
-
When subjected to external mechanical trauma, pathogen invasion, or animal browsing, various tree species secrete complex resinous secondary metabolites that aid in wound repair and regeneration[1,2]. After injury, plants of the Aquilaria genus within the Thymelaeaceae family produce resins containing a variety of chromone and sesquiterpene compounds, which emit a pleasant aroma[3,4]. Furthermore, as these substances permeate and accumulate within the wood, they form a dark brown-to-black material that is denser than water, hence the term 'agarwood', which is also used to describe this specialized wood[5]. In China, the naturally distributed Aquilaria species include Aquilaria sinensis, commonly known as 'white wood agarwood' or 'native agarwood', which is the sole botanical source of traditional Chinese agarwood and is predominantly found in regions such as Hainan, Guangdong, Guangxi, Yunnan, and Fujian. This native agarwood is a precious medicinal plant unique to China, and it is classified as a second-level protected plant in the 'National Key Protected Wild Plant List'[6,7].
In China, the utilization of agarwood has a long history, with the earliest records dating back nearly 2000 years to the Han Dynasty[8]. The unique fragrance produced when agarwood is burned, coupled with its physical property of being denser than water, rendered it a precious fragrance in ancient courts and among literati, leaving behind a rich legacy of poetic and literary works[9]. The application of new technologies has enabled researchers to delve into the medicinal components of agarwood[10]. For example, technologies such as gas chromatography–mass spectrometry (GC–MS) and high-performance liquid chromatography (HPLC) have identified a variety of sesquiterpenes and chromones in agarwood, including agarwood spiralane-type sesquiterpenes and benzene-2-(2-phenylethyl) chromones[11,12]. Among them, 2-(2-phenylethyl) chromone compounds exhibit anti-inflammatory activity, β-caryophyllene possesses antioxidant and antimicrobial effects, and diterpenoid compounds have antidepressant effects[13,14].
Traditional agarwood is mainly produced from natural wood of aquilaria and related trees, that is, farmers search for and collect agarwood produced when agarwood trees are physically or biologically damaged in nature[15]. This method has extremely low yield and requires cutting down trees to obtain the agarwood deep in the trunks, causing great damage to the naturally distributed wild agarwood resources. Furthermore, a variety of physically traumatic methods, such as drilling, cutting, or burning[16], are used to destroy and stimulate trees to produce agarwood. However, these methods result in low agarwood production rates and extended formation periods, failing to meet the socio-economic demands for agarwood[17]. Consequently, agarwood remains a scarce resource, leading to high market prices. In recent years, to satisfy market demands, researchers and producers have invented methods that utilize chemical damage or fungal stimulation to shorten the agarwood formation time and increase production yields[18,19]. Nonetheless, these methods inflict considerable harm on agarwood trees, and the subsequent harvesting process involves the felling of entire trees, resulting in the loss of forest resources. Therefore, research into the mechanisms of agarwood formation, improvement of agarwood production methods, and the cultivation of new varieties are of paramount importance for sustainable development of the agarwood industry[20,21].
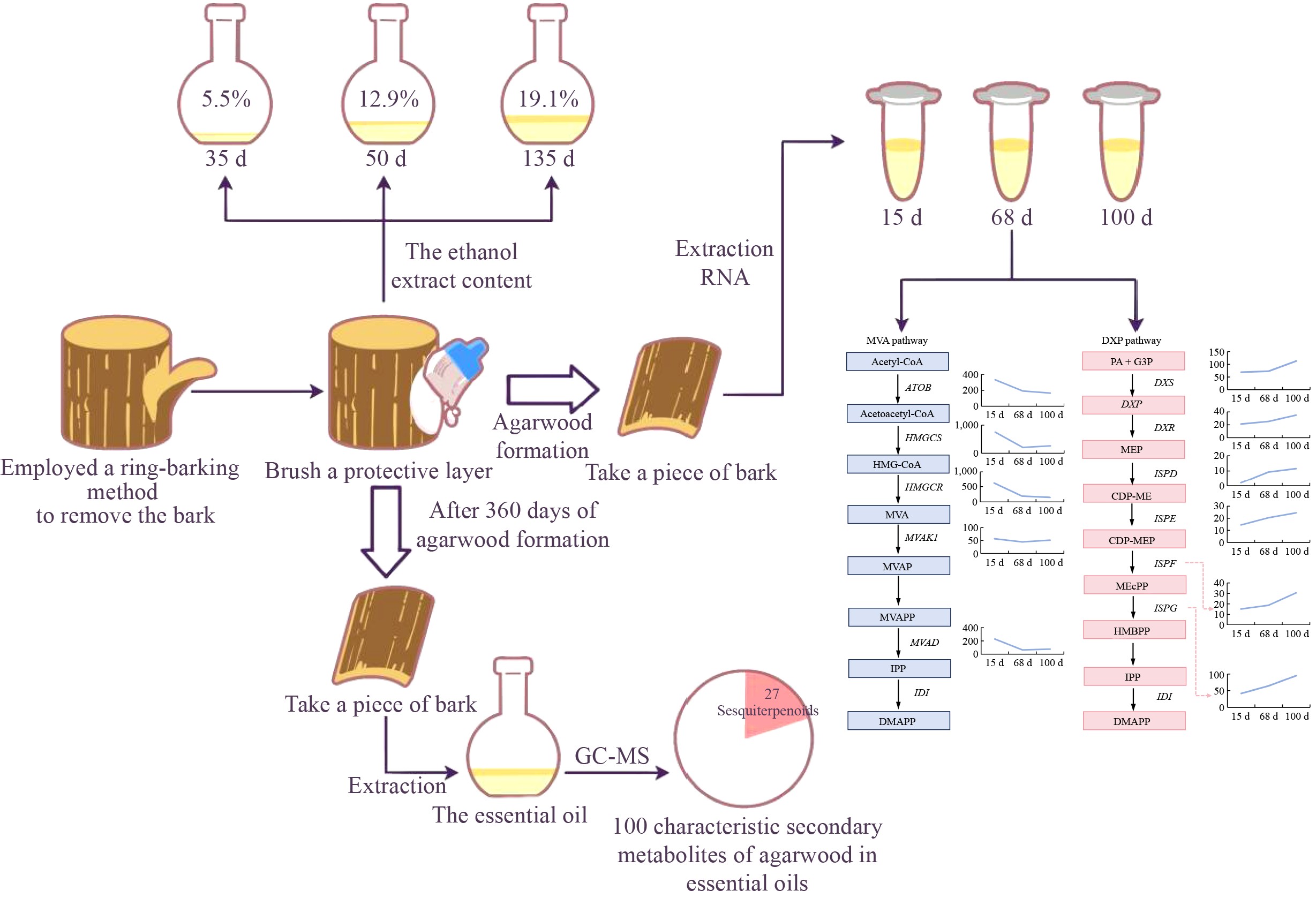
In recent years, significant progress has been made in the molecular mechanisms underlying the formation of agarwood[22−24]. So far, transcriptome sequencing has described the expression of terpene synthesis– or stress response–related genes during the agarwood formation process, but the molecular mechanisms of agarwood formation are not yet fully understood[25]. Notably, 2-(2-phenylethyl) chromones and their derivatives have been identified as key markers in agarwood formation, and their biosynthetic pathways have been hypothesized[26]. Furthermore, high-quality whole-genome analysis of A. sinensis was completed, and transcriptome sequencing analysis related to agarwood formation was also conducted[27,28]. However, the exact gene sequences of enzymes related to the biosynthesis of aroma substances are still unclear. It has been noted that genes related to the 1-deoxy-D-xylulose-5-phosphate (DXP) pathway and the mevalonic acid (MVA) pathway were identified in the A. sinensis genome and actively expressed in the agarwood formation areas[28]. In the present study, a ring-barking method was employed to facilitate the formation of agarwood. This approach involved the removal of the entire bark from A. sinensis trees and the application of a self-formulated silicate-based protective gel to ensure the survival of the agarwood tree. The encumbering process was continuously monitored, analyzing the agarwood formation and the gene expression trends of the regenerating cork layer within 135 d after bark removal. After one year of agarwood formation, the cork layer was collected from the regenerated bark and the essential oil extracted for further compositional analysis. This study provides a novel method for agarwood formation and offers new insights into understanding the agarwood formation mechanisms of A. sinensis.
-
Three 8-year-old Aquilaria sinensis trees grown in the Dongping Farm in Yangjiang Town, Qionghai City, Hainan Province (China), were used in this study. Permission was obtained from the Agricultural Research, Education and Extension Organization of China to collect plant samples. The specimens were stored at the Hainan Key Laboratory for Sustainable Utilization of Tropical Bioresources in Hainan University (specimen numbers 21612-21613).
