








-
Fungi comprise diverse lineages of heterotrophic eukaryote organisms which serve an array of key roles in nutrient cycling, plant and animal health, human well-being, and the food and pharmaceutical industries[1,2]. However, only about 5% of the estimated 2.2−3.8 million extant species of fungi have been described to date[3,4]. Traditionally, species identification was performed by trained mycologists through careful examination of the phenotypic properties. However, retention of shared ancestral character states and convergence of similar character states among unrelated species (i.e., unrecognized cryptic speciation) has led to a severe underestimation of species diversity[5,6].
Since Hebert et al.[7] first introduced the concept of DNA barcoding as a molecular tool for species identification, mycologists have explored genes with high primer fidelity and adequate resolution power across taxonomic levels. White et al.[8] were the first to propose the internal transcribed spacer (ITS) region of the nuclear rRNA gene cluster as a suitable marker for phylogenetic research across the fungal kingdom, and the ITS has since been ratified as the universal barcode for fungi[9,10]. However, a few caveats limit the utility of the ITS for species identification: (1) sequences are only available for about half of the 140,000 or so known fungal species[5], (2) the presence of intragenomic variation may affect the accuracy of species identification[11], and (3) there is a shortage of reliable high-quality reference sequences from type specimens[12]. The vast collections of historical type and other voucher fungal specimens preserved in fungaria around the world therefore represent an invaluable source of DNA for barcoding and phylogenetic studies. DNA barcoding becomes an especially important tool in identification of dried voucher specimens for which few or no morphological records exist. Although isolation and amplification of DNA from fungarium specimens is substantially more challenging than working with fresh source material, it is fully possible to successfully amplify barcode sequences from a specimen more than 100 years old[13−17].
The Guyana Shield is a highly biodiverse region of northeastern South America. It is characterized by an ancient geological formation that features several unique table-top mountain plateaus (tepuis) rising above the surrounding tropical savannas, lowland and montane forests, and montane scrub, resulting in high endemism of plants and animals. In 1983, the Smithsonian Institution's National Museum of Natural History (NMNH) initiated the Biological Diversity of the Guyana Shield (BDG) program to study, document, and preserve the biological diversity of this unique biogeographical region of South America. With most of its diverse habitats still intact, the relatively unexplored country of Guyana has attracted special attention. Between 1983 and 2005, the BDG led several expeditions across Guyana to map its flora and mycota[18−23]. The many thousands of fungal specimens collected during these expeditions were distributed to herbaria across the world, where they serve as an invaluable source of reference material for future mycology studies[13]. Thus, the exhaustive collection efforts by the BDG program and subsequent follow-up studies in this region of South America have greatly enhanced our knowledge of tropical fungal biodiversity, resulting in the discovery and description of numerous new species[24−39].
A subset of the fungal collections from Guyana (~550 samples of dried fruiting bodies) was distributed to the United States National Fungus Collections (BPI), where they have remained in storage until the present. Roughly half of these collections were indeterminates (indets); i.e., specimens that could not be identified to the species or genus level at the time of collection. Unfortunately, few indets were accompanied by morphological records to aid in later identification efforts. The growing record of fungal discoveries coming out of Guyana since the initiation of the BDG program (see above) demonstrates that much remains to be learned about the mycota of the Guyana Shield. The large number of fungal specimens from Guyana kept in storage at the National Fungal Collections awaiting accessioning into the fungarium therefore holds the potential to harbor several rare and undescribed species from this biodiverse region of South America. The objectives of this study were (1) to examine the collection of dried fruiting bodies with the specific aim of providing determinations of the many indets by means of DNA barcoding and (2) obtaining accession samples for which molecular determinations could be made in the United States National Fungus Collections (BPI).
-
This study included dried fruiting bodies (sporocarps) from 551 unique samples collected at multiple locations and habitats across Guyana and the border region with Brazil over a long time period (1982–2005). The fungal collection effort varied greatly from expedition to expedition, and the number of specimens collected peaked during the time periods 1990–1994 and 1998–2002 (Fig. 1). The details of each expedition, including route maps and descriptions of collecting sites and habitats are freely available in a series of Smithsonian Contributions to Botany publications[18–23,40,41].
