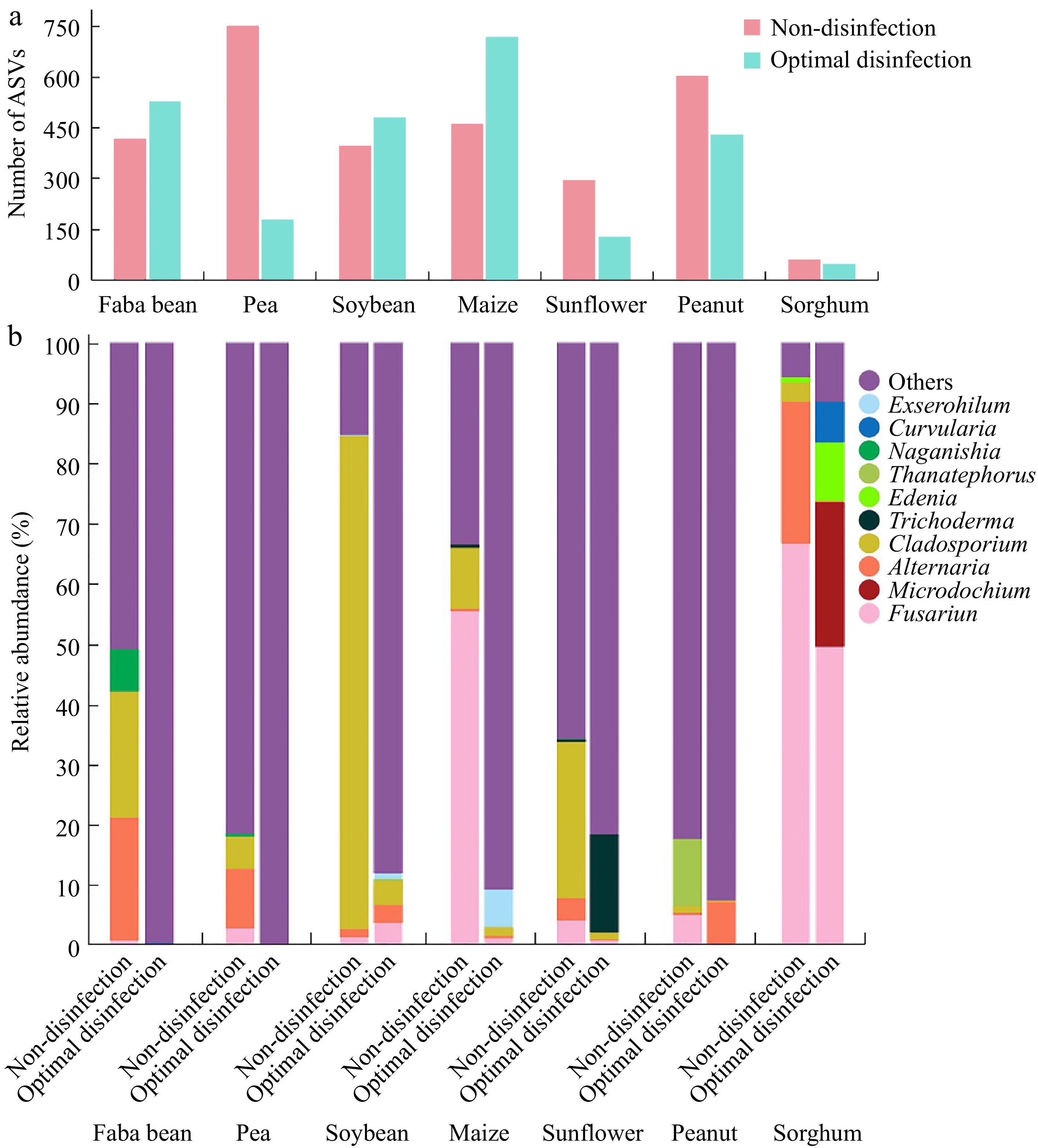
To investigate the impact of the NaClO disinfection protocol on fungi in seeds, the diversity and abundance of fungi in seven crop seeds after disinfection were analyzed using high-throughput sequencing. Compared to pre-disinfection levels, the number of ASVs decreased from 749 to 176 in pea ('Zhongwan 06'), from 603 to 426 in peanut ('Native species'), from 292 to 125 in sunflower ('Dwarf edible sunflower'), and from 57 to 44 in sorghum ('CP21'). In contrast, the number of ASVs increased in faba bean ('Tongcanxian 6'), maize ('Nongkeyu 368'), and soybean ('Zhonghuang 13')—rising from 417 to 525 in faba beans, from 460 to 715 in maize, and from 396 to 478 in soybean. However, the abundance of unannotated sequences significantly increased (Fig. 4). The relative abundance of fungi (such as Alternaria, Cladosporium, Fusarium, and Thanatephorus) found before disinfection was significantly reduced post-disinfection. Additionally, except for sorghum ('CP21') seeds, the growth of surface fungi on PDA plates was markedly reduced in the seeds of seven crops treated with the disinfection protocol, compared to untreated seeds, after rolling a few times on the PDA plates (Supplementary Fig. S1).
