








-
Bougainvillea spp., native to South America and also known as the Chinese trumpet vine, leaf flower, or paper flower, constitute evergreen woody vines in the Nyctaginaceae family[1]. They are now widely distributed across tropical and subtropical regions worldwide and play a significant role in landscaping, bonsai cultivation, and garden ornamentation[2,3]. Their true, botanical flowers are small and inconspicuous, clustered at the branch tips, while the brilliantly colored 'petals' are visually prominent and widely appreciated, specialized, modified leaves, bracts. These bracts exhibit considerable chromatic diversity, including red, pink, orange, white, purple, yellow, and complex color patterns. They also display morphological diversity, such as single and double (multi-bracted) forms, which confer substantial ornamental and commercial value to Bougainvillea[4].
For a long time, Bougainvillea breeding has primarily depended on traditional hybridization and sport selection, which are time-consuming and characterized by inherent unpredictability[5]. Given the complex formation mechanisms of its key ornamental traits, including bract development, floral color diversity, and floral characteristics, early studies were primarily devoted to genetic diversity analysis[6,7] and the exploration of physiological and biochemical characteristics[8,9]. Meanwhile, understanding the formation mechanisms of core ornamental traits (e.g., floral color and floral architecture) and stress resistance physiology remained largely descriptive, limited to phenotypic and physiological observations. In recent years, the rapid development of molecular biology techniques, particularly the decoding of genomic information[10,11], has created unprecedented opportunities to systematically elucidate the regulatory networks governing bract development, floral color formation, and floral trait variation in Bougainvillea. With a core focus on bract development, floral color diversity, and floral trait variation, this review aims to provide a comprehensive review of recent advances regarding their evolutionary origins, morphological characteristics, developmental processes, metabolic foundations, and regulatory mechanisms. By integrating relevant physiological and biochemical evidence, a comprehensive framework bridging macroscopic phenotypes and microscopic regulatory mechanisms is established, providing a theoretical foundation for molecular design-based breeding and genetic improvement of Bougainvillea.
-
Bougainvillea species are native to South America, with their current natural distribution concentrated in the tropical and subtropical regions of Brazil, Peru, Argentina, and adjacent areas[1,12]. The earliest wild populations were collected and described in Brazil in 1767, which predominantly exhibited purple flowers[12]. This region is recognized as the primary center of genetic diversity for Bougainvilleas, where the characteristic tropical climate and habitats have provided the environmental basis for the formation and maintenance of the species' original traits[13].
From the 19th to the 21st century, Bougainvilleas were widely introduced to numerous tropical and subtropical regions worldwide, including the United Kingdom, the Netherlands, the United States, Kenya, India, and China, initiating their global cultivation[13]. In the early stages of introduction, key species such as Bougainvillea spectabilis, Bougainvillea glabra, and Bougainvillea peruviana were introduced to Europe and other regions[13,14]. Through introduction and hybridization, significant cultivars such as 'Mrs. Butt' were developed at an early stage. By the 1960s, a wide range of cultivars covering a spectrum of white, red, purple, pink, yellow, and orange hues had been established[14]. In China, Bougainvillea was first introduced to Taiwan Province in 1872 and later became naturalized in Fujian, Guangdong, Yunnan, and Guangxi. Designated as Xiamen's municipal flower in 1986, highlighting its prominent role in subtropical Chinese horticulture[12,13,15].
Ecological adaptation and trait differentiation
-
The widespread cultivation of Bougainvillea worldwide reflects its exceptional adaptability to diverse habitats. This adaptability manifests not only in its tolerance of adverse conditions, such as drought and high temperatures, but also in its geographically differentiated ornamental traits. Owing to its exceptional drought resistance and rapid growth, Bougainvillea is extensively utilized in urban landscaping and horticulture[16,17].
Bougainvillea tolerates high light intensity, salinity, and arid conditions, and thrives in tropical and subtropical regions. It can even grow at elevations up to 2,300 m, although it remains sensitive to frost, entering dormancy during colder seasons. This adaptability further drives trait differentiation and is reflected in the growth habits of cultivated varieties (e.g., erect, semi-erect, trailing, or climbing), leaf characteristics (such as leaf shape ranging from elliptical to ovate and color from pale to deep green), spine traits (density, length, and curvature), and floral traits (such as bract color, shape, and size). These variations represent the genetic diversity of Bougainvillea shaped by environmental adaptation. This differentiation not only aids cultivar identification but also underpins their diverse applications in horticultural landscapes, such as container plants, hedges, or climbers, and underlines the role of ecological pressures in shaping morphological diversity[18,19].
-
Bougainvillea exhibits considerable floral color diversity, with coloration primarily originating from the bracts. These bracts, resembling petals, exhibit vivid hues and are conventionally referred to as petal-like bracts in morphology[20,21]. As shown in Fig. 1, the bract colors of Bougainvillea can be classified into six primary series: purple, red, orange, pink, yellow, white, and an additional multicolored or variegated series[22].

Figure 1.
