-

Figure 1.
Representative varieties of different color series in Bougainvillea. (a) 1a. 'Brazil Lilac', 2a. 'President', 3a. 'Royal Purple', 4a. 'Sao Paulo', 5a. 'Magic Gold Leaf Purple'. (b) 1b. 'Red Ribbon', 2b. 'Flame', 3b. 'Sasha', 4b. 'Xueye Hong', 5b. 'Unicorn'. (c) 1c. 'Orange Ice', 2c. 'Flame Orange', 3c. 'Louise Wathen Mediopicta', 4c. 'Firecracker Orange', 5c. 'Orange Queen'. (d) 1d. 'Bois de Rose', 2d. 'Pink Panther', 3d. 'Kayata', 4d. 'Batik Pink', 5d. 'Liuli Pink'. (e) 1e. 'Yellow Wonder', 2e. 'Fantasy Yellow', 3e. 'California Gold', 4e. 'Golden Glow', 5e. 'Rainbow Foxtail'. (f) 1f. 'Bambino Beesnees', 2f. 'Jamaica White Variegata', 3f. 'Snow White', 4f. 'Jamaica White', 5f. 'White Flame'. (g) 1g. 'Makris', 2g. 'Green Leaf Cherry Blossom', 3g. 'Sophia', 4g. 'Wajid Ali Shah', 5g. 'Pink Bubbles'.
-
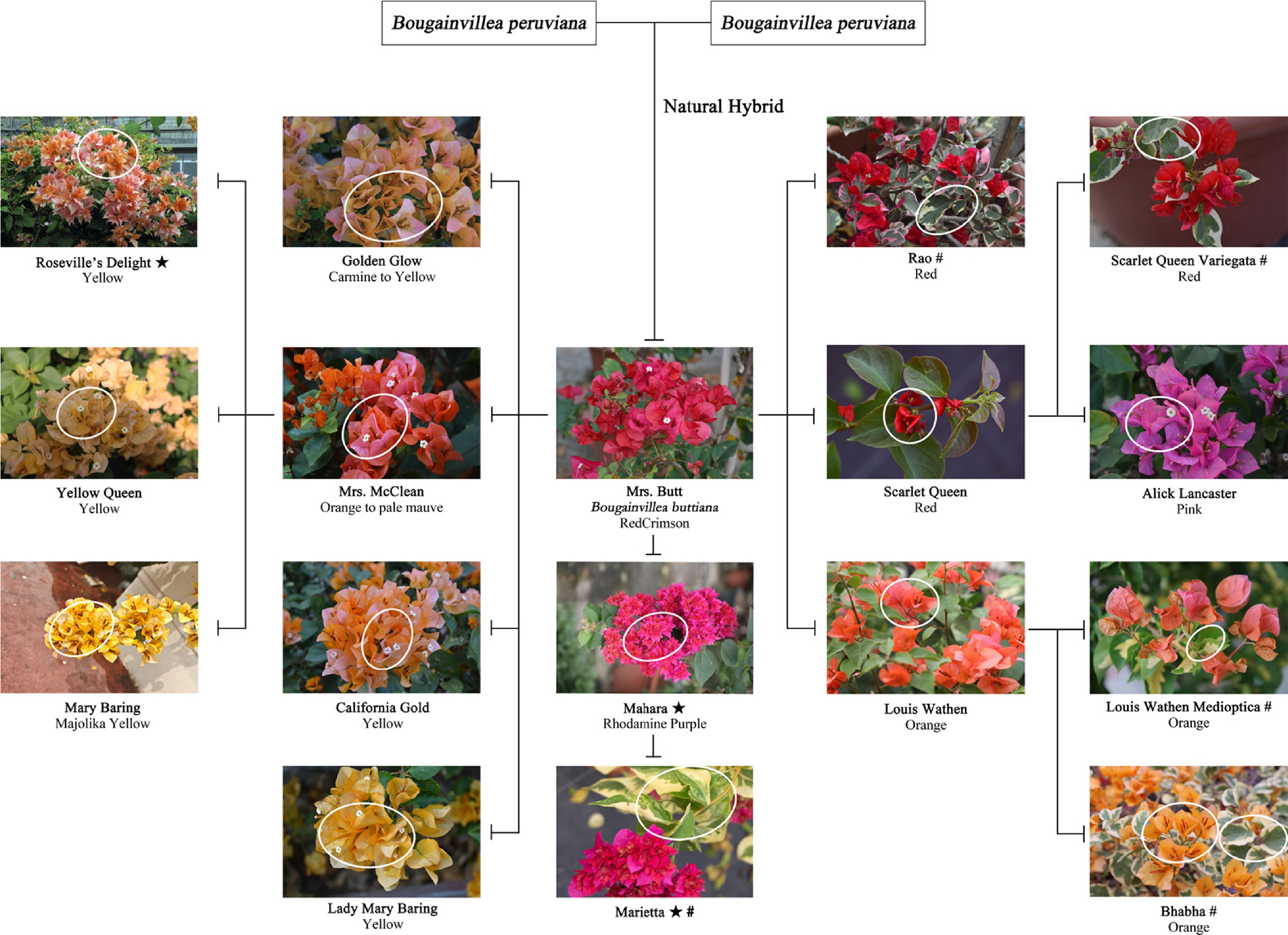
Figure 2.