The experimental materials consisted of 8-year-old Aquilaria sinensis trees with a healthy trunk and with a diameter ranging from 15 to 18 cm at a height of 1.2 m high with three individuals as replicates. The agarwood planting commenced in February 2023, followed by the continuous collection and measurement of the regenerated bark. The planting date was selected based on clear weather conditions, ensuring no rainfall within 3 d after planting. During the planting process, a complete ring of agarwood bark was removed using a ring-barking knife at a height of 10 cm from the ground on the lower part of the trunk and approximately 2 m on the upper part. Subsequently, the bark was entirely stripped between the upper and lower ring-baring sites, and a self-formulated protective gel was applied, composed of 5% glacial acetic acid, 2 g/L sodium silicate, 5 mg/L glycine, 5 mg/L nicotinic acid, 1 mg/L naphthaleneacetic acid, and 1 mg/L indole-3-acetic acid. Fifteen days after planting, a peelable layer of regenerated bark began to form, marking the initiation of ongoing related research. The agarwood harvesting process paralleled the planting procedure, which involved stripping the regenerated bark and applying the protective gel to ensure the survival of the trees.
Determination of agarwood quality
-
The identification of samples was conducted in accordance with the forestry industry standard LY/T 2904—2017 for 'Agarwood'. The determination of ethanol extract content in agarwood was performed as follows: Two g of the pulverized sample was weighed. The sample was placed in a 250-mL conical flask, to which 100 mL of 95% ethanol was added using a pipette, sealed, weighed, and sonicated for 1 h before being allowed to cool. After cooling, the conical flask was removed, and the contents were filtered through filter paper. A 25-mL aliquot of the filtrate was pipetted into a previously dried and weighed evaporation dish, which was then placed in an oven at 65 °C for 3 h to dry. The dish was removed and quickly reweighed. The ethanol extract content was calculated using the following formula, and the measurement was conducted twice. The absolute error between the two determinations should not exceed 0.3%, and the arithmetic mean was taken as the result of the determination. The result was displayed to two decimal places.
Formula:
X(%)=m1−m2mx×(1−w)×400 where, m1 is the mass of the ethanol extract and evaporation dish; m2 is the mass of the evaporation dish; mx is the mass of the sample; and w is the moisture content in the sample. HPLC fingerprinting analysis: A total of 0.2 g of the pulverized agarwood sample was weighed and placed into a capped centrifuge tube. Then, 10 mL of 95% ethanol was added using a pipette, and the tube was weighed and subjected to ultrasonic treatment in a water bath for 1 h. The mixture was allowed to cool, and reweighed. Any weight loss was replenished with 95% ethanol, followed by mixing well and incubation. The supernatant was taken and filtered through a 0.45-μm membrane filter for subsequent analysis. Alternatively, 2 mL of the ethanol extract filtrate was taken, filtered through a 0.45-μm membrane filter, and used as the sample solution.
Chromatographic conditions and system suitability testing were conducted using acetonitrile as mobile phase A and 0.1% formic acid solution as mobile phase B, following the gradient elution program specified in the table below; the flow rate was set at 0.7 mL/min; the column temperature was maintained at 31 °C; and the detection wavelength was 252 nm. The theoretical plate count, calculated based on the peak of agarwood tetrol, should not be less than 6000. Ten μL of the agarwood sample solution was accurately aspirated and injected it into the HPLC system for analysis.
Microstructural analysis of cork
-
Microstructural analysis of the agarwood cork layer involved the following staining procedure[29]: Sections of the agarwood cork layer were obtained which were approximately 5 mm thick and up to 3 cm in maximum length, they were fixed in 70% formaldehyde-acetic acid (FAA) for 48 h. The samples were dehydrated through a graded series of alcohol concentrations (60%, 70%, 80%, 90%, and 95%) for 24–72 h per gradient, followed by 100% (v/v) for 12-24 h, 100% isopropanol (I) for 12-24 h, and dimethylbenzene for 8–24 h. The samples were infiltrated with infiltration solution I at 4 °C for 24–72 h, infiltration solution II at 4 °C for 24–72 h, and infiltration solution III at 4 °C for 24–72 h. The samples were placed in embedding capsules. An appropriate amount of embedding medium was added, and the samples were vacuum-infiltrated for 5 h and polymerized in a 37 °C water bath. The samples were sectioned using a hard tissue microtome to a thickness of 10 μm, and the slides were baked at 60 °C overnight. The deparaffinization process of the sections is as follows: First, immerse the sections in Eosin Y Xylene I for 6 h at a temperature maintained at 37 °C; then, left overnight in Eosin Y Xylene II, also at 37 °C; next, the sections were treated in Eosin Y Xylene III at room temperature for 10 to 15 min, followed by another 10 to 15 min at room temperature in Eosin Y Xylene IV. Afterward, the sections were sequentially immersed in 100% Ethanol I, 100% Ethanol II, 95% Ethanol, 90% Ethanol, and 80% Ethanol for 10 min each, and finally rinse thoroughly with tap water. For toluidine blue staining, the sample sections were stained in the dye solution for about 2 min, then rinsed with water, and examined under a microscope. Appropriate differentiation or no differentiation were performed based on the depth of staining, rinsed with tap water, the slides were then dried in a 60 °C oven. The sections were clarified in clean dimethylbenzene for 5 min and mounted with neutral resin. The sections were examined under a light microscope, and image capture and analysis were performed.
RNA extraction and cDNA preparation from the regenerating bark of A. sinensis
-
At three distinct stages after implantation, namely at 15, 68, and 100 d, samples of A. sinensis bark were rapidly frozen using liquid nitrogen and stored at −80 °C. Subsequently, the cork layer was ground into a fine powder using liquid nitrogen and subjected to RNA extraction. The integrity of the nucleic acid samples was assessed through agarose gel electrophoresis, followed by the construction of transcriptome libraries. Utilizing transcriptome sequencing technology, nine A. sinensis samples, with three replicates at each of the three stages (15, 68, and 100 d after implantation), underwent paired-end transcriptome sequencing, yielding raw sequencing data. The proportion of clean data obtained after filtering the raw data from each sample was above 99.2%, while the percentages of low-quality data, reads with a proportion of N bases greater than 10%, reads consisting entirely of A bases, and adapters were all at very low levels, meeting the requirements for analysis. The percentage of base quality reaching Q20 and above, and Q30 and above was greater than 97% and 93%, respectively, for the filtered samples. The GC content ranged between 45% and 48%. Additionally, the balanced base distribution indicated high data quality, suitable for subsequent data analysis.
cDNA was synthesized from RNA using the FastKing RT Kit in accordance with the manufacturer's instructions and used for real-time fluorescent quantitative analysis. Gene expression analyses were performed using BMKCloud (
www.biocloud.net ).Supercritical carbon dioxide extraction
-
Twenty kg of dried samples and pulverized bark were loaded it into an extraction vessel. The flow rate of CO2 was controlled at 20 L/h, with an extraction pressure of 25 MPa and an extraction temperature of 40 °C. The extraction duration was set at 2 h, after which the sample was collected[30].
GC-MS analysis
-
The mass of the sample for analysis was 1 g. The chromatographic column used was an HP-5ms Ultra Inert (30 m × 250 μm × 0.25 μm). The temperature program was initiated at 50 °C and was held for 0 min, then ramped up at a rate of 5 °C/min to 310 °C, where it was maintained for 10 min. The injection port temperature was maintained at 250 °C, with an injection volume of 1 μL using a non-split injection method. The mass spectrometry conditions involved an electron ionization ion source with an electron energy of 70 eV. The ion source temperature was set at 230 °C, and the MS detector transfer line temperature was maintained at 280 °C.