Representative varieties of different color series in Bougainvillea. (a) 1a. 'Brazil Lilac', 2a. 'President', 3a. 'Royal Purple', 4a. 'Sao Paulo', 5a. 'Magic Gold Leaf Purple'. (b) 1b. 'Red Ribbon', 2b. 'Flame', 3b. 'Sasha', 4b. 'Xueye Hong', 5b. 'Unicorn'. (c) 1c. 'Orange Ice', 2c. 'Flame Orange', 3c. 'Louise Wathen Mediopicta', 4c. 'Firecracker Orange', 5c. 'Orange Queen'. (d) 1d. 'Bois de Rose', 2d. 'Pink Panther', 3d. 'Kayata', 4d. 'Batik Pink', 5d. 'Liuli Pink'. (e) 1e. 'Yellow Wonder', 2e. 'Fantasy Yellow', 3e. 'California Gold', 4e. 'Golden Glow', 5e. 'Rainbow Foxtail'. (f) 1f. 'Bambino Beesnees', 2f. 'Jamaica White Variegata', 3f. 'Snow White', 4f. 'Jamaica White', 5f. 'White Flame'. (g) 1g. 'Makris', 2g. 'Green Leaf Cherry Blossom', 3g. 'Sophia', 4g. 'Wajid Ali Shah', 5g. 'Pink Bubbles'.
Evolutionary origins of bract color diversity
-
As shown in Fig. 2, the evolution of bract coloration in Bougainvillea has been driven by natural hybridization and gene flow among South American wild species. Interspecific hybridization among the wild species Bougainvillea glabra, Bougainvillea spectabilis, and Bougainvillea peruviana has substantially contributed to the expansion of bract color diversity[23].
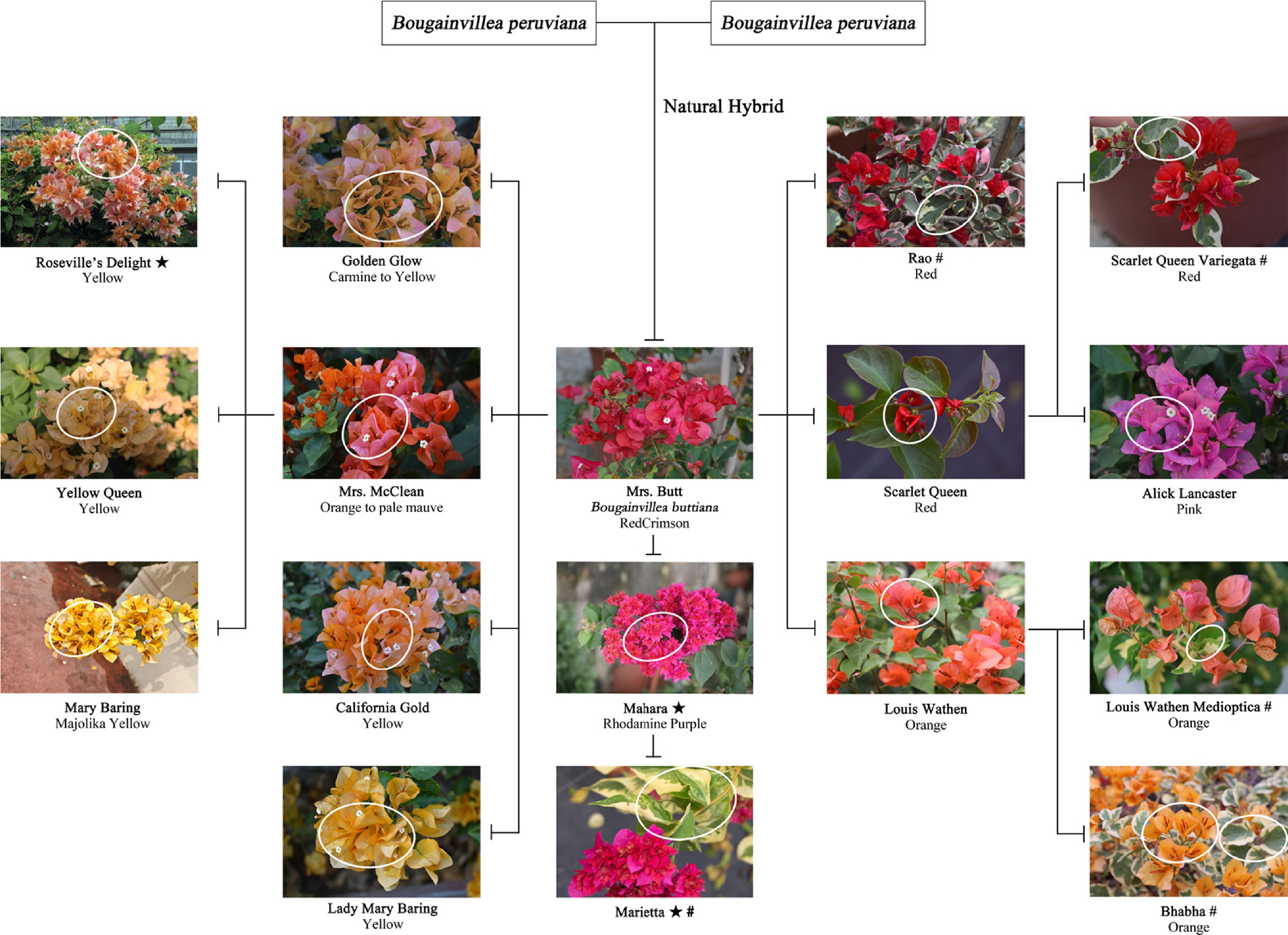
Figure 2.
