








-
Tomato (Solanum lycopersicum L.) is an important source of antioxidants, carbohydrates, carotenoids, nicotinic acid, lycopene, vitamins, and several phyto-compounds[1−4]. Tomato consumption has been found to contribute to the prevention of cancer, coronary and emerging cardiovascular disorders[3,4]. In molecular biology, tomato is a model plant for studying fleshy fruit biology and fruit quality[5−7]. Tomatoes are free of cholesterol and a good source of fiber and natural phenols as they produce less fat and calories. Intensive domestication and breeding practices have enhanced tomato yield, fruit quality, color formation, mechanical harvesting, pest and disease resistance[8]. These strategies have albeit, decreased genetic diversity while improving the productivity of this crop, as some of the breeding efforts were counterproductive to the nutritional and sensory qualities of new commercial varieties, which are mostly seen by consumers as less flavorsome[2,3,9,10]. Fruit quality is measured based on external and internal characteristics. The external quality factors are fruit size, color, and texture which are easily detected by the naked eye[2]. The level of sugar and vitamins, and bioactive compounds (anthocyanin, lycopene, and malate) constitute the internal fruit quality attributes[11−13]. Flavor is an important characteristic in the domestication of fruit crops[14]. The tomato flavor palette has changed substantially as a result of its domestication and diversification[15−17]. It was the wild, red-fruited species, Solanum pimpinellifolium that gave rise to the cultivated tomato species, Solanum lycopersicum var. lycopersicum in South America, and it was from this variety that the domesticated tomato, Solanum lycopersicum var. lycopersicum emerged in Mexico[14,18].
Sugars, along with phytohormones, control the growth, development, and metabolism of organs to determine sugar dynamics in tomato fruits[19]. The key constituents that contribute to the flavor of tomatoes include sugars (sweetness), organic acids (acidity), volatile compounds (aroma), and texture (firmness and juiciness). Volatiles in the fruit interact with olfactory receptors in the nose, and the brain is able to recognize such distinct characteristics as 'sweet', 'smoky', or 'fruity'[20,21]. Volatile compounds are present in both primary and secondary metabolites in ripe tomato. More than 400 volatile compounds have been detected in ripe tomato fruits. Biosynthesized volatiles and metabolites during tomato maturation account for the taste and flavor of most commercial genotypes[22,23]. Cis-3-hexanal, cis-3-hexanol, hexanal, 3-methylbutanal, 6-methyl-5-hepten-2-one, methyl salicylate, 2-isobutylthiazole, 1- pentan-3-one, trans-2-hexanal, and b-ionone are among the prime contributors to ripe tomato flavor[22]. Multiple genes, including LIN5, ALMT9, AAT1, CXE1, and LoxC influence the amounts of these metabolites in tomato fruit.
New commercial tomato varieties possess substantially lower concentrations of essential flavor chemicals than older varieties as they have not been the focus of modern breeding programs[24−26]. The multipart and distinct metabolite regulation in tomato subspecies has been demonstrated by QTL study, genome-wide association studies, and targeted metabolome analysis in several cultivars and accessions[25−27]. Sucrose makes up less than 1% of the dry weight of fruits, whereas fructose (25%) and glucose (22%) are the main accumulated soluble sugars[28]. During the early stages of fruit development, the amount of glucose and fructose in tomato fruits increase considerably[29]. Hexose phosphates are primarily used in the synthesis of starch in green fruits until 13 days post anthesis. In the early stages, starch accumulation in the pericarp and columella tissues is a critical factor in determining the total soluble solid content of mature fruits[22].
Invertase (EC 3.2.1.26, INV) is indispensable for sugar metabolism, growth, and stress responses in tomato. Soluble acid invertase is responsible for the majority of invertase activity in tomato[9]. Tomato acid invertase isoforms have been isolated and cloned[30], while invertase inhibitor proteins have also been isolated and cloned[31−33]. Tomato apoplastic invertases are encoded by at least four distinct genes, each of which has a very unique organ expression pattern[34]. It is difficult to create unambiguous techniques for the study of compartmentation and the complex control of invertase by inhibitors, which makes it impossible to establish the activity and role of each of the acid invertase isoforms[35,36].
This review discusses recent research advances that sheds light on the roles of invertase in sucrose metabolism and flavor regulation in tomato fruits. In this paper, we provide evidence from recent research on the essential role of invertase in the regulation of sugar metabolism and fruit flavor in tomato.
-
Sugar is one of the critical molecules modulating quality of tomato fruit and flavor, although this depends on the genotype and maturation stage[37−40]. Sugars comprise about 55%−65% of fruit soluble solids and contribute to the overall flavor of tomato fruit. Stommel[41] found that green-fruited tomatoes (sub-species eulycopersicon) accrue high sugar in contrast to red-fruited tomatoes (sub-species eriopersicon), which mainly store reduced sugar. Polysaccharides comprise about 0.7% of tomato juice, with pectins, arabinogalactans, xylans, and cellulose as the main constituents[42]. The remaining fruit carbohydrates are accounted for by soluble reducing sugars, mainly glucose and fructose[43]. Glucose and fructose levels increase in ripening fruits[39]. An invertase enzyme activity breaks down sucrose into D-glucose and D-fructose[39,44].
The extracellular invertase substrate that hydrolyzes sucrose to glucose and fructose monomers is apoplastic sucrose. Monosaccharide transporters import hexoses into carbon sink cells[45]. Invertase is vital in regulating assimilate partitioning in the three protein-mediated steps (two transporters and invertase)[19,46]. Extracellular invertase activities and hexose transporters cushion the cells to transport phloem carbohydrates and strengthen carbon sink tissues[19]. These are typically enriched by a co-ordinated cytokinin induction[47], co-expression of extracellular invertase, and hexose transporters[48]. Invertase is also vital in carbon source-sink regulation and developmental processes as reported by several studies, including overexpression of yeast invertase in the apoplast of transgenic tobacco plants[49], invertase deficient maize mutant[50], regulation of seed development[51], antisense suppression of extracellular invertase in transgenic carrot plants[52], and an anther-specific isoenzyme of tobacco[53].