-
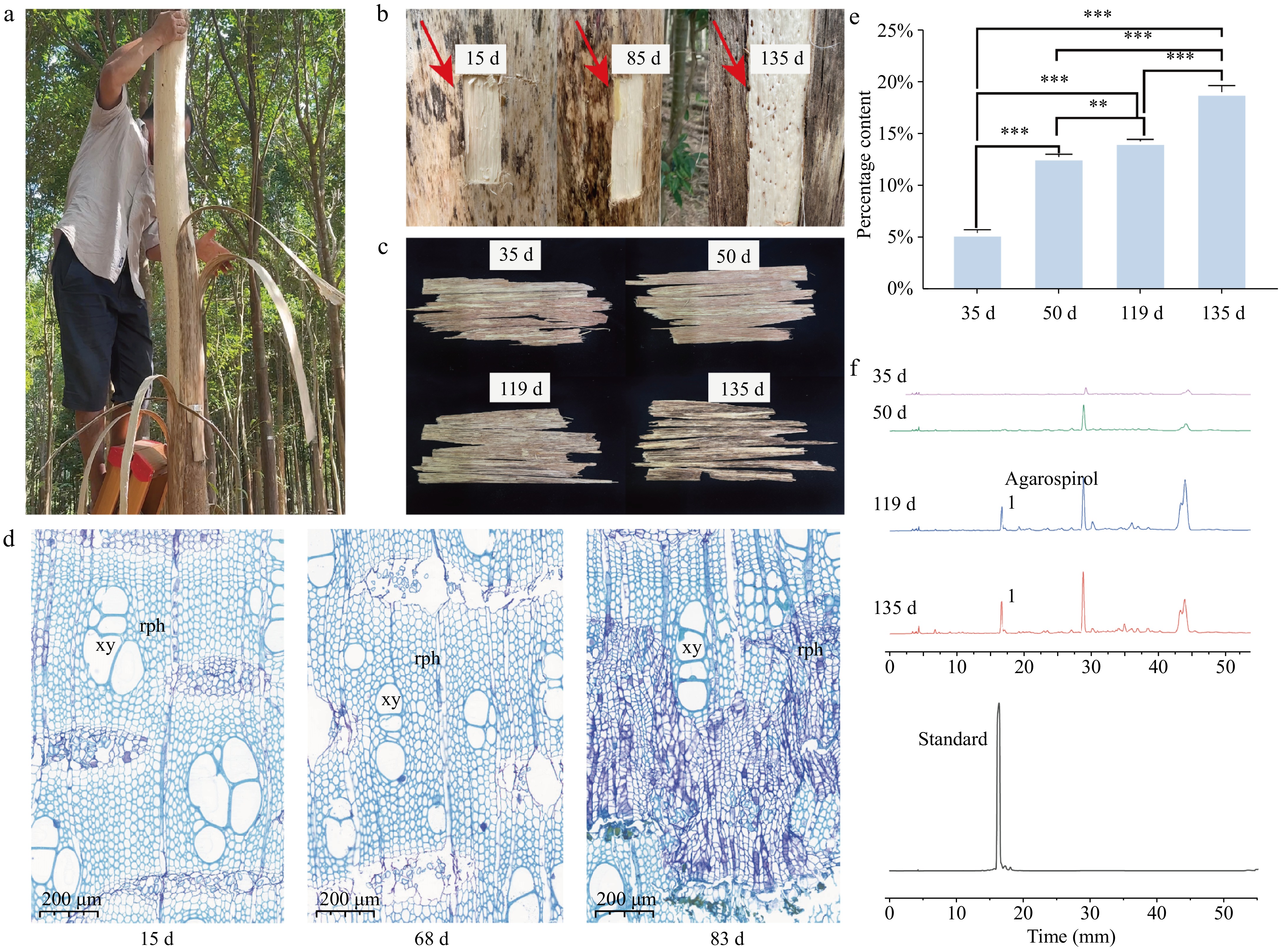
In this study, A. sinensis trees were treated using the bark-ringing method to induce agarwood formation (Fig. 1a). Bark regeneration samples were successfully obtained at different stages of the planting process. A distinct color change was observed in the newly formed cork layer at the junction between the trunk and the bark in the agarwood formation area. Upon heating, this area emitted the characteristic aromatic scent of agarwood (Fig. 1b, indicated by the arrow). The color change in the cork layer of the regenerating bark was slower at 35 and 50 d after planting, but by 119 d after planting, the wood at the agarwood formation site turned into a deep yellow-brown color, and by 135 d after planting, the color deepened to dark brown (Fig. 1c). To track and understand the bark regeneration process of A. sinensis following the bark-ringing method, regenerating bark samples were collected at 15, 68, and 83 d after planting for sectioning, staining, and microscopic observation (Fig. 1d). Fifteen days after bark removal, the bark began to regenerate, and an increase in the number of phloem vessels was observed in the regenerating cork layer. By 83 d, the phloem vessels had developed extensive secretory cells. Furthermore, the regenerating bark samples from 35 d after planting were processed using ethanol extraction. The ethanol extract content increased with the duration of the planting process (Fig. 1e). At 35 d after planting, the extract yield was 5.5%. The extract yield at 50, 119, and 135 d after planting was 12.9%, 14.3%, and 19.1%, respectively. HPLC analysis revealed that the characteristic peak of agarwood tetrol in the samples gradually increased over time, and the chromatograms at 119 and 135 d after planting exhibited increased complexity, indicating a growing diversity of compounds (Fig. 1f).
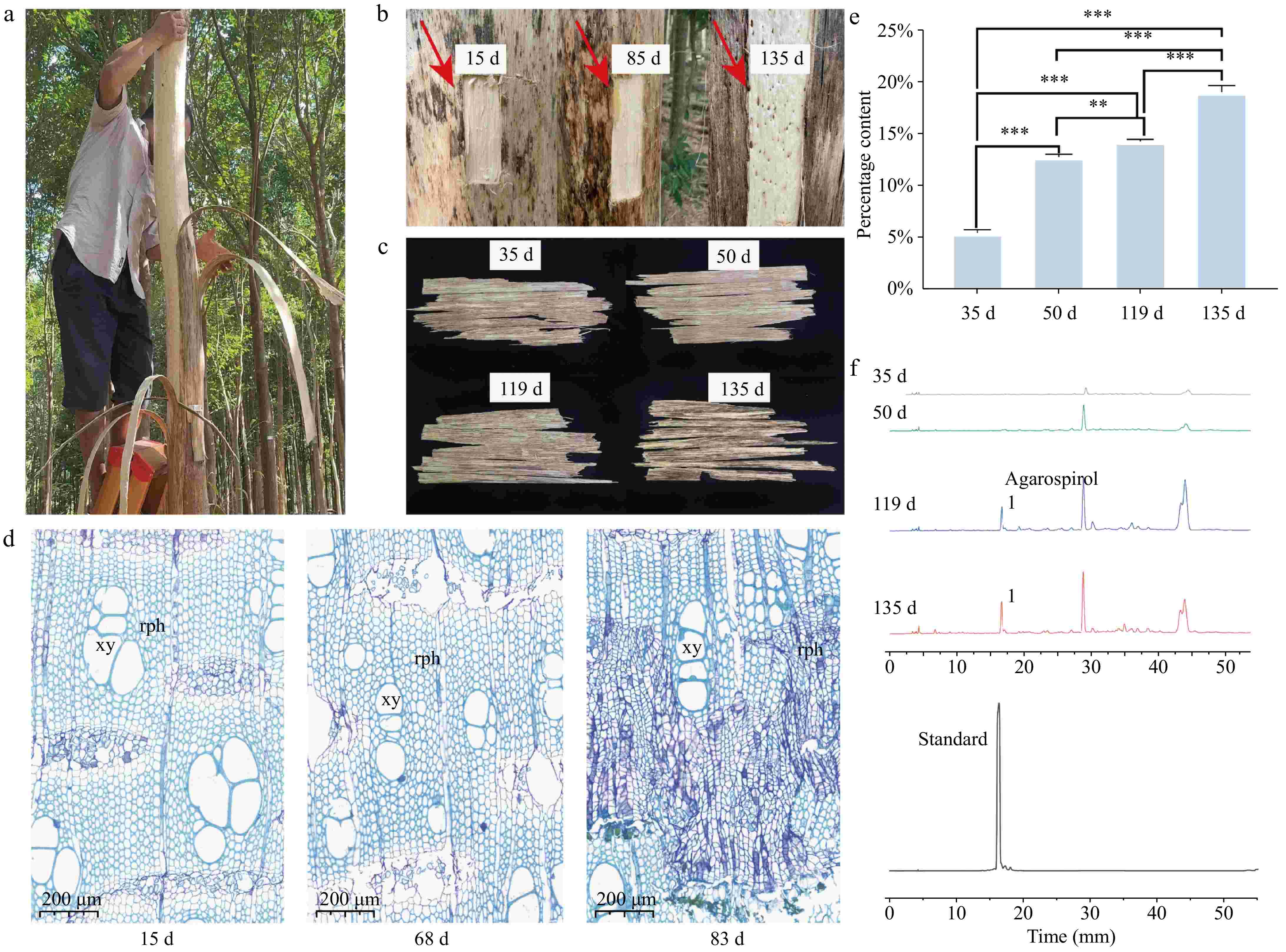
Figure 1.
Cambial regeneration process after ring-barking for agarwood formation. (a) On-site bark ring-barking of agarwood; (b) the trunk sections with regenerated phloem layers removed at 15, 85, and 135 d after bark removal; (c) samples of the regenerated phloem layers at 35, 50, 119, and 135 d after bark removal; (d) abbreviations: rph - regenerated phloem; xy - xylem vessels; (e) bar chart of ethanol extract content: changes from 35 to 135 d. * Indicates p < 0.05, ** indicates p < 0.01, *** indicates p < 0.001. (f) High-performance liquid chromatography (HPLC) characteristic peak chart.