Bud mutation relationship diagram of Bougainvillea × buttiana 'Mrs. Butt' and its bract colouration[28]. ★ denotes a double-bract variety, # denotes variegated foliage, and a circular mark represents the bud mutation site of the variety.
-

Figure 3.
Schematic diagrams of different bract structures. (a)−(d) Terminal branch structure. (e)−(h) Flower bud structure. (i)−(l) Bract morphology.
-
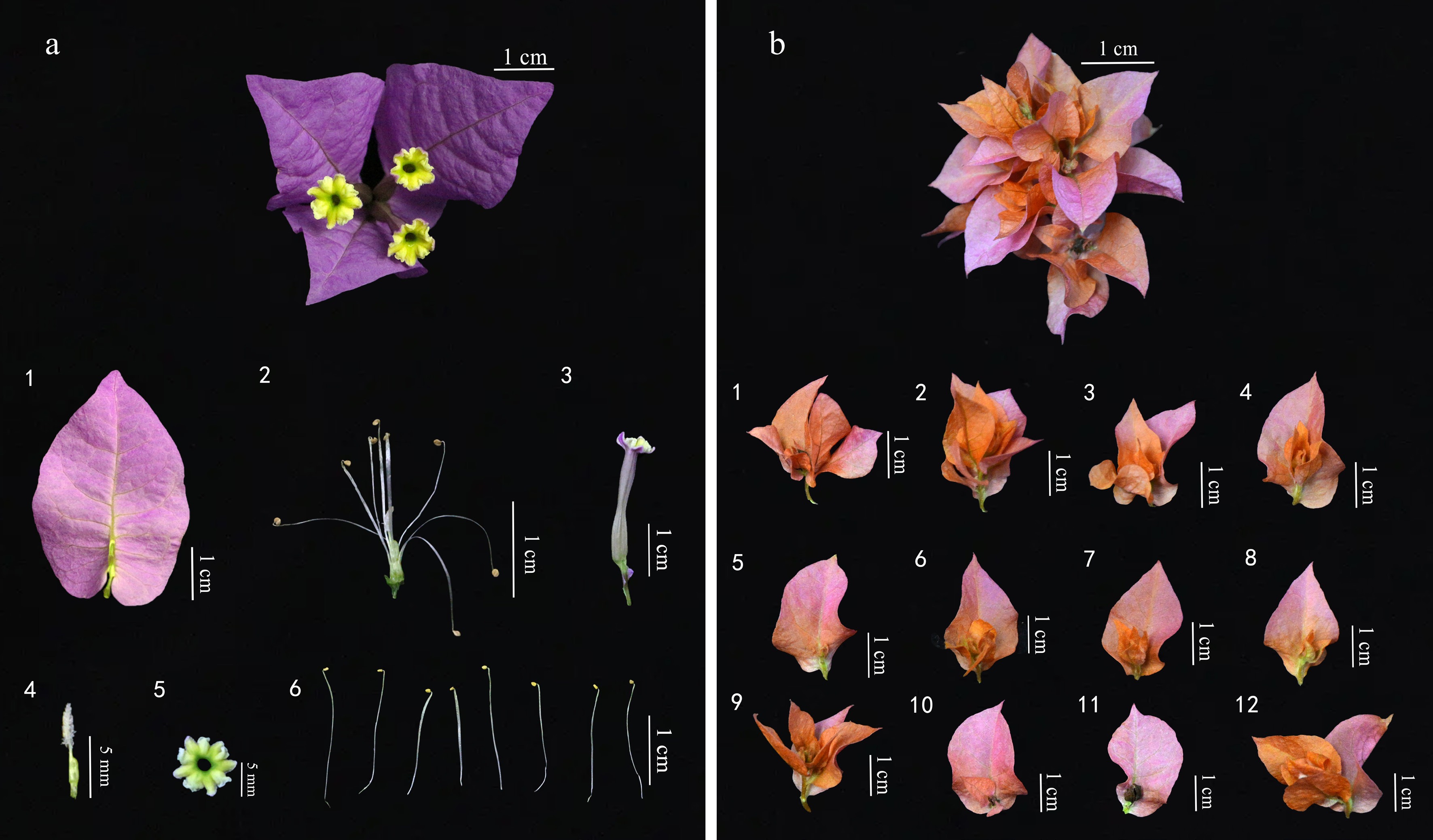
Figure 4.