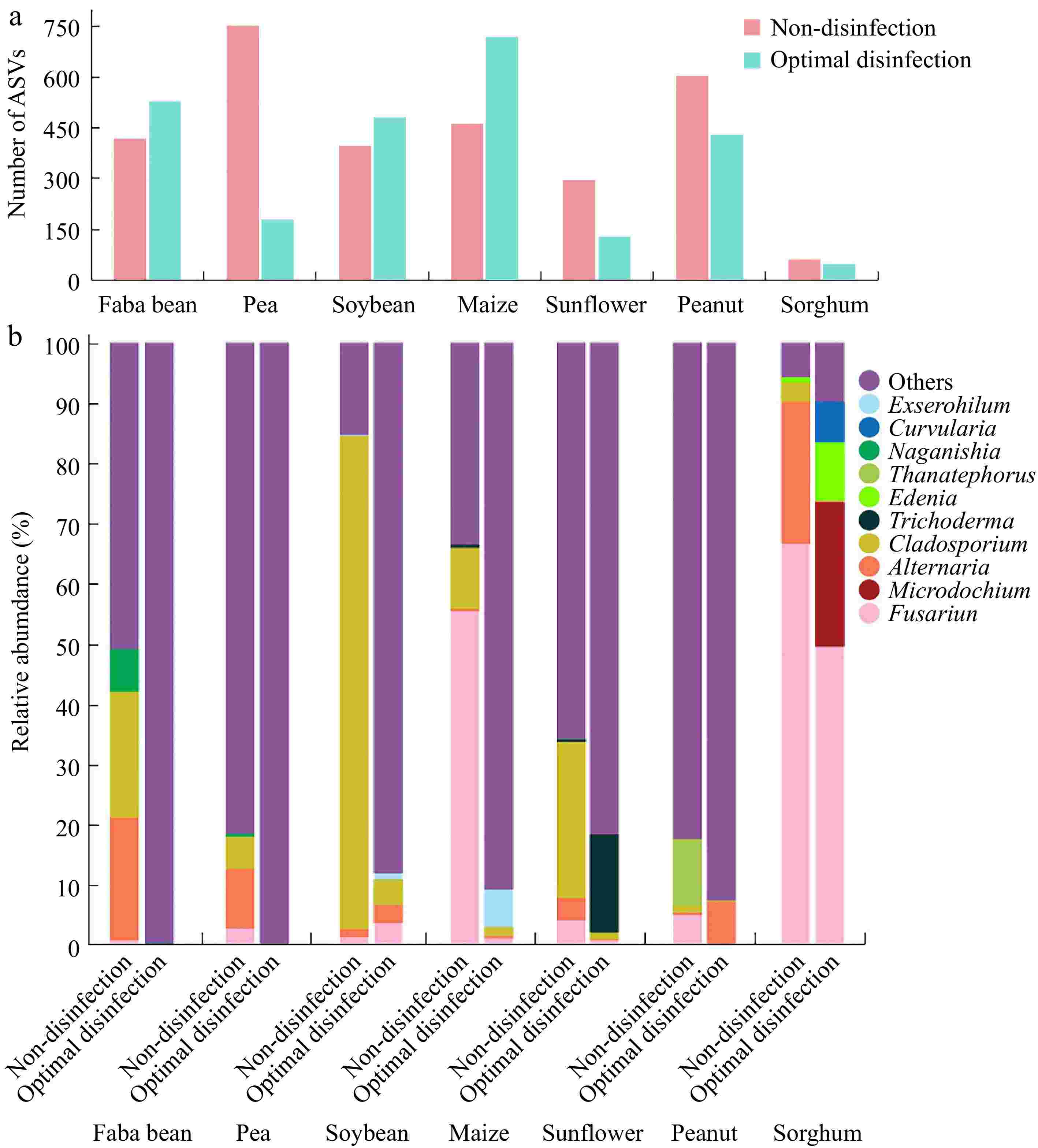
Figure 4.
