-
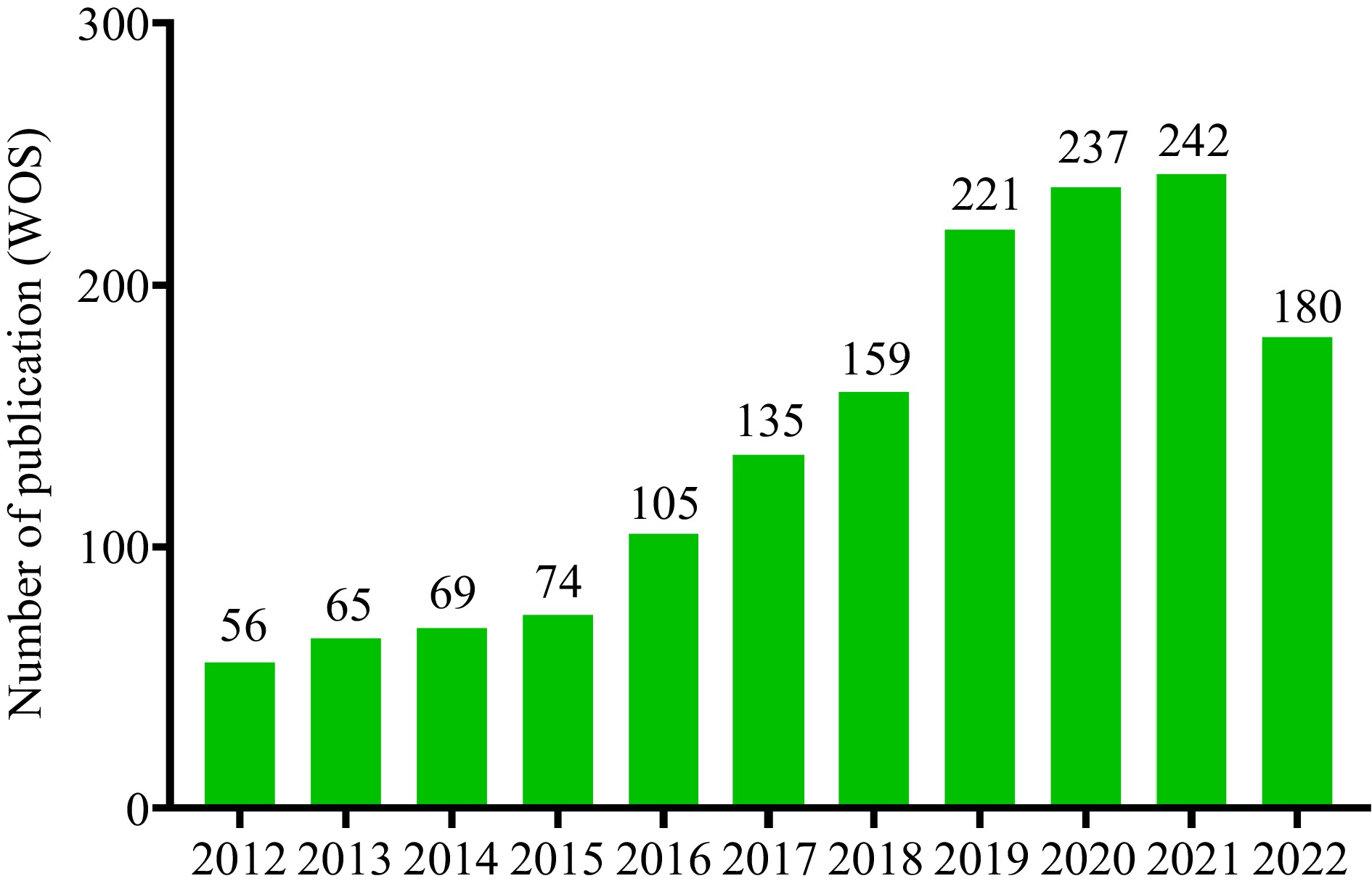
Figure 1.
The number of publications on 'Alzheimer's disease and dietary phytochemicals' (indexed by Web of Science) significantly increased since 2012 and the breakout increase occurred in the last four years (2018−2021).
