









-
Trichomes, originating from the epidermis, are widespread on the aerial organs of plants and can be found in most vascular plants. They play an important role in plant adaptation to different environments and resistance to various stresses[1−5]. Trichomes can be divided into unicellular trichomes and multicellular trichomes based on the cell number. Cotton (Gossypium spp.) fibers and Arabidopsis trichomes are unicellular, while trichomes in most vascular plants are multicellular in structure. Trichomes can also be divided into two functionally distinct types: non-glandular trichomes and glandular trichomes. In many plants including tomato (Solanum lycopersicum), Artemisia annua, Vitis and Cotton, glandular trichomes have been found to produce a large array of secondary metabolites, such as glandular trichomes on tomato, Artemisia annua and Cotton[1, 6−8].
Although unicellular trichomes in Arabidopsis have been extensively studied, we still have little understanding of the formation of multicellular trichomes, especially the glandular trichomes. To date, some studies on multicellular trichomes have been conducted on horticultural plants including cucumber and tomato. There are eight morphologically distinct types (I–VIII) on cucumber fruits, including two types of glandular trichomes and six types of non-glandular trichomes[9]. In cucumber, several transcription regulators for Arabidopsis trichome development, such as GLABROUS1 (GL1), GLABROUS3 (GL3), TRANSPARENT TESTA GLABRA1 (TTG1) and TRIPTYCHON (TRY) have been found to perform conserved functions in cucumber[10−13]. In tomato, seven types of trichomes have been identified which include four types of glandular trichomes and three types of non-glandular trichomes. Type I and type II are the longest trichomes with multicellular stalk and base, with type I containing a unicellular gland on the top. Type III trichomes are non-glandular with a unicellular base and are shorter than type II trichomes. Type IV and type V trichomes are similar in length, with a multicellular stalk and a unicellular base. A major difference is that type IV trichomes have a unicellular gland at the top. Type VI trichomes can be easily recognized as they exhibit a large four-celled gland and unicellular stalk. Type VII trichomes are often curved and are the smallest trichomes with 8−12 celled glands[14, 15].
A number of transcriptional factors have been reported to regulate multicellular trichome formation in tomato. Woolly, encoding a HD-ZIP transcriptional factor, regulates type I trichome formation[16]. Hair absent (H), encoding a C2H2 transcriptional factor, also regulates type I trichome formation[17]. SlMYC1 mediates JA signaling during type VI trichome development and terpene biosynthesis[18, 19]. HOMEODOMAIN PROTEIN8 is involved in trichome elongation, promoted by JA signaling[20, 21]. In addition, SlbHLH95 was found to act as a negative regulator to repress the trichome initiation in tomato[22]. Despite the identification of those regulatory factors, the detailed regulatory mechanisms are still unknown. An essential step before the exploration of the mechanisms involved, is a clearer understanding of spatiotemporal development and differentiation of tomato trichomes.
In this study, we conducted comprehensive observation of trichome development in different developmental stages of tomato leaves. We also observed the progression of gland differentiation and analyzed the DNA content in each trichome cell in tomato using multiple live-imaging markers. We found that spatial distribution of tomato trichomes changes over the course of leaf development with concurrent reduction of type IV trichomes and increase of other types of trichomes. We also found that the gland of type I/IV trichomes and the multicellular base is initiated as early as the two-cell stage. Through visualizing the nuclei of the stalk cells, we found a positive correlation between the length of stalk cells and nuclear size. Finally, we found two morphologically distinct forms of type VII glandular trichomes with the majority being covered by a wax-like substance which could help to form extra storage space for secretory metabolites.
-
It has been previously reported that type IV trichomes were mainly formed on embryonic (cotyledons) and juvenile leaves of cultivated tomato, and type IV trichome formation appeared to be associated with different developmental stages[23]. To further understand how spatiotemporal dynamics of trichome development occur over different developmental stages, we quantified all types of trichomes on the adaxial leaf surface at three different developmental stages, including 1 cm in size (the first stage), 2 cm in size (the second stage) and mature terminal leaflets (the third stage) (Fig. 1a). For this analysis, we grouped type II, III and V trichomes into one category (II+III+V) because all three types are non-glandular trichomes and could be easily recognized under scanning electron microscope (SEM) (Supplemental Fig. S1). We then grouped type I and IV trichomes into another category (I+IV) because both types are morphologically similar with a unicellular gland on top, easily recognized under SEM. As most type VII trichomes were found on the leaf vein, we didn’t include this type in the analysis.
