








-
Soybean (Glycine max (Linn.) Merr.) is an important food and oil crop[1], which is not only a significant source of edible oils and proteins for humans and animals, but also widely used in industrial fields such as biofuel production[2]. Mutagenesis refers to the use of physical and chemical means to induce genetic mutations in organisms under artificial conditions, and through artificial screening and identification, ultimately achieving the breeding of new varieties or germplasm resources[3]. Spontaneous mutations occur in nature at a very low frequency (~10−6)[4], whereas mutagenesis has a mutation frequency that is hundreds of times higher than spontaneous mutations, and is capable of inducing a wide variety of types of mutant genes[5]. Physical mutagenesis is one of the most used modalities in the field of mutation breeding[6] and includes both ionising and non-ionising radiation, such as X-rays, gamma rays and beta rays[7]. X-rays have a wavelengths range from 0.001 to 0.01 nm[8], and are an electromagnetic wave generated by accelerated electrons hitting a metal target without the need for a radioactive source, and have the characteristics of easy operation and low cost of use[9]. X-rays have been recognised as a common means of mutagenesis since 1928 when Muller, an American geneticist, experimentally demonstrated that exposure to X-rays caused genetic mutations[10,11]. X-rays were then considered a common means of mutagenesis. Soon after, X-rays were also used in plant research, and Stadler demonstrated for the first time that X-rays could induce mutations in maize and barley[12]. Chlorina was the world's first tobacco mutant variety successfully bred by scientist Tollene using X-rays, and it was successfully released in the 1930s[13]. Now, the IAEA's Mutant Variety Database (
https://mvd.iaea.org , accessed 18 October 2024) contains more than 3,448 officially registered mutant varieties, of which 539 were developed directly by X-ray irradiation, and about 50% by direct or indirect use of gamma rays, compared to less than 1% of the varieties produced by ion beam irradiation[14]. The history of irradiation mutagenesis breeding in soybean can be traced back to 1931, when scientists in the former Soviet Union used X-rays as a mutagenic source to treat soybean seeds and soybean plants that had already flowered, pioneering the use of X-rays in irradiating soybeans[15].Soybean is a strictly self-pollinating crop with less than one percent natural self-pollination[16]. In the last decade China started to rely on importing soybeans from other countries[17]. The breeding, and application of high-quality, high-yielding, stress- and disease-resistant soybean varieties is a fundamental prerequisite for the development of the soybean industry. Although many reports have been published on the effects of radiation mutagenesis on soybean phenotypes, there is limited information on the effects of electron accelerator treatments at the level of plant gene sequences. Therefore, understanding the mechanisms of plant response to radiation is essential for improving plant resistance and productivity. To study the mutagenic effects of different ionising radiation-directed genomes, we irradiated different varieties of soybeans using an electron accelerator and collected samples from irradiated M1 generation soybeans, as well as from unirradiated controls, for genomics testing and analysis. The electron accelerator is the first electron accelerator in China and even in the world to combine linear electron accelerator technology with agricultural breeding technology, and it is the first electron accelerator used for crop mutation breeding in China. This study relies on the X-rays generated by the electron accelerator to carry out mutagenesis research on soybeans. Taking the mutagenic treatment of soybean seeds as a starting point, this study aims to gain an in-depth understanding of the mutagenic effects of soybeans at the genome level after being irradiated by the electron accelerator, to provide a theoretical basis for the innovation of soybean germplasm resources and radiation breeding.
-
Soybean seeds (Tianlong No.1 and Williams 82) with full grains and uniform size were selected for radiation mutagenesis treatment. The samples were irradiated using a BVE-2MV electron accelerator from Sanya Mingyao Germplasm Innovation Technology Co. Ltd (Sanya, Hainan, China), which has a maximum energy of 2.0 MeV, a rated current/dose rate of 0.5 mA/Gy/h, with radiation doses set at 180 Gy and 200 Gy. The irradiated soybeans were sown in a field in Yazhou District, Sanya City, Hainan Province, China (109° E 18° N).
Hyper-seq library construction and sequencing
-
The DNA of soybean seedlings was extracted by the magnetic bead method using the following steps: (1) Sampling and grinding. Add about 0.1g of healthy and disease-free soybean leaves to a deep well plate (96 wells), and add grinding steel beads and liquid nitrogen to fully grind the samples, so that the samples appear powdery; (2) Water bath. Immediately add 600 μL of GEA1 to the ground powder, and then heat the sample in a water bath at 65 °C for 15 min add 200 μL of GEA2, and then leave it at room temperature for 15 min; (3) Centrifugation. Centrifuge at 4,000 rpm for 45 min, transfer 400 μL of supernatant to a new deep-well plate, and add RNase A; (4) DNA extraction. Put the deep-well plate into the automatic nucleic acid extraction robot and add ddH2O, GEW1, GEW2, GEB, and magnetic beads for DNA extraction according to the method; (5) Concentration detection. Evaluate the integrity of DNA samples by 1% agarose gel electrophoresis, use Qubit to detect the concentration of DNA samples, and store it in the refrigerator at −20 °C; (6) Library construction and sequencing. The libraries were then constructed from the population DNA samples using Hyper-seq[18] technology and the Illumina NovaSeq 6000 platform from Shanghai Personal Biotechnology Co. Ltd (Shanghai, China) was used for sequencing.