Composition of the major fungi in faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species'), and sorghum ('CP21') seeds treated with optimal disinfection protocol and without disinfection as determined through high-throughput sequencing. (a) The number of Amplicon Sequence Variants (ASVs). (b) Relative abundance of major fungal genera.
Compared to the fungi in sorghum ('CP21') seeds before disinfection, the relative abundances of Alternaria, Cladosporium, and Fusarium decreased after disinfection with 10% NaClO for 35 min. However, the relative abundances of genera such as Curvularia, Edenia, and Microdochium increased, with values of 24.0%, 9.7%, and 6.8%, respectively. These genera have all been confirmed as pathogenic[51−53] (Fig. 4). Similarly, Sanger sequencing analysis identified the fungal species in the tissue blocks of sorghum ('CP21') seeds treated with the ISTA protocol and optimal germination protocol on PDA plates as Bipolaris zeicola (55.6%), Epicoccum sorghinum (22.2%), Alternaria alternata (11.1%), and Fusarium fujikuroi (11.1%). Alternaria, Bipolaris, Curvularia, and Edenia belong to the Pleosporaceae family, which includes many pathogenic fungi of grass[52,54] (Supplementary Table S4). The results suggest that the cortical structure near the embryo of sorghum seeds shrinks, which may lead to incomplete disinfection. Placing the disinfected seeds in a 7 g/L solid agar medium for viability monitoring can mitigate the impact of fungi on seedling growth.