Identification of differentially expressed genes
-
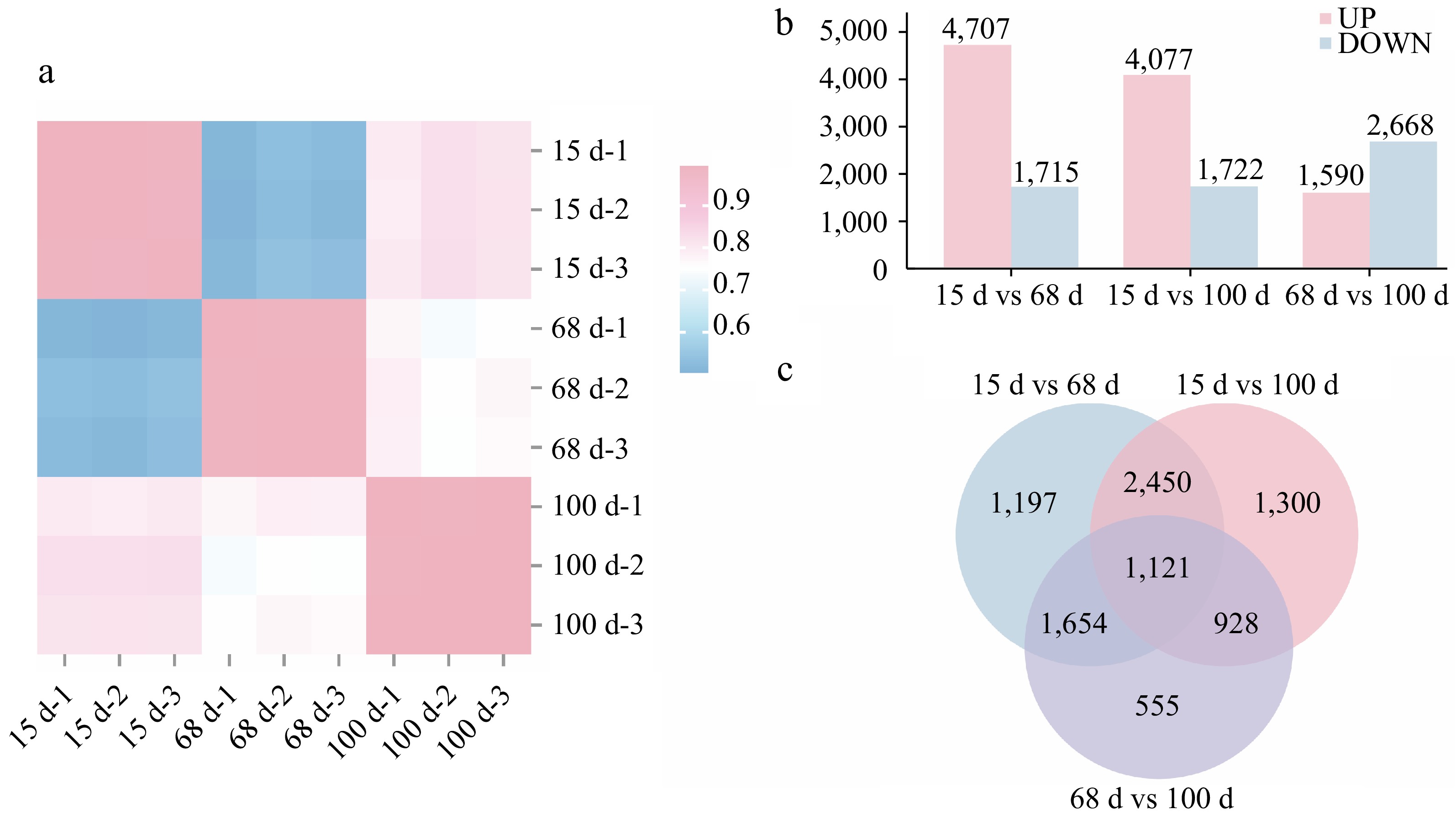
To further investigate the molecular mechanisms underlying agarwood formation, RNA samples were extracted from the continuously sampled materials as shown in Fig. 1d, and transcriptome sequencing analysis conducted. Upon examination, the RNA extraction quality and the base quality of the samples met the sequencing standards. The results of Pearson correlation analysis indicated that for the three groups of replicate samples at different stages after planting (15, 68, and 100 d), the correlation coefficients among samples within each group were all greater than 0.99 (Fig. 2a). Differentially expressed genes between different samples were also identified (Fig. 2b). There were 4,707 upregulated Unigenes and 1,715 downregulated Unigenes between 15 and 68 d after planting. There were 4,077 upregulated Unigenes and 1,722 downregulated Unigenes between 15 and 100 d after planting. Between 68 and 100 d after planting, 1,590 Unigenes were upregulated and 2,668 Unigenes were downregulated. The results revealed a higher number of differentially expressed Unigenes between 15 and 68 d after planting, suggesting that many pathways and Unigenes were involved in the agarwood formation process during this stage. The Venn diagram of the differentially expressed gene sets showed that there were 1,121 common differentially expressed genes among the three sample sets (Fig. 2c).
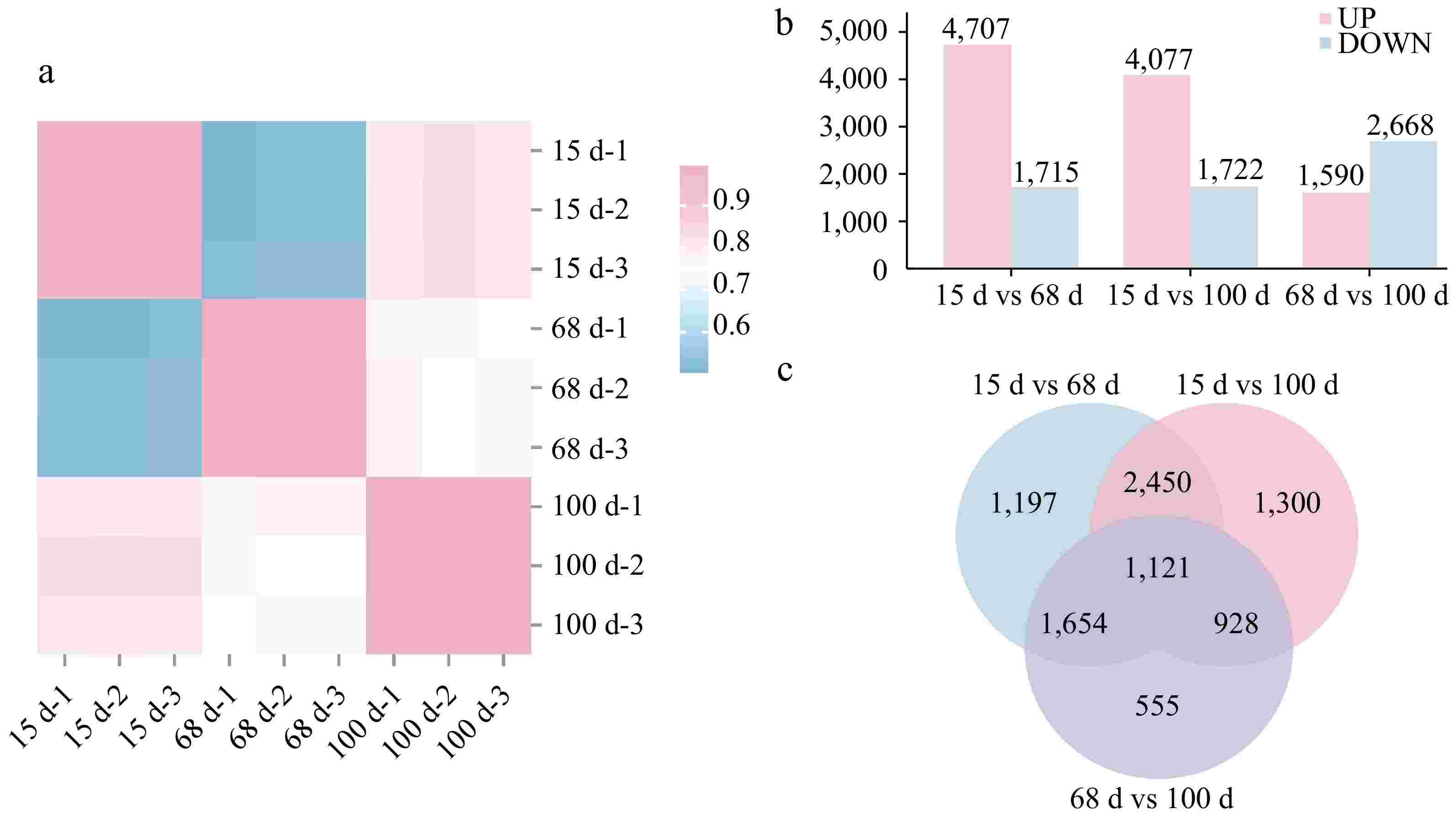
Figure 2.
Differential gene expression among samples at 15, 68, and 100 d after agarwood formation. (a) Pearson correlation heat map for three different stages at 15, 68, and 100 d after implantation, with three replicates, totaling nine white Aquilaria samples; (b) bar chart of differentially expressed genes at three different stages after implantation. UP indicates the number of upregulated genes, DOWN indicates the number of downregulated genes; (c) Venn diagram of the differentially expressed genes at three different stages after implantation at 15, 68, and 100 d.