Comparison of flower bud structures between (a) single-bract Bougainvillea ('President') and (b) double-bract Bougainvillea ('Roseville's Delight'). (a1) Bract. (a2) Stamens and pistil. (a3) Perianth tube. (a4) Pistil. (a5) True flower. (a6) Stamen. (b1)−(b12) Structure of individual bracts.
-
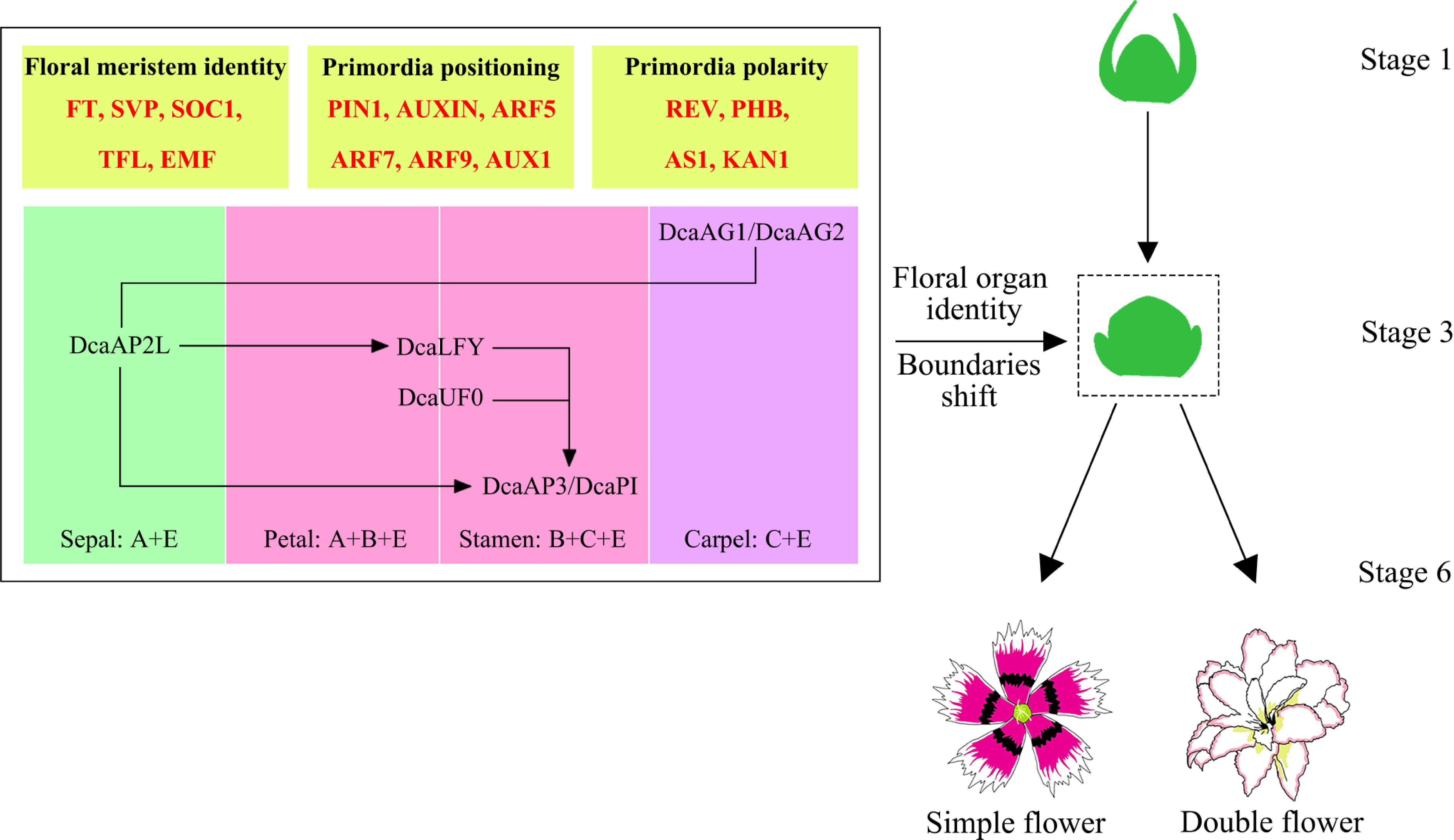
Figure 5.