-
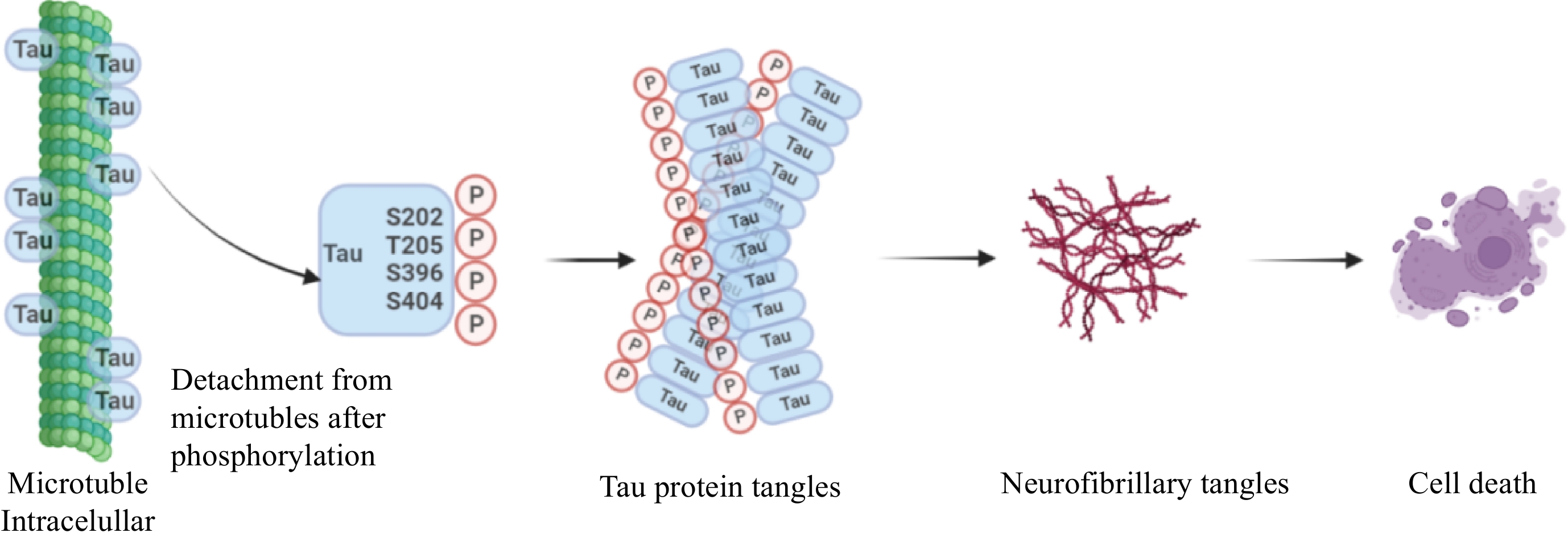
Figure 2.
Hypothesis of tau propagation. The tau proteins are usually hyperphosphorylated by binding to amino residues, typically Ser202, Thr205, Ser396, and Ser404. Then these monomers aggregate to form tau protein tangles, a complex oligomer, that eventually form neurofibrillary tangles, resulting in cell death. Drawing on
https://app.biorender.com/ . -
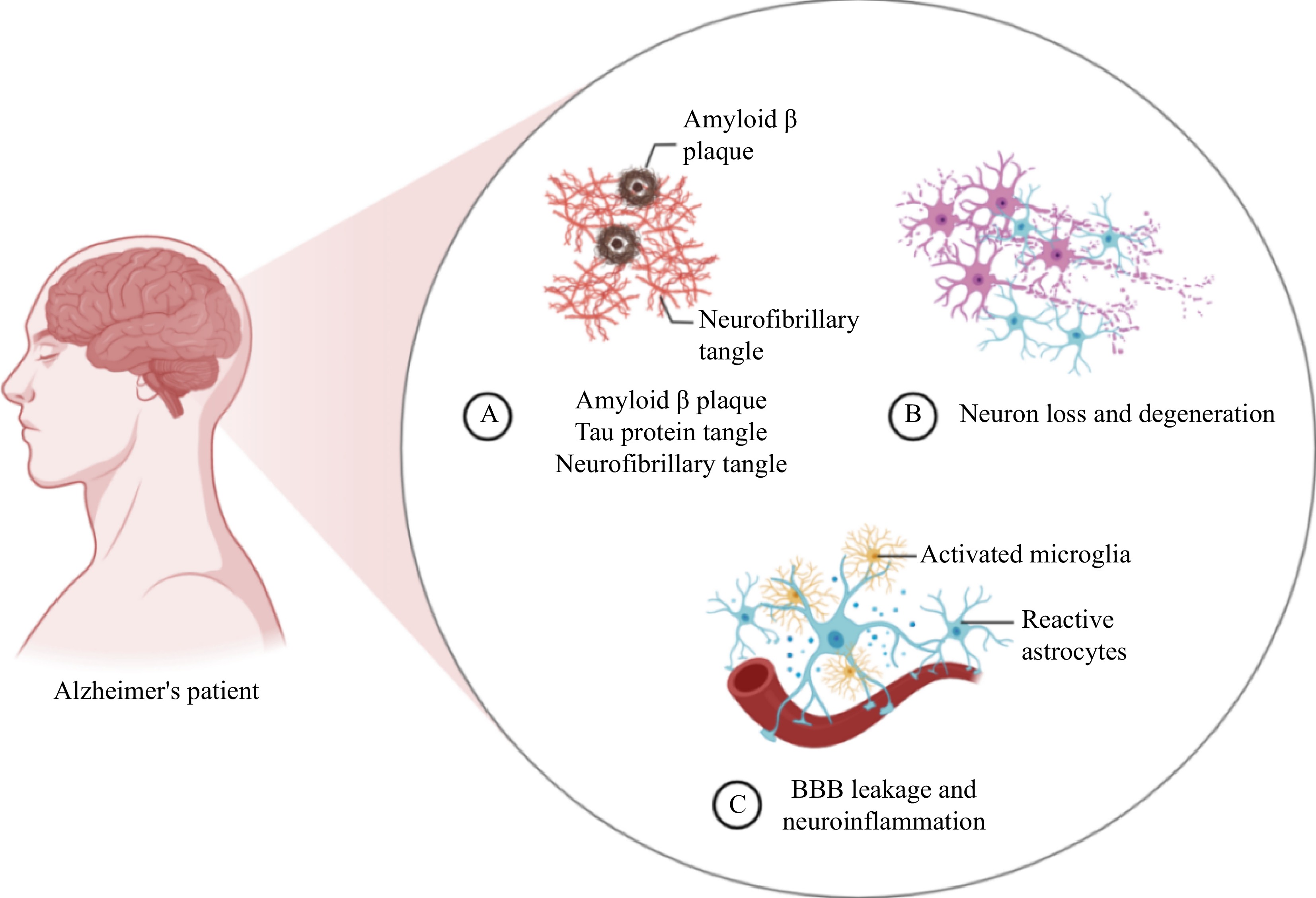
Figure 3.
(A) Three main pathological features of Alzheimer's disease (AD), amyloid beta plaques (Aβ), tau protein tangle and neurofibrillary tangle in the brains of AD patients. (B) The accumulation of neurofibrillary tangle leads to