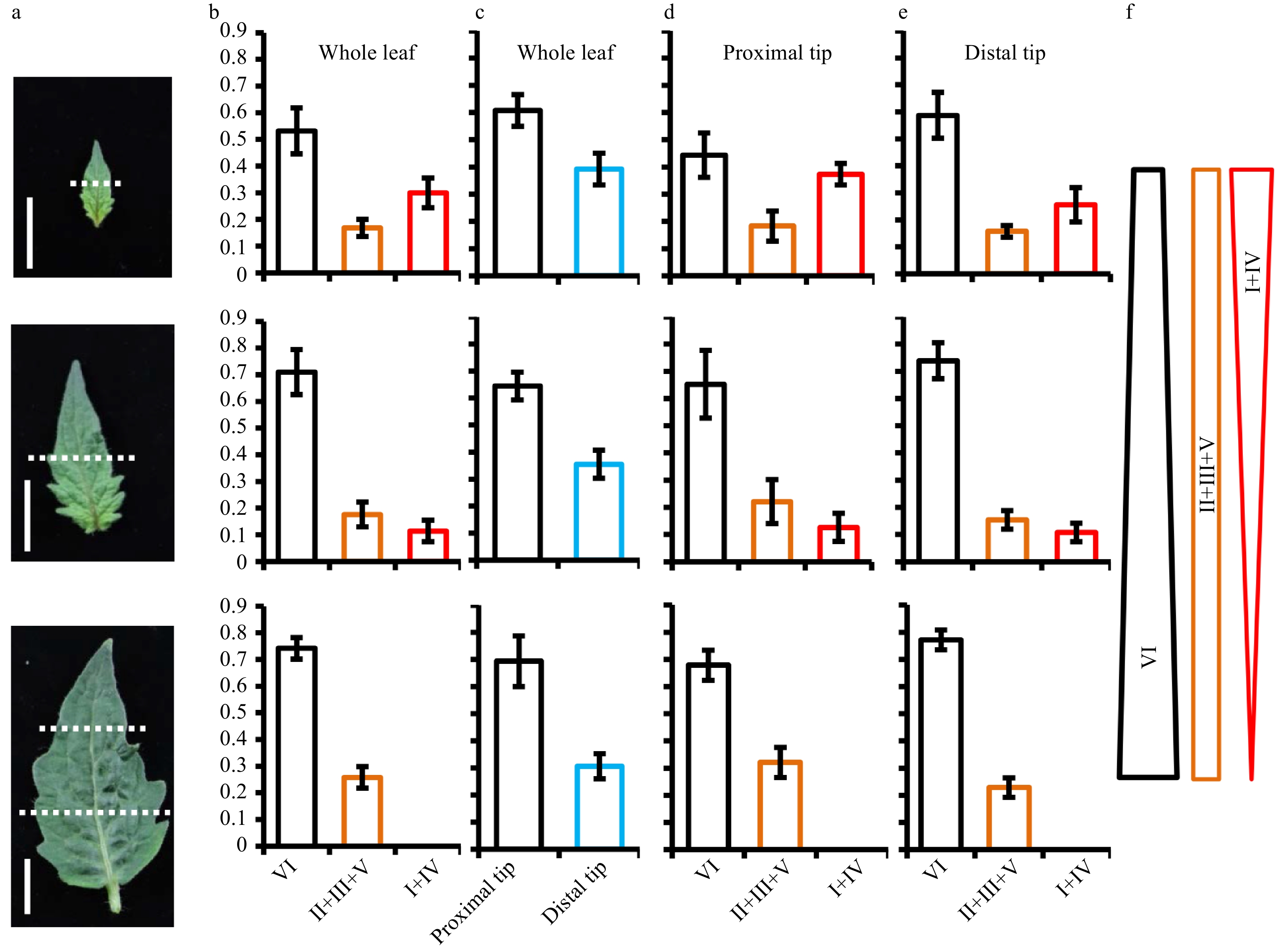
Figure 1. Spatiotemporal distribution of trichomes in the developing leaf. (a) Leaves at three different stages were used for analyzing the spatiotemporal distribution of trichomes. At the first and second stage, leaves were divided into two sections (proximal region and distal region) with approximate equal length and the dotted lines show the boundary of proximal region and distal region. At the third stage, leaves were divided into three parts (distal, middle, and proximal) with approximate equal length and the upper and lower parts of leaves were used to quantify the trichome number. Bars: 1 cm. (b) The percentage of each category trichome on the whole leaves. (c) The proportion of trichomes on the proximal region and distal region of whole leaves. (d−e) The percentage of each category trichome on the proximal region (d) and distal region (e) of leaves. (f) The schematic diagram summarizing the changing trend of each category of trichome over the leaf expansion. Type II, III and V were grouped into one category (marked as II+III+V); type I and IV were grouped into one category (marked as I+IV); type VI was a separate category. Five leaves at different stages were used to quantify the trichome number. The bars represent the standard deviation (SD) of 5 biological replicates.
Our results showed that nearly half of the trichomes were type VI glandular trichomes on 1 cm leaves. In the rest, 20% were non-glandular trichomes (II+III+V) and 30% were type I and IV glandular trichomes (Fig. 1b and Supplemental Table S1). The proportion of type VI trichomes increased while the frequency of type I and IV decreased with the transition from first to second stage. From the second stage to the third, type I and IV trichomes continued to reduce and had almost completely disappeared from mature terminal leaflets, while type II, III and V trichomes were concomitantly increased (Fig. 1b and Supplemental Table S1). Furthermore, we counted the trichome number on the proximal region and distal region of leaves respectively. Our result showed that more than 60% of trichomes were concentrated on the proximal region of leaves, with less than 40% of trichomes formed on the distal region throughout the whole developmental (Fig. 1c and Supplemental Table S1). In contrast to this overall trend, we found type VI trichomes showed a higher proportion on the distal region of leaves than the proximal region (Fig. 1d, e and Supplemental Table S1). In contrast to type I and IV trichomes, the proportion of type II, III and V trichomes was unchanged at the first and second stages. On mature leaves, the proportion of type II, III and V reached a peak on the proximal region (Fig. 1d, e and Supplemental Table S1). These results indicated that the differentiation into type I and IV trichomes is suppressed over the progression of leaf expansion, while the development of type VI trichomes and non-glandular trichomes continued.
Glandular cell development of type I/IV trichomes
-
It has been previously reported that the gland of type I/IV trichomes is the factory producing acyl sugars, which play an important role in insect resistance in tomato[24]. To track the development of the gland of type I/IV trichomes, we developed a specific type I/IV gland marker, SlAT2 (Solyc01g105580) which encodes an acyltransferase enzyme[25]. We generated pSlAT2: GUS-YFP construct by fusing the promoter of the SlAT2 with a β-glucuronidase (GUS) and yellow fluorescent protein (YFP) reporter gene and developed transgenic tomato consistently expressing this gene. Histochemical GUS staining and fluorescent observation by confocal microscope showed the clear signal in the gland cells of type I/IV trichomes (Fig. 2 and Supplemental Fig. S2). We observed the differentiation of type I/IV gland on the hypocotyl and juvenile leaves where type I/IV trichomes are mostly formed[23]. No GUS-YFP expression was detected when the trichome was initiated and this signal was rarely detected at the one cell stage (Supplemental Fig. S2b−f and Fig. 2a). From the two-cell stage to the mature stage, the expression of GUS-YFP became clearly visible in the top cell of the trichomes (Fig. 2 and Supplemental Fig. S2). Occasionally, we found the expression of GUS-YFP in the two top cells, with the signal weaker in the basal cell than the top cell, suggesting the process of cell fate transition (Supplemental Fig. S2h, i). These results suggest that the type I/IV gland could produce the ability to activate biosynthesis of acyl sugar as early as the two-cell stage. Interestingly, there was no morphological difference between type I/IV trichomes and non-glandular trichomes (mainly type II and III trichomes) at the two-cell stage (Fig. 2b), suggesting the cell fate was likely specified prior to the morphological change of the cell.