Membrane transporters are gateways to the transport of metabolites, and their intercompartmental compounds can exert extensive effects on fluxes[54]. Their functions in sugar transporters and tomato fruit growth have not yet been thoroughly studied. Evidence for regulating tomato sugar transporters by endogenous sugars via kinases adds an extra layer of complexity to delineating their role in transporters and carbohydrate accumulation in fruits[44]. Sugar signaling and sensing by invertase, hexokinases, and unidentified proteins are often implicated in carbohydrate accumulation and modulation of fruit flavor[1,3,4,39]. Invertase transforms sucrose into hexoses used for energy, and signaling molecules for growth and development (Table 1), including carbon partitioning into sink tissues[55,56]. Earlier studies on fruit flavor reveal correlations between sugar content and fruit development, fertility, and phytohormones[39,57]. Hexokinases are sugar receptors central to the regulation of sugars in tomato[58−60]. The consequence of overexpressing a native hexokinase using a fruit-specific promoter is yet to be deciphered[61].
Table 1. Major roles of different invertases and sucrose synthase in tomato.
Invertase/sucrose Role Source Cell wall invertase Modulates tomato fruit flavor [8, 62, 63] Adjusts sucrose allotment between source and sink organs [31, 64] Enhances plant response to stress/signal transduction [57, 65−67] Modulates total soluble solid content in mature fruits [10, 68, 69] Regulates cell differentiation and fruit development [44] Sustains the apoplastic glucose and fructose content at an optimum level [43, 70] Vacuolar invertase Influences fruits sugar composition and storage organs [10, 44, 71, 72] Regulates plant response to osmotic stress [73] Modulates tomato response to cold stress [74, 75] Cytoplasmic invertase Adjusts sucrose metabolism in tomato [74−76] Regulates biosynthesis of volatiles in matured fruits [25, 77, 78] Modulates sugar gene expression in matured fruits [71] Sucrose synthase Adjusts sucrose partitioning between source and sink organs [35, 48, 67, 68, 79] Regulates sucrose metabolism in matured fruits [41] Enhances tomato response to cold stress [66] Modulates tomato fruit flavor [29, 31, 62, 80] -
Sucrose is the major translocated photoassimilate in tomato, usually transported from carbon source to sink tissues in response to a pressure gradient precipitated by the osmotic potential at the phloem loading/unloading sites[19]. Fruit cells are the major receptacles for sucrose transported from the symplast[19]. Sucrose may also be metabolized in the apoplast by a cell wall invertase, and the resultant hexoses are imported via plasma membrane hexose transporters (Table 1, Fig. 1). Previously, it was thought that fresh fruits were the preponderant organs for sucrose symplastic loading, while ripening fruits housed the apoplastic hexose loading[48]. Apoplastic loading has recently been found during fruit growth, thus debunking the earlier proposition[66].
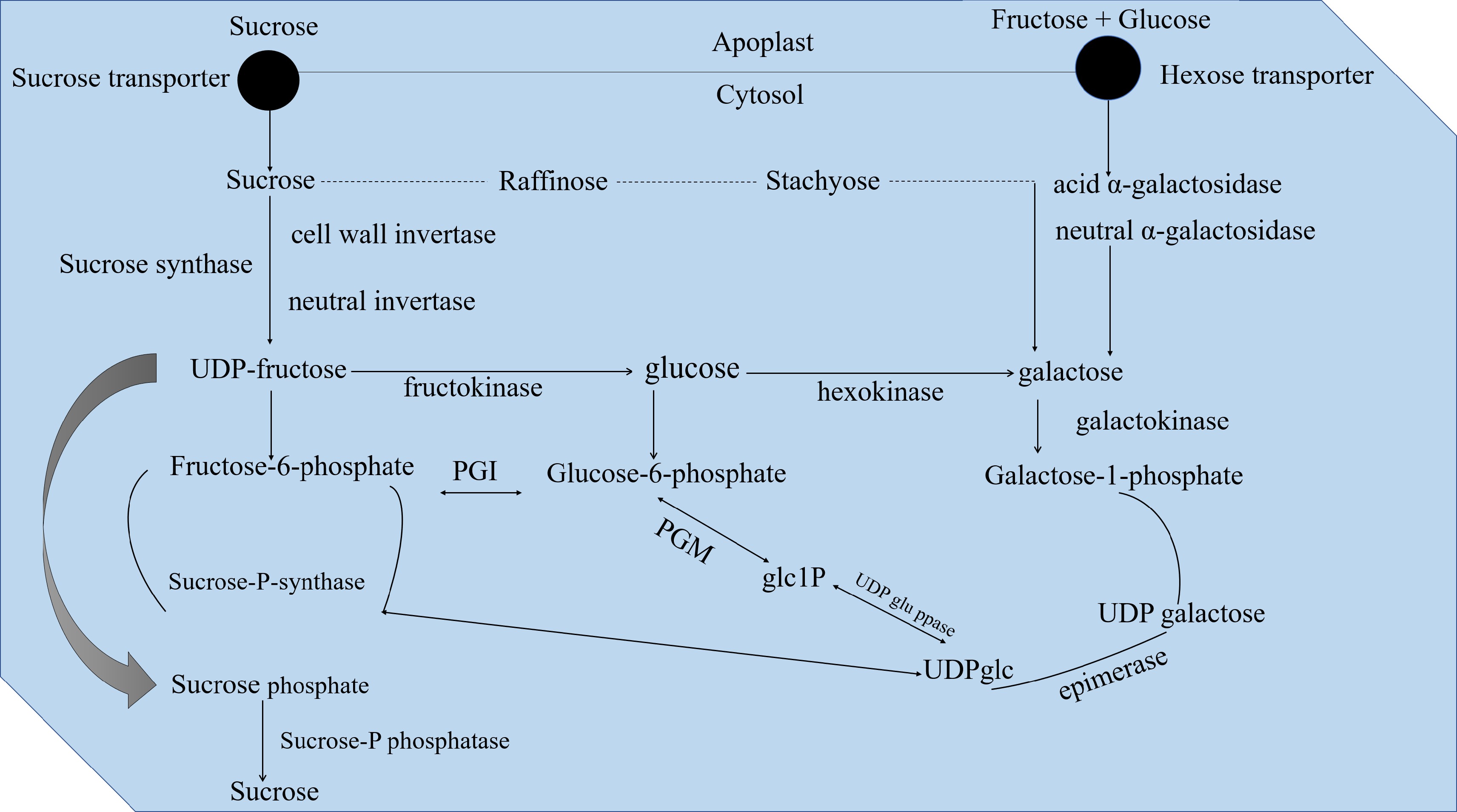
Figure 1. Invertase catalyzes the pivotal step in sugar metabolism. Sucrose unloading in tomato fruit is squarely a controlled process. The sucrose and hexose transporters modulate synthesis, loading, unloading, membrane transport, metabolic conversion, and compartmentalization of translocated sugars and sucrose. Sucrose is degraded in vacuole by invertase and re-synthesized in the cytosol catalyzed by sucrose synthase. The final stage encompasses sucrose degradation in the apoplast by invertase following sucrose synthesis in the cytosol and subsequently catalyzed by the sucrose synthase, UDP, Uridine diphosphate. Phosphoglucomutase (EC 5.4.2.2), also written as PGM is an enzyme that transfers a phosphate group on a -D-glucose monomer from position 1 to position 6 in the forward direction, or from position 6 to position 1 in the reverse direction. More specifically, it aids in the conversion of glucose 1-phosphate into glucose 6-phosphate. Phosphoglucose isomerase (PGI; EC 5.3. 1.9) is a cytosolic glycolytic enzyme that catalyzes the reversible isomerization of D-glucose 6-phosphate (G6P) to D-fructose 6-phosphate (F6P).