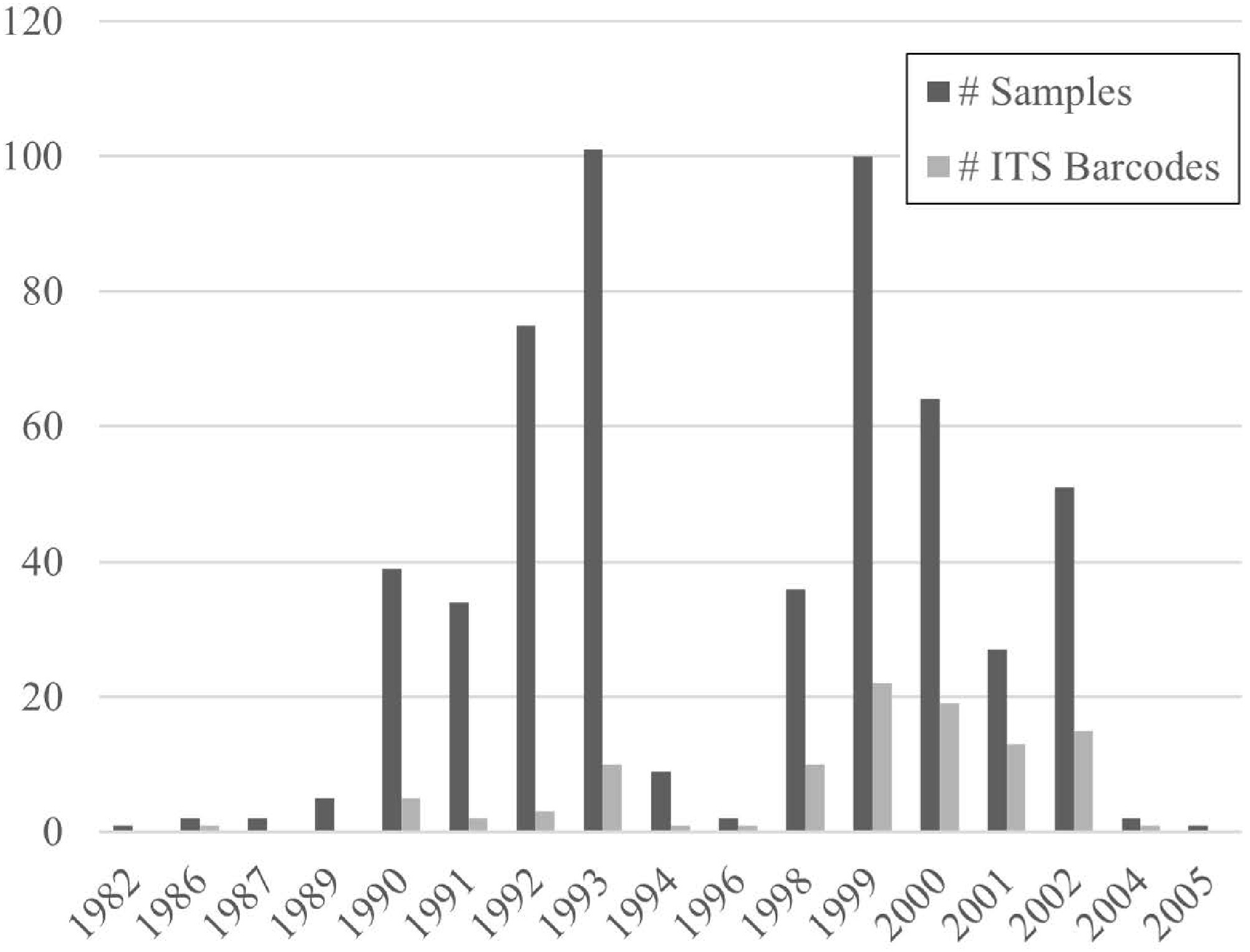
Figure 1.