neuronal loss and degeneration, a form of cell death. (C) The continued neuronal loss and degeneration activates microglia and reactive astrocytes, which further contributes to blood brain barrier (BBB) leakage and neuroinflammation. Drawing on
https://app.biorender.com/ . -
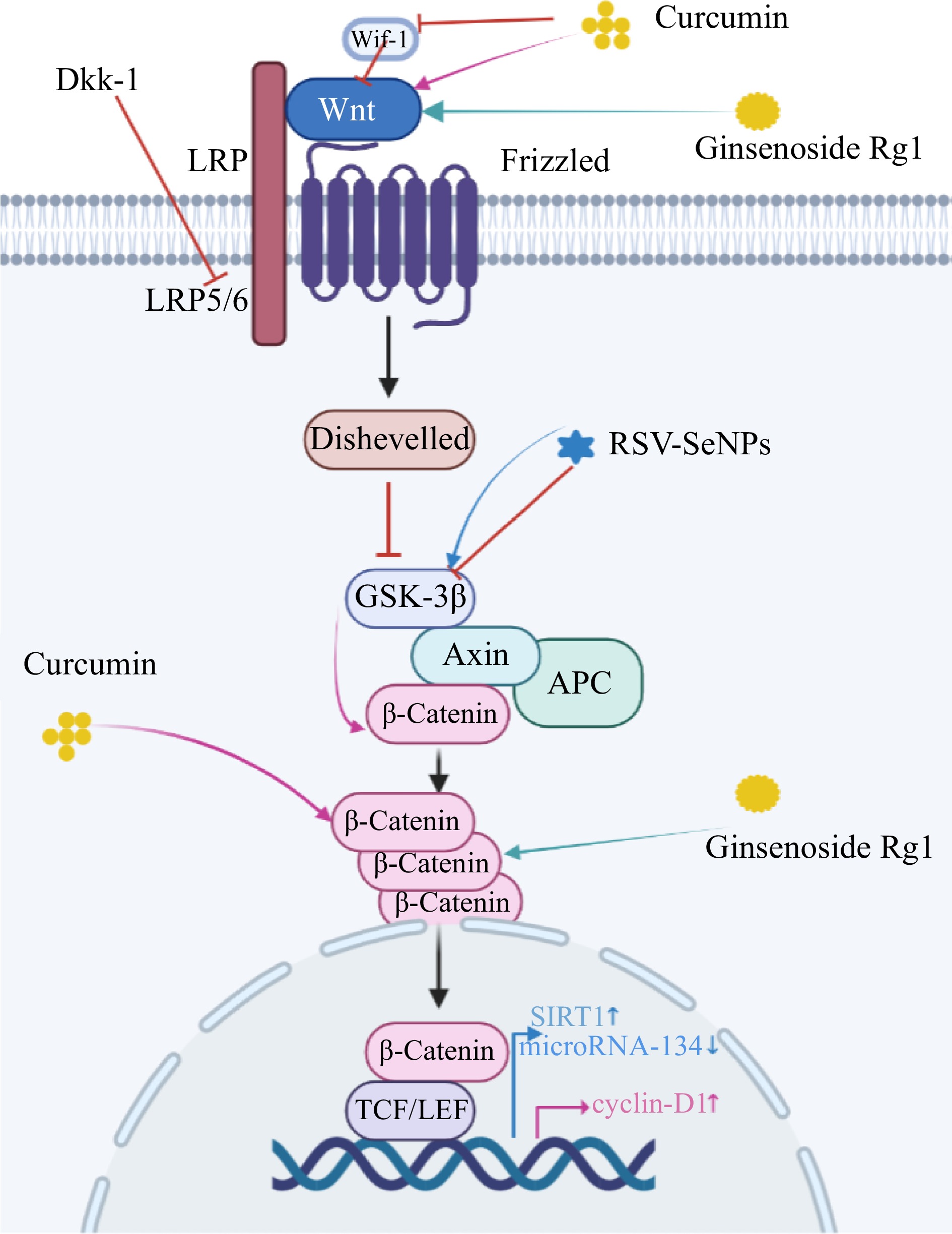
Figure 4.
Schematic model of the role of dietary phytochemicals in neurogenesis through activation of the Wnt/ β-catenin signaling pathway. Pink arrows: Curcumin interacts with Wif-1 and Dkk-1 to increase Wnt levels and activate the Wnt pathway. Wnt interacts with frizzled receptors to down-regulate low-density lipoprotein (LRP-5/6) expression and trigger cytoplasmic