Variant detection and mutation site selection
-
The raw data were quality controlled and filtered by fastp[19] and the clean data obtained was mapped to the reference genome of soybean (GCA_030864155.1) obtained from the National Center for Biotechnology Information (NCBI). Duplicate sequences were removed from the BAM format file using Burrows-Wheeler-Alignment[20] tool and SAMtools[21] using PicardMarkDuplicates, and then detected and screened for SNPs and InDels (small insertions and deletions) using the Gatk[22] analysis process for the detection and screening of SNPs and InDels, When the variant types of the treated material differed from those of the control group, the site was selected as a mutation site using vcftools[23]. After the above steps, all variant sites were annotated with SnpEff[24]. A total of 321 datasets were obtained, including 161 samples of the Tianlong No.1 M1 generation (59 samples of TL180 Gy, 86 samples of TL200 Gy, and 16 control group samples), as well as 160 samples of the Williams 82 M1 generation (64 samples of W180 Gy, 85 samples of W200 Gy, and 11 control group samples).
-
Preliminary trials were conducted with the Tianlong No.1 soybean cultivar, employing radiation gradients of 180, 200, and 220 Gy. Each treatment group comprised 120 seeds. Following a 53-d growth period, the lethal rate was assessed, and it was found that the mortality rate at 180 Gy and 200 Gy was 57.5%, which was approximately the lethal dose (Fig. 1c). After most of the soybeans reached the R5−R8 stage, the height of soybean plants did not continue to increase. The plant height of soybean was measured at the R8 period, and it was found that after radiation treatment, the height of the irradiated group's plants was significantly changed compared with that of the control group, and the plant height showed a dwarfing state in general (Fig. 1b, d), of which the shortest average plant height of soybean in the 200 Gy irradiation group under three irradiation gradients was only 11.33 cm, and the size of the leaves gradually became smaller with the increase of the dosage (Fig. 1c), which indicated that the growth of soybean was inhibited by electron accelerator irradiation. The selection of irradiation dose is particularly important in irradiation mutagenesis breeding, to ensure a sufficiently large screening population with a high mutation rate, the semi-lethal dose (LD50) is usually selected as the optimal irradiation dose[25,26], so it is considered that 180 to 200 Gy is a more suitable irradiation dose for soybean seeds, 180 and 200 Gy were used in the subsequent experiments.
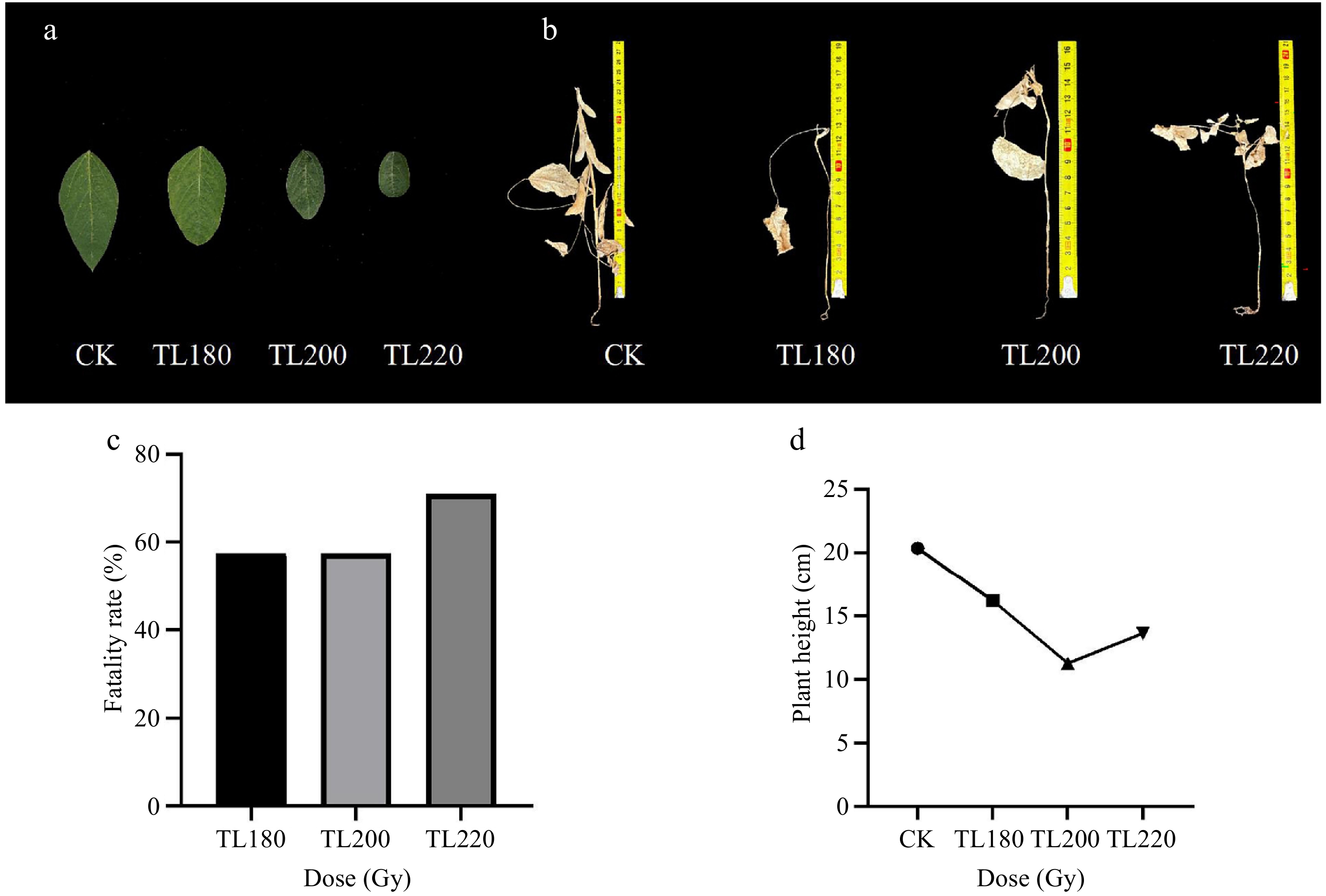
Figure 1.