Transcriptional regulation diagram of single-petalled and double-petalled flowers in Dianthus chinensis[56].
-
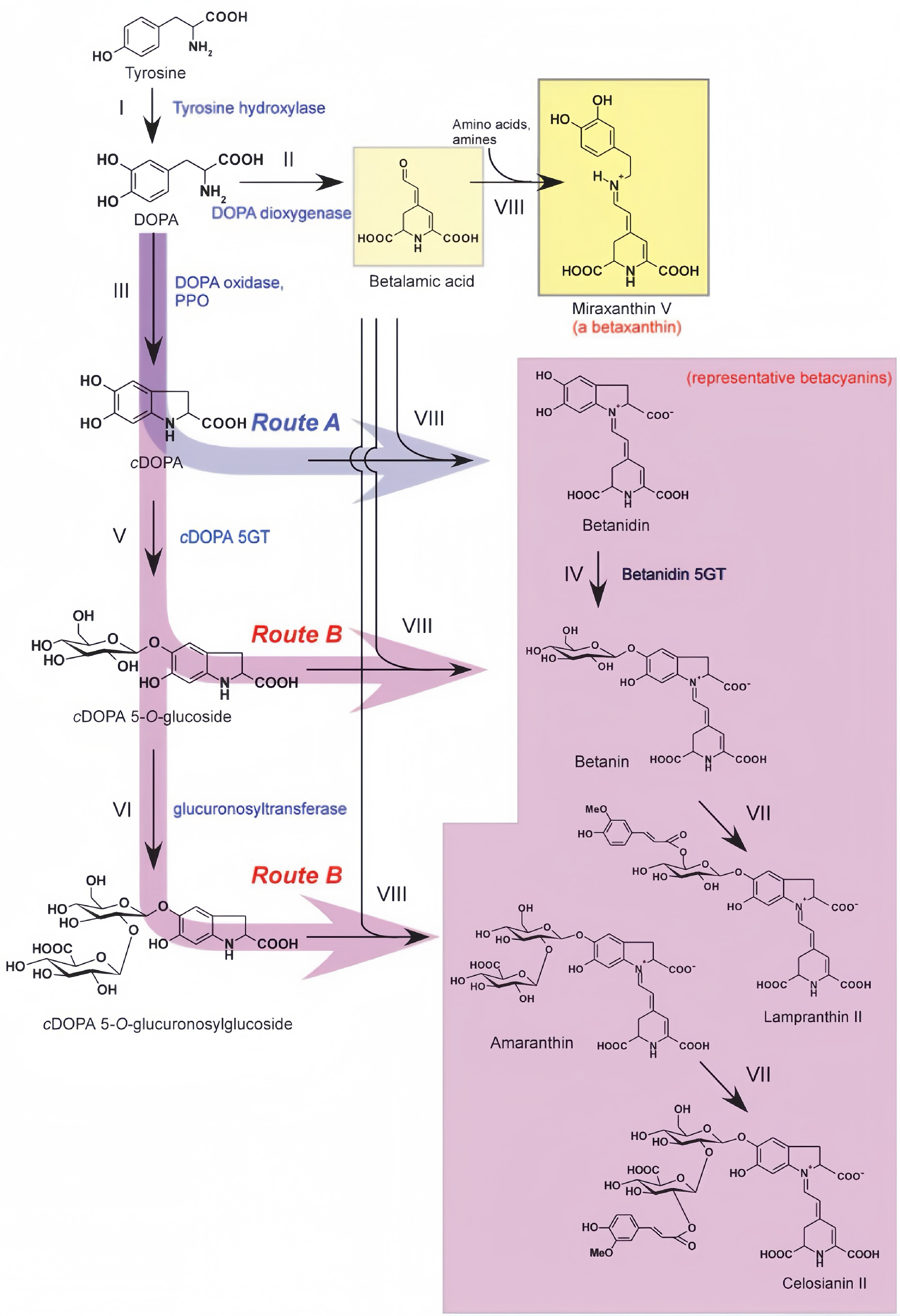
Figure 6.