Sucrose synthase (Susy) triggers sucrose metabolism in the cytoplasm into UDP-glucose and fructose or fructose and glucose by neutral cytoplasmic invertase[71,72]. Susy and hexokinases mobilize carbon from sucrose for the hexose phosphate pool, while the invertase-led metabolized sucrose is usually fated for vacuolar storage[73,74]. Susy and invertase activities are critical determinants of fruit sink strength[43,72]. Hexose phosphates are critical for starch synthesis, usually before 13 days post-anthesis overlapping with peak mitotic activity in the fruit[74−76]. Starch accumulation peaks at ~40 days post-anthesis (DPA) and subsequently degrade upon ripening[77,78]. Sucrose phosphate synthase (SPS) and Susy causes sucrose re-synthesis during fruit growth[79,80]. However, enzymes contributing to the degradation of starch during re-synthesis are still unknown[81−83]. Substrate cycles between sucrose and starch offer flexibility to preserve the fruit as a carbon sink[84].
Sucrose is either imported through the symplast or 'hexosized' into the apoplast for further import into the cell[44,72,85]. Sucrose and hexose can be stored in the vacuole, with sucrose flux occurring from anthesis to approximately 20 – 25 DPA[44,85,86]. The intermediates of hexose phosphate are then imported into the plastid to synthesize starch (Fig. 1). Susy and hexokinase activities are decreased compared to invertase, and apoplastic imports of hexose are increased when sugar is stored in the vacuole. Starch biosynthesis is minimal, and active starch degradation may enhance storage sugar content at this stage[85].
-
Invertase is a significant macromolecule for fructose and glucose hydrolysis. Invertase occurs in isoforms with varying biochemical characteristics and subcellular localizations[6]. Based on current knowledge, they are mainly involved in regulating sucrose transport to distinct tissues in crucial development processes such as carbohydrate partitioning, plant response to abiotic and biotic stresses unassisted or in conjunction with phytohormones[43,67]. Every isoform of invertase in the fruit–apoplast, cytoplast, and vacuole is independently and jointly inhibited. The modified LIN5-a cell wall invertase is the foundation for high total soluble solids (TSS) in S. pennellii[60]. Higher TSS is correlated with increased starch metabolism in wild tomato; however, starch alteration is not a universal route to high TSS[73,87].
Invertase compartmentalization as a precursor protease vesicle (PPV) introduces a new dimension of invertase regulation in vivo[88,89]. PPV sequesters some vacuolar invertase for release into acidified vacuoles[82−84,90]. It is unclear yet whether this invertase is active within PPV of acidic sucrose-confining vacuoles before vesicular fusion. Vacuolar invertase may be regulated by cell wall-associated kinases (WAKs)[91,92]. If WAKs are dysfunctional, vacuolar invertase activity in the roots decreases to less than 50%, and under low-osmolyte conditions, growth is inhibited[10]. The WAKs are ideally used as status indicators for the interface between the plasma membrane and cell wall as each WAK has an extracellular (possibly pectin) N-terminus and a cytoplasmic (signaling) C-terminal serine/threonine kinase domain[10].
Plants display sessile life-forms and possess well-developed regulatory mechanisms to respond to environmental stress[2,93]. These processes are mainly triggered by the transport of assimilates from source tissues into sink tissues under strict molecular control using different sugar transporters and invertases[75,94,95]. The carbon source tissues are the exporters of net sugars while the carbon sinks are the net importers of sugars. For example, matured leaf mesophyll cells are the primary reservoirs for carbon fixation and are considered photosynthetically active carbon source(s)[67]. Ho[96] reported that storage sinks might store imported photosynthates in specific organs (seeds) and use the roots or meristems to import carbohydrates to sustain growth and development. Variations in carbon source tissues usually limit the transport of carbohydrates, hormones, and regulatory apparatuses that activate a response to adverse conditions[46].
Based on their biochemical properties and subcellular localization[97−99], invertases are classified as acid/vacuolar, neutral/cytoplasmic, and extracellular invertases[80,93]. Invertase and other sucrose cleaving enzymes determine carbon sink strength as they form the sucrose gradient used to transport sucrose from phloem under a steeped concentration gradient[44,93]. We have summarized functions of invertase based on recent functional genomic studies on this family[43,48,49,71,81,86,97,100], which underlines the role of extracellular invertase in intermediate defense responses, fruit flavor, and plant development processes (Fig. 2).