disheveled (Dvl). Then it breaks the Axin/APC/GS K-3β homeostasis and down-regulates the expression of GSK-3β. This sequence of reactions ultimately up-regulates cytoplasmic β-catenin expression and transfers it from the cytoplasm to the nucleus. After cellular internalization curcumin directly upregulates cytoplasmic β-catenin levels. In the nucleus, TCF/LEF and cyclin d1 promoter activity were enhanced. Green arrows: Rg1 activates Wnt/GSK-3β/β-Catenin signaling pathway by inhibiting the activation of GSK-3β and phosphorylation of β-Catenin. Wnt signaling pathways are critical in the pathogenesis of the AD. Blue arrows: RSV-SeNPs upregulate the expression of GSK3β, Sirtuin-1 (SIRT1) and decrease that of microRNA-134, consequently increasing neurite outgrowth. Drawing on
https://app.biorender.com/ . -
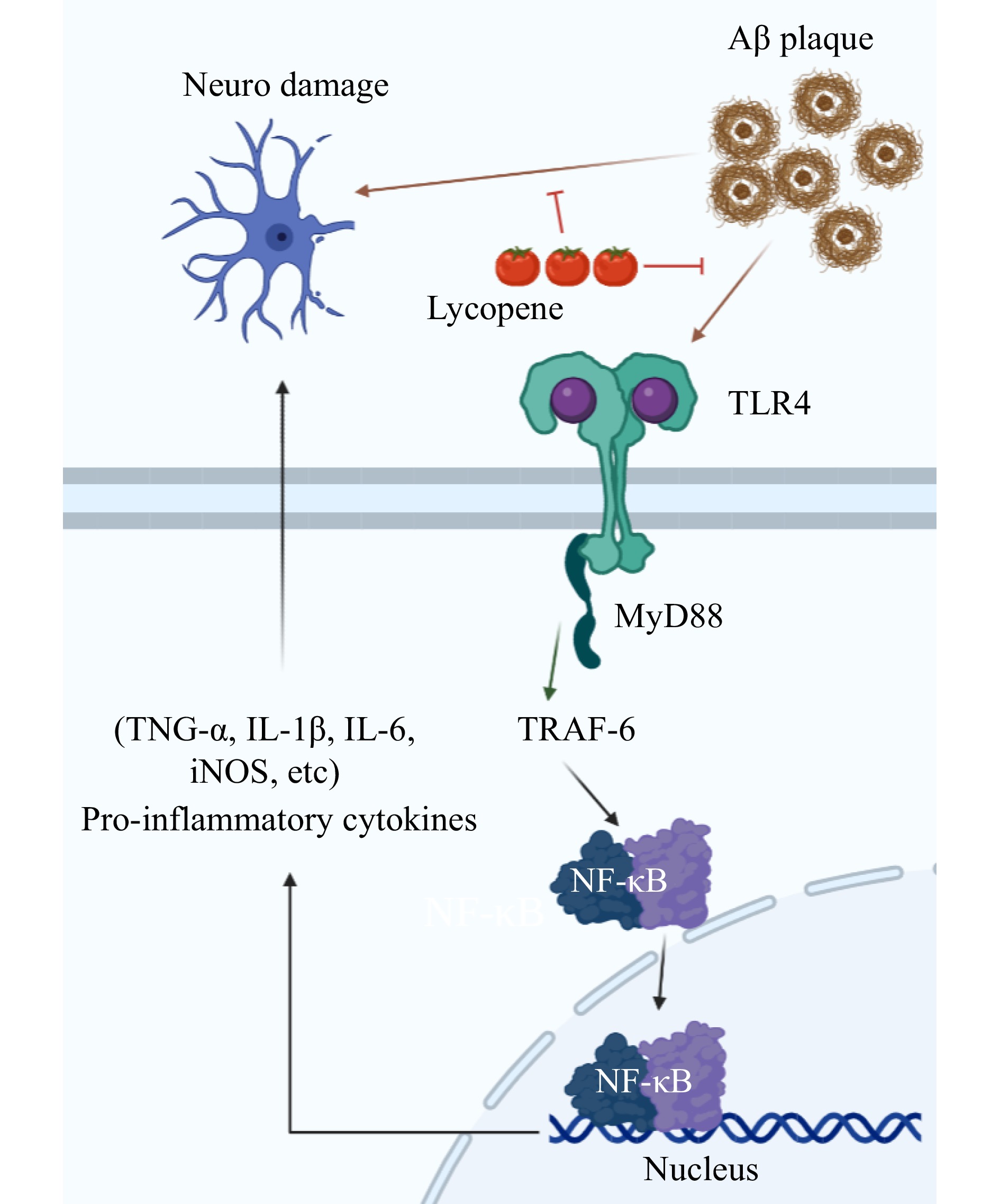
Figure 5.
Lycopene inhibits the down-regulation of TLR4 (Toll-like receptors 4) by Aβ, which further affects MyD88 (Myeloid differentiation primary response gene 88) and TRAF6, thereby activating the NF-κB pathway. On the other hand, lycopene directly inhibits Aβ induced neuronal damage, as shown by decreased levels of serum inflammatory cytokines and increased expression of the p65 subunit and TLR4. Drawing on
https://app.biorender.com/ . -
Dietary phytochemicals Model Dosage Molecular mechanism Ref. 