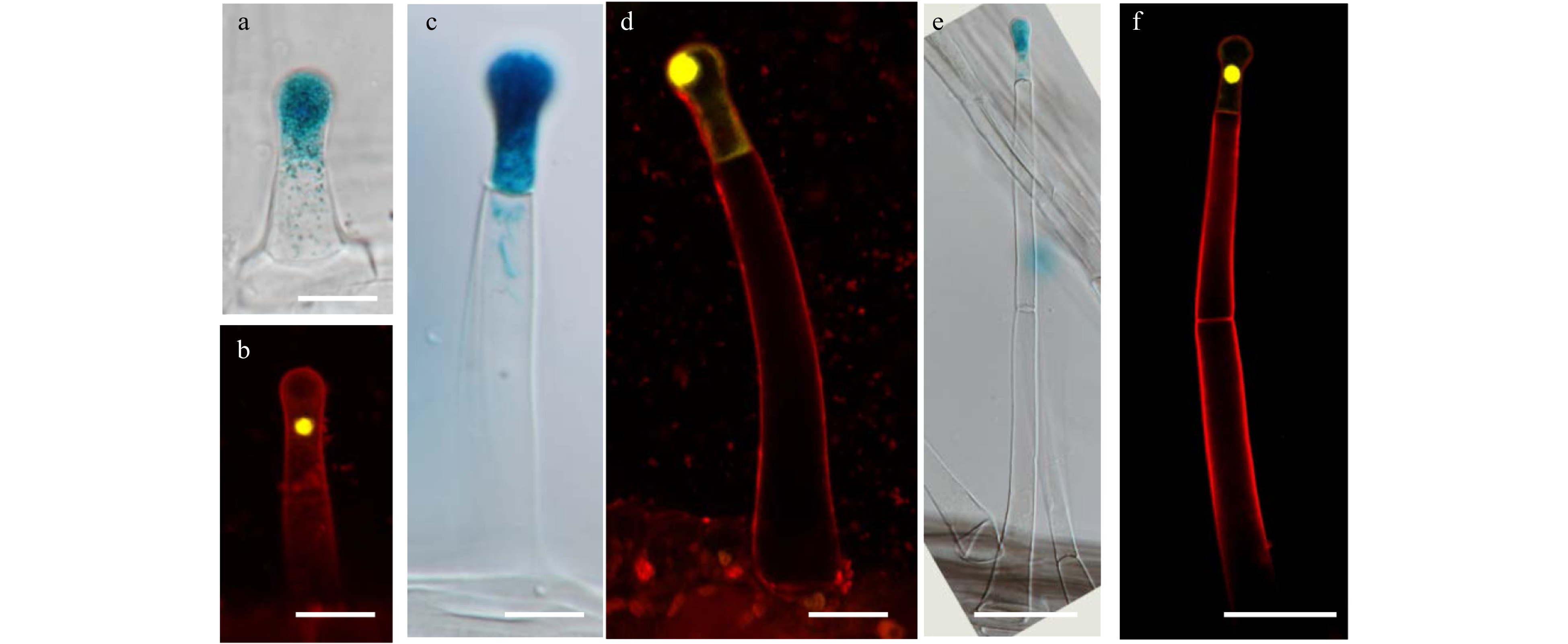
Figure 2. The expression of GUS and YFP driven by pSlAT2 in type I/IV trichomes. (a) GUS is expressed in the top cell of the trichome at the two-cell stage. The picture was taken by differential interference contrast microscope (DIC). Bar: 20 μm. (b) YFP is expressed in the top cell of the trichome at the two-cell stage. The sample was stained with 5 μg/ml propidium iodide (PI) (red fluorescence). Bar: 20 μm. (c−d) GUS (c) and YFP (d) are expressed in the top cell of type I/IV trichomes with one stalk cell. Bar: 20μm. (e−f) GUS (e) and GFP (f) are expressed in the top cell of the type I/IV trichomes with two stalk cells. Bar: 50 μm.
The formation of trichome base cells
-
Base cells are an indispensable component of trichomes, however their development has not been studied. In tomato, only type I and type II trichomes, the two longest trichomes, have multicellular bases, while the other shorter trichomes (type III, IV, V, VI and VII trichomes), have unicellular bases[14,26]. We speculate there may be a positive correlation between the base cell number and trichome length. We initially observed the developing leaves under SEM and found that the smallest trichomes with multicellular bases often contained two stalk cells (Fig. 3a), suggesting that the formation of trichome bases may be initiated at the two-stalk cell stage. We next examined the cell number and arrangement of the trichome bases under DIC microscopy using decolorized leaves. Our observation showed that the cell number in the trichome bases varied from 2 to 10, and these base cells varied in size. Interestingly, the base area seemed to change during development. Basal area was expanded as the trichome cell number was increased. In addition, the trichome basal cells on more mature tissues, including stems and mature leaves, were arranged in two or more layers (Supplemental Fig. S3).