Bud mutation relationship diagram of Bougainvillea × buttiana 'Mrs. Butt' and its bract colouration[28]. ★ denotes a double-bract variety, # denotes variegated foliage, and a circular mark represents the bud mutation site of the variety.
The purple flower was the earliest floral coloration documented in Bougainvillea. Initially documented by French botanist Philippe Kemmer during expeditions in Brazil, it corresponded to Bougainvillea glabra, representing the dominant floral hue in the native population[24]; the red variety was subsequently identified by R.V. Butt and designated 'Mrs. Butt'. As the most vivid hue among the seven primary color groups, it was later established as a natural hybrid between Bougainvillea peruviana and Bougainvillea glabra. The cultivar displays intermediate traits, with leaves similar to B. peruviana and bracts resembling B. glabra. Spontaneous mutations contributed significantly to the evolutionary development of the 'Mrs. Butt' cultivar, giving rise to numerous novel bract colors[25]. Orange Bougainvillea arose from natural bud mutations of red cultivars, first observed in 'Mrs. Butt' and 'Scarlet Queen'[26]. Pink-toned Bougainvilleas mainly derive from bud mutations of red and purple varieties and are characterized by soft hues, commonly referred to as 'the most romantic color'. Yellow-toned varieties originated as bud mutations within the orange spectrum; the earliest yellow Bougainvillea, 'Mary Baring', originated as a mutation from the orange-toned 'Mrs. McClean'[26]. White Bougainvilleas mainly arise through bud mutations of purple and red varieties and are commonly found in low-light habitats. Their bracts are white due to the absence of pigment-synthesizing genes, whereas bicolored varieties constitute the most chromatically diverse group, exhibiting bracts in multiple hues such as white, pink, red, and purple. These colors appear in patchy or mottled patterns, a morphological trait termed chimerism, commonly resulting from bud mutations or interspecific hybridization[27].
-
As illustrated in Fig. 3, Bougainvillea bracts exhibit substantial morphological diversity. Based on size and petal-like form, they can be classified into four primary types: single-bract, double-bract, butterfly-shaped, and irregular bracts. The single-bract type represents the typical morphology of natural wild species or traditional cultivars, with each inflorescence bearing only three bracts that envelop central florets, which are white or pale-colored. These bracts are proportionally harmonious with the true flowers and retain intact corollas. Representative varieties include 'Mrs. McClean' (Fig. 3a). The double-bract type arises through natural bud mutation or artificial induction and is characterized by a substantial increase in bract number, ranging from 18 to 40 per inflorescence, with the floral tube either reduced or absent, forming a 'tubeless flower' structure. Representative varieties include 'Roseville's Delight' (Fig 3b). Butterfly-type bracts arise from bud mutations of the single-bract type, exhibiting elongated bracts, typically lanceolate or oblong, while retaining true floral structures. Representative varieties include 'Ratana Yellow' (Fig. 3c). Irregular bracts also arise from bud mutations of the single-bract type, frequently exhibiting inward curling or outward flipping of bracts and irregular shapes while maintaining true floral structures[26,29]. Representative varieties include 'Mahsuri Red' (Fig. 3d).

Figure 3.
Schematic diagrams of different bract structures. (a)−(d) Terminal branch structure. (e)−(h) Flower bud structure. (i)−(l) Bract morphology.
Structure and function of bracts
-
Bracts serve not only as ornamental structures but also perform important physiological functions. Visually, the bracts of Bougainvillea mimic petals through their vivid colors, thereby attracting pollinators, including butterflies and bees. Specific bract colors may also draw particular pollinator species, facilitating co-evolution and promoting a stable ecological niche[26]. Furthermore, compared to true leaves, bracts typically exhibit higher light reflectance, which may protect inner flowers from photoinhibition under intense sunlight[30]. Bracts retain photosynthetic capacity, although differing in chloroplast structure and photosynthetic pigments from leaves, contributing partially to the plant's carbon assimilation[31]. Their abundant pigments, such as betacyanins and flavonoids, possess antioxidant activity and potentially mitigate photo-oxidative stress[32].
-
The 'petals' of Bougainvillea typically refer to its bracts, which are derived from leaves and have evolved into specialized structures retaining distinct venation patterns[33]. The flower bracts are the primary ornamental feature of Bougainvillea, with both external form and internal architecture contributing to ornamental value. The external morphology of a single-bracteate Bougainvillea (Fig. 4a1−a6) inflorescence comprises three enclosing leaf-like bracts. The middle portion of the perianth tube constricts, ending in five lobes that constitute the true flower. The pedicel of the true flower is anchored centrally to form an integrated 'true flower–bract' structure with an overall triangular shape. Its internal structure comprises bracts, a perianth tube, a true flower, stamens, and pistils[34]. The double-bract type Bougainvillea (Fig. 4b1−b12) flower bud is composed of multiple overlapping bract layers, forming a pompom-like floral structure. It exhibits a spatially nested structure with multiple concentric layers. The outer bracts provide support and protection, while the inner layers progressively decrease in size, with progressively smaller inner layers exhibiting delicate morphology and often showing color gradients. The floral organs, including pistils, stamens, and perianth tubes, are reduced, resulting in the absence of typical floral structures[35].