Impact of different radiation doses on the growth of soybeans. (a) Size of leaves at each dose. (b) Plant height at each dose. (c) Statistics of lethality at each dose level. (d) Average plant height at each dose level. TL180 refers to the Tianlong No.1 soybean population at 180 Gy, TL200 refers to the Tianlong No.1 soybean population at 200 Gy, and TL220 refers to the Tianlong No.1 soybean population at 220 Gy.
Molecular characterization of induced mutations in different soybeans by different irradiation doses
-
In this study, we analyzed in detail the data from 321 samples of two soybean varieties, Tianlong 1 and Williams 82, at different doses. The molecular characterisation of electron accelerator-induced mutagenesis was further investigated for the four M1 generations, and a total of 1,002,604 mutation sites were detected, including 871,817 SNPs, 67,451 INSs, and 63,336 DELs, with the number of SNPs accounting for about 86.96%. Further analysis revealed that the distribution of these variant loci at the genome-wide level showed heterogeneity (Fig. 2). SNPs were the most abundant mutations in each treatment group, accounting for about 85% to 90% of the four M1 populations, followed by INS at about 5.3%−7.3% and DEL at 4.50%−7.43% (Fig. 3a). The number of mutation sites induced by increasing radiation dose gradually increased from 180 to 200 Gy, but at the same radiation dose, Williams 82 soybeans produced more mutations, and the average number of mutations produced by Williams 82 soybean varieties was 749 more than that produced by Tianlong No.1 soybean varieties at 180 Gy, and 3,561 more than that produced by Tianlong No.1 soybean varieties at 200 Gy. Under the same varieties, Tianlong No.1 soybean had 611 more mutant loci at 200 Gy than the average mutation produced, and Williams 82 soybean had 3,422 more mutant loci at 200 Gy than the average mutation produced at 180 Gy (Fig. 3b; Table 1).
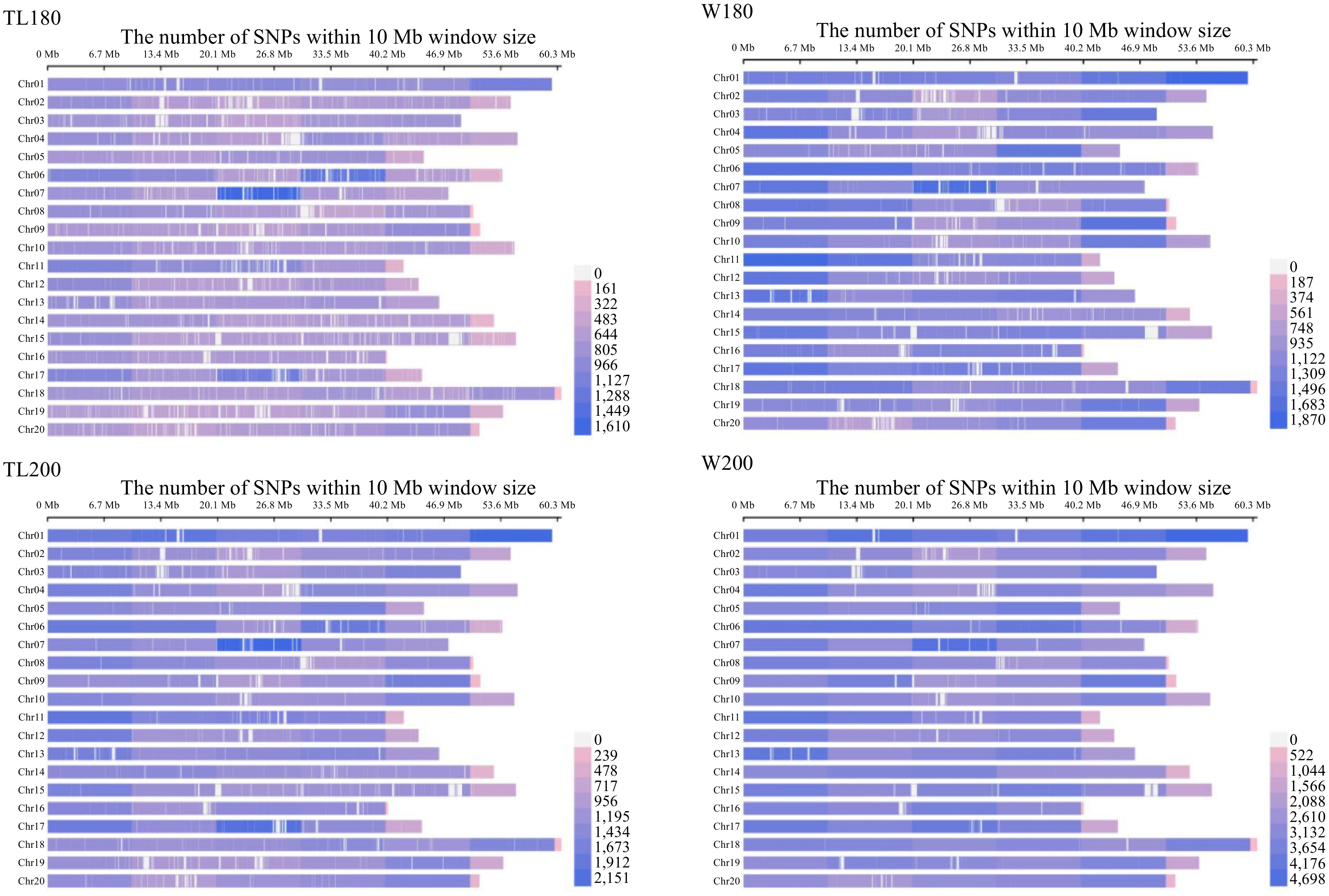
Figure 2.
Distribution of genomic mutations induced by an electron accelerator at various dose levels. TL180 represents the Tianlong No.1 soybean population at 180 Gy, TL200 represents the Tianlong No.1 soybean population at 200 Gy, W180 represents the Williams 82 population at 180 Gy, and W200 represents the Williams 82 population at 200 Gy.
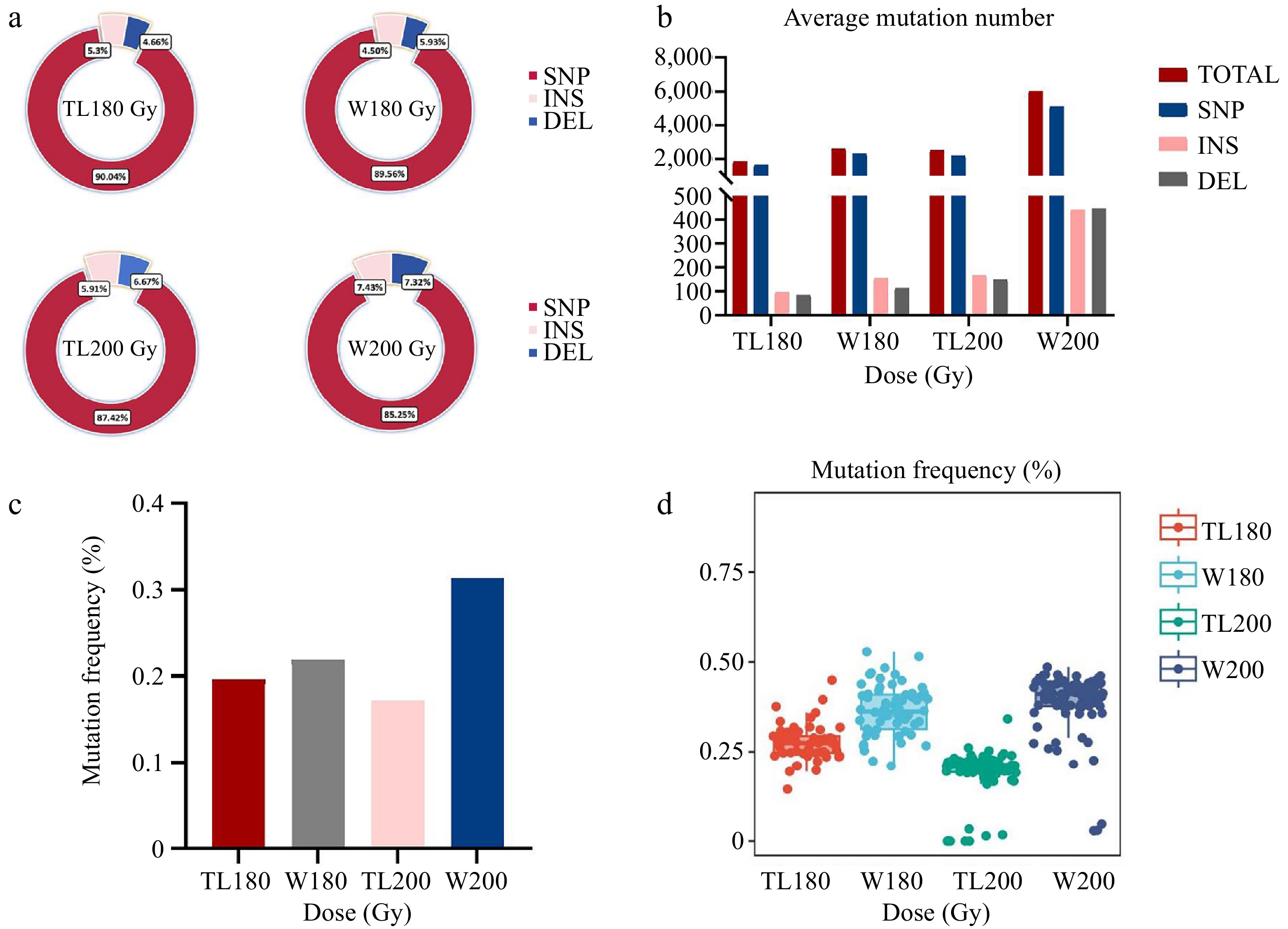
Figure 3.