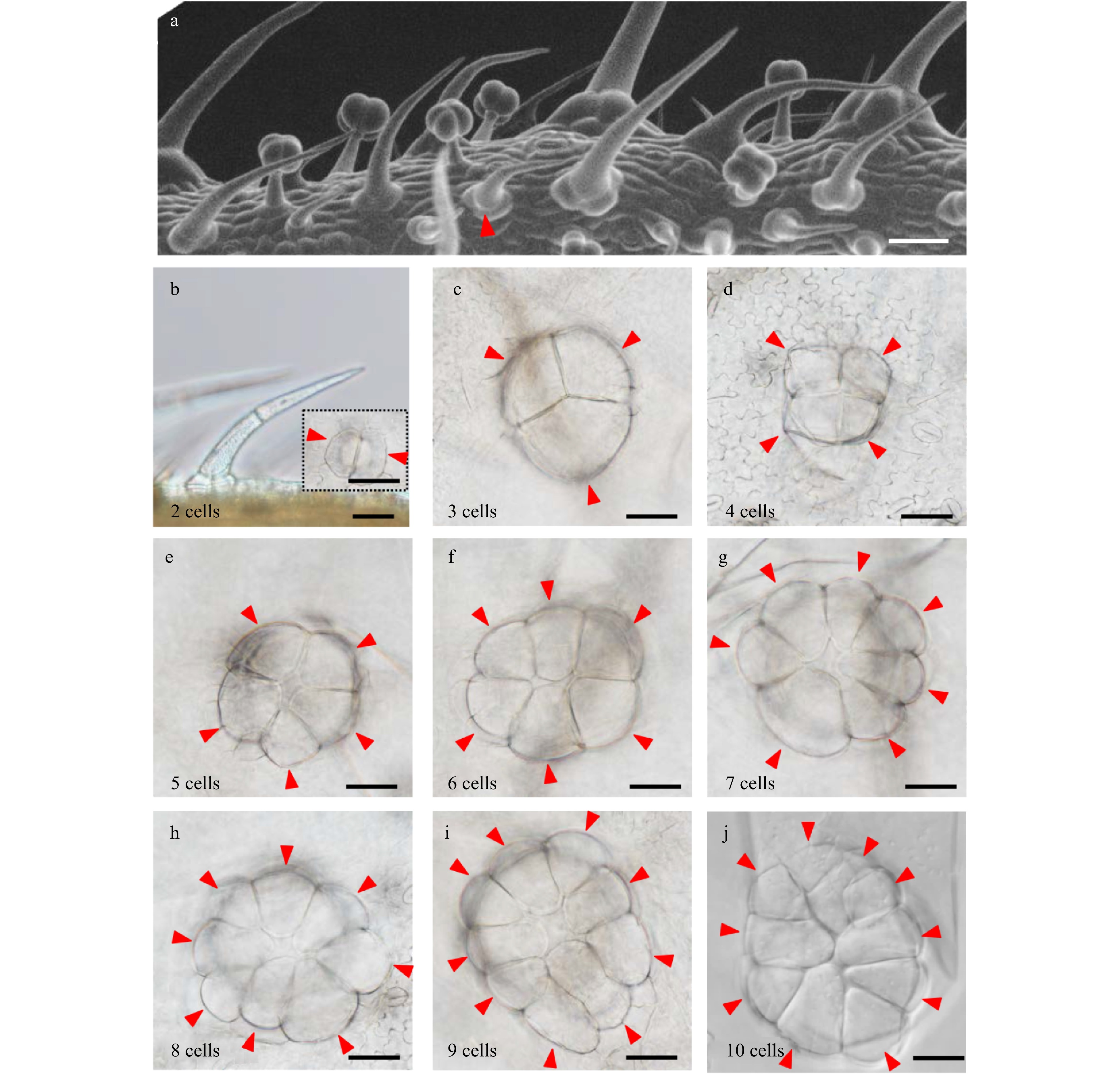
Figure 3. Number and arrangement of trichome base cells. (a) Observation of the trichome base at different developmental stages by SEM. The red triangles show the trichome with two stalk cells and two base cells. Bar: 40 μm. (b) Observation of the trichome with two stalk cells and two base cells by DIC. The picture surrounded by black dotted lines shows the base with two cells. The red triangles show the base cell of trichomes. Bars: 40 μm. (c−j) Trichome bases with three to ten cells. The numbers in the lower left corner of the picture show the number of the base cell. Bars: 40 μm.
DNA content of trichome cells
-
Trichome formation was initiated as a series of cellular events including cell fate specification and endoreduplication-driven cell expansion. In Arabidopsis, the DNA content of leaf trichomes can reach 16-32C through endoreduplication[5, 27]. From previous studies it is known that the initial trichome structure results from a number of periclinal divisions of trichome initial cells (TIC)[14, 28, 29]. However, little is known about endoreduplication and DNA content of tomato trichomes. To address this, we developed transgenic tomato plants expressing p35S:H2B-GFP to visualize the nuclei. We found that the nuclei of trichomes were generally larger than those in the leaf (Supplemental Fig. S4). Through quantifying the nuclei size, we estimated the DNA content of type I/IV glandular trichomes with 1−4 stalk cells and type II/III trichomes with 4−6 stalk cells. We did not quantify type V trichomes because it was difficult to distinguish these from the developing type II/III trichomes at the three-cell stage.
Our quantification showed that the nuclei size of the stalk cells was positively correlated with both trichome length and DNA content. The first stalk cells (1st SC) have higher DNA content than the top stalk cells in type II and II trichomes (Fig. 4). In detail, the 1st SC was about six times larger than that of stomata at the four-cell stage as well as eleven times as large as stomata at the six-cell stage (Fig 4a, c). However, the nuclei size of the apical cells of different trichomes had no correlation with trichome length (Fig. 4). Occasionally, we saw the nuclei of some apical cells were significantly larger than the sub-apical cells, which presumably represent the to-be-dividing apical cells (Supplemental Fig. S5a−c). In addition, the nuclei of two neighboring apical cells were very close, which might result from cells that had just finished division (Supplemental Fig. S5d). For type I/IV trichomes, the nuclei size of the stalk cells was also gradually reduced from the 1st SC to the top, and the stalk cell toward the base was about three times as large as that of stomas at the one-stalk cell stage and about five times as large as stomas at the four-stalk cell stage. Besides, the nuclei size of the glandular cells was similar to stomas at the one to three-stalk cell stages (Fig. 5a−c). Unfortunately, we were unable to quantify the nuclei size of the glandular cells at the four-stalk cell stage or more as no fluorescence signal was detected in these cells, which might be due to the inefficient expression of CaMV 35S promoter in these cells (Fig. 5d). Together, our quantification suggested that endoreduplication occurred during stalk cell expansion and that nuclear ploidy level was correlated with cell size.