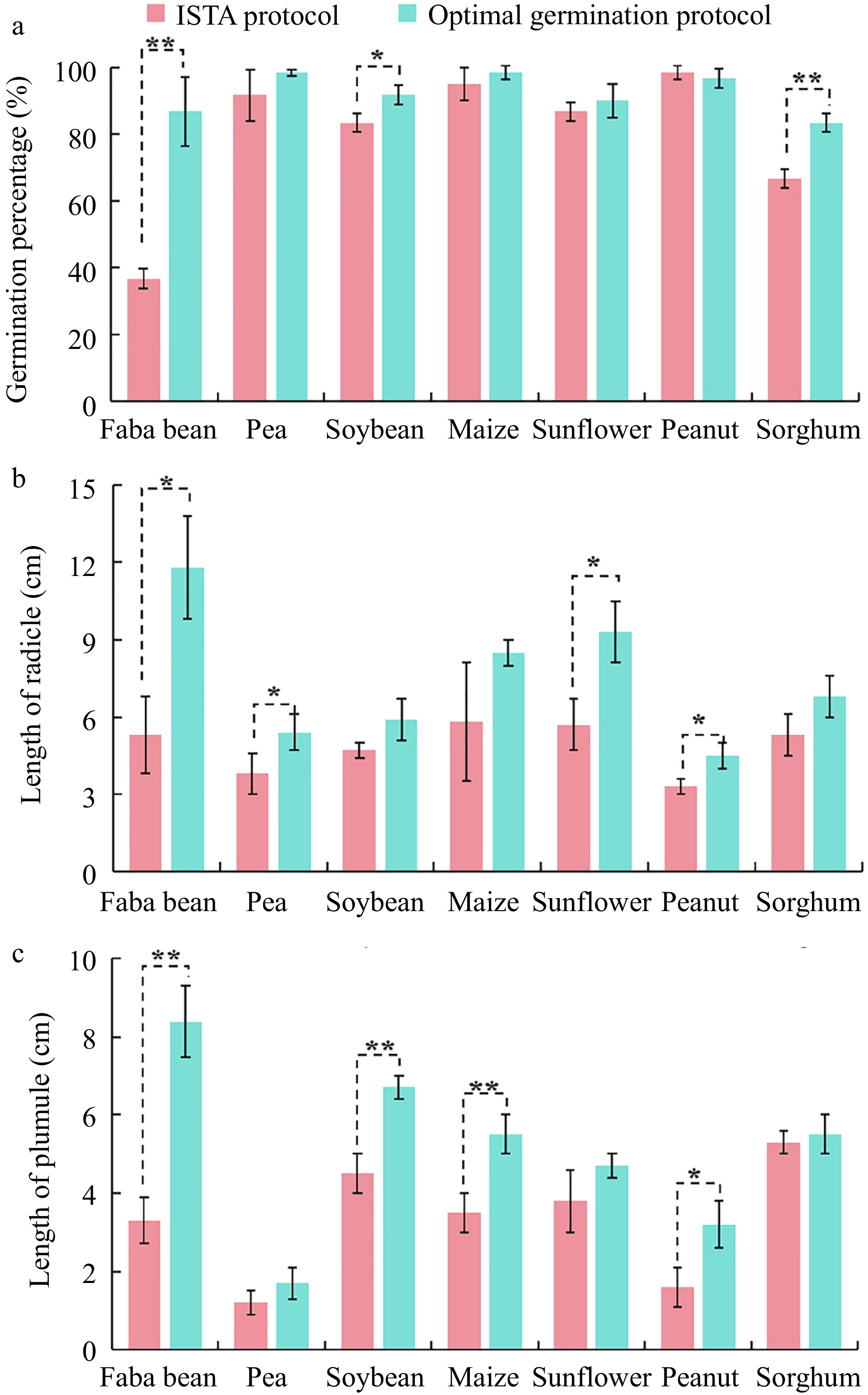
To better demonstrate the advantages of the optimal germination protocol for viability monitoring of seven crop seeds compared to the ISTA protocol, the optimized protocol significantly increased the germination percentages of faba bean ('Tongcanxian 6') and sorghum ('CP21') by 2.36-fold and 1.25-fold, respectively. Additionally, the optimized protocol effectively enhanced the radicle length of the seeds; faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), sunflower ('Dwarf edible sunflower'), and peanut ('Native species') showed increases of 2.23-fold, 1.42-fold, 1.63-fold, and 1.36-fold, respectively. The plumule lengths of faba bean ('Tongcanxian 6'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), and peanut ('Native species') were significantly greater than those of the ISTA protocol, with increases of 2.55-fold, 1.49-fold, 1.57-fold and 2-fold, respectively (Fig. 5). The results of this study confirm that the developed NaClO disinfection protocol can effectively remove fungi from seeds, promoting better growth during seed germination. Furthermore, to assess the general applicability of the optimal germination protocol across different varieties of the seven crop species, various varieties showed effectively reduced fungal infection and improved seed germination index (Supplementary Fig. S2). In conclusion, the results confirm that the optimal germination protocol can effectively eliminate fungi in seeds and improve the accuracy of seed viability monitoring.