Distribution of genomic mutations induced by an electron accelerator in the M1 population at each dose level. (a) Proportion of SNPs, INSs, and DELs in different dose treatment groups. (b) Average number of mutations induced in different dose treatment groups. (c) Average mutation frequency in different dose treatment groups. (d) Mutation frequency in individual samples induced at each dose level.
Table 1. Soybean M1 generation mutation information statistics.
Sample Number Total loci Variant loci Mutation frequency % Average SNPs Average insertions Average deletions TL180 59 55,938,993 109,878 0.1964 1676.86 98.64 86.83 W180 64 75,979,433 167,151 0.2200 2339.14 155.00 117.59 TL200 86 123,112,699 212,685 0.1728 2161.94 164.87 146.27 W200 85 163,648,886 512,890 0.3134 5144.12 441.55 448.33 The average number of mutations in Tianlong No.1 soybean was 1,862 at 180 Gy, with an average mutation frequency of 0.1964% (total number of mutated loci/total number of loci of all samples combined at this dose), and the average number of mutations at 200 Gy was 2,473, with an average mutation frequency of 0.17%. Williams 82 soybean had an average number of mutation 2,611 and an average mutation frequency of 0.22 % at 180 Gy, and an average mutation number of 6,034 and an average mutation frequency of 0.31 % at 200 Gy (Fig. 3c). The single sample with the highest mutation rate in Williams 82 soybeans at 180 Gy was W180-91 with a mutation frequency of 0.53% (number of mutations in a single sample/total number of loci in that sample), and the single sample with the highest mutation rate in Tianlong No.1 was TL180 Gy-83 with a mutation frequency of 0.45%. The single sample with the highest mutation rate in Williams 82 soybeans at 200 Gy was W200 Gy-27 with a mutation frequency of 0.49%, and the single sample with the highest mutation rate in Tianlong No.1 was TL200 Gy-83 with a mutation frequency of 0.34% (Fig. 3d).
Analysis of gene structure mutations in soybean M1 at different doses
-
We conducted an exhaustive analysis of genetic variations to investigate the distribution patterns of gene mutations induced under different varieties and radiation dosages within the gene structure. The results showed that the distribution pattern of mutations in the gene structure showed a high degree of consistency and did not produce significant differences, regardless of the dose or species. Specifically, the intergenic region became a hotspot for mutation occurrence, occupying about 30% of all detected mutations, followed by about 21% in the downstream region, 18%−19% in the upstream region, about 12% in the intronic region, and about 11% in the exonic region (Fig. 4a). Transitions (Ts) and Transversions (Tv) are a measure of how often SNP-specific types of mutations occur. We found that the Ts/Tv ratios ranged from 2.12 to 2.43 in the irradiated experimental groups of the four M1 populations (Fig. 4b). Thus, Ts was more common than Tv in the soybean M1 generation after irradiation by the electron accelerator, with C/G to T/A being the most abundant.