Temporal distribution of specimens' sampling and sequencing success.
DNA extraction, polymerase chain reaction, and sequencing
-
About 10–100 mg of dried sporocarp tissue was taken from each sample and placed in 2-mL tubes containing 500-µm garnet and 6-mm zirconium Satellite beads (Ops Diagnostics, Lebanon, New Jersey) and subsequently ground to a fine powder using a three-dimensional (3D) Precellys Evolution homogenizer (Bertin, Montigny-le-Bretonneux, France). Genomic DNA was subsequently extracted using a Qiagen DNeasy Plant Mini Extraction Kit (Qiagen, Valencia, California), following the manufacturer's protocols.
The highly variable ITS regions between the 18S, 5.8S, and 28S nuclear ribosomal genes were amplified using a suite of primer combinations, targeting basidiomycetes and ascomycetes (Table 1). Amplification reactions included 12.5 µL of KAPA2G Robust HotStart ready-mix (KAPA Biosystems, Wilmington, Massachusetts), 1.25 µL of each primer (10 µM/L), 1−2 µL of template DNA, and distilled H2O for a total reaction volume of 25 µL. Polymerase chain reactions (PCRs) were performed on a Bio-Rad C1000 Touch Thermal Cycler (Life Science, Hercules, California) under the following conditions: 3 min of initial denaturing at 95 °C, followed by 35 cycles of 30 s of denaturing at 95 °C, 15 s of annealing at 50 °C and 1 min of extension at 72 °C, and a final extension at 72 °C for 5 min. Amplification success was assessed by running 12 µL of the PCR products on an automated capillary QIAxcel Advanced electrophoresis system (Qiagen, Valencia, California). Amplicons were cleaned enzymatically using the following protocol: a mixture of 1 µL of Exonuclease I, 1 µL of shrimp alkaline phosphatase, and 3 µL of the PCR product was placed in a thermocycler and digested at 37 °C for 15 min and 80 °C for 15 min. Cleaned amplicons were sequenced using BigDye v.3.1 chemistry (Applied BioSystems, Foster City, California) with the same primers used for amplification. Sequencing products were purified using ethanol–ethylenediaminetetraacetic acid (EDTA) precipitation and run on a SeqStudio Genetic Analyzer (Applied Biosystems, Foster City, California). Sequence contigs were assembled and edited in Sequencher v. 5.5 (GeneCodes Corp., Ann Arbor, Michigan). Finally, BLASTn searches were conducted in GenBank (
https://blast.ncbi.nlm.nih.gov/ ) to verify or determine species' identification using the traditional 97% ITS sequence identity as a DNA barcoding threshold[42].Table 1. Fungal ITS (internal transcribed spacer) primers used in this study.
Primer name Oligo sequence (5'→3') Ref. ITS1 TCCGTAGGTGAACCTGCGG [8] ITS1F CTTGGTCATTTAGAGGAAGTAA [67] ITS4 TCCTCCGCTTATTGATATGC [8] ITS4b CAGGAGACTTGTACACGGTCCAG [67] ITS5 GGAAGTAAAAGTCGTAACAAGG [8] NLB3 GGATTCTCACCCTCTATGA [68] NLB4 GGATTCTCACCCTCTATGAC [68] NSI1 GATTGAATGGCTTAGTGAGG [68] LB-W CTTTTCATCTTTCCCTCACGG [69] LA-W CTTTTCATCTTTCGATCACTC [69] -
One hundred and three (103) high-quality ITS sequences were retrieved from the 551 specimens included in this study. Comparison with other ITS sequences in GenBank identified 76 fungal taxa distributed into 30 families (Table 2). From this, we uncovered 3 genera and 11 species that had never before been recorded in Guyana, according to data in MyCoPortal (
www.mycoportal.org ), GenBank (www.ncbi.nlm.nih.gov ), Global Biodiversity Information Facility (GBIF,www.gbif.org ) and iNaturalist (www.inaturalist.org ). DNA barcoding provided identifications to the species level for 33 unknown samples (i.e., indets), and 5 specimens to the genus level, and added identifications to the species level for specimens for which only generic determinations and few morphological records existed previously. Another 34 ITS barcode sequences confirmed prior morphological determinations made by the original collectors. Finally, in 10 instances, the ITS barcode sequence conflicted with the previous determination made on the basis of the morphology and/or chemical characteristics. The ITS barcode sequences provided closely matched congeners or closely related genera, warranting close re-examination of the specimens involved. None of the identifications provided by ITS barcoding represent potentially undescribed species. The designation to genus level rather reflected a lack of sequence differentiation among congeners than a lack of close matches in GenBank BLAST searches. The ITS sequences generated by this study have been deposited in GenBank with the accession numbers shown in Table 2.Table 2. Fungal taxa identified by BLASTN queries of ITS sequences in GenBank.