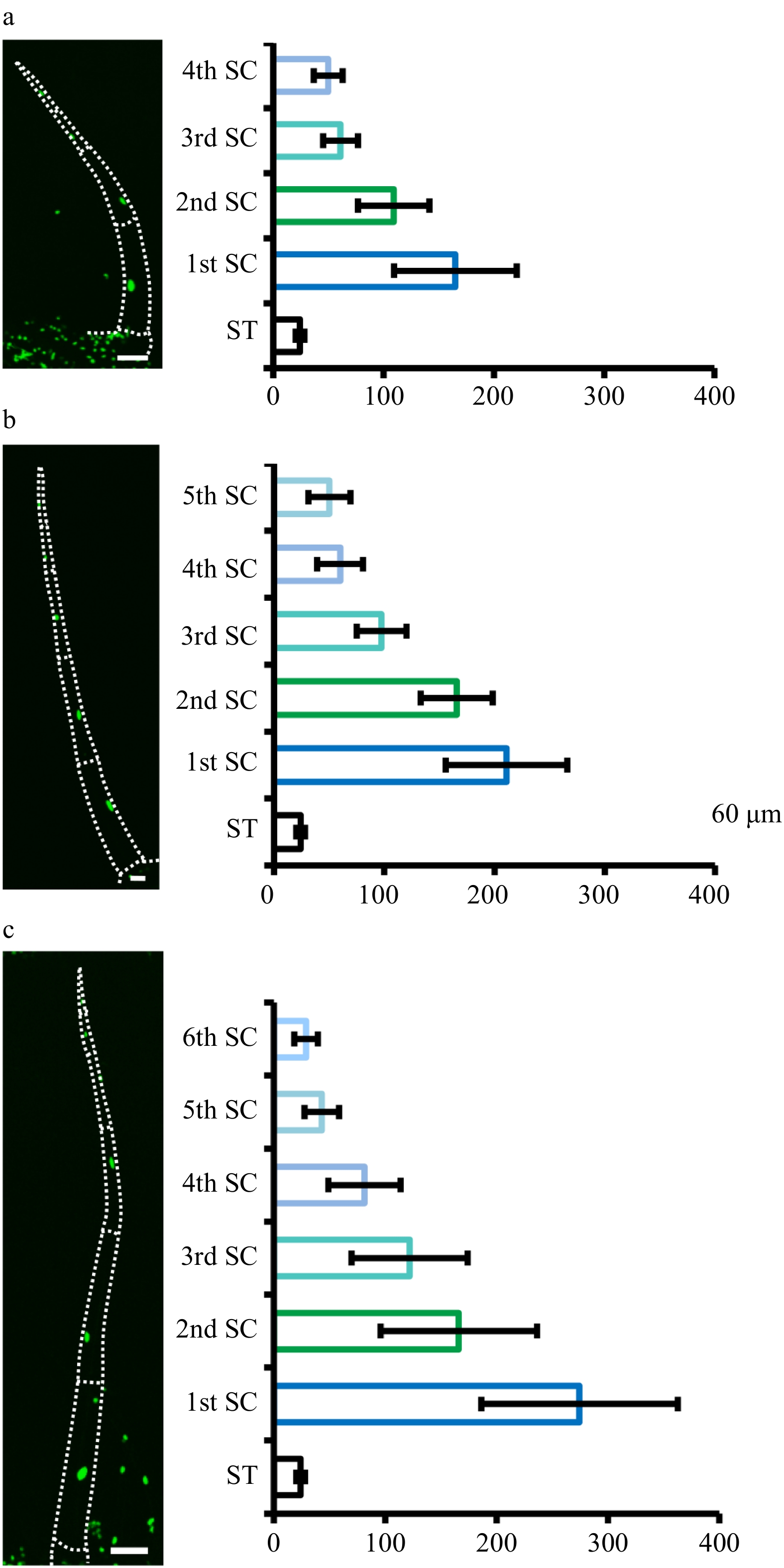
Figure 4. Quantification of nuclear size of type II/III. Quantification of nuclear size of type II/III trichomes with four (a), five (b) and six (c) stalk cells. Pictures were taken on transgenic plants expressing 35S::H2B-GFP (OE-H2B-GFP) under confocal microscope. The dotted lines show the outline of trichomes. The bar charts show the statistical results of nuclear size. The nuclei of the stomata (ST) are the control. The stalk cells (SC) towards the leaf epidermis are marked as the first stalk cell (1st SC) and the next as the second stalk cell (2nd SC), and so on. The error bars on the charts represent the standard deviation (SD) of more than ten biological replicates.
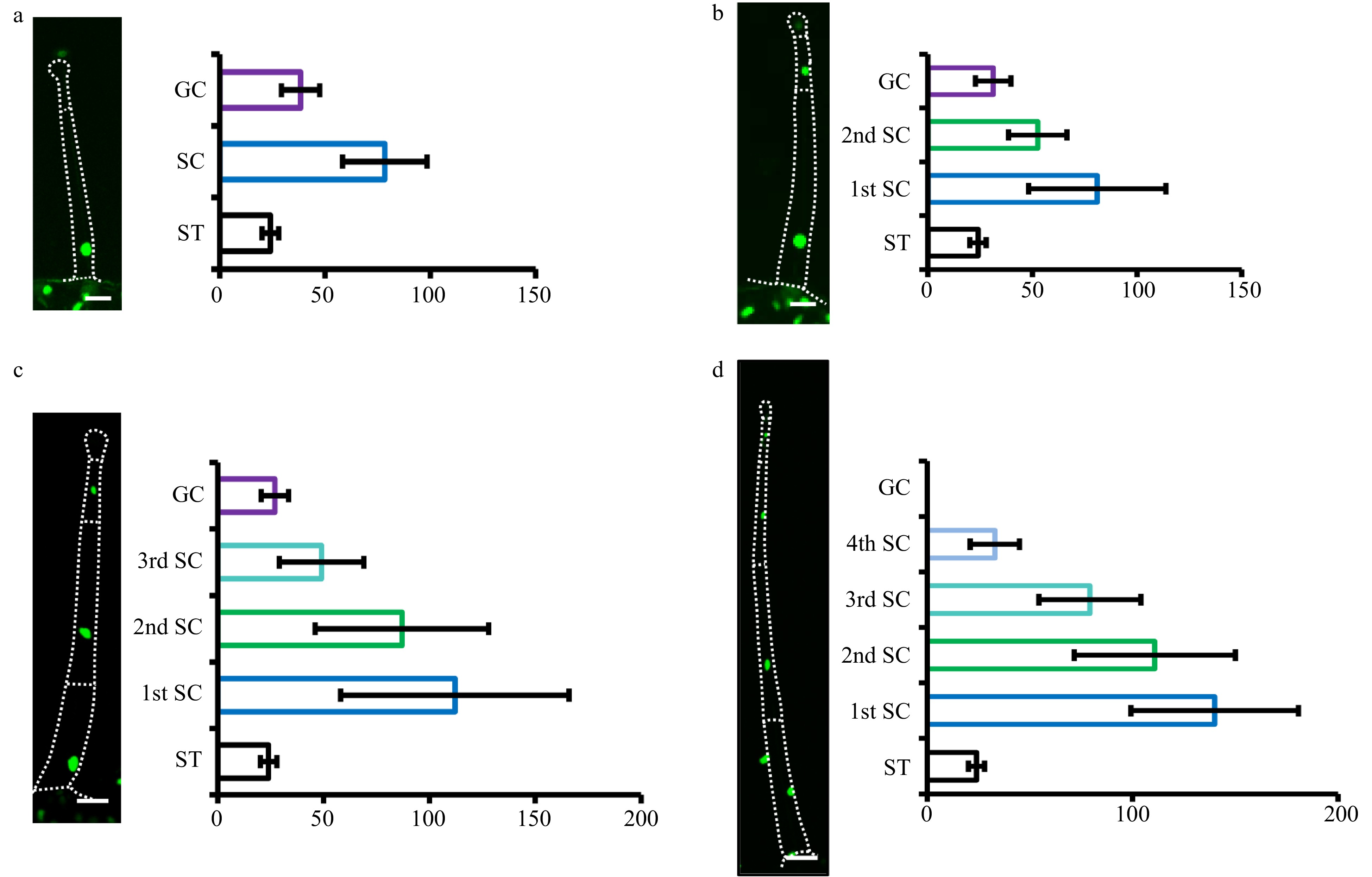
Figure 5. Quantification of nuclear size of type I/IV. The nuclei of type I/IV trichomes with one to four stalk cells were quantified (a−d). Pictures were taken on transgenic OE-H2B-GFP plants under confocal microscope. The dotted lines show the outline of trichomes. The gland cells and stalk cells of type I/IV trichomes with one to three stalk cells were analyzed, while only stalk cells of type I/IV trichomes four stalk cells were analyzed, as GFP fluorescence was difficult to detect in the gland of type I/IV trichomes with four stalk cells. The nuclei of the stomata (ST) are the control. The stalk cells (SC) towards the leaf epidermis were marked as the first stalk cell (1st SC) and the next one as the second stalk cell (2nd SC), and so on. The gland cells were labeled as GC. The error bars on the charts represent the standard deviation (SD) of more than ten biological replicates.
The morphology of type VII trichomes in Micro-tom
-
Although the morphology of I-VI trichomes has been well elucidated, type VII glandular trichomes are rarely described due to their extremely small size. In our previous study, we found that type VII were the second most abundant glandular trichomes in Micro-tom[17]. This type of trichome, like the type I-VI trichomes, can also be described as a typical trichome with a base, stalk and gland (Fig. 6). To further characterize the morphology of type VII trichomes, we examined them with multiple approaches including tissue sectioning, SEM and live-imaging with the 35S:H2B-GFP marker. We found that the glandular cells of type VII trichomes varied from eight to twelve and these cells were arranged on two or three planes (Fig. 6a−d and Supplemental Fig. S6a−g). Based on the gland cell organization, we classified them into two main types named as type VII-a containing two layers of gland cells and type VII-b containing three layers of gland cells (Fig. 6 and Supplemental Fig. S6). Interestingly, both types were surrounded by the substance that forms balloon-like structures (the left images of Figure 6a, b; Fig. 6e, f and Supplemental Fig. S5j). The