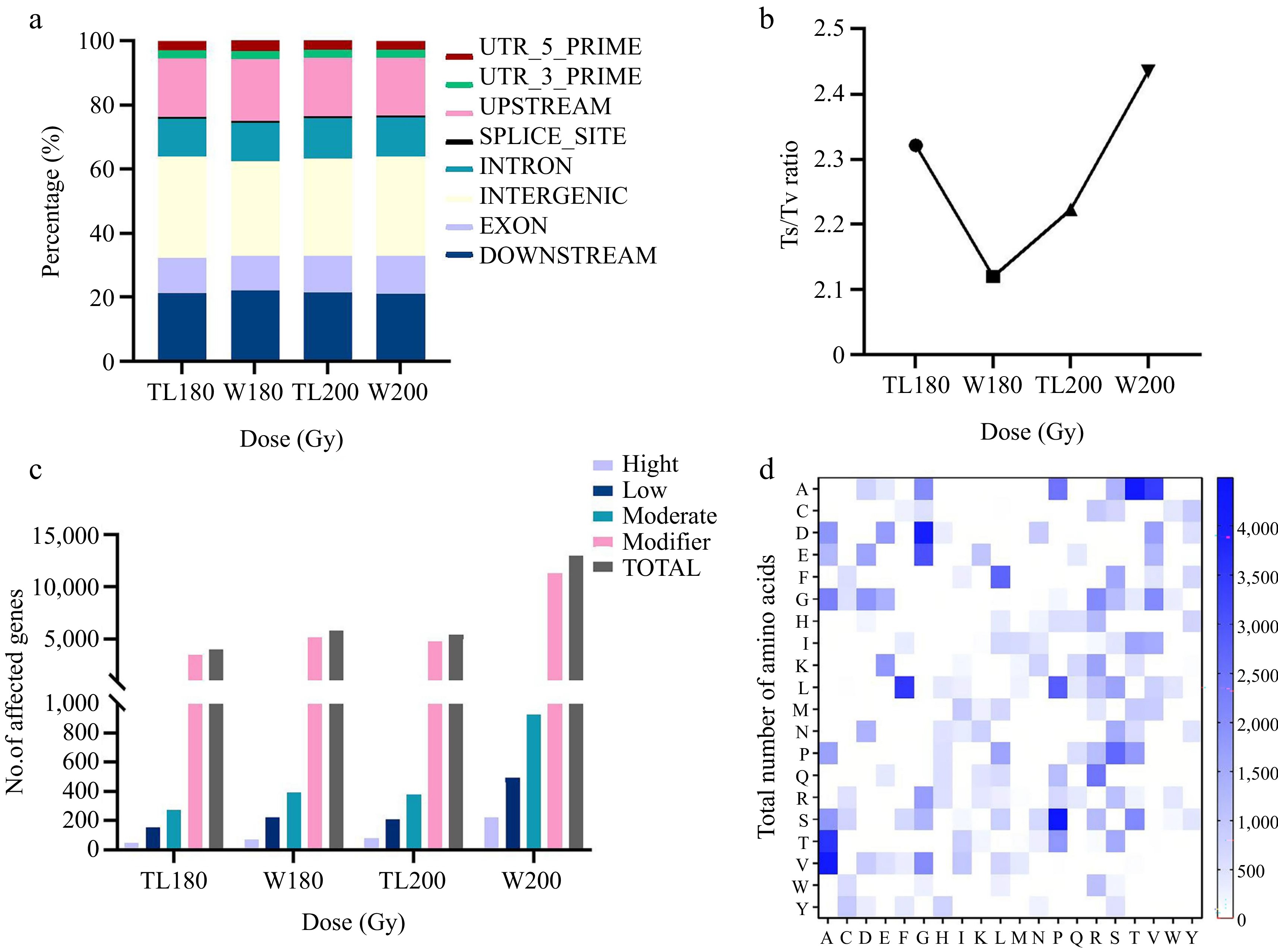
Figure 4.
Comprehensive analysis of mutations induced by various dose treatment groups. (a) Distribution of mutations in gene structure across different doses. (b) Ts/Tv ratio at various doses. (c) Average number of genes affected by mutations at different doses. (d) Total number of amino acid alterations induced at various doses.
In this study, we conducted a comprehensive mutation analysis on a total of 321 samples from four M1 populations, and Table 2 shows the number of mutated genes induced under different dose treatment groups. A total of 2,172,206 genes were identified as being affected by mutations to varying degrees. Of these, 33,236 genes were highly affected (high (destructive) impact on proteins, potentially causing protein truncation, loss of function, or triggering nonsense-mediated degradation). 152,851 genes were moderately affected (a non-destructive variant that may alter the effectiveness of the protein). 83,671 genes were only lightly affected (the variant is considered mostly innocuous or unlikely to alter the behaviour of the protein), and in addition, 1,902,448 genes were labelled as modifier types (Table 2), which usually refers to variants affecting non-coding regions or variants affecting non-coding genes, which tend to be more difficult to predict their effects (see
http://snpeff.sourceforge.net for details), with more genes affected in the Williams 82 soybean variety at the same dose (Fig. 4c).Table 2. The number of mutated genes induced under different dose treatment groups.
Sample High Low Moderate Modifier Total TL180 2,800 9,138 16,249 206,267 234,454 W180 4,563 14,274 25,191 327,616 371,644 TL200 6,812 17,991 32,613 408,042 465,458 W200 19,061 42,268 78,798 960,523 1,100,650 Changes in amino acids induced in M1 plants by different doses
-
In the four M1 electron accelerator irradiation groups we further analyzed the amino acid variation due to radiation-induced SNPs. We observed that a total of 298 amino acid variants were induced in the four M1 groups, accounting for 74.5% of all possible amino acid variants (Fig. 4d). These data show that the pattern of amino acid mutations induced under different doses of irradiation from the electron accelerator exhibits a high degree of consistency, suggesting that the irradiation dose, while affecting the number of mutations, has little effect on the specificity of the type of mutation. The most abundant amino acids change as follows: proline (P) to serine (S), threonine (T) to alanine (A), alanine (A) to valine (V), glycine (G) to aspartic acid (D), alanine (A) to threonine (T), and so on. It is worth noting that there is a correlation between the irradiation dose and the number of amino acid variants, which shows an increasing trend as the irradiation dose increases. A total of 280 times amino acid mutations were induced in Tianlong No.1 soybean at 180 Gy and 393 times amino acid mutations were induced at 200 Gy. A total of 402 times amino acid mutations were induced in Williams 82 soybean at 180 Gy, and 951 times amino acid mutations were induced at 200 Gy.