Family Taxon GenBank Agaricaceae Leucoagaricus sp. PP102299 Amanitaceae Amanita cyanopus C.M. Simmons, T.W. Henkel & Bas PP102312 Amanita fulvoalba Mighell & T.W. Henkel PP102313 Auriculariaceae Auricularia delicata
(Mont. ex Fr.) Henn.PP102310 Auricularia fuscosuccinea1 (Mont.) Henn. PP102308−102309 Boletaceae Brasilioporus rufonigricans (T.W. Henkel) A.C. Magnago & T.W. Henkel PP102325 Austroboletus rostrupii
(Syd. & P. Syd.) E. HorakPP830805 Chalciporus sp. PP830806 Chalciporus trinitensis, aff. (Heinem.) Singer, I.J.A. Aguiar & M.H. Ivory PP102328 Singerocomus inundabilis (Singer) T.W. Henkel & Husbands PP102315 Tylopilus sp. PP102324 Tylopilus balloui (Peck) Singer PP102323, PP830803 Tylopilus exiguus T.W. Henkel PP830800−830801 Tylopilus orsonianus Fulgenzi & T.W. Henkel PP102319−102321, PP830802 Tylopilus potamogeton var. irengens T.W. Henkel PP102322 Tylopilus vinaceipallidus (Corner) T.W. Henkel PP102316−102318 Xerocomus potaroensis T.W. Henkel & Husbands PP102326−102327, PP830804 Callistosporiaceae Guyanagarika pakaraimensis Sánchez-García,
T.W. Henkel & AimePP102307 Cantharellaceae Cantharellus guyanensis
Mont.PP102304 Clavariaceae Clavaria zollingeri Lév. PP102362 Clavulinaceae Clavulina sp. PP830810 Clavulina caespitosa T.W. Henkel, Meszaros & Aime PP102358 Clavulina connata1, cf.
(Berk.) CornerPP102355 Clavulina craterelloides Thacker & T.W. Henkel PP102356−102357 Clavulina dicymbetorum T.W. Henkel, Meszaros & Aime PP102360 Clavulina kunmudlutsa T.W. Henkel & Aime PP102361 Clavulina tepurumenga T.W. Henkel & Aime PP102359 Cordycipitaceae Cordyceps sp. PP102372 Cordyceps polyarthra1 Möller PP102371 Cordyceps takaomontana Yakush. & Kumaz. PP102370 Coriolaceae Agaricus sp. PP102298 Dacrymycetaceae Dacryopinax spathularia (Schwein.) G.W. Martin PP102354 Elaphomycetaceae Elaphomyces sp. PP102365 Pseudotulostoma volvatum O.K. Mill. & T.W. Henkel PP102366−102369 Entolomataceae Entoloma sp. PP830796 Nolanea sp. PP830797 Rhodocybe sp. PP102314 Incertae sedis Ductifera pululahuana (Pat.) Donk PP102311 Geastraceae Geastrum sp. PP102376 Gloeophyllaceae Gloeophyllum sp. PP102330 Gloeophyllum striatum
(Fr.) MurrillPP102329 Hygrophoraceae Hygrocybe sp. PP102295−102296 Hygrocybe conica var. brevipora (Dennis) S.A. Cantrell & Lodge PP830807 Hymenochaetaceae Fuscoporia gilva (Schwein.)