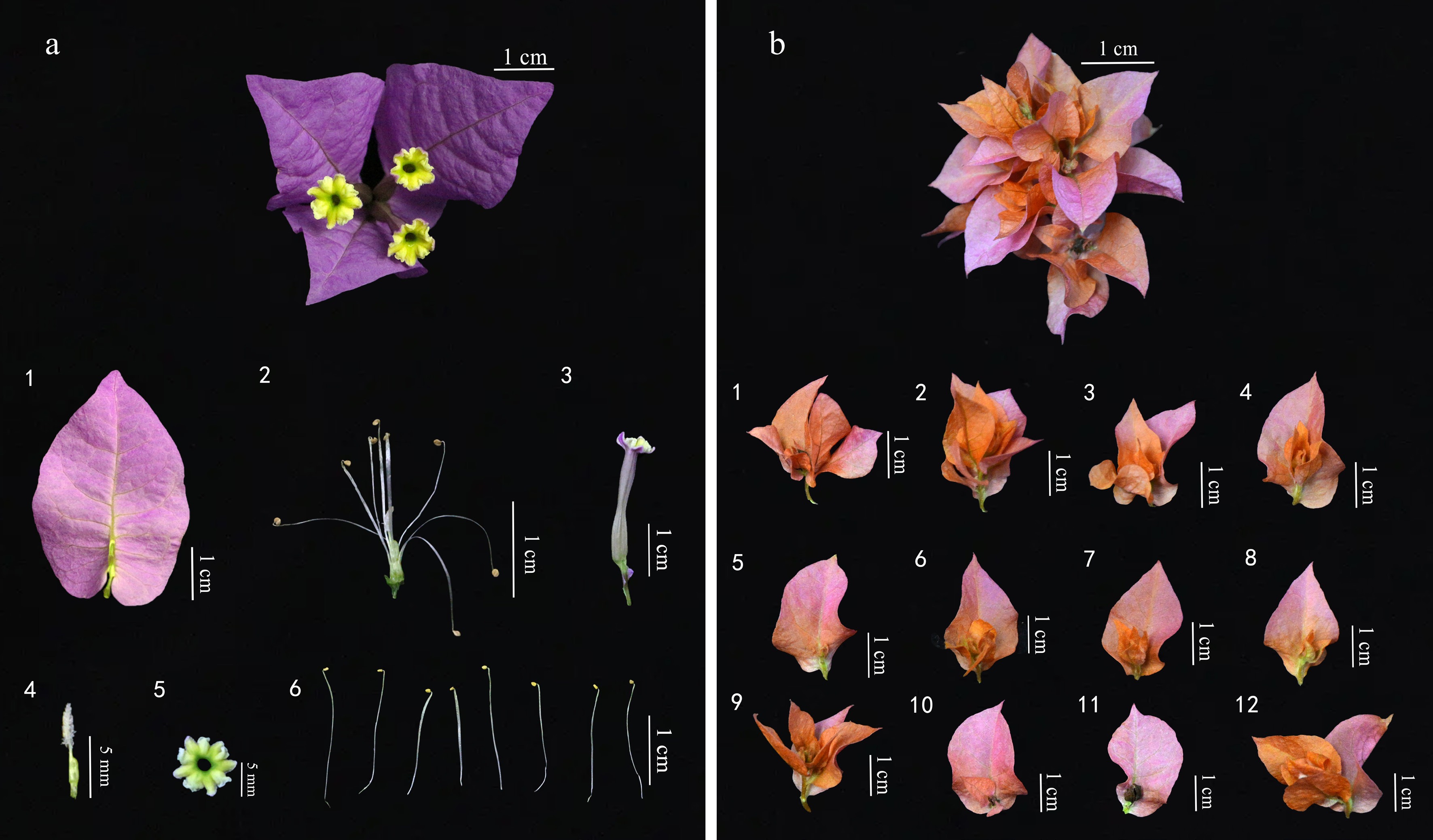
Figure 4.
Comparison of flower bud structures between (a) single-bract Bougainvillea ('President') and (b) double-bract Bougainvillea ('Roseville's Delight'). (a1) Bract. (a2) Stamens and pistil. (a3) Perianth tube. (a4) Pistil. (a5) True flower. (a6) Stamen. (b1)−(b12) Structure of individual bracts.
Developmental process of floral organs
-
The morphogenesis of bracts is tightly regulated by hormonal signaling networks. Key hormones, including gibberellins, abscisic acid, cytokinins, and ethylene, undergo dynamic fluctuations during the early stages of floral bud differentiation. Their synergistic and antagonistic interactions jointly regulate the initiation, expansion, and ultimate morphology of bract primordia. Gibberellins GA7 and GA4 are markedly elevated during early floral differentiation, directly promoting bract morphogenesis. Abscisic acid, trans-zeatin riboside, and the ethylene precursor ACC specifically regulate lateral bract development, whereas dihydrozeatin and salicylic acid contribute to apical bract differentiation. Jasmonic acid influences color differentiation through dynamic expression of JAZ genes, leading to white bracts, whereas downregulation results in purple bracts. Gibberellins and cytokinins synergistically inhibit chlorophyll metabolism, thereby delaying bract greening and extending flowering duration. Concurrently, these hormones coordinate pigment synthesis and bract morphogenesis by regulating cell division and expansion[36].