Validation by quantitative fluorescent detection
-
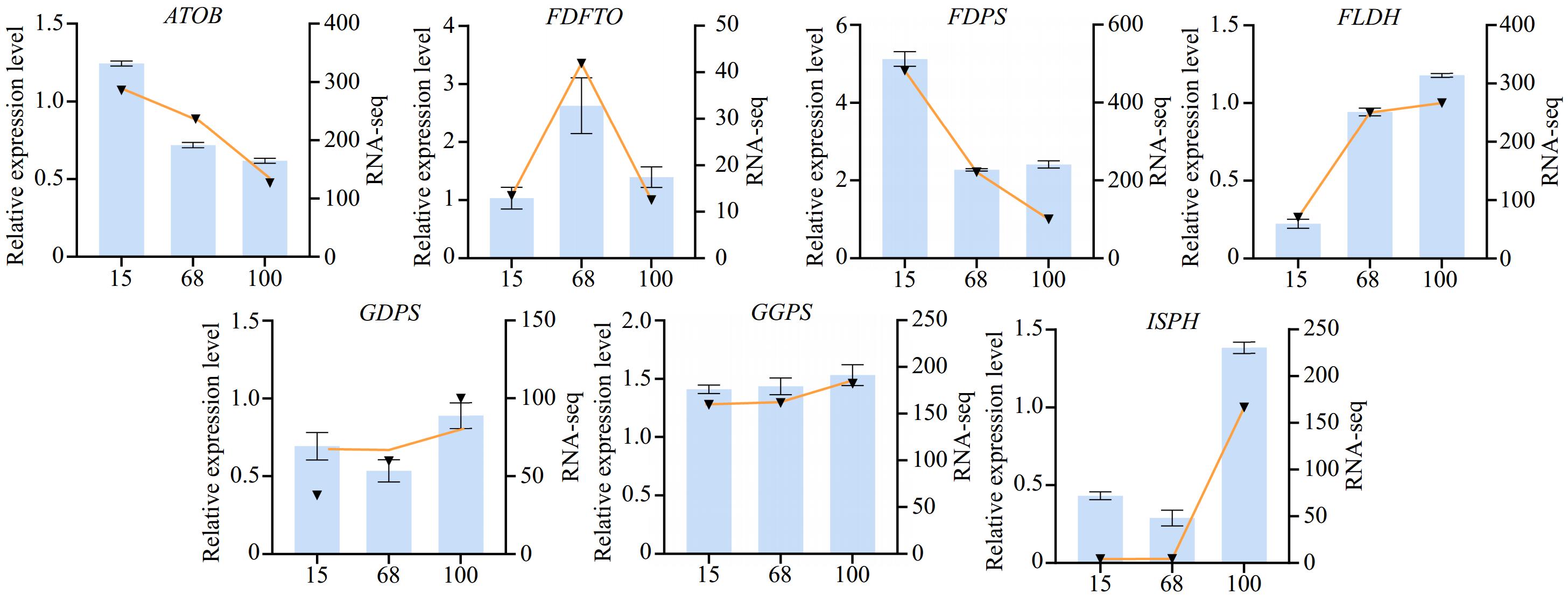
During the agarwood formation process, to gain a deeper understanding of the expression patterns of the differentially expressed genes at various stages, quantitative real-time polymerase chain reaction (qPCR) was employed to validate the selected genes. These genes were screened from transcriptome data and were considered to play a key role at different stages of agarwood formation. Samples were collected at three stages of plant agarwood formation (15, 68, and 100 d), and their total RNA was extracted and reverse-transcribed into cDNA. Subsequently, using quantitative fluorescent PCR technology, differentially expressed genes that had been determined in advance were analyzed for quantification. These genes are involved in key steps of the agarwood biosynthesis pathway, including the synthesis, transport, and metabolism of aromatic compounds. For example, acetyl-CoA C-acetyltransferase (ATOB) is involved in the transfer of acetyl-CoA and catalyzes the production of acetoacetyl-CoA by acetyl-CoA. Furthermore, geranylgeranyl diphosphate synthase (GGPS) is an important terpenoid synthetase, which can catalyze the binding of isopentenyl pyrophosphate (IPP) and dimethylallyl pyrophosphate (DMAPP). It produces geranyl diphosphate and releases pyrophosphate (PPi). The experimental results showed that the expression patterns of most genes were consistent with those obtained from previous transcriptome sequencing data, further confirming the accuracy of the transcriptome data (Fig. 3).
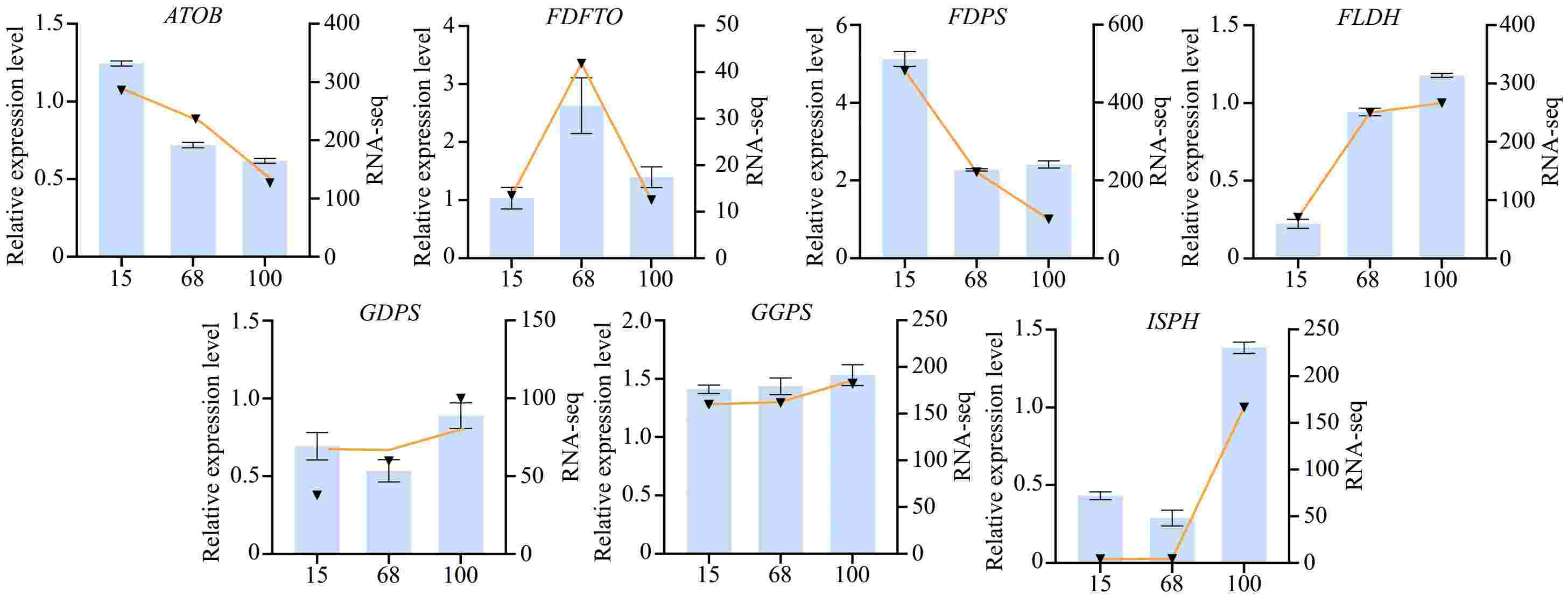
Figure 3.
Expression levels of differential agarwood genes at various stages of formation. The bar chart represents the transcriptomic expression levels, while the line chart represents the fluorescence quantitative expression levels. Gene symbols: ATOB: acetyl-CoA C-acetyltransferase; FDFTO: farnesyl-diphosphate farnesyltransferase; FDPS: farnesyl-diphosphate synthase; FLDH: NAD+-dependent farnesol dehydrogenase; GDPS: geranyl-diphosphate synthase; GGPS: geranylgeranyl diphosphate synthase; ISPH: 4-hydroxy-3-methylbut-2-en-1-yI diphosphate reductase.