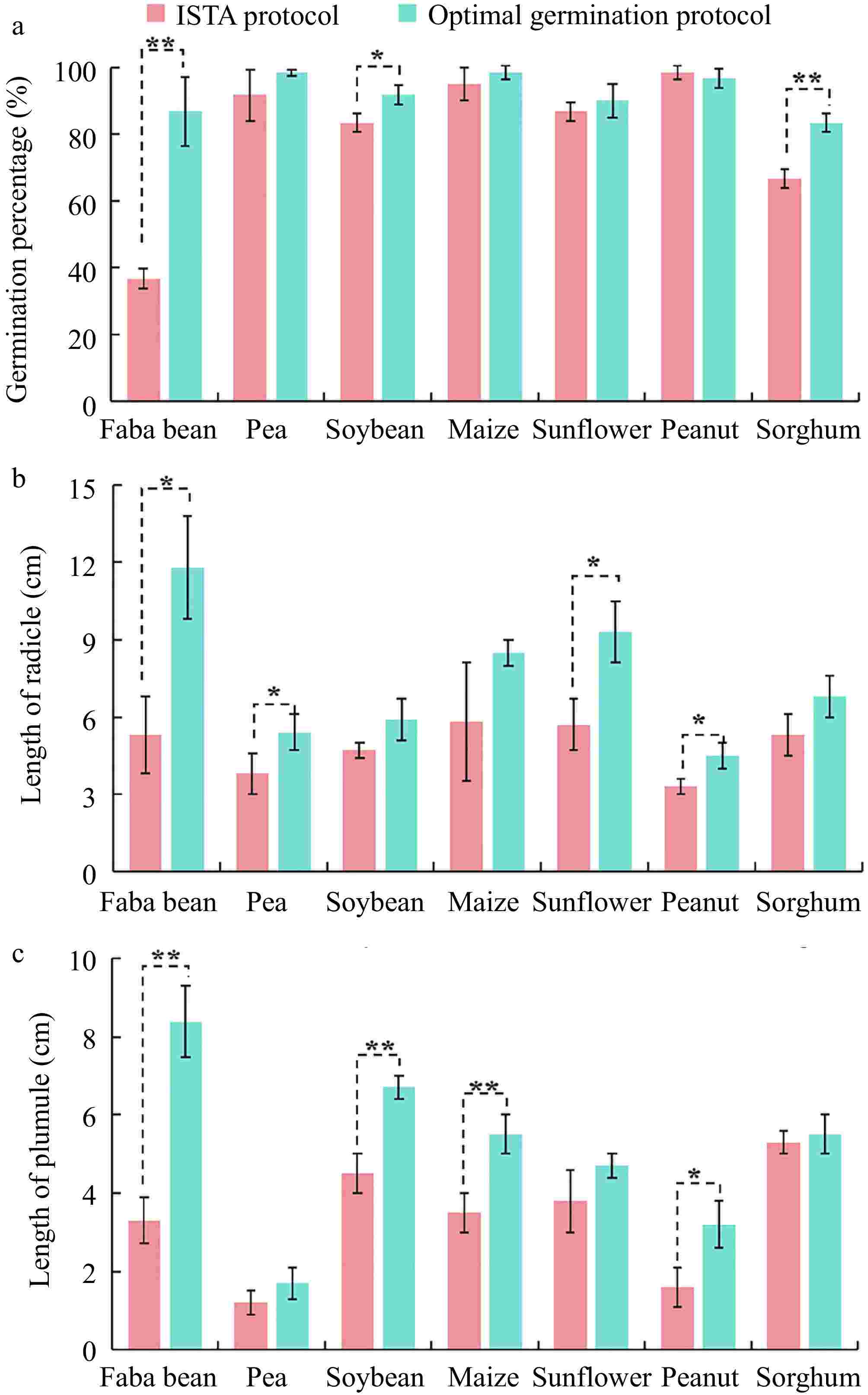
Figure 5.
Comparison of the effect of the optimal disinfection protocol and the ISTA protocol on (a) germination percentage, (b) radicle length, and (c) plumule length of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species'), and sorghum ('CP21') seeds. Values are presented as means ± SD (n = 3). Significant differences were determined using one-way ANOVA (*, p < 0.05; **, p < 0.01).