-
Through mutation breeding, new varieties that are rare in nature or difficult to obtain through conventional breeding can be produced[27]. Radiation mutagenesis has many advantages such as promoting genetic recombination, increasing mutation rate, breaking trait chains, overcoming plant self-incompatibility, shortening the breeding period, and facilitating gene transfer through long-distance hybridization[28,29]. Radiation mutagenesis has now been applied to several species including Arabidopsis, wheat, watermelon, perennial gentian, oilseed rape, chrysanthemum[30−35], and many other mutants that are difficult to obtain by other mutagenesis methods. The electron accelerator mutation breeding system is a high-tech piece of equipment specially developed and manufactured for mutation breeding, and it is the first electron accelerator used for crop mutation breeding in China, to date, only a few research studys have been done on it. Therefore, exploring the genomic mutagenic effect of soybean after irradiation by electron accelerator is not only conducive to the innovation of soybean germplasm resources, but also of vital scientific significance to the field of research on the application of electron accelerators in crop breeding. In this study, the mutagenic effects of Williams 82 and Tianlong No.1 soybean M1 generation treated by electron accelerator irradiation were comprehensively analyzed, and 180 and 200 Gy were considered to be more appropriate irradiation doses. Only SNPs and InDels induced by different soybean varieties at different doses were explored here, and other types of mutations were not addressed. We found that SNPs were the most abundant genomic mutation type in the M1 generation cultured by electron accelerator irradiation, accounting for about 85%−90%, while InDels accounted for less than 8% (Fig. 3a), with the C/G to T/A mutation type being the most abundant. This finding is similar to that obtained previously, where Shirasawa et al. performed whole genome resequencing using sodium ethyl methane sulfonate (EMS) and gamma-irradiation-induced tomato Micro-Tom, and found that single-nucleotide variants (SNVs) were the major mutations[36]. Xiao et al. identified a total of 1,076,010 SNPs and 183,421 InDels in aubergine with the most common mutation types being C/G to T/A and A/T to C/G transitions[37]. Yuan et al. radiated sorghum using fast neutrons, and SBS accounted for about 90.0% of the small mutations in the M1 generation[38].
The current study demonstrates that the mutation effect is influenced by not only the type of radiation source employed but also the specific mutagenic material and radiation dose. The ability to select appropriate mutagenic materials and doses for different species is crucial for increasing the mutation frequency and obtaining effective mutants in mutagenesis. Following the treatment of different soybean varieties with varying doses of radiation, the data were processed and it was found that the Williams 82 soybean variety exhibited greater sensitivity to radiation than the Tianlong No.1 soybean variety. In particular, the Williams 82 soybean variety exhibited a more pronounced radiation response when subjected to the same radiation treatment. This manifested as an intensification of genetic variation, with the average mutation rate of the Williams 82 soybean variety exceeding that of the Tianlong No.1. The number of mutation sites produced by the Williams 82 soybean variety was found to be 1.40 times higher than that of the Tianlong No.1 soybean variety at 180 Gy, and 2.44 times higher at 200 Gy. The Williams 82 soybean variety exhibited a significantly higher number of genes that were heavily affected at 180 Gy, with 4,563 genes affected, 1,763 more than the number affected in the Tianlong No.1 soybean variety at the same dose. At 200 Gy, the Williams 82 soybean variety exhibited 19,061 genes that were heavily affected, which represents 12,249 gene increases compared to the number of heavily affected genes observed in the Tianlong No.1 soybean variety at the same dose. The data indicate that the Williams 82 soybean displays a high degree of radiosensitivity following electron accelerator irradiation treatment. Therefore, it is important to consider variety specificity in radiation breeding and genetic improvement studies. For example, Li et al. determined the LD50 and growth of five varieties of Gladiolus gandavensis after being irradiated by X-rays to derive the radiation sensitivity of the different varieties[39], and Liu et al. used different doses of 60Co-γ rays on the macadamia nut varieties 'HAES900' and 'HAES951' in a fresh pollen irradiation treatment and found that the pollen of 'HAES951' was more sensitive to 60Co-γ ray irradiation[40].