morphology of these trichomes appeared to be similar to the peltate trichomes in peppermint (Mentha × piperita), basil (Ocimum basilicum), and lavender (Lavandula spp.), which are covered by cuticles serving as extracellular storage space for secondary metabolites[30−33]. It is possible that the glandular cells of VII trichomes contain similar structure to those with storage roles. In support of this, we saw droplet like substances attached to the type VI glands, with some even secreted onto the leaf surface, which were likely secondary metabolites (Supplemental Fig. S6c, d, I).
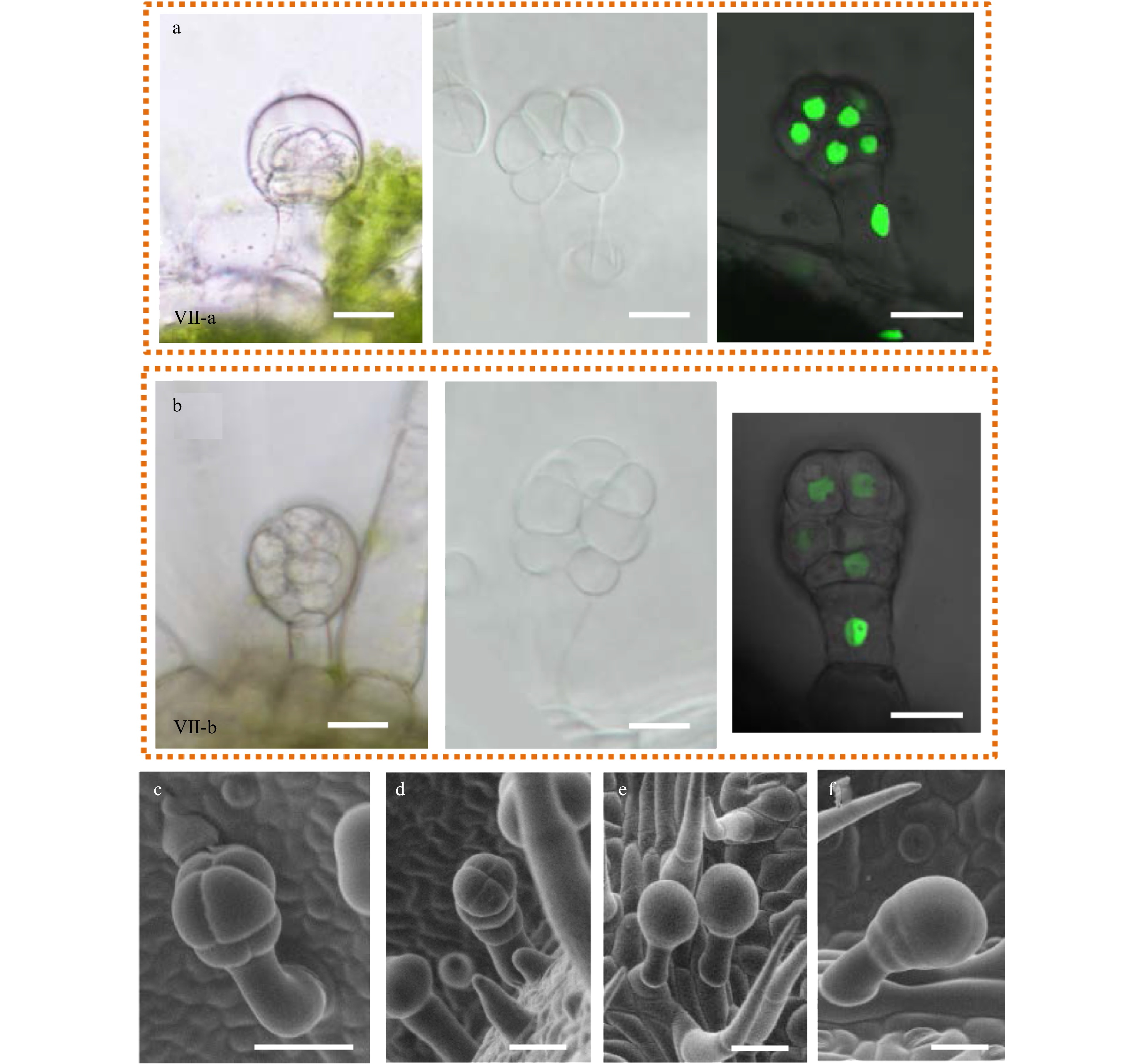
Figure 6. The morphology of type VII glandular trichomes. (a−b) The morphology of type VII glandular trichomes was observed by DIC (left and middle pictures) and confocal microscope (right picture). The left pictures were taken on the fresh section and the middle pictures were taken on the decolored leaves. The right pictures were taken on the leaves from OE-H2B-GFP transgenic plants. Two different types of VII trichomes were found, named as type VII-a (a) and type VII-b (b). (c−f) Type VII trichomes with different morphology were observed under SEM.
-
Cell division and cell expansion are two important processes during organ morphogenesis[34, 35]. Tomato trichomes are initiated as a protrusion from the epidermal surface, but eventually form appending multicellular structures, providing an ideal system for studying cell division and cell expansion[14, 29]. The initiation of multicellular trichomes involves multiple rounds of periclinal divisions. To reveal the regulatory mechanism behind the cell division in multicellular trichomes, we need to understand the exact division pattern during trichome formation. Our previous study showed that the trichome gradually becomes mature from the top down to the basal cells[14]. Accordingly, it is possible that only the top cells divide during trichome formation. In this study, we found that the nuclear size of the basal cells was larger than the top cells, suggesting the ploidy level of the basal cells was higher. We also found frequent division of the top cells, which could prevent endoreduplication[36]. Furthermore, we often saw cell division in the top cells but not in the base cells in the H2B-GFP marker lines (Supplemental Fig. S4). Therefore, we provide more evidence in this study supporting that the top cells keep dividing during tomato multicellular trichome development, which leads to an increase in trichome cell number.
Trichome cells differentiate as early as the two-cell stage
-