Wistar rats Cur-PLGA-NPs
(5−20 mg/kg body weight, 3 weeks)Cur-PLGA-NPs causes enhances the nuclear translocation of β-catenin, decreases GSK-3β levels, and increases promoter activity of the TCF/LEF and cyclin-D1. [18] Transgenic APP/PS1
miceCurcumin
(160 ppm, 6 months)Curcumin reduces the level of neuropro-inflammatory miR-146a, up-regulates the expression of CFH protein, and inhibits the phenotype of M1 microglia. [20] 
ICR mice TGN-Res@SeNPs
(50 mg/kg body weight, 16 weeks)kappa B↓ / protein kinase↓ / Akt↓
NF-κB/ mitogen-activated protein kinase/Akt signal pathway.[26] Wistar rats RSV-SeNPs (200 mg/kg body weight,
8 weeks)RSV-SeNPs up-regulates the expression of GSK3β and SIRT1, and down-regulates the expression of microRNA-134, consequently increasing neurite outgrowth. [25] 
Transgenic APP/PS1 mice Lycopene (4 mg/kg body weight, 5 days) LXR↑ / PI3K↑ / AKT↑
Lycopene alleviates neurovascular changes in APP/PS1 mice by activating the LXR–PI3K–AKT signaling pathway.[6] Wistar rats Lycopene
(1−4 mg/kg body weight, 2 weeks)Lycopene decreases NF-κB expression and downregulates IL-1β and TNF-α production. [28] 
Transgenic APP/PS1 mice Gallic acid (20 mg/kg body weight,
6 months)Gallic acid increases the ADAM10 proprotein convertase furin, activates ADAM10 and directly inhibits BACE1 activity, does not alter ADAM10 or BACE1 transcription. [7] 
Transgenic APP/tau/PS1 mice Berberine
(100 mg/kg body weight, 4 months)Berberine ameliorates cognitive deficits, reduces the Aβ accumulation, inhibits the apoptosis of neurons, and promotes the formation of microvessels in the mouse brain by enhancing brain CD31, VEGF, N-cadherin, and Ang-1. [8] 
Sprague dawley rat hippocampal neurons Ginsenoside Rg1
(60 μM, 24 h)CDΚ5↓ / IDE↑ / BACE1↑
Ginsenoside Rg1 significantly decreases CDK5 expression, inhibits PPARγ phosphorylation at serine 273, elevates IDE expression, downregulates BACE1 and APP expression.[44] Tree shrews Ginsenoside Rg1
(30 mg/kg body weight, 8 weeks)Bcl-2/Bax↑ / Wnt↑ / GSK-3β↓ / β-cateni↑
Rg1 increases the ratio of Bcl-2 to Bax and the expression of neuronal markers MAP2 and NeuN Rg1 regulates oxidative stress, cell apoptosis, and neuroinflammation by the Wnt/GSK-3β/β-catenin signaling pathway.[45] 
Wistar rats Pseudoginsenoside-F11 (2−8 mg/kg body weight, 4 weeks) Calpain I↓ / CDK5↓ / GSK-3β↓
Pseudoginsenoside-F11’s decreased GSK-3β (Ser9) phosphorylation and CDK5 activity.[46] 
ICR mice Ginsenoside Rh2
(12.5 and 25 mg/kg, 14 days)ERK↑ / CREB↑ / BDNF↑
Rh2 upregulates the phosphorylation of the ERK-CREB-BDNF pathway in the hippocampus.[47] 
PC12 cells Artemisinin
(0−50 μM, 0−80 min)ERK1/2↑
Artemisinin suppresses LDH release;
Artemisinin restraines the production of intracellular ROS;
Artemisinin modulating Δψm and caspase 3/7 dependent pathway;
Artemisinin activates ERK1/2 signaling.[48] 
ICR mice Torularhodin
(0.5 and 1.5 mg/kg body weight,
4 weeks)Nrf2↑ / NF-κB↓
Torularhodin ameliorates neuronal oxidative damage via the activation of Nrf2 translocation, upregulation of HO-1, and inactivation of NF-κB.[49] Table 1.
Structures of dietary phytochemicals and their potential anti-AD mechanisms.
Figures
(5)
Tables
(1)