Expression of genes related to sesquiterpene biosynthesis pathways at different stages of agarwood formation
-
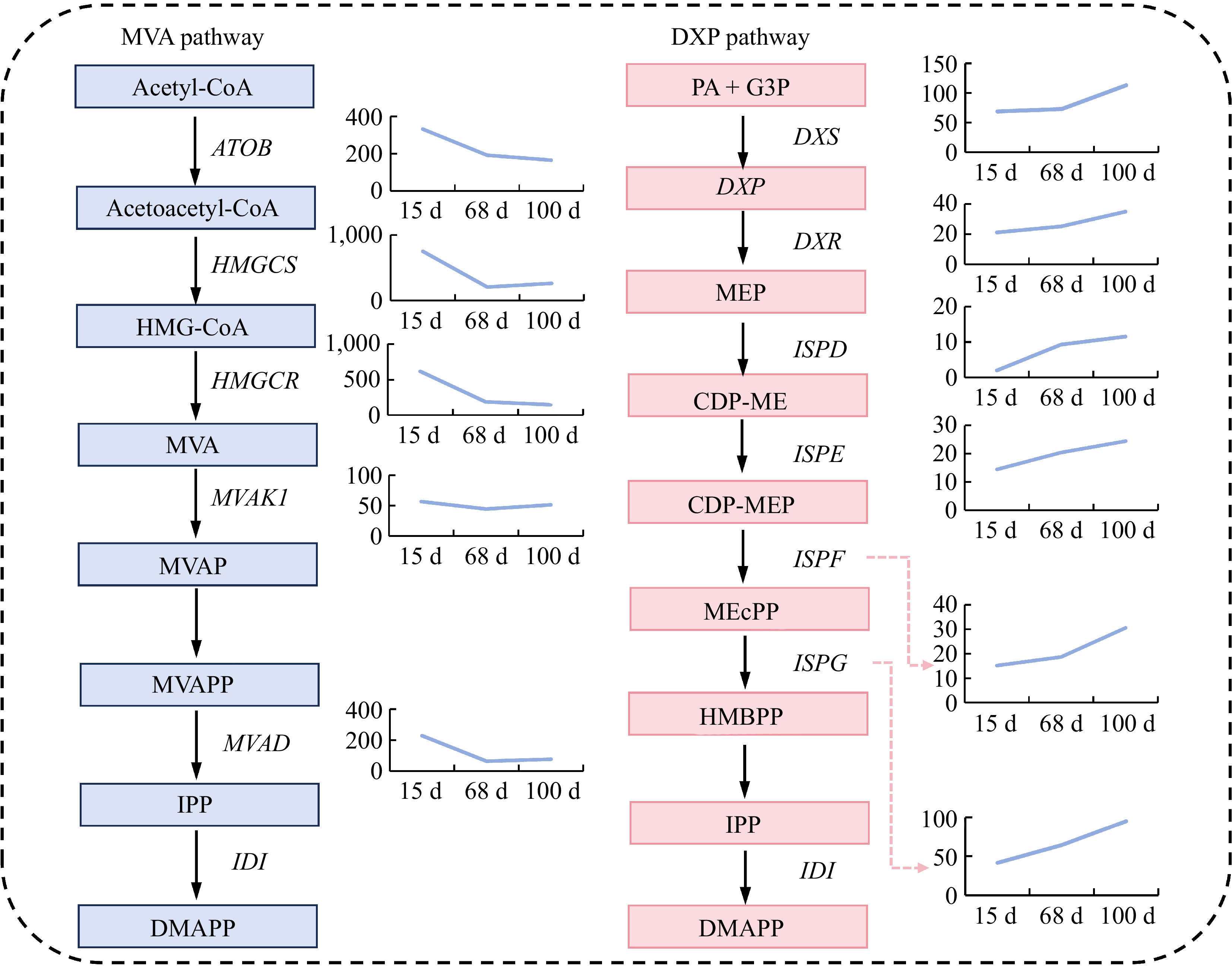
To further analyze the expression trends of the key genes in sesquiterpene synthesis during different stages of agarwood formation, this study focused on the MVA pathway and the DXP pathway within sesquiterpene biosynthesis. The results revealed that the expression levels of the differentially expressed genes involved in the MVA pathway showed a declining trend, with significant decreases in the expression of ATOB, HMGCS, HMGCR, and MVAD genes from 15 to 68 d after planting (Fig. 4). However, there were no significant changes in expression from 68 to 100 d after planting, although the MVAK1 gene exhibited a downward trend, albeit not significant, from 15 to 68 d after planting. As for the DXP pathway, the expression levels of the differentially expressed genes all showed an increasing trend. Specifically, the DXS, DXR, and ISPF genes exhibited significant upregulation from 68 to 100 d after planting, while the ISPE and ISPG genes demonstrated a stable upward trend in expression from 15 to 100 d after planting. The expression level of the ISPD gene showed a marked increase from 15 to 68 d after planting, and a gradual upward trend in expression from 68 to 100 d after planting (Fig. 4).
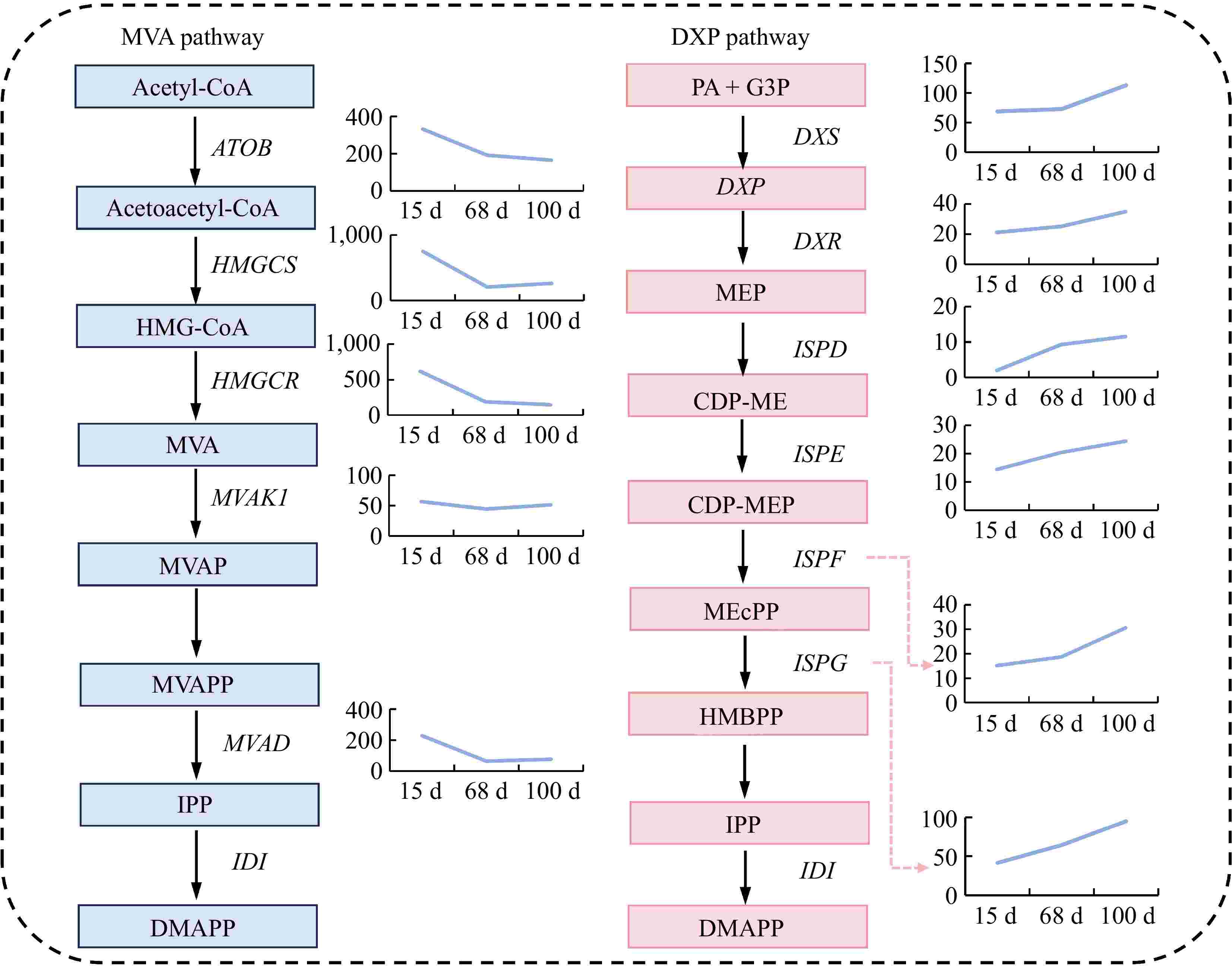
Figure 4.
Expression of differential genes in the sesquiterpene biosynthetic pathway. Abbreviations: Acetyl-CoA: acetyl-CoA; Acetoacetyl-CoA: acetoacetyl-CoA; HMG-CoA: 3-hydroxy-3-methylglutaryl-CoA; MVA: mevalonic acid; MVAP: phosphomevalonic acid; MVAPP: mevalonate diphosphate; IPP: isopentenyl diphosphate; DMAPP: dimethylallyl diphosphate; PA + G3P: pyruvic acid + glyceraldehyde 3-phosphate; DXP: 1-deoxy-D-xylulose 5-phosphate; MEP: 2-C-methyl-D-erythritol 4-phosphate; CDP-ME: cytidine diphosphate-2-C-methyl-D-erythritol; CDP-MEP: cytidine diphosphate-2-C-methyl-D-erythritol 2-phosphate; MEcPP: 2-C-methyl-D-erythritol 2,4-cyclodiphosphate; HMBPP: 1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate. ATOB: acetyl-CoA C-acetyltransferase; HMGCS: hydroxymethylglutaryl-CoA synthase; HMGCR: hydroxymethylglutaryl-CoA reductase (NADPH); MVAK1: mevalonate kinase; MVAD: diphosphomevalonate decarboxylase; IDI: isopentenyl-diphosphate Delta-isomerase; DXS: 1-deoxy-D-xylulose-5-phosphate synthase; DXR: 1-deoxy-D-xylulose-5-phosphate reductoisomerase; ISPD: 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase; ISPE: 4-diphosphocytidyl-2-C-methyl-D-erythritol kinase; ISPF: 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase; ISPG: (E)-4-hydroxy-3-methylbut-2-enyl- di phosphate synthase.