T. Wagner & M. FischPP102353 Hysterangiaceae Hysterangium sp. PP102293−102294 Inocybaceae Inocybe lilacinosquamosa Matheny, Aime & T.W. Henkel PP102297 Meruliaceae Cymatoderma dendriticum (Pers.) D.A. Reid PP102351 Irpex lacteus (Fr.) Fr. PP102350 Phlebia1 radiata1Fr. PP102349 Mycenaceae Panellus stipticus1 (Bull.)
P. Karst.PP102300−102301 Physalacriaceae Cyptotrama sp. PP102302 Polyporaceae Favolus brasiliensis (Fr.) Fr. PP102322 Fomes fasciatus (Sw.) Cooke PP102333−102334 Hexagonia variegata Berk. PP102331 Hexagonia hydnoides (Sw.)
M. FidalgoPP830809 Lentinus crinitus (L.) Fr. PP102337 Lentinus scleropus (Pers.) Fr. PP102338 Lentinus swartzii1 Berk. PP102335−102336, PP830808 Panus strigellus (Berk.) Chardón & Toro PP102339−102340 Trametes sp. PP102347−102348 Trametes sanguinea
(Klotzsch) Pat.PP102341−102345 Trametes trogii Berk. PP102346 Stereaceae Stereum sanguinolentum1
(Alb. & Schwein.) Fr.PP102352 Russulaceae Russula sp. PP830798 Russula gelatinivelata S.L. Mill., Aime & T.W. Henkel PP102305, PP830799 Russula puiggarii
(Speg.) SingerPP102306 Sarcoscyphaceae Cookeina speciosa (Fr.) Dennis PP102364 Cookeina tricholoma
(Mont.) KuntzePP102363 Sclerodermataceae Scleroderma sp. PP102303 Squamanitaceae Squamanita guyanensis
T.W. Henkel & I. SaarPP102378 Xylariaceae Daldinia eschscholtzii1 (Ehrenb.) Rehm PP102373 Xylaria sp. PP102375, PP830811 Xylaria ophiopoda1 Sacc. PP102374 Nectriaceae Corallonectria1 jatrophae1 (Möller) C.S. Herrera &
P. ChaverriPP102377 Neophysopellaceae Neophysopella1 vitis1 (P. Syd.) Jing X. Ji & Kakish. PP830812 1Taxa new to Guyana. Successful amplification of the ITS region from these 20- to 30-year-old field-collected fruiting bodies proved very challenging. Although the overall success was low, the amplification and sequencing success improved significantly over time when comparing the proportion of ITS barcode sequences obtained relative to the number of samples examined from the early years of the BDG program (1982–1994) with the later years (1996–2005) (Fig. 1, p = 0.03, Mann–Whitney U-test). False positives were rampant due to microbial contamination (e.g., Aspergillus, Malassezia, Metarhizium, Penicillium, and Physisporinus). Typically, repeated rounds of amplifications under varying PCR conditions (i.e., annealing temperature, amount of DNA template, different primer combinations) were necessary before clean amplicons of appropriate length (600−1,100 bps) were acquired. Curiously, tissue samples from certain fungal groups (especially boletes and carbon fungi such as Xylaria) repeatedly produced very dark DNA extracts. These samples consistently failed to amplify the ITS region. Diluting the extracts in double-distilled water did not improve the amplification success of these samples.
-
This study underscores the value of well-curated voucher specimens as essential resources for building reliable DNA reference libraries and linking names to sequences, which, in turn, form the foundation for biodiversity studies, conservation planning, and environmental monitoring. The challenges of extracting high-quality DNA from historical fungaria specimens also reiterates the need for continued collection of fresh specimens from poorly studied regions of the world, as well as targeted collection of fungal groups that are poorly represented in herbaria[43−46].