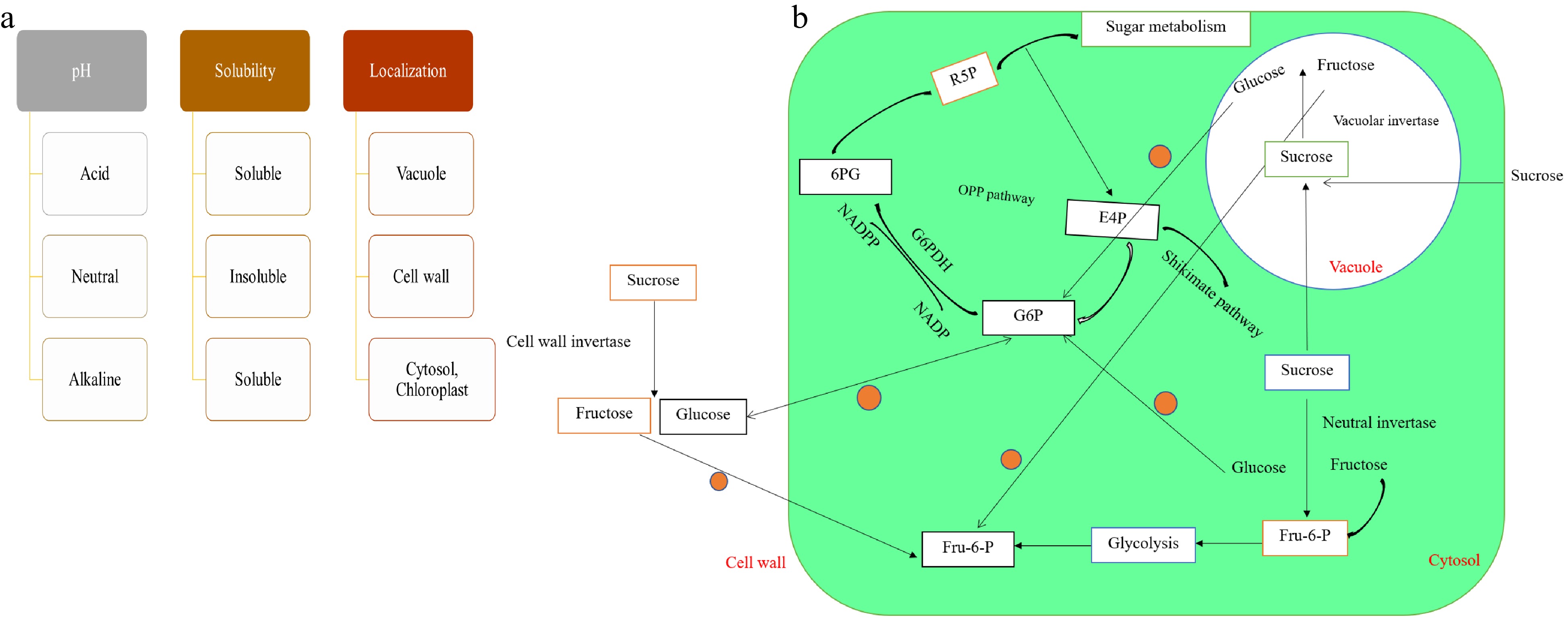
Figure 2. (a) Classification of invertase based on pH, solubility, and subcellular localization. (b) Functions of invertase, role in sucrose metabolism and their biosynthetic pathways in tomato. The mitochondrial NADPH pool is maintained by oxidative pentose phosphate pathway (OPP pathway) and glucose supply when mitochondria are subjected to oxidative stress. Glucose 6-phosphate (P6G) is a glucose molecule that has its hydroxyl group on carbon 6 phosphorylated. Due to the fact that the vast majority of glucose that enters a cell gets phosphorylated in this manner, this dianion is quite frequent in cells. The pentose phosphate route produces ribose 5-phosphate (R5P), which is also used as an intermediary. The formation of ribulose 5-phosphate is the final stage in the oxidative processes in the pentose phosphate pathway. Adapted from Chibbar[44], Huang et al.[10], Fotopoulos[57], Roitsch and González[93], Tauzin and Giardina[66].
-
In tomato, the Glycoside hydrolase family 32 (GH32) includes enzymes that are associated with invertase/fructofuranosidase (EC: 3.2.1.26); inulinase (EC: 3.2.1.7); levanase (EC: 3.2.1.65); exo-inulinase (EC: 3.2.1.80); sucrose: sucrose 1-fructosyltransferase (EC: 2.4.1.99); and fructan: fructan 1-fructosyltransferase (EC: 2.4.1.100)[100]. Invertase is organized functionally in gene families comprising various organ and growth-specific isoenzymes[57,97,98]. Godt and Roitsch[65] cloned and characterized four isoenzymes (LIN5, LIN6, LIN7, and LIN8) of the tomato extracellular invertase gene family. Twenty-four invertase genes were recently identified comprising nine invertase genes localized in the cell wall (CWINV), two localized in the cell membrane (CMINV), 11 were localized in the chloroplast (ChlINV), one each localized in the cytosol (CyINV), and vacuole (VaINV) distributed on eight of the 12 chromosomes (Fig. 3) of the tomato genome[94,101]. We also detected 143 invertase genes from four different plant species (Fig. 3) and constructed a phylogenetic tree using the FastTree MAFFT plugin in Geneios 11 and clustered the 143 (five species) invertase genes into six clades, one with a possible pseudogenic outgroup (Fig. 3). The combined 143 invertase genes were clustered into six clades of dicotyledonous species (A. thaliana, Capsicum annuum, S. tuberosum and S. lycopersicum) and one monocotyledonous species (Z. mays) (Fig. 3). Clade 1 includes genes from three dicots, with clades 2, 3 and 4 mostly containing monocots and dicots (Arabidopsis, pepper, tomatoes and maize), while clades 5 and 6 were mostly dicots (Solanaceae). Invertase genes in clade 6 were vacuolarly localized with possible tomato pseudogene (Solyc10g061980, this gene may have lost its conserved domain with no functional annotation in the tomato genome (see Fig. 3, clade 1)). Thus, the divergence of invertase underlines their different functions in tomato plant. Invertase is reported to regulate stress, organ developments and carbohydrate metabolism in plants[93]. Specifically, our phylogenetic analysis of invertase genes from tomato clustered them into three subcellular-specific clades (chloroplast, cell wall, and vacuole)[79] (Fig. 3). Similar dichotomies between cell-wall invertases from monocots and dicots have been reported[101−104].