Detection of secondary metabolites in essential oil of agarwood
-
To further understand the quality of agarwood obtained by the bark-ringing method used in this study, samples of the regenerated cork layers were collected 360 d after agarwood formation. The agarwood essential oil was extracted using the supercritical carbon dioxide method. GC–MS analysis detected a total of 100 volatile components in the white wood agarwood essential oil sample, with their retention times, names, and relative percentage content are presented in Table 1. Notably, the sample contained 27 types of sesquiterpene components (Table 2).
Table 1. Chemical composition and relative content of agarwood essential oil.
No. Retention time (min) Name CAS Relative content 1 6.296 Benzaldehyde 100-52-7 0.61 2 13.645 Benzaldehyde, 4-(1-methylethyl)- 122-03-2 1.25 3 21.409 Isoaromadendrene epoxide 1000159-36- 6 0.06 4 21.885 Cyclohexane, 1-butenylidene- 36144-40-8 0.08 5 22.14 Bicyclo[2.2.1]heptane-2,5-diol, 1,7,7-trimethyl-, (2-endo,5-exo)- 10359-41- 8 0.14 6 23.066 Alloaromadendrene 10359-41-8 0.1 7 23.125 (+)-2-Carene, 2-acetyl 1000151- 09-7 0.07 8 23.321 .beta.-Clovene 1000163- 00-2 0.06 9 23.423 .alpha.-Maaliene 489-28-1 0.16 10 23.499 .beta.-Panasinsene 1000159- 39-0 0.07 11 23.61 cis-Thujopsene 470-40-6 0.45 12 23.797 Agarospirol 1460-73-7 1.08 13 23.899 Patchoulene 1405-16-9 0.07 14 24.162 Aromadendrene oxide-(1) 1000151- 98-4 0.43 15 24.23 Arctiol 36061-11- 7 0.38 16 24.323 3,5,11-Eudesmatriene 193615- 07-5 0.06 17 24.476 Guaiol 489-86-1 0.15 18 24.587 .gamma.-Gurjunenepoxide-(1) 1000156- 15-0 0.33 19 24.697 Longipinocarvone 1000151- 87-1 0.31 20 24.782 .alpha.-Guaiene 3691/12/1 0.07 21 24.884 Caryophyllene 87-44-5 0.36 22 25.224 Tetrahydrofuran-2-ol, 3,4-di[1-butenyl]- 1000131-84- 0 0.72 23 25.377 cis-Z-.alpha.-Bisabolene epoxide 1000131- 71-2 0.37 24 25.666 .beta.-Oplopenone 28305-60- 4 0.09 25 25.734 13-nor-Eremophil-1(10)-en-11-one 54275-21- 7 0.09 26 25.946 Spiro[adamantane-2,5'-[1.2]dioxolan]-3'-one, 4'-methylene- 126255- 68-3 0.73 27 26.056 Nerolidol 142-50-7 0.37 28 26.15 Isospathulenol 88395-46- 4 0.06 29 26.252 .beta.-Humulene 116-04-1 0.12 30 26.379 .alpha.-Cadinol 481-34-5 0.85 31 26.456 Dehydroxy-isocalamendiol 1005276- 30-1 0.19 32 26.558 Selin-6-en-4.alpha.-ol 118173- 08-3 0.33 33 26.617 1,4,4,7a-Tetramethyl-2,4,5,6,7,7a-hexahydro-1H-indene-1,7-diol 121747- 53-3 0.12 34 26.804 9-Tetradecenal, (Z)- 53939-27- 8 0.07 35 26.872 Neointermedeol 5945-72-2 0.3 36 27.059 Hexadecahydro-benzo[de]anthracene 1000371- 48-1 0.56 37 27.161 E,E-6,8-Tridecadien-2-ol, acetate 1000131- 35-4 0.68 38 27.203 Isolongifolol 1139-17-9 0.43 39 27.331 1-Bromo-3-(2-bromoethyl)-nonane 70928-49- 3 0.39 40 27.552 2,6,10-Dodecatrienal, 3,7,11-trimethyl-, (E,E)- 502-67-0 0.88 41 27.704 Nootkatone 4674-50-4 5.63 42 27.823 .beta.-Santalol 77-42-9 0.56 43 28.019 1,1,4,7-Tetramethyldecahydro-1H-cyclopropa[e]azulene-4,7-diol 1212211- 43-2 0.34 44 28.121 Arteannuin b 1000212- 12-6 0.9 45 28.299 Isoshyobunone 1000360- 30-1 0.21 46 28.444 1,8-Cyclopentadecadiyne 4722-42-3 0.75 47 28.563 cis-Z-.alpha.-Bisabolene epoxide 1000131- 71-2 0.51 48 28.656 Valerenol 101628- 22-2 2.76 49 28.775 1,8,11,14-Heptadecatetraene, (Z,Z,Z)- 10482-53- 8 0.79 50 28.834 Isobutyl (5-isopropyl-2-methylphenyl) carbonate 1000378- 27-3 0.25 51 28.936 7-(2-Hydroxypropan-2-yl)-1,4a-dimethyldecahydronaphthalen-1-ol 92857-25- 5 0.72 52 29.072 Nootkaton-11,12-epoxide 1000374- 20-3 0.17 53 29.183 Nerolidyl acetate 2306-78-7 0.38 54 29.429 6-(p-Tolyl)-2-methyl-2-heptenol, trans- 39599-18- 3 2.58 55 29.59 Humulenol-II 19888-00- 7 0.78 56 29.76 Silphiperfol-6-en-5-one 20405-58- 7 1.2 57 29.905 L-Glutamine, N-ethoxycarbonyl-, methyl ester 1000452- 41-9 0.1 58 29.973 6-Decen-4-yn-3-ol, 2-methyl-9-(2-methoxyethoxymethoxy)-, (E)- 1000156- 83-6 0.24 59 30.1 Bohlmann k2631 80367-94- 8 4.55 60 30.194 (3R,3aR,4aS,5R,9aS)-3,5,8-Trimethyl-3a,4,4a,5,6,7,9,9a-octahydroazuleno[6,5-b]furan-2(3H)-one 66873-38- 9 1.3 61 30.279 1-(1-Adamantyl)-1-phenylethanol 303797- 64-0 0.68 62 30.372 Valerenic acid 3569/10/6 0.87 63 30.482 Ambrosin 509-93-3 3.08 64 30.567 Isoaromadendrene epoxide 1000159- 36-6 1.99 65 30.729 Acetate, 2-cyclohexenyl-3-[1-(2-oxopropyl)ethenyl]-2,4,4-trimethyl 1000196- 78-1 0.97 66 30.831 Benzene, 1-isocyanato-2-methoxy- 700-87-8 0.2 67 30.941 Valerenic acid, ethyl ester 1000453- 13-8 0.38 68 31.001 Oxacyclotetradeca-4,11-diyne 6568-32-7 0.29 69 31.145 Octanoic acid, 1-ethenyl-1,5-dimethyl-4-hexenyl ester 10024-64- 3 0.19 70 31.298 Estr-4-en-3-one 4811-77-2 0.67 71 31.434 Zederone 7727-79-9 0.68 72 31.527 Spirafolide 130838- 06-1 0.36 73 31.587 3-(Benzyloxymethyl)hex-5-ene-1,2-diol 1000192- 76-5 1.08 74 31.689 7,10,13-Hexadecatrienoic acid, methyl ester 56554-30- 4 0.49 75 31.893 1,8-Cyclopentadecadiyne 4722-42-3 0.07 76 32.012 6-(1-Hydroxymethylvinyl)-4,8a-dimethyl-3,5,6,7,8,8a-hexahydro-1H-naphthalen-2-one 1000190- 51-4 0.78 77 32.122 2,9-Heptadecadiene-4,6-diyn-8-ol, (Z,E)- 50816-77- 8 0.23 78 32.216 Z,Z,Z-4,6,9-Nonadecatriene 1000131- 11-7 0.28 79 32.309 trans-Valerenyl acetate 101527- 74-6 0.35 80 32.7 Benzene, 1,1'-(1,6-hexanediyl)bis- 1087-49-6 0.17 81 33.176 Cedran-diol, 8S,13- 8S, 13- 88588-48- 1 0.1 82 33.286 7-Benzofuranol, 2,3-dihydro-2,2-dimethyl- 1563-38-8 0.16 83 33.405 9-Octadecenal, (Z)- 2423/10/1 1.53 84 33.906 1-Pentyn-3-ol, 1,4-diphenyl- 1000162- 87-4 0.38 85 34.441 Picrotoxin 124-87-8 0.12 86 34.509 1,1'-Butadiynylenedicyclohexanol 5768/10/5 0.14 87 35.282 Reynosin 28254-53- 7 0.23 88 36.098 1,1-Diacetoxy-9,9-diformylnona-2,4,6,8-tetraene 104970- 92-5 0.05 89 36.812 8-Naphthol, 1-(benzyloxy)- 326875- 68-7 17.43 90 38.103 Neoisolongifolene-8-ol 1000159- 36-9 0.06 91 39.224 1-Heneicosanol 15594-90- 8 0.23 92 40.116 Phthalic acid, propyl 6-ethyl-3-octyl ester 1000315- 17-3 0.11 93 40.473 2-(2'-Methoxyphenethyl)chromone 1000422- 88-7 0.15 94 40.864 7-Methoxy-2-phenethylchromone 1000422- 88-8 2.83 95 40.991 Acetamide, 2-methoxy-2-phenyl-N-[2-(4-chlorophenyl)ethyl]- 327991- 45-7 9.12 96 41.509 8-Methoxy-2-phenethylchromone 1000422- 89-0 0.12 97 43.497 Fumaric acid, 2,4,4-trimethylpentyl dodec-2-en-1-yl ester 1000405- 61-4 12.16 98 43.786 13-Docosenamide, (Z)- 112-84-5 0.57 99 44.203 Squalene 111-02-4 0.48 100 44.772 6,7-Dimethoxy-2-phenethyl-4H-chromen-4-one 84294-87- 1 1.84 Table 2. Twenty-seven sesquiterpene molecules.