The specimens examined here were collected over an extended time period and under various storage conditions, two factors which clearly affected the quality of the source material, the degree of DNA degradation, and ultimately the ability to successfully amplify and sequence the > 600-bp ITS region from these old, dried fungal specimens. As expected, the proportion of samples that successfully yielded a high-quality barcode sequence declined significantly over time. Although vouchered type specimens and historical fungarium collections represent invaluable sources of reference material for linking DNA sequences to species names[47,48], efforts to retrieve ITS barcode sequences from historical vouchers can be a challenging and costly exercise[6,15]. As demonstrated here, successful amplification of a 600-bp ITS barcode sequence from older fungarium specimens requires more rigorous DNA extraction procedures and PCR troubleshooting[13,49]. For poorly preserved specimens, a time-consuming and laborious alternative to direct amplification is to amplify the two subunits of the ITS region (ITS-1 and ITS-2) separately and sequence shorter overlapping fragments that are later assembled in silico[50]. In addition to degrading the DNA's quality, the age of fungarium specimens and the method used to preserve them also significantly increases the risk of contamination[13,51]. The specimens examined in this study were all collected, sorted, and dry-preserved in the field during extended expeditions into remote tropical areas of South America, and subsequently stored in boxes on office shelves for nearly two decades until the present day. These are all factors which likely contributed to the pervasive levels of microbial (primarily by molds and yeasts) and biochemical contamination in our DNA samples, which, in many instances, severely hampered our ability to amplify uncontaminated ITS sequences from dried sporocarp tissues. Finally, it has been shown that residual polysaccharides and secondary metabolites such as phenolic compounds can adversely affect the DNA extraction process[52]. Certain fungal groups contain more of these compounds than others and will often yield DNA extracts that appear cloudy or stained from pigmentation[49]. The dark pigmentation observed in several DNA extracts obtained from bolete specimens belonging to the genera Lactarius, Tylopilus, and Xerocomus is suggestive of the presence of such metabolites and could help explain why we repeatedly failed to amplify the ITS barcode region from these specimens. In spite of all of these complicating factors, the seemingly low PCR and sequencing success rate of 18.7% (103/551 samples) achieved in this study is actually in line with success rates reported elsewhere on similarly aged specimens[15].
As so eloquently stated by Begarow et al.[12], "Any species identification effort employing DNA barcoding will be only as good as the available reference sequences permit". This challenge has prompted the development of independent databases such as UNITE (
http://unite.ut.ee ), with the aim of reliably identifying fungi to the species level using carefully curated high-quality reference sequences[53,54]. The wealth of reference specimens available in herbaria around the world should serve a key role in achieving this goal[17,55]. However, not all fungal lineages are equally represented in fungaria. Most historic collection efforts have focused on sampling fruiting bodies of mushroom-forming macrofungi that are only available temporally and thus are easily overlooked. In order to capture unknown fungal diversity and improve the representation of under-sampled lineages in public DNA repositories, it is therefore necessary to support continued efforts to systematically collect, document, and extract high-quality DNA from fresh specimens in the field, especially from geographical regions where comparatively fewer collections exist, such as South America[56].The ITS region displays great variability in DNA sequence divergence across fungal lineages, which complicates its utility as a universal barcode for fungi[42,57,58]. This variability further limits our ability to assign a species identification to environmental samples and unidentified dried specimens with poor morphological descriptions. Since sequence variation in the ITS region is not always sufficient for accurate species identification, it has been argued that a multi-locus phylogenetic approach is needed to accurately resolve species' relationships and assign a taxonomic affiliation to unknown samples[59]. Several additional nuclear loci have demonstrated great promise as complementary secondary barcodes (e.g., nLSU, TEF1, and RPB2) when the ITS alone fails to resolve species-level relationships[5,60]. However, rapid advances in next-generation sequencing technology and drastic cost reductions have made genomic barcoding an increasingly feasible alternative to the traditional reliance on a single universal barcode marker[12,61−66].