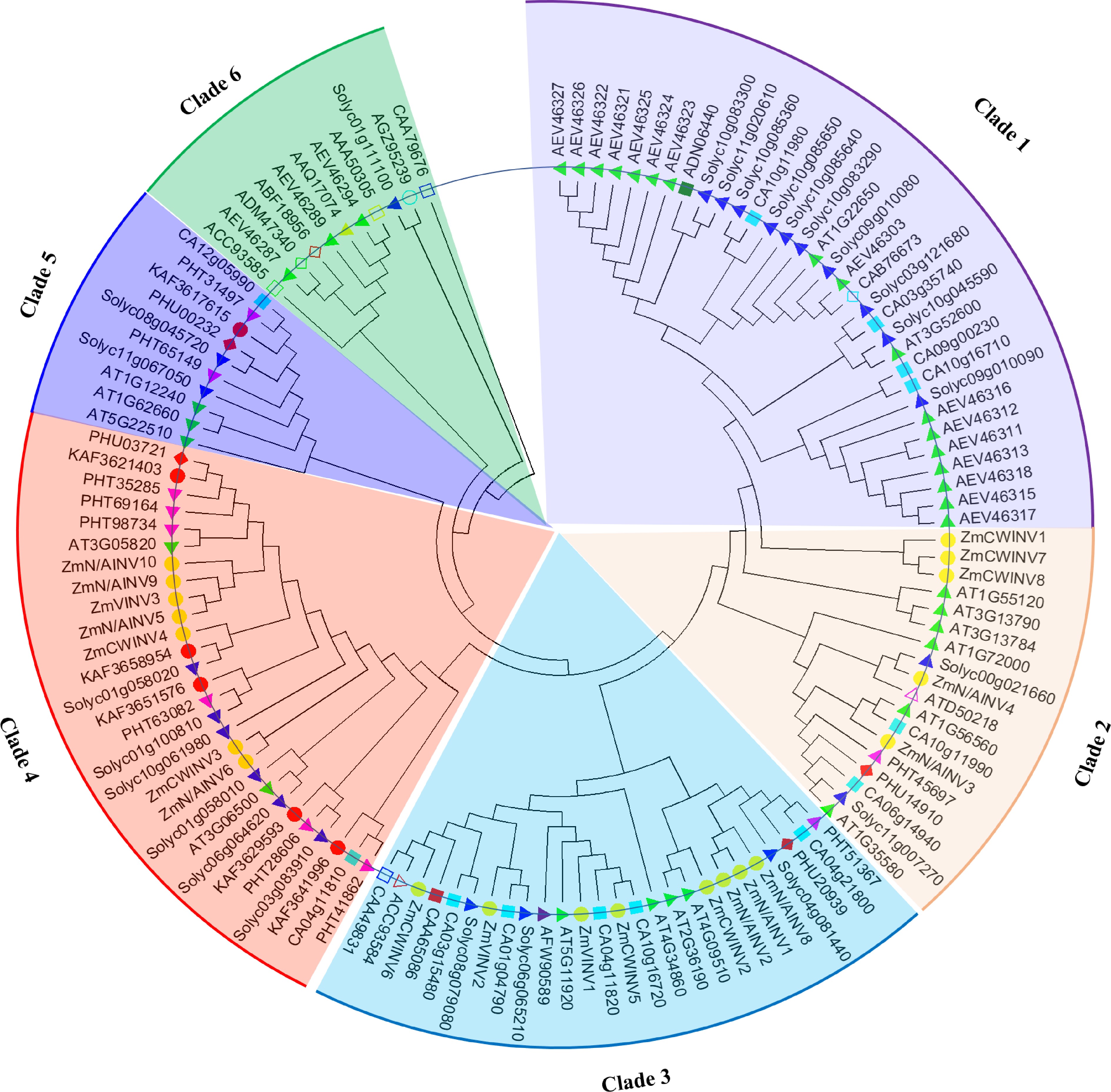
Figure 3. Evolutionary divergence and relationships of tomato invertase genes within plant species. Arabidopsis thaliana (AT), Solanum lycopersicum (Sl or sol), Pepper (CA), Zea mays (Zm), Solanum tuberosum (AE), Potato (AA, PHU or PHT).
Several gene families originated either by duplication or by evolutionary divergence from common ancestry. Approximately 41% and 77% of the predicted proteins in Arabidopsis thaliana[5] and Oryza sativa[105] are members of different gene families. Four invertase genes localized in the cell wall and vacuole (Atβfruct1, Atβfruct2, Atβfruct3, and Atβfruct4) in Arabidopsis thaliana (L.) were identified and functionally studied by Tymowska-Lalanne and Kreis[106]. The four invertase genes localized in the cell wall and vacuole were expressed in developmental processes in an organ-specific manner. Rice invertase genes comprising eight alkaline/neutral, nine-cell wall, and two vacuolar invertases were identified by Ji et al.[116]. In the vacuolar group, a sequence similar to a complete N-terminal motif that targets alkaline phosphatase post-translationally in yeasts vacuolar membrane was replaced by the N-terminal signal peptide and co-directs cell-wall invertases into the endoplasmic reticulum for secretion[107]. The two invertase families co-evolved rapidly via gene duplication and gene loss, but more than ten intron losses occurred in the acid invertase family than one single intron gain in the alkaline/neutral invertase[108−115]. Yao et al.[70] cloned six cell wall invertase genes (MeCWINV1-6) in cassava and confirmed their carbon export regulation from source leaves and sucrose to hexose in the apoplast. In Populus, five invertase genes localized in the cell wall (PtCWINV1-5), three in the vacuole (PtVINV1-3), and 16 neutral/alkaline (PtNINV1-16) were identified on 14 chromosomes[116,117]. Again, the sugarcane genome has six neutral/alkaline (ShN/AINVs) and eight acid invertases (ShAINVs) as recently identified and functionally characterized by Wang et al.[118]. In maize, three vacuolar isogenes (InvVR), eight invertases localized in the cell wall (InvCW), and ten alkaline/neutral (InvAN) invertases were cloned in three different phases of maize development, with varied molecular characteristics in nine tissues[119]. In pepper, nine acid invertase genes were identified and functionally characterized[120]. These genome-wide functional characterizations of invertase were made possible by the availability of the fully sequenced genomes of these crops. Genomic approaches are being deployed to harness the potential of invertase for flavor improvement programs in tomato and other economically important vegetables[2,7,12,121,122].