No. Retention time (min) Name CAS Relative content 1 21.409 Isoaromadendrene epoxide 1000159-36- 6 0.06 2 23.066 Alloaromadendrene 10359-41-8 0.1 3 23.321 β-Chloranil 1000163- 00-2 0.06 4 23.423 α-Terpinene 489-28-1 0.16 5 23.499 β-Caryophyllene 1000159- 39-0 0.07 6 23.61 Desulfurized ketene 470-40-6 0.45 7 23.797 Agarospirol 1460-73-7 1.08 8 23.899 Patchoulene 1405-16-9 0.07 9 24.23 Arctiol 36061-11- 7 0.38 10 24.323 3,5,11-Dodecamethyltriene 193615- 07-5 0.06 11 24.476 Guaiol 489-86-1 0.15 12 24.697 Longipinocarvone 1000151- 87-1 0.31 13 24.884 Caryophyllene 87-44-5 0.36 14 26.056 Nerolidol 142-50-7 0.37 15 26.252 β-Himachalene 116-04-1 0.12 16 26.379 α-Cadinol 481-34-5 0.85 17 27.203 Isolongifolol 1139-17-9 0.43 18 27.704 Nootkatone 4674-50-4 5.63 19 28.121 Arteannuin b 1000212- 12-6 0.9 20 28.299 Isoshyobunone 1000360- 30-1 0.21 21 29.183 Nerolidyl acetate 2306-78-7 0.38 22 29.59 Himachalol II 19888-00- 7 0.78 23 30.1 Bohlmann k2631 80367-94- 8 4.55 24 30.372 Valerenic acid 609870 0.87 25 30.482 Ambrosin 509-93-3 3.08 26 31.434 Zederone 7727-79-9 0.68 27 31.527 Spirafolide 130838- 06-1 0.36 -
In this study, a new method for the formation of agarwood, whole-tree debarking method is reported. This method induces the regeneration of agarwood bark and stimulates the formation of secretory cells in the regenerated cork layer, leading to the production of agarwood. Compared with traditional methods of agarwood formation, this approach is characterized by rapid and continuous agarwood formation. Following the removal of the bark in A. sinensis, a silicate sol formed by sodium silicate and acetic acid was employed as a protective film matrix to prevent the loss of moisture and microbial invasion in the short term after bark removal. Additionally, an appropriate amount of vitamins were added to this matrix as nutritional supplements for the cell regeneration process, along with a suitable concentration of plant auxins as signaling molecules for the regeneration of cork formation layer cells. This protective film for A. sinensis bark effectively prevented tree mortality in practice and, in the process of promoting bark regeneration, accompanied by the proliferation of vascular bundle cells and the formation of secretory cells led to the acquisition of more agarwood secretions.
The variation in ethanol extract content within the regenerating bark indicates that the content of agarwood secretions increases progressively over time following the planting process. The content already surpassed the requirements for qualified agarwood as stipulated in the Chinese forestry industry standard LY/T 2904-2017, reaching over 10%, after 50 d after planting. Concurrently, HPLC analysis revealed an increase in the number of characteristic peaks over time, particularly in the samples collected at 119 and 135 d after planting, where the complexity of the chromatograms increased, indicating an enhanced diversity of secondary metabolites during this stage. Concurrently, the area of the characteristic peak for agarwood tetrol also gradually increased, indicating an improvement in the quality of agarwood with the extension of the planting time.
The agarwood formation site in this study is located beneath the regenerating bark, where the barrier of the bark reduces the influence of external microorganisms on the formation area. After 100 d of planting, numerous cracks appeared on the periphery of the regenerating bark, and the samples collected after this period were often contaminated with microorganisms, making it challenging to extract qualified RNA samples. Therefore, transcriptome analysis was only possible on the regenerating bark samples collected within 100 d after debarking. Considering the weather condition in the forestry, samples from 15, 68, and 100 d after debarking were collected to elucidate the expression patterns of synthesis genes for important sesquiterpene compounds in agarwood secretions. In agarwood, the biosynthesis of sesquiterpenes is closely related to the synthesis of isoprenoid precursors. The synthesis of these precursors can occur through the MVA pathway, which takes place in the cytoplasm, as well as the DXP pathway, which takes place in the plastids. Under the conditions of this agarwood planting technique, the expression levels of the genes related to the MVA pathway showed a declining trend, while the expression levels of the genes associated with the DXP pathway increased, suggesting that the DXP pathway may play a more critical role in this agarwood formation process. Additionally, qPCR analysis of multiple key sesquiterpene genes and significantly differentially expressed genes identified through transcriptome analysis further supported the accuracy of the transcriptome results.
To assess the practical benefits of this agarwood formation method in production practice, samples of regenerated agarwood bark were obtained 360 d after formation and the supercritical carbon dioxide extraction method utilized to obtain agarwood essential oil. The oil composition included 27 types of sesquiterpene components, with a relative content of 1.08% for agarospirol, which is one of the sesquiterpenoid compounds possessing significant biological activity and pharmacological effects in agarwood essential oil. According to the Chinese Pharmacopoeia, this harvested agarwood is qualified in traditional Chinese medicine applications. Compared to the whole-tree agarwood induction technology, this whole-tree debarking method produces relatively low fragrance content. However, since the harvest of fragrant woods from regenerated barks is sustainable, the whole-tree debarking method may have high potential in future applications. Therefore analyzing and understanding the content and characteristics of these compounds is of great importance.
We thank Omicshare (
www.omicshare.com ) for RNA-seq analysis. This research was funded by the National Key R&D Program of China (2021YFA0909600), and the Hainan Normal University Talent Research Start-up Fund Project Funding (HSZK-KYQD-202436, HSZK-KYQD-202421).-
The authors confirm contribution to the paper as follows: the majority of the experimental work conducted, draft manuscript preparation: Yan M, Zhou G; experimental work: Yan M, Lu Z, Li P, Xie M; supervised this study supervision: Wan Y. All authors reviewed the results and approved the final version of the manuscript.
-
Sequence data that support the findings of this study have been deposited in the NCBI database with the primary accession code PRJNA1102621.
-
The authors declare that they have no conflict of interest.
-
Received 22 August 2024; Accepted 25 November 2024; Published online 24 February 2025
-
Here a ring-barking method is reported to remove the bark of Aquilaria sinensis over a large area to increase the Agarwood secretion.
High-performance liquid chromatography and gas chromatography-mass spectrometry analysis revealed a stable improvement in agarwood quality over time.
Transcriptome sequencing identified the DXP pathway as dominant in agarwood formation, with decreased expression in the MVA pathway.
A total of 100 volatile components were identified in agarwood essential oil, including a high proportion of sesquiterpenoid substances.
-
# Authors contributed equally: Mengyao Yan, Zhongliang Lu
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yan M, Lu Z, Li P, Xie M, Zhou G, et al. 2025. Expression analysis of sesquiterpenes biosynthesis-related genes in Aquilaria sinensis during bark regeneration. Tropical Plants 4: e006 doi: 10.48130/tp-0024-0048 

Figure 1.
Cambial regeneration process after ring-barking for agarwood formation. (a) On-site bark ring-barking of agarwood; (b) the trunk sections with regenerated phloem layers removed at 15, 85, and 135 d after bark removal; (c) samples of the regenerated phloem layers at 35, 50, 119, and 135 d after bark removal; (d) abbreviations: rph - regenerated phloem; xy - xylem vessels; (e) bar chart of ethanol extract content: changes from 35 to 135 d. * Indicates p < 0.05, ** indicates p < 0.01, *** indicates p < 0.001. (f) High-performance liquid chromatography (HPLC) characteristic peak chart.