-
This study demonstrates both the promise and the challenges of using DNA barcoding to inventory and re-evaluate historical fungal collections from biodiverse yet underexplored regions such as the Guyana Shield. By successfully recovering 103 high-quality ITS sequences from 551 fungarium specimens collected in Guyana several decades ago, we were able to document 76 fungal taxa, including 3 genera and 11 species previously unrecorded in that country. DNA barcoding provided new or confirmed identifications for numerous specimens, highlighting the critical role of molecular tools in resolving the substantial fraction of indeterminate collections that remain in fungaria around the world. Despite the significant obstacles posed by degraded DNA, microbial contamination, and the inhibitory secondary metabolites inherent in older specimens, the sequencing success rate achieved here is consistent with comparable studies of historical collections. However, our findings also illustrate the limitations of relying solely on the ITS as a universal barcode. Inconsistent sequence divergence across fungal lineages, coupled with incomplete coverage in public databases, often constrains the resolution of taxonomic identifications. To address this, a multilocus or genomic barcoding approach should be prioritized in future studies to enhance discriminatory power and improve confidence in species-level assignments. Finally, this work reinforces the need for renewed and systematic collection of fresh fungal material, particularly from biodiversity hotspots such as the Guyana Shield, where fungal diversity remains vastly underestimated. Investments in expanding taxonomic expertise, sequencing efforts, and curation of both new and legacy specimens will be essential to close persistent knowledge gaps and better capture the true scope of fungal diversity on Earth.
This project was supported by funds appropriated to the United States Department of Agriculture (USDA) (Agricultural Research Service [ARS] National Program 301 project #8042-22000-327-000-D). This project was supported in part by an appointment (research fellowship to Frode Jacobsen) to the ARS Research Participation Program administered by the Oak Ridge Institute for Science and Education (ORISE) through an interagency agreement between the US Department of Energy (DOE) and the USDA. ORISE is managed by Oak Ridge Associated Universities (ORAU) under DOE contract No. DE-SC0014664. All opinions expressed in this paper are the author's and do not necessarily reflect the policies and views of USDA, DOE, or ORAU/ORISE. Special thanks go to the Smithsonian Institution's National Museum of Natural History for providing the specimens included in this study. Catalina Salgado-Salazar and John Hall provided valuable assistance in the laboratory.
-
This research did not involve any studies with human participants, and no ethical approval was required according to the institutional and national regulations.
-
The authors confirm contributions to the paper as follows: study conception and design: Jacobsen F, Castlebury LA; data collection: Jacobsen F, Kelloff C; analysis and interpretation of results: Jacobsen F, Castlebury LA; draft manuscript preparation: Jacobsen F, Kelloff C, Castlebury LA. All authors reviewed the results and approved the final version of the manuscript.
-
The genome sequences of fungi utilized in this research project are available online at the GenBank Nucleotide database of the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov). Fungal specimen records are available online at the MyCoPortal (www.mycoportal.org) and GBIF (www.gbif.org) databases.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Jacobsen F, Kelloff C, Castlebury LA. 2025. New additions to the fungal species of Guyana. Studies in Fungi 10: e019 doi: 10.48130/sif-0025-0019 

New additions to the fungal species of Guyana
- Received: 06 May 2025
- Revised: 16 July 2025
- Accepted: 05 August 2025
- Published online: 23 September 2025
Abstract: DNA barcoding is today a standard method used for species identification, preferably in conjunction with supporting morphological data. Here, we employed DNA barcoding using the highly conserved nuclear ribosomal internal transcribed spacer (ITS) region to identify a large set of dried fungal sporocarp specimens that were collected in Guyana between 1982 and 2005. Nearly half of the 551 specimen collections sampled lacked determination below family level (indets). Our study yielded 103 ITS barcode sequences and resulted in the identification of 11 species and 3 genera previously not recorded in Guyana. Thirty-three of the indets were identified to the species level and another five were identified to the generic level. None of the ITS sequences generated in this study represented potentially undescribed species. Our study highlights the importance of fungarium reference specimens as sources of reliable reference barcode sequences for accurate species identification of unknown source DNA. Future barcoding efforts would benefit greatly from utilizing a multilocus approach and next-generation sequencing platforms.
-
Key words:
- DNA barcoding /
- Fungarium specimens /
- ITS /
- Sporocarp /
- Tropical fungi