-
Sugar unloading is an organized process in tomato fruits, and its pattern varies throughout fruit development. Sugar is primarily unloaded by the symplast in developing fruits[2]. There are several plasmodesmata and cell connections at this stage, but they gradually decline[123]. Only a small amount of sucrose is unloaded by apoplastic invertase and transported into the fruit cells by hexose transporters during this early stage of development[19,85]. Despite the fact that sucrose does not unload in the tomato pericarp until 35 days after anthesis, a putative role for apoplastic invertase has been proposed based on kinetic properties of a moderate QTL for brix index[124,125].
According to Kataoka et al.[126], the activation of vacuolar acid invertase and neutral invertase by gibberellic acid shortly after anthesis can increase the sink size of individual pericarp cells. Apart from Susy, acid invertase (EC 3.2.1.26) may also be involved in sucrose cleavage, implying that sucrolytic activity occurs not only in the cytosol but also within the vacuole[127−133]. According to Beauvoit et al.[85], acid invertase is responsible for the majority of sucrose cleavage in dividing cells, whereas cytosolic neutral invertase and SuSy are primarily involved in subsequent cell expansion phase.
During the later stages of fruit development, wild species frequently exhibit an increased import of sugar from source leaves[134−145]. Conversely, sucrose accumulation is restricted because invertase activity increases during ripening in cultivated tomato[30]. Since fructose is sweeter than other sugars, metabolic engineering was used to increase fructose content in commercial tomato fruits by using fructokinase targets[29,127]. Schaffer et al.[128] reported that the trait of high fructose to glucose ratio is inherited separately from sucrose accumulation. Several wild tomato species differ from domesticated tomato cultivars in their total soluble solid content, a convenient proxy for sugar content.
Wild relatives of S. lycopersicum could be a good source of genes for improving fruit sugar composition. Mutations in enzymes involved in carbon metabolism have been discovered in S. chmielewskii and S. habrochaites, resulting in specific sugar compositions. The sucr mutation in an invertase gene causes sucrose to replace glucose and fructose in the fruit of S. chmielewskii[129]. The S. habrochaites allele of the ADP glucose pyrophosphorylase enzyme was significantly more efficient than the allele found in cultivated species, resulting in a higher fruit sugar content[128]. The LIN5 gene, which encodes apoplastic invertase, has been found to have a QTL modulating sugar partitioning, with the S. pennellii allele producing higher sugar concentrations than the S. lycopersicum allele[130]. An invertase-like enzyme was identified in the S. pennellii acylglucose biosynthesis pathway, located in the trichome gland cells (Sopen03g040490). The enzyme functions on the pyranose ring–acylated acylsucroses that are found in wild tomatoes, but not on the furanose ring–decorated acylsucroses of cultivated tomatoes. The introduction of the metabolic enzymes led to the modification of the main acylsucrose biosynthetic pathway, which previously resulted in the loss of furanose ring acylation[131].
In fruit ripening, an important metabolic change from glucose to fructose preserves fruit flavor by removing the bitterness-inducing alkaloid, α-tomatine. GORKY, a nitrate/peptide family transporter mediating α-tomatine subcellular localization during fruit ripening, has a base deletion in which GORKY mediates the α-tomatine localization in the vacuole during fruit ripening, allowing the export of α-tomatine and its derivatives into the cytosol, which promotes the total conversion of the α-tomatine pool into non-bitter forms and thus, improving fruit flavor[8]. The QTL, Brix9-2-5 is associated with a single amino acid deletion in the third exon of the LIN5 gene (ASP348 to GLU)[132,133]. RNAi-silenced LIN5 significantly decreased fruit yield, lowered fruit size, seed size, and seed quantity[60]. Sugar metabolism alterations were mostly restricted to sucrose accumulation, with a simultaneous decrease in the amount of glucose and fructose. Silencing of the vacuolar invertase TIV1 gene resulted in reduction in fruit size, increased sucrose accumulation, and lower levels of hexose sugars throughout late development stage in tomato[31]. A chaperone-like DnaJ encoding gene, sugar partitioning-affecting protein (SPA) was extracted and cloned. Subsequent study showed that transgenic plants had a large increase in fruit weight, fruit per plant, and harvest index when SPA was silenced[64]. This SPA-attributed chaperone protein was found to influence the supply-to-demand carbon flow, governing phosphoglucomutase, sugar kinase, and invertase enzyme activities, which affects fruit development and consequently the harvest index[64].
-
Invertase regulates several facets of tomato growth either independently or jointly, ranging from gene expression to long-distance nutrient translocation, carbohydrate partitioning, growth processes, phytohormone, abiotic and biotic responses[2,29,39,62,93]. As a common characteristic in higher plants, carbon autotrophy plays a pivotal role in sucrose metabolism[93]. Carbohydrate synthesized in carbon source tissues are translocated into carbon sink tissues to maintain heterotrophic metabolism and growth or deposited as sucrose or starch[90−93]. The hydrolytic cleavage of sucrose into hexose monomers is made possible by invertase. Variations in carbon source-sink relationships define tomato growth and development such that regulatory mechanisms of photo-assimilates partitioning can be established in specific environments. There has been abundant evidence that sucrose and its cleavage products are required metabolic signals influencing diverse gene expression and regulation in tomato growth and fruit quality[72].