Tomato produces seven morphologically and functionally distinct types of trichomes[14, 15, 26]. The type and density of trichomes vary at different developmental stages and on different tissues[23], indicating that the differentiation of trichomes is a complex regulatory process. A previous study showed that pSlAT2: GUS-YFP was expressed in the tip cells of type I/IV trichomes[25]. Using the promoter of SlAT2, we constructed a reporter line in which we can monitor the differentiation of the tip cells. We found such differentiation at the one-cell stage. This suggests that these trichome cells are specified in early stages and do not wait until most divisions finish. Interestingly, we found that the stalk cell of the type I/IV trichomes near the leaf epidermis elongated quickly at the two-cell stage, which generates the clearly different morphology among trichome cells at the end of the two-cell stage. Together, we think the differentiation of the different trichome cells occurs prior to the two-cell stage. In addition, we found that there were some curved trichomes at the one-cell stage that could develop into type VII trichomes which are typically curved. Therefore, our results support that trichome fate determination likely occurs in the early stages of trichome initiation.
Often overlooked type VII trichomes could be important
-
The most valuable and meaningful function of glandular trichomes is to biosynthesize and secrete metabolites, which requires enough storage space[37]. Glandular trichomes have evolved morphological features that allow them to accumulate considerable amounts of metabolites, such as lavender (Lavandula spp.), Artemisia annua, sunflower (Helianthus annuus) and peppermint (Mentha x piperita) whose glands are located under the cuticle, forming a storage space for metabolites[30−33]. In tomato, type VI trichomes have an intercellular space for metabolite storage[38]. Tomato type VII trichomes are covered with some uncharacterized substance that forms a similar storage space on the top of the glandular cell, suggesting type VII trichomes have the potential to store more secondary metabolites. Compared to other tomato cultivars, Micro-tom has a very high proportion of type VII trichomes[14]. Micro-tom may therefore serve as the ideal system for studying the development of type VII trichomes. The study of this type of trichomes could provide new insights into the increase of secondary metabolites and enhancing insect resistance in cultivated tomato.
-
Tomato, Solanum lycopersicum L cv Micro-Tom, was used as the wild type for all experiments. The wild type and transgenic plants were grown in a green house under the conditions of 25 °C, 16 h : 8 h, light : dark cycle and 60% relative humidity.
Construction of marker lines
-
Acyltransferase 2 (Solyc01g105580) has previously been described and demonstrated to encode an acyltransferase with specific expression in type I/IV tomato trichome gland[25]. To figure out the differentiation of the gland of type IV trichomes, we constructed transgenic plants expressing GUS-YFP fusion protein under the SlAT2 promoter as type I/IV gland marker lines.