At the genetic regulatory level, bract development shares regulatory pathways with leaf development yet displays distinct features. As shown in Table 1, gene families such as KNOX, YABBY, and ARP, involved in establishing leaf proximal–distal polarity and organogenesis, are critically involved in bract development[37,38]. The highly specific expression of YABBY family genes is closely associated with bract flattening and morphological diversity[39]. Genes related to auxin transport carriers (e.g., PINs), response factors (e.g., ARFs), and gibberellin synthesis and signaling pathways exhibit marked upregulation during bract development[40]. Auxin concentration gradients likely determine bract morphology, whereas exogenous gibberellin application promotes bract enlargement[41,42]. Cytokinins modulate bract size and number through regulation of the cell division cycle[43].
Table 1. Key genes in floral organ development and their characteristics.
Gene name Functional description Mode of expression Associated traits YABBY (DL, FIL) Regulate apical-meristematic polarity of floral organs; promote flattened growth. Specifically expressed on the apical surface of the bract primordium. YABBY overexpression results in flattened bracts and morphological diversification; mutant bracts show curling or asymmetry. KNOX (STM, BP) Maintain meristem activity;
inhibit differentiation.Expressed in the apical meristem, Downregulated in bracts. Ectopic expression of KNOX induces bract lobes or nodular protuberances and regulates bract number. ARP (AS1, AS2) Antagonizes KNOX;
promotes organ differentiation.In the bract primordia, full expression occurs. AS1/AS2 mutations result in irregularly lobed bracts, affecting bract symmetry. Auxin signalling pathways (PINs, ARFs) Regulate auxin distribution gradients; determine bract morphology. Expressed at the apex or margin of the bract. The PIN mutation causes bract distortion; ARF overexpression results in enlarged bracts. Gibberellin pathway (GID1, DELLA) Mediates GA signalling;
promotes cell elongation.High expression of bracts during the rapid elongation phase. DELLA mutant bract dwarfism; GID1 overexpression increases bract size. Cell cycle genes (Cyclins, CDKs) Regulating cell division. Expression during the active phase of the bract meristem. Downregulation of expression reduces bract size and affects the number of bract cells. Cell wall-associated genes (EXPANSINS, XTHs) Promote cell wall relaxation;
regulate expansion.Highly expressed during the bract expansion stage. EXPANSINS silences hardening of bracts; XTH overexpression confers flexibility to bracts. At the cellular level, the final size of bracts is determined by the coordinated activities of cell division and cell expansion. Cell cycle regulatory genes, such as CYCLINS and CDKs, alongside cell wall relaxation-related genes like EXPANSINS and XTHs, exhibit dynamic expression during bract development, thereby directly regulating bract growth[44,45]. These genes collectively shape the final morphology and size of bracts through the coordinated regulation of cell division and expansion.
Primary types of floral phenotypic variation
-
As a prominent ornamental genus within the family Nyctaginaceae, Bougainvillea displays extensive phenotypic diversity. This not only reflects the species' genetic richness but also serves as a critical basis for germplasm resource characterization, the development of new varieties, and horticultural applications. Phenotypic variation primarily occurs in three traits: bracts, true flowers, and overall plant structure. These variations arise through multiple pathways, including natural hybridization, bud sport mutations, and artificial breeding, forming the key features of Bougainvillea germplasm resources[46].
In terms of bracts, Bougainvillea exhibits substantial variation in shape, color, and structure. Bract shapes include ovate, elliptical, and lanceolate forms. Double-flowered cultivars display complex, multilayered, overlapping structures, whereas single-flowered varieties exhibit simpler structures[47]. Color variation ranges from white, yellow, pink, red, purple, to multicolored patterns. In some cultivars, bract color dynamically adjusts in response to environmental factors such as light exposure, thereby enhancing ornamental value through dynamic visual effects[48]. Bract size and posture also vary significantly, including straight, twisted, or spreading forms. These variations are used not only for cultivar identification but also as key traits for parental selection in hybrid breeding[30].
Variation in true flowers, though less pronounced than in bracts, is important for classification and breeding. True flowers primarily refer to tubular florets, whose floral structures, such as the corolla and perianth tube, vary in length and shape. For instance, the perianth tube may appear slender or swollen at the base[47]. Fertility represents another key aspect of true flower variation. Many cultivated varieties exhibit pollen or ovule sterility, thereby constraining the use of sexual reproduction. However, fertility may be partially restored via approaches such as induced polyploidy[30]. The position and density of inflorescence attachment also vary considerably between cultivars, including compact and loose inflorescence types. These characteristics are closely linked to plant adaptability and ornamental appeal[46].
Overall floral structure variation includes macroscopic traits, such as inflorescence arrangement, attachment patterns, and spatial relationships among floral components. Inflorescence attachment sites include terminal, apical, or distributed along branches, with flowering density ranging from dense to sparse. These characteristics influence both the overall ornamental appeal and the taxonomic value of the flower[47]. For instance, certain cultivars exhibit compact inflorescences suited for container landscaping, whereas others display loose arrangements suited to open growth conditions. The relative positioning of bracts to true flowers and morphological variations in the perianth tube (such as constriction or swelling) represent additional important aspects of overall floral structure. These variations reflect the influence of genetic and environmental interactions[46]. Concurrently, floral structural variation is closely linked to plant adaptability; for example, inflorescence architecture may optimize pollination efficiency or contribute to stress resilience[25].