Nonetheless, recent work on the invertase route of sucrose usage has also established critical new regulatory mechanisms[39,44,134]. Li et al.[43] reported that high invertase activities increased sucrose imports into young tomato fruits. Sugar signaling regulates high sucrose import pathways and cell wall[43]. Different invertase family members show different response to varied stresses and activities in different organs[93]. Transcriptionally and post-translationally, invertase regulation is influenced by hormones, oxygen supply, pathogens, protein inhibitors, and sugars[135−139]. Predominant among the several mechanisms identified in the modulation of expression or regulation of invertase are differential transcript formation[43,71,74], exon-skipping[67,93,131], protein-inhibitor binding[85,93,132], and a recent compartmentalization-control and breakdown mechanisms[10] (Fig. 4). These regulatory mechanisms further indicate that invertase plays a vital role in sucrose metabolism under the influence of stressors[142,143].
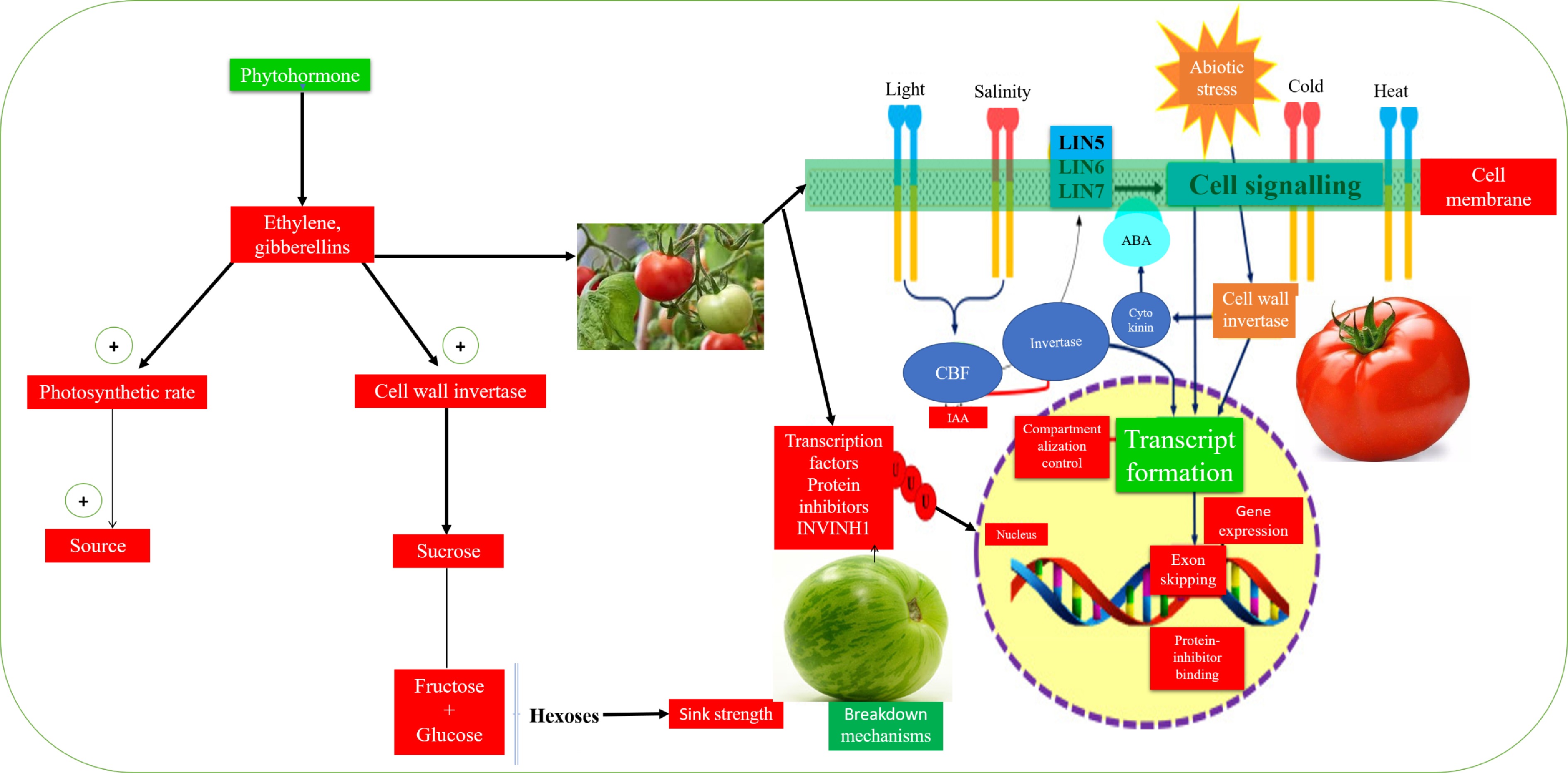
Figure 4. Key stress-induced invertase regulatory mechanisms in tomato fruit development. High invertase activity increases sucrose imports into young tomato fruits. Sugar signaling regulates high sucrose import pathways and cell wall. Phytohormones, protein inhibitors, and other abiotic stressors influence invertase regulatory mechanisms. Prime invertase regulatory mechanisms under stress conditions range from differential transcript formation[43,71,74], exon-skipping[67,93,131], protein-inhibitor binding[85,93,132], compartmentalization-control and breakdown mechanisms[10].
It was recently reported that cold stress inhibits the transcription factor, invertase inhibitor-1 (INVINH1), and enhances cell wall invertase genes, LIN6 and LIN8 transcriptions in tomato[142]. Furthermore, silencing INVINH1 expression in tomato increases invertase activities and enhances cold tolerance[142]. As transgenic INVINH1 tomatoes were less involved in invertase expression, they were more susceptible to cold stress, glucose, fructose, and hexose production[142]. The in-vitro repression of the C-repeat binding factor genes (CBF) was regulated by INVINH1 or glucose[37]. Auxin has also been found to stimulate the activity of cell wall invertase[144]. Invertase activity correlates with the peak of indolyl-3-acetic acid concentration during stem cell development, whereas exogenous use of indolyl-3-acetic acid stimulates rising vacuolar invertase activity[145]. Likewise, there was partial evidence of the induction of cell wall invertase by abscisic acid (ABA) with a fusion between the LIN6 promoter and the glucuronidase reporter gene in tomatoes[93]. Cytokinins impair invertase activity which was evident in the high cell wall activity and cytokinin concentrations, especially in organs that overgrew[65].