Histone H2B with fluorescence lines is a new biological reagent for visualizing chromatin structure, chromosome morphology, and nuclear dynamics in fixed and living cells in a model plant genetic system. To visualize the nuclei of trichome cells, we inserted AtH2B-GFP (AT3G45980) driven by CaMV 35S into pHellsgate8. Tomato stable transformation was conducted by Agrobacterium-mediated transformation as previously described.
Quantification of trichomes
-
To quantify the trichome number on the different developing leaves, the terminal leaflets of the fourth leaves were sampled to count the trichomes in leaves. The trichomes in the leaf are divided into three types: type VI, type II+III+V and type I+IV. The photos were taken by scanning electron microscope (SEM, Hiiachi TM3030 PLUS) and densities were counted by Image J software. Error bars represent the standard deviation (SD) of the biological replicates.
DIC microscopy
-
To obtain the interference contrast (DIC) images of the trichomes, cotyledons and leaves were treated with destaining (75% ethanol [v/v] and 25% acetic acid [v/v]) at room temperature until the chlorophyll was clear (about 30 min). After treatment with transparent solution (7% NaOH [g/v] in 60% ethanol [v/v]) for 1 h, the samples were rehydrated through an ethanol series (40%, 20% and 10%) for 15 min at each step. Finally, seedlings were placed on glass slides and photographed under DIC microscope (Nikon, ECLIPSE Ni-U).
For GUS observation, plant materials were incubated in GUS staining solution (10 mM EDTA, 0.1% [v/v] Triton X-100, 2 mM potassium ferricyanide, 100 mg/mL chloramphenicol, 1 mg/mL X-Gluc in 50 mM sodium phosphate buffer, pH 7.0) at 37 °C overnight. The transparent method described above was used for transparency and observation.
Confocal microscopy
-
YFP signal of pSlAT2: YFP-GUS transgenic plants was visualized using Zeiss LSM 880. Before observation, seedlings were stained with 5 μg/ml propidium iodide (PI) for 10 min. Fluorescence from YFP was detected by excitation at 488 nm and PI signal was detected with a 543 nm long-pass emission filter. The expression of H2B-GFP was detected using an Olympus FluoView 1000 laser scanning confocal microscope. The cell nuclei size was quantified using the 3D reconstructed images. All three-dimensional images were processed with Olympus software and Zeiss software.
- This work is supported by the National Key Research and Development Program of China (2018YFD1000800) to S.W, the National Natural Science Foundation of China (32000157) to Jiang Chang, and the Natural Science Foundation of Fujian Province (2019J01379) to Jiang Chang.
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Leaves used for analyzing spatiotemporal distribution of trichomes in the developing leaf by SEM. (A B) The leaf in the first stage; (C D) The leaf in the second stage; (E F) The leaf in the third stage. Bars: 2 mm.
- Supplemental Fig. S2 The expression of GUS and YFP driven by pSlAT2. (A) GUS driven by SlAT2 promoter is expressed in the top cell (gland) of type I/IV glandular trichomes on the leaf. (B&C) The GUS signal could not be detected in the initial cells of trichomes. Bar: 200μm. (D&E) The trichomes with one stalk cell could occasionally be stained by 1 mg/mL X−Gluc (This phenomenon was rare). (F) Weak YFP signal could be detected in the trichome at one cell stage (This phenomenon was rare). (G) GUS is expressed in the top cell of the trichomes with two smaller stalk cells. (H&I) YFP signal could be found in the two cells (This phenomenon was very rare). Bar: 20μm.
- Supplemental Fig. S3 The arrangement of trichome base cells on mature leaves and stems. (A&B) The base cells are arranged in one layer on the mature leaves under SEM.The red dotted boxes show the trichome bases.The images on the right are an amplified view of the trichome base in the left image. (C) Some basal cells are arranged in one layer and others arranged in two layers on the mature leaves under SEM.The red triangle represents the base cells arranged in two layers. (D) The base cells are arranged in one layer on the stems under SEM. (E−F) The base cells are arranged in two or more layers on the stems under SEM. Bars: 100μm.
- Supplemental Fig. S4 The nuclei of trichomes cells (A) and stomas (B) . The pictures were taken on the leaves from transgenic plants of p35S::H2B−GFP. The sample was stained with 5 μg/ml propidium iodide (PI) (Red fluorescence). Bar: 70μm.
- Supplemental Fig. S5 Dividing trichome cells. The white arrows indicate cells that may be dividing (A−C) or have just finished division (D) . The pictures were taken on the leaves of OE−H2B−GFP transgenic plants stained by PI. Bar: 40μm.
- Supplemental Fig. S6 The morphology of type VII glandular trichomes. The red arrows represent droplet like substances attached to the type VI glands. Bars: 40 μm.
- Supplemental Table S1 The total number of each type of trichomes in the developing leaf.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chang J, Yang J, Wang J, Wu S. 2021. Comprehensive observation of trichome development in Micro-tom tomato. Vegetable Research 1: 6 doi: 10.48130/VR-2021-0006

Comprehensive observation of trichome development in Micro-tom tomato
- Received: 03 April 2021
- Accepted: 11 June 2021
- Published online: 23 June 2021
Abstract: Trichomes help plants defend against various stresses by formation of a protective barrier. Although the unicellular trichomes in Arabidopsis have been well studied, little is known of the development of multicellular trichomes in tomato. With seven morphologically and functionally different types, tomato trichomes are more complex than the unicellular trichomes in Arabidopsis. Here, we comprehensively analyzed the spatiotemporal pattern of tomato trichome development. Our observations show that the gradual reduction of type I/IV trichomes during leaf development is accompanied by an increase of other trichome types. We further examined the gland development of type I/IV trichomes by constructing a marker reporter, pSlAT2: GUS-YFP which is specifically expressed in the top cell of the trichomes. Interestingly, we found the multicellular base of trichomes is initiated as early as the two-cell stage. Through tracking a nuclear marker, H2B-GFP in tomato trichomes, we found the correlation between stalk cell length and nuclear size, suggesting that cell ploidy level could affect trichome cell size. Finally, we found type VII glandular trichomes exhibit two morphologically distinct forms, with the majority covered by a wax-like substance, which could help to form extra storage space for secretory metabolites. Our results provide the basis for further mechanistic study of trichome development in tomato.
-
Key words:
- trichomes /
- development /
- tomato