Mechanism of double-petalled flower formation
-
The double-petalled flower form represents a significant ornamental trait in Bougainvillea, resulting from alterations in the specification of floral organ identity. In model plants such as Arabidopsis thaliana and Antirrhinum majus (Snapdragon), floral organ identity is controlled by interactions among three classes of functional genes: A, B, and C. Primarily belonging to the MADS-box gene family, these genes regulate downstream targets through the formation of protein complexes[48]. Recent studies proposing the 'flower tetramer model' have clarified that MADS-box proteins assemble into tetrameric complexes, interact with CArG-box motifs in target genes, and regulate floral organ development via DNA looping[49].
As shown in Table 2, analysis of the Bougainvillea genome has identified numerous MADS-box genes, with their phylogenetic relationships and expression patterns characterized[50,51]. Research indicates that ectopic expression or loss of function of Class C genes (e.g., AGAMOUS, AG) plays a central role in petalization and double-flowered phenotype formation[52,53]. In single-petalled Bougainvillea, Class C genes are specifically expressed in stamens and pistils, maintaining normal reproductive organ development, whereas in double-flowered cultivars, C-class genes (e.g., BglAG) exhibit significantly reduced expression levels or impaired function in floral organs fated to develop as stamens. This results in the double-flowered phenotype, characterized by multiple layers of bracts[52,54]. Altered expression boundaries of Class B functional genes (e.g., APETALA3, PISTILLATA) also contribute to double-petal morphogenesis. In Bougainvillea, shifts in Class B gene expression patterns are closely associated with petalization, potentially facilitating double-petal formation through modulation of floral organ identity[53,55]. Furthermore, Class E genes (e.g., SEPALLATA), acting as 'glue' proteins, play a crucial role in tetrameric complex formation, with expression changes indirectly affecting double-petalled flower development[48].
Table 2. Functional and expression characteristics of key MADS-box family genes.
Gene name Genetic category Functional description Mode of expression Associated traits BgAP1 Class A Regulates the development of sepals and petals, negatively regulating Class C genes through negative feedback inhibition. Highly expressed in bracts and petals, moderately to lowly expressed in stamens and carpels. Maintain the identity of single sepals and petals; expression imbalance may lead to bract formation. BgAP3 Class B Forms heterodimers with BgPI, determining petal and stamen identity. Highly expressed in petals and stamens, moderately to lowly expressed in bracts. In double-flowered varieties, expression of boundary expansion participates in stamen petalisation. BgPI Class B Interacts with BgAP3 to regulate petal and stamen development. Its expression pattern is similar to that of BgAP3, being enriched in petals and stamens. Alterations in Class B gene expression may induce homoeotic transformation in floral organs. BgAG Class C Determines the identity of stamens and carpels, inhibiting the indefinite growth of floral meristems. Highly specifically expressed in stamens and carpels; downregulated in double-flowered cultivars. Functional loss leads to stamen petalisation, forming a double-petalled flower form. BgSEP Class E As a 'glue' protein, it promotes MADS tetramer formation and regulates the development of all floral organs. Widely expressed in all floral organs, with higher expression in bracts. Altered expression of Class E genes affects protein complex stability, indirectly leading to double petals. BgFULL Class C related Participates in carpel and fruit development, with partial functional overlap with AG. Highly expressed during early floral development, associated with bract expansion. May compensate for the loss of apical meristem function and promote the formation of bract-like structures. Although Bougainvillea and Dianthus chinensis belong to the families Nyctaginaceae and Caryophyllaceae, respectively, they are both members of the order Caryophyllales, with a highly conserved MADS-box regulatory network controlling floral organ development. Therefore, research findings on double-flower formation, floral meristem size, primordium redifferentiation, and boundary gene regulation in carnation provide valuable reference points and testable hypotheses for understanding the mechanisms underlying double-flower formation in Bougainvillea. As shown in Fig. 5, studies on Dianthus chinensis indicate that double-flower formation is closely associated with meristem size and primordia redifferentiation, with stage 3 of floral development (S3) representing a critical period for the initiation of double-flower traits. The floral meristem of double-flowered forms is larger, facilitating the development of additional petal primordia. Concurrently, abnormal division and redifferentiation of primordia during development lead to an increased number of petals. During this process, genes such as DcaAP2L, DcaLFY, and DcaUFO are expressed at petal–stamen boundaries. Alterations in their expression patterns potentially induce stamen-to-petal conversion, resulting in double-petal structures. These genes may also contribute to double-petal morphogenesis by modulating the expression of class B genes (e.g., DcaAP3/PI) within the ABC model[56]. It is thus speculated that the molecular mechanism underlying double-flower formation in Bougainvillea is similar to that characterized in Dianthus.