Prior to the widespread use of genome editing technologies, RNAi-mediated gene suppression or identification of genes responsible for spontaneous mutations was frequently utilized for analyzing gene function in tomato[6,13,146]. Many important tomato genes in fruit development, ripening and stress responses have been functionally confirmed using genome editing-mediated targeted mutagenesis, mainly CRISPR/Cas9[147]. Several elements of tomato fruit flavor could be studied as recent results of CRISPR/Cas9-mediated re-evaluation of transcription factors and cell wall modifying enzymes in fruit ripening were promising[148−152].
Modern commercial tomato varieties deteriorate in flavor quality compared to heirloom varieties, which could be attributed to the long-standing focus on yield improvement in tomato domestication and breeding[141,147]. A large number of genes involved in fruit characteristics, including flavor, are revealed by a recent work on panSV genome, tomato pan-genome construction using 725 phylogenetically and geographically representative tomato accessions, and recent tomato genome sequencing[5]. In tomato breeding, genome editing has been used to produce mutants, and the majority of the mutants are knockout mutants in which a gene of interest has been inactivated[148−152,154,155]. Most loss-of-function mutants do not normally generate agriculturally-useful phenotypes[154]. Contrary, gain-of-function mutations caused by base substitution or targeted transgene insertion, have enormous promise for direct application in tomato flavor improvement[154]. Tomato mutants became more resistant to abiotic stressors when modified using the DNA base editor CBE, and CRISPR/Cas9 gene editing system[156−160]. The new prime editing methods, which can provide precision genome editing by installing desired substitutions and insertions, are being utilized in many research programs, employing chemically modified DNA as a donor in CRISPR/Cas9 which can significantly increase the effectiveness of flavor gene insertion in tomato[20]. Gain-of-function mutations resulting from genome editing have the potential to be used in variety of applications, including precision tomato breeding for improving flavor[154].
MYB12 mutation using CRISPR/Cas9 successfully enabled pink tomato fruit formation[4]. In addition, SP5 G for day-length responses[144−146], AGL6 for fruit parthenocarpy[147], SEP4 members for inflorescence architecture[144], RIN for fruit ripening[148] and SlALC for fruit shelf-life[149] were studied through mutants produced by CRISPR/Cas9 gene editing. The resulting gene-edited tomato plants exhibited altered phenotypes, somewhat close to those of cultivated tomato plants[149,151]. CRISPR/Cas9 genome editing can also be employed to study the regulation of invertase or flavor-related gene expression[161−165]. SlCLV3, S, and SP cis-regulatory regions were mutated, and several novel cis-regulatory alleles of these genes show different degrees of phenotypic changes in tomatoes[152]. These experiments provide demonstrable ways to obtain knock-in mutants for enhancing flavor and stress tolerance by replacing a promoter with increased expression of invertase or stress response gene.
-
It is evident that invertases play significant roles in sugar metabolism, fruit development, and flavor in tomato with several genes encoding their proteins. The use of functional genomic approaches has improved our knowledge of invertases. Despite these milestones, specific important questions remain unanswered, such as why are invertases found with varied properties in different subcellular compartments? and, how do these enzymes interact with each other to regulate total soluble solids and flavor in tomato?
Multiple phosphatases and kinases are involved in invertase regulation, which is compatible with the variety of their signaling networks. Different mechanisms can also mediate opposing reactions to sugars and invertase gene activity, with some sugar-repressed invertases usually involved[153,161−165], but not in the usual kinetic forms induced by sugar[58,166]. Many modifications in the tomato genome have been created, using gene editing approaches for cultivar-specific changes. The broad application of CRISPR/Cas editing system in tomato breeding allows us to engineer invertase or pyramid targeted QTLs to accelerate new genotypes with improved flavor and fruit quality. CRISPR/Cas-mediated engineering holds the potential to boost invertase activity or sugar metabolism to enhance tomato fruit flavor and quality.
Increasing molecular marker development is useful for marker-assisted breeding for both qualitative and quantitative traits in tomato[163−165]. Nevertheless, while in practice, markers have been used widely to enhance genetically-inherited tomato traits, but they are yet to be utilized to develop complex traits such as flavor in tomatoes. With recent developments in tomato genome and transcriptome sequencing, modern PCR-based markers such as single nucleotide polymorphisms (SNPs) can be produced for improving fruit flavor in tomatoes. More markers are also expected to be available through new sequencing and genotyping technologies such as genotype by sequencing (GBS). Further research may be needed to identify invertase allele- and population-specific markers to expand marker-assisted selection in breeding for flavorful tomatoes.
- This work was supported by grants from the National Key Research and Development Plan of China (2018YFD1000800); National Natural Science Foundation of China (31991182; 31972426); Hubei Hongshan Laboratory Project (2021hszd007); Wuhan Frontier Projects for Applied Foundation (2019020701011492).
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ahiakpa JK, Karikari B, Magdy M, Munir S, Mumtaz MA, et al. 2021. Regulation of invertase and sucrose for improving tomato fruit flavor: A review. Vegetable Research 1: 10 doi: 10.48130/VR-2021-0010

Regulation of invertase and sucrose for improving tomato fruit flavor: A review
- Received: 19 August 2021
- Accepted: 23 November 2021
- Published online: 14 December 2021
Abstract: Tomato (Solanum lycopersicum L.) is a commercially farmed vegetable belonging to the Solanaceae family, the third most important vegetable after potato (Solanum tuberosum L.) and onion (Allium cepa L.). It is cultivated for its fresh fruits and processed paste, with over 153 million metric tons of global production. However, modern tomato cultivars have limited sugars, acids, and volatiles allelic diversity as flavor has generally been less prioritized in breeding programs. Invertase is an essential regulator of flavor and sugar metabolism in tomato. Genetic control of tomato flavor is still incomplete without a clear understanding of the roles of invertase and sucrose metabolism. This review provides an overview of our current understanding of the invertase mode of action in sucrose metabolism, their evolutionary and functional divergence in the tomato genome, role in stress response, genetic and hormonal control of fruit flavor and quality. We summarized the primary roles of invertase in sugar metabolism and fruit flavor.
-
Key words:
- Solanum lycopersicum /
- Solanaceae /
- Cell wall invertase /
- Fruit taste /
- Total soluble solids