Biosynthetic pathway of betalain[65].
-
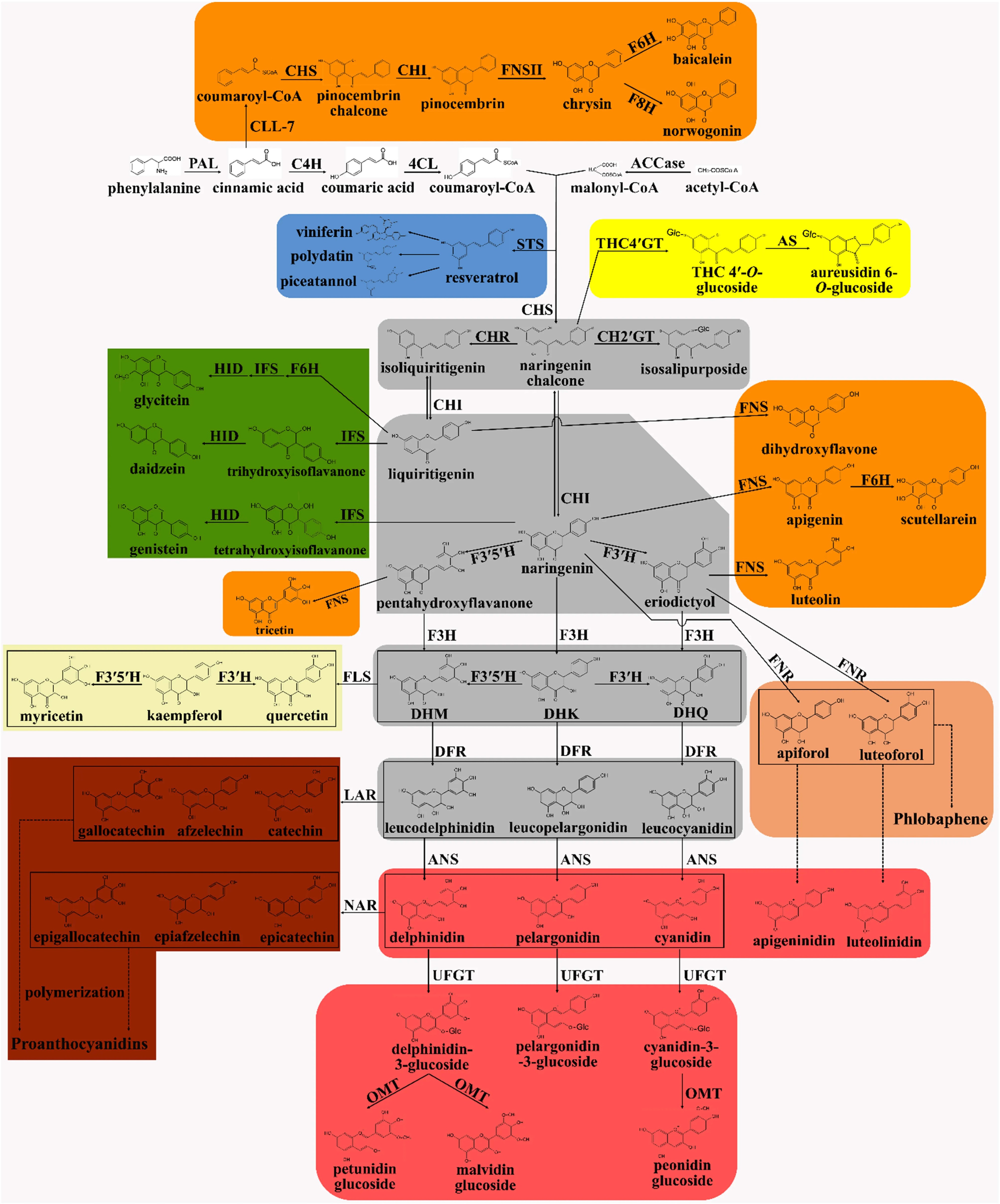
Figure 7.