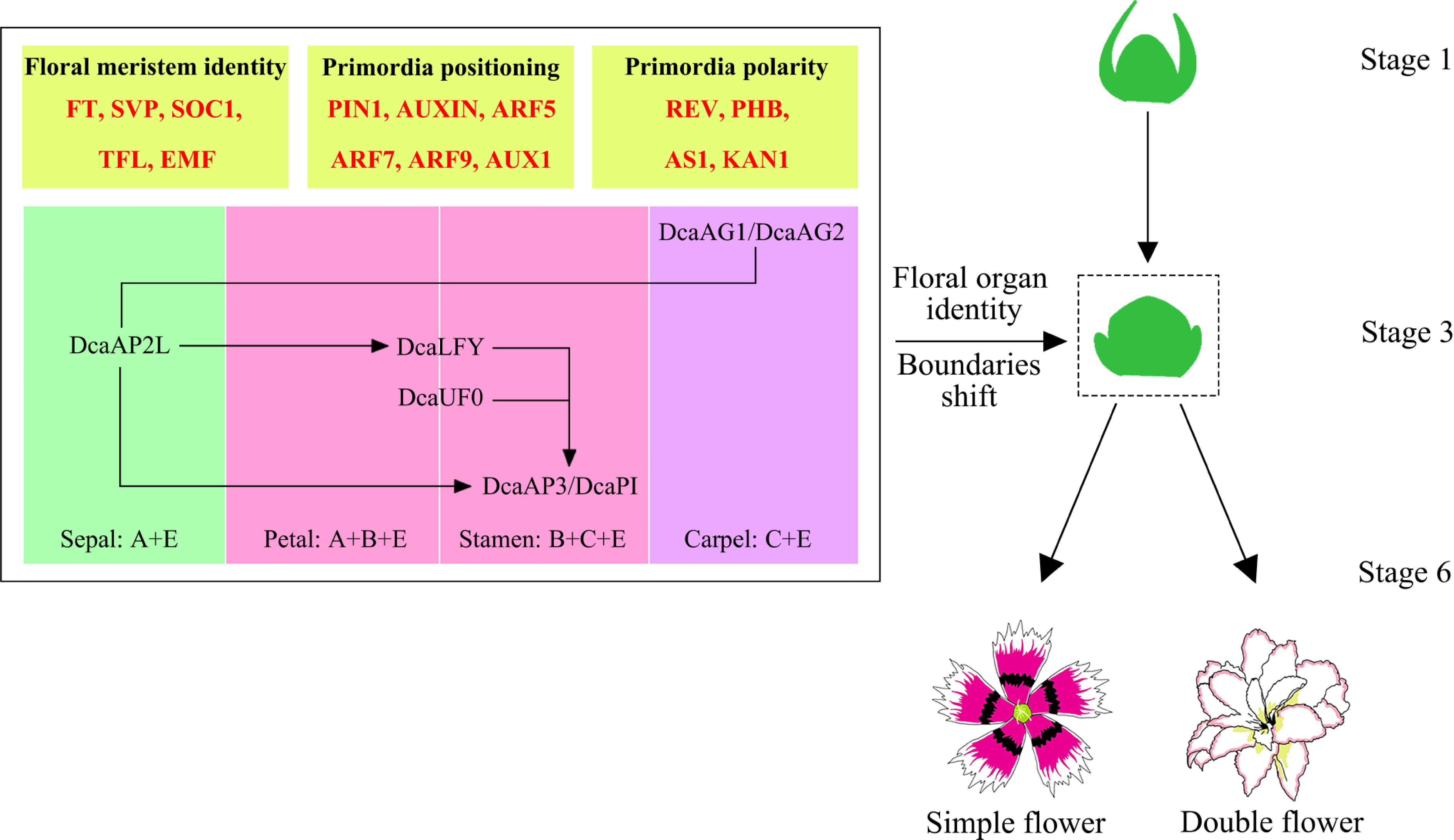
Figure 5.
Transcriptional regulation diagram of single-petalled and double-petalled flowers in Dianthus chinensis[56].
-
Bougainvillea bracts contain a rich variety of pigments, exhibiting distinct pigment compositions and concentrations among color groups. Common pigments include betalains, flavonoids, chlorophylls, and carotenoids, whereas anthocyanins are absent, reflecting the mutual exclusivity of betalains and anthocyanins[57]. The formation and variation of Bougainvillea flower colors are notable within the plant kingdom. While the coloration of most ornamental flowers relies on dynamic metabolic changes in anthocyanins within petals[58−60], Bougainvillea utilizes a distinctive pathway dominated by betalain and flavonoid metabolism[61].
Betalain is a nitrogen-containing, water-soluble pigment with multiple biological activities, first identified in beetroot. As a pyridine derivative, its stability is influenced by pH, light exposure, temperature, oxidants, enzymes, and metal ions. It is categorized into betanin and betaxanthin[62,63]. Betalains are rare in nature and confined to plants of the order Caryophyllales, and no plant is known to produce both pigment types concurrently. For instance, Mirabilis jalapa accumulates vivid betalains in its petals while entirely lacking anthocyanins[64]. As depicted in Fig. 6, betalain biosynthesis uses tyrosine as its precursor via a sequence of enzymatic reactions[65]. Tyrosine is first hydroxylated into L-DOPA, catalyzed by tyrosine hydroxylase or cytochrome P450 enzymes such as CYP76AD1, representing a key step in betalain synthesis[65,66]. L-DOPA is subsequently oxidatively cleaved by DOPA 4,5-dioxygenase (DOD) to yield betalamic acid, which serves as the chromophore for betalain. This compound can spontaneously or enzymatically condense with cyclo-DOPA derivatives to form betanin, resulting in deep red to purplish-red bracts. Alternatively, it may combine with amino acids and amines to generate betaxanthin; higher betaxanthin accumulation produces yellow to orange bracts in Bougainvillea[65,67]. Betalain biosynthesis also includes several post-synthetic modification steps. For instance, cyclo-DOPA can form cyclo-DOPA glucoside, catalyzed by glucosyltransferase, subsequently condensing with betanin to form betanin glycoside. Acylation modifications via acyltransferase further increase pigment stability[57,65]. Transcriptional regulation also contributes to this pathway; for example, WRKY transcription factors promote betalain accumulation through binding to the W-box in the CYP76AD1 promoter region[57]. The entire pathway occurs in the cytoplasm, involving multiple oxidation, condensation, and modification reactions, producing structurally diverse betalain derivatives[68].