-
This study found that the accuracy of seed viability evaluation is affected by fungal infection during germination. The genera of fungi in different crop seeds have many commonalities by high-throughput and Sanger sequencing. Fusarium species are important and widely prevalent fungi that inhibit seed germination of all seven crop species. A three-factor, three-level orthogonal experiment was used to optimize suitable germination conditions as follows: faba bean, pea, sunflower, and sorghum seeds were disinfected with 10% NaClO for 35 min; soybean seeds were disinfected with 8% NaClO for 15 min; peanut and maize seeds were disinfected with 6% NaClO for 25 min. It is recommended to culture on a 7 g/L agar medium. The optimal germination protocol could effectively reduce the affection of fungal on seed germination, providing technical support for improving the accuracy of seed viability monitoring in Genebanks.
This work was supported by the Agricultural Science and Technology Innovation Program and the National Natural Science Foundation of China (31860073). We thank Mr. Yixin Lin and Ms. Niwen Tang from the Chinese Academy of Agricultural Sciences for their advice during the experimental design process and Novogene who provided the high-throughput sequencing.
-
The authors confirm contribution to the paper as follows: study conception and project supervision: Yin G, Deng H, Deng Z; study design: Li Y, Wang Y; experiments performing: Xu X, Cheng C; experiments assisting: Xin X, Zhang J; data analysis: Xu X, Cheng C, Xin X, Zhang J; drafted manuscript preparation: Xu X, Cheng C, Li Y, Wang Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its Supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xinyue Xu, Chongyang Cheng
- Supplementary Table S1 The name of eleven varieties in each faba bean, pea, soybean, maize, sunflower, peanut and sorghum species.
- Supplementary Table S2 Fungal genera identified through Sanger sequencing during seed germination of 11 varieties from each species: faba bean, pea, soybean, maize, sunflower, peanut, and sorghum.
- Supplementary Table S3 Comparison and assessment of germination percentage, radicle and plumule length of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species') and sorghum ('CP21') seeds seeds treated with NaClO solutions at concentrations of 6%, 8%, and 10%, disinfection times of 15 min, 25 min, and 35 min, and agar concentrations of 1.75 g/L, 3.5 g/L and 7 g/L in the culture medium, with a pH range of 5.8–6.0. The values are mean ±SD (n = 3). Means in same letter are not significantly different at p < 0.05 according to one-way ANOVA.
- Supplementary Table S4 Fungal genera identified through Sanger sequencing in sorghum ('CP21') seed tissue blocks cultured on potato dextrose agar medium, with optimal and non-optimal disinfection treatments.
- Supplementary Fig. S1 Comparison of epiphytic fungal growth in seeds of faba bean ('Tongcanxian 6'), pea ('Zhongwan 06'), soybean ('Zhonghuang 13'), maize ('Nongkeyu 368'), sunflower ('Dwarf edible sunflower'), peanut ('Native species') and sorghum ('CP21'), with optimal disinfection and non-disinfection treatments, after a few turns of rolling on potato dextrose agar medium.
- Supplementary Fig. S2 Comparison of germination percentage, germination index, and vitality index of faba bean, pea, soybean, maize, sunflower, peanut and sorghum seeds, following germination by the ISTA protocol and the optimal germination protocol. Values are presented as means ± SD (n = 3). Significant differences were determined using one-way ANOVA (*, p < 0.05; **, p < 0.01).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xu X, Cheng C, Li Y, Xin X, Wang Y, et al. 2025. Optimization and analysis of the seed viability monitoring for seven crop species to inhibit fungal infection. Seed Biology 4: e004 doi: 10.48130/seedbio-0025-0004 

Figure 1.
Detection germination percentage and mold incidence percentage in seeds from 11 varieties of each faba bean, pea, soybean, maize, sunflower, peanut, and sorghum, germinated according to the ISTA protocol. Values are presented as means ± SD (n = 3).