Biosynthetic pathway of flavonoids[70].
-
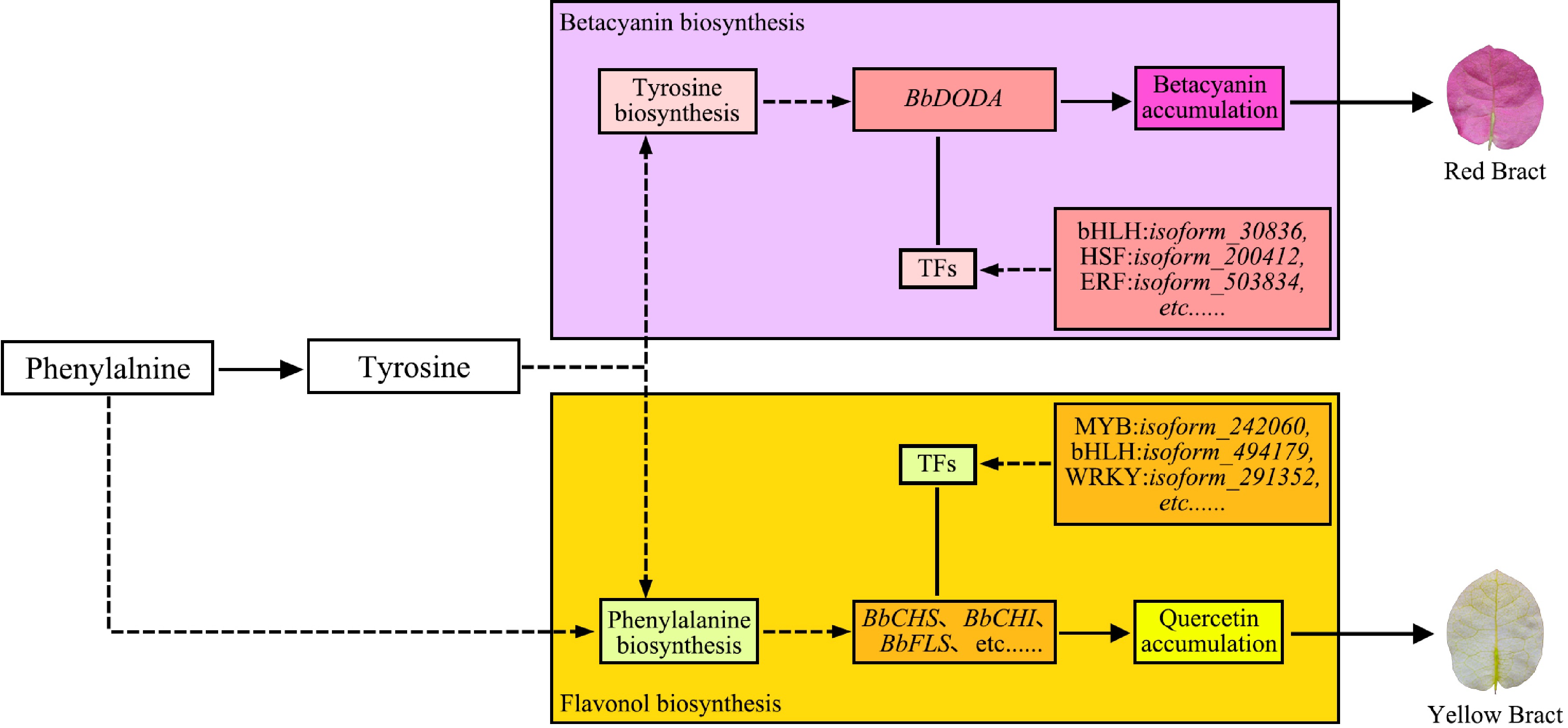
Figure 8.
Molecular mechanism model of bract colour formation in 'Chitra'[74].