In recent years, high-throughput sequencing technology has been developing continuously, which can provide rich mutation information at the genome level by sequencing means, and can provide rich mutation information at the genome level, providing a possibility for further in-depth study of mutation frequency and mutation spectrum of mutant groups. The results of sequence analysis can not only reveal the mutation frequency and mutation sequence characteristics of individual genes more reliably but also obtain genome-wide information on the composition and distribution of different types of mutations[41−46]. Hyper-seq sequencing and DNA mutation detection is a method that utilizes high-throughput sequencing technology for mutation detection and functional gene resolution. By sequencing a large number of samples simultaneously, it greatly improves the efficiency of mutation detection. In addition, mutation analysis using sequencing data can obtain more accurate mutation information, including SNPs and InDels, which greatly improves the high efficiency and accuracy of mutation detection. In this study, the high-throughput sequencing data obtained from Hyper-seq sequencing of a total of 321 samples from four M1 lines were enriched with data analysis and integration, and a total of 1,002,604 mutation sites were detected, including 871,817 SNPs, 63,336 DELs, and 67,451 INSs, with a maximum mutation rate of 0.53% in a single sample (Fig. 3d). Hyper-seq technology is characterized by its high throughput, rapidity, and ultra-low-cost features. By adopting a large-scale Barcode hybrid sequencing strategy, Hyper-seq technology can significantly reduce the sequencing cost, which is more than 50% lower than that of other simplified sequencing technologies and molecular breeding microarray technologies[47]. The use of Hyper-seq technology with the mutation detection technology system is a reliable method for detecting early mutants, and the detection of radiation-mutated soybean materials can quickly screen out mutant materials with DNA mutations. In the future, we can eliminate materials with no mutation or low mutation and keep the materials with high DNA mutation in the M1 generation to continue growing for the M2 generation, thus accelerating the breeding process.
-
This study investigated in detail the gene mutations induced by electron accelerators in soybeans. The results indicated that the electron accelerator is a powerful tool for soybean mutation breeding, with different varieties of soybeans exhibiting varying sensitivities to electron accelerator radiation, and that Hyper-seq is capable of rapidly detecting a wide range of mutations induced by electron accelerator radiation. This study identified that the radiation dose of the electron accelerator can benefit plant breeders in further optimizing doses and planning large-scale mutation breeding programs for soybeans, as well as for other crops. In the future, we plan to conduct high-throughput phenotypic and genomic analyses of the soybean M2 mutagenic population to further understand the mechanisms of radiation mutagenesis and their application in gene function analysis.
This study was supported by the Biological Breeding-National Science and Technology Major Project (2023ZD04073) and Hainan Yazhou Bay Seed Lab (B23YQ0002).
-
The authors confirm contribution to the paper as follows: study conception and design: Liang T, Xia Z, Zou M; material cultivation and collection: Liang T, He M, Xu R, Wang Q, Chen J, Yang L, Xia Y, Luo X, Zhu M; electron accelerator irradiation: Li W, Ren J; data collection: Liang T, He M, Xu R, Wang Q; analysis and interpretation of results: Liang T, He M, Xu R, Wang Q; draft manuscript preparation: Liang T, Xu R. All authors reviewed the results and approved the final version of the manuscript.
-
The genome sequence data reported in this paper have been deposited in the Genome Warehouse in China National Center for Bioinformation, under Accession No. PRJCA032391 that is publicly accessible at https://ngdc.cncb.ac.cn/gwh/.
-
The authors declare that they have no conflict of interest.
-
Received 15 November 2024; Accepted 22 January 2025; Published online 24 February 2025
-
SNPs were the most abundant type of genetic variation following irradiation with the electron accelerator.
Hyper-seq technology can be used to rapidly screen the radiation-induced soybean materials for DNA variants and accelerate the breeding process.
Under the same radiation treatment, Williams 82 soybean showed more obvious radiation response than Tianlong No.1 soybean.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liang T, He M, Xu R, Wang Q, Chen J, et al. 2025. Study on the mutagenic effects in the soybean genome accelerates breeding. Tropical Plants 4: e007 doi: 10.48130/tp-0025-0003 

Study on the mutagenic effects in the soybean genome accelerates breeding
- Received: 15 November 2024
- Revised: 18 January 2025
- Accepted: 22 January 2025
- Published online: 24 February 2025
Abstract: Soybean (Glycine max (Linn.) Merr.) is a significant grain, oil, and food crop, originating in China. It is of vital strategic significance for the development of the national economy. In this study, we used an electron accelerator to treat Tianlong No.1 and Williams 82 soybean varieties with different doses of mutagenesis treatments, and Hyper-seq technology was used to rapidly detect the mutation loci in 321 radiated soybean M1 generation materials, and a total of 1,002,604 variant sites were detected, including 871,817 single nucleotide polymorphisms (SNPs), 67,451 insertions (INSs), and 63,336 deletions (DELs), of which the number of SNPs accounted for about 87%. In addition, the Williams 82 soybean variety was more sensitive to radiation than the Tianlong No.1 soybean variety under the same radiation treatment and possessed more variable loci. Although soybeans originated in China, limitations in soybean varieties, planting patterns, and available planting areas have led to a situation where China's total soybean production falls short of meeting domestic demand. Consequently, imports have risen, posing a significant threat to China's food security. The results of this study provide a reference for the rapid screening of radiation mutagenesis materials, and through the in-depth analysis of the mutation loci, the response mechanism of soybean to radiation mutagenesis can be better understood, which will in turn accelerate the improvement of soybean varieties.