-
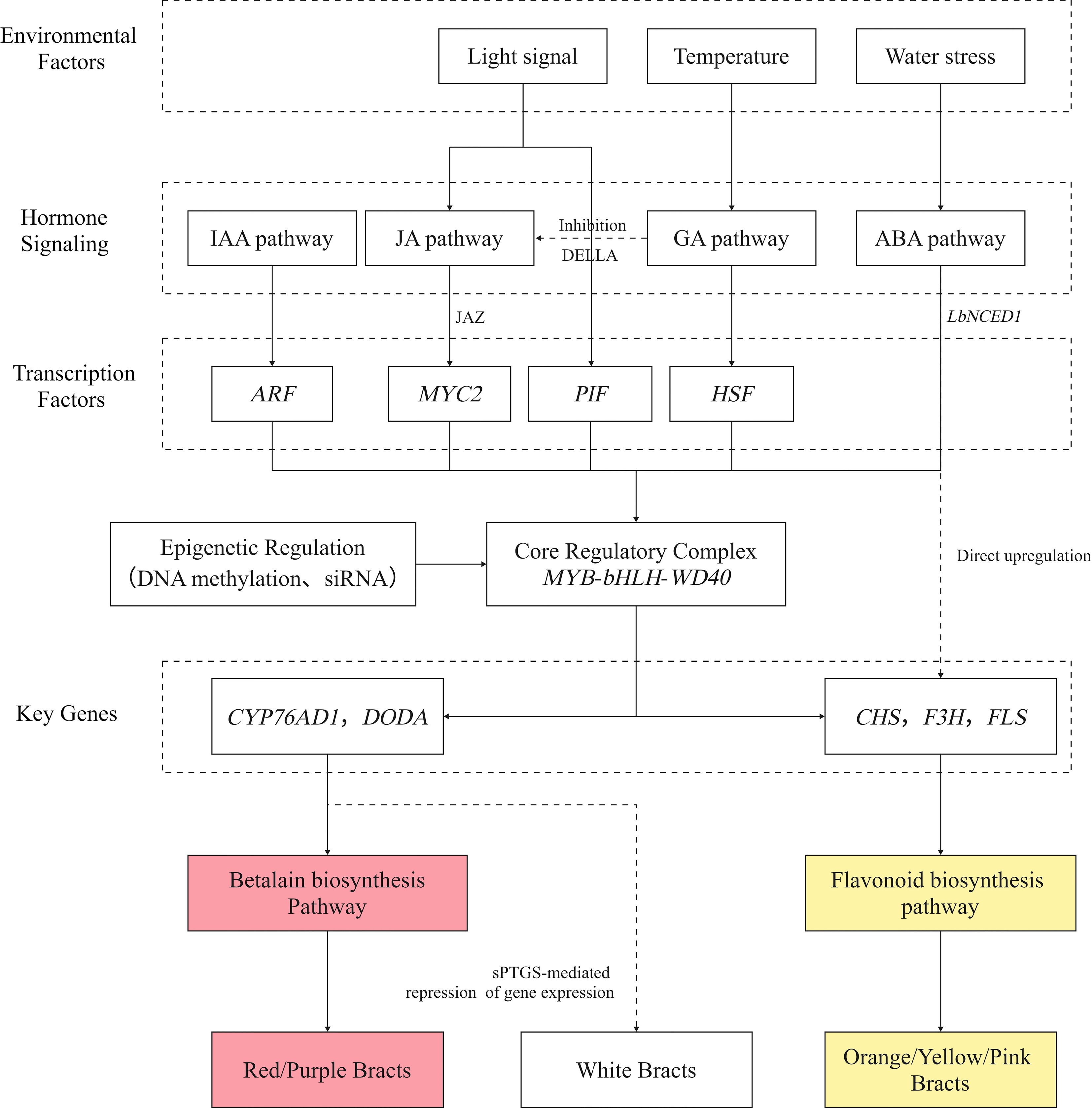
Figure 9.
Regulatory diagram illustrating how genes, hormones, and environmental factors influence the formation of Bougainvillea bract coloration.
-
Gene name Functional description Mode of expression Associated traits YABBY (DL, FIL) Regulate apical-meristematic polarity of floral organs; promote flattened growth. Specifically expressed on the apical surface of the bract primordium. YABBY overexpression results in flattened bracts and morphological diversification; mutant bracts show curling or asymmetry. KNOX (STM, BP) Maintain meristem activity;
inhibit differentiation.Expressed in the apical meristem, Downregulated in bracts. Ectopic expression of KNOX induces bract lobes or nodular protuberances and regulates bract number. ARP (AS1, AS2) Antagonizes KNOX;
promotes organ differentiation.In the bract primordia, full expression occurs. AS1/AS2 mutations result in irregularly lobed bracts, affecting bract symmetry. Auxin signalling pathways (PINs, ARFs) Regulate auxin distribution gradients; determine bract morphology. Expressed at the apex or margin of the bract. The PIN mutation causes bract distortion; ARF overexpression results in enlarged bracts. Gibberellin pathway (GID1, DELLA) Mediates GA signalling;
promotes cell elongation.High expression of bracts during the rapid elongation phase. DELLA mutant bract dwarfism; GID1 overexpression increases bract size. Cell cycle genes (Cyclins, CDKs) Regulating cell division. Expression during the active phase of the bract meristem. Downregulation of expression reduces bract size and affects the number of bract cells. Cell wall-associated genes (EXPANSINS, XTHs) Promote cell wall relaxation;
regulate expansion.Highly expressed during the bract expansion stage. EXPANSINS silences hardening of bracts; XTH overexpression confers flexibility to bracts. Table 1.
Key genes in floral organ development and their characteristics.
-
Gene name Genetic category Functional description Mode of expression Associated traits BgAP1 Class A Regulates the development of sepals and petals, negatively regulating Class C genes through negative feedback inhibition. Highly expressed in bracts and petals, moderately to lowly expressed in stamens and carpels. Maintain the identity of single sepals and petals; expression imbalance may lead to bract formation. BgAP3 Class B Forms heterodimers with BgPI, determining petal and stamen identity. Highly expressed in petals and stamens, moderately to lowly expressed in bracts. In double-flowered varieties, expression of boundary expansion participates in stamen petalisation. BgPI Class B Interacts with BgAP3 to regulate petal and stamen development. Its expression pattern is similar to that of BgAP3, being enriched in petals and stamens. Alterations in Class B gene expression may induce homoeotic transformation in floral organs. BgAG Class C Determines the identity of stamens and carpels, inhibiting the indefinite growth of floral meristems. Highly specifically expressed in stamens and carpels; downregulated in double-flowered cultivars. Functional loss leads to stamen petalisation, forming a double-petalled flower form. BgSEP Class E As a 'glue' protein, it promotes MADS tetramer formation and regulates the development of all floral organs. Widely expressed in all floral organs, with higher expression in bracts. Altered expression of Class E genes affects protein complex stability, indirectly leading to double petals. BgFULL Class C related Participates in carpel and fruit development, with partial functional overlap with AG. Highly expressed during early floral development, associated with bract expansion. May compensate for the loss of apical meristem function and promote the formation of bract-like structures. Table 2.
Functional and expression characteristics of key MADS-box family genes.
-
Gene name Functional description Mode of expression Associated traits PAL Catalyzing the conversion of phenylalanine to cinnamic acid. Highly expressed in magenta bracts and less so in pale yellow bracts. Indirect regulation of red and yellow phenotypes in bracts. CHS Catalyzing the formation of cinnamoyl-CoA from chalcone. Significantly highly expressed in pale yellow bracts, with lower expression in red bracts. Regulating the formation of pale yellow bracts. F3H Catalyzing the conversion of naringenin to dihydroflavonol. Expression is higher in pale yellow bracts and lower in red bracts. Promote the accumulation of yellow pigment. DFR Reduction of dihydroflavonol to form colorless anthocyanins. Highly expressed in orange or purple bracts, with lower expression in red bracts, which are dominated by betacyanins. Promote the formation of orange or purple bracts. ANS Catalyzing the formation of anthocyanins from colorless anthocyanidins. In beetroot pigment plants, it is typically expressed at low levels or not expressed at all. Anthocyanins are absent in Bougainvillea. TYR Catalyzing the hydroxylation of tyrosine to produce L-DOPA. Highly expressed in red bracts and less so in pale yellow bracts. Promoting the formation of red bracts. DOPA Catalyzing the conversion of L-DOPA into betacyanin precursors. Significantly highly expressed in red bracts and lowly expressed in pale yellow bracts. Promoting the formation of red bracts. CYP76AD Participation in L-DOPA hydroxylation. Highly expressed in red bracts and negatively correlated with the expression of flavonoid pathway genes. Influencing the colour ratio of red and yellow bracts. Table 3.
Functional and expressive characteristics of key genes in the beet pigment and flavonoid metabolic pathways.
Figures
(9)
Tables
(3)