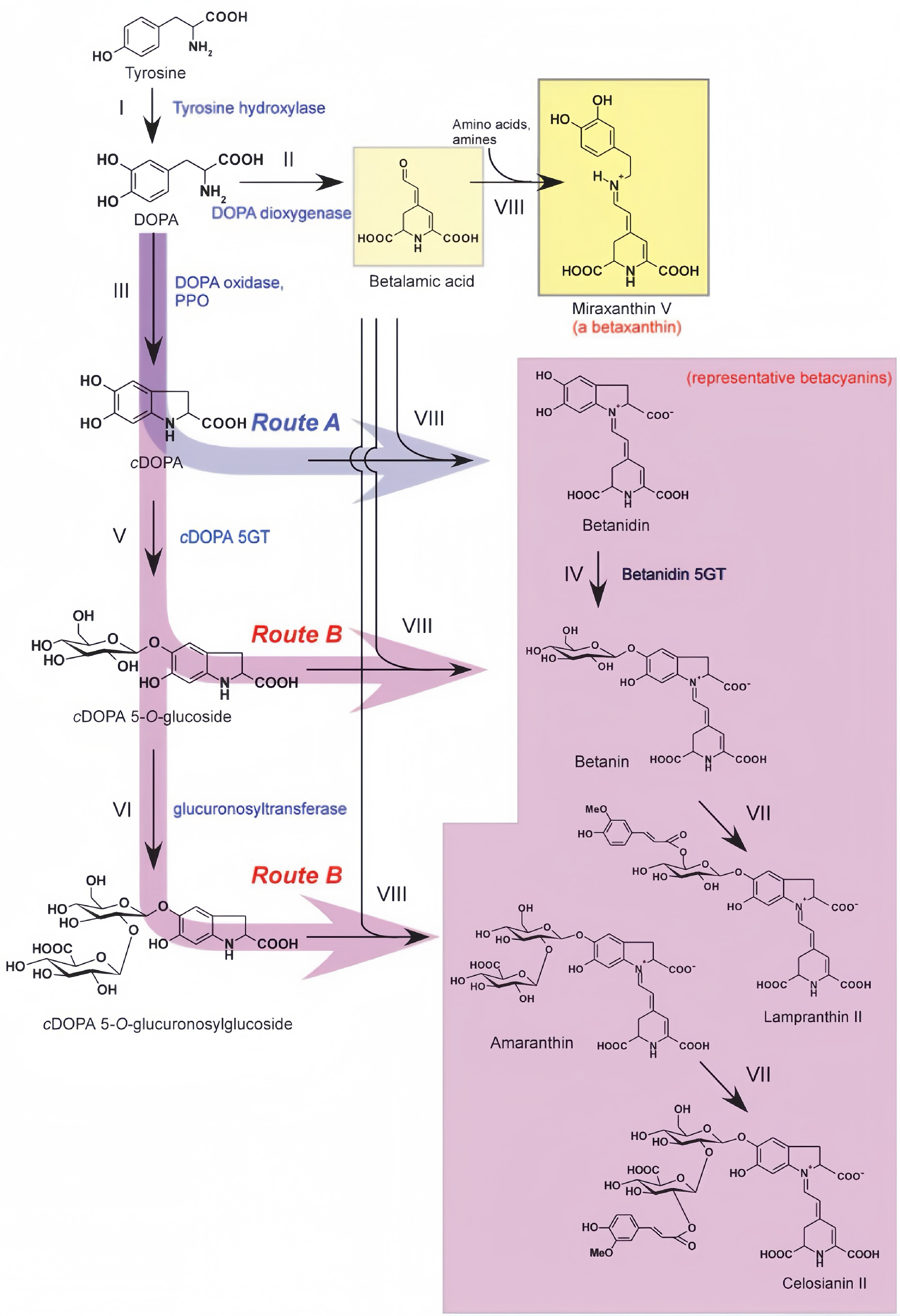
Figure 6.
Biosynthetic pathway of betalain[65].
As shown in Fig. 7, the flavonoid biosynthesis pathway originates from the phenylpropanoid pathway, where phenylalanine is deaminated by phenylalanine ammonia-lyase (PAL) to generate cinnamic acid. Cinnamic acid is then hydroxylated by cinnamic acid 4-hydroxylase (C4H) and converted by 4-coumaroyl-CoA ligase (4CL) into 4-coumaroyl-CoA[65,69]. The expression levels of these enzymes directly impact bract pigment biosynthesis. 4-coumaroyl-CoA condenses with three molecules of malonyl-CoA, catalyzed by chalcone synthase (CHS), to generate chalcones, including naringenin chalcone. Chalcones are subsequently isomerized by chalcone isomerase (CHI) into the flavone hesperetin, forming the central branching point of the C6–C3–C6 flavonoid framework[65,70]. In bract color regulation, CHS, the initial enzyme in the flavonoid pathway, exhibits expression levels positively correlated with total flavonoid content, thus influencing bract color intensity[65]. Conversely, altered CHI expression may promote chalcone accumulation, resulting in enhanced yellow pigmentation, as supported by the association between elevated flavanol accumulation and yellow coloration in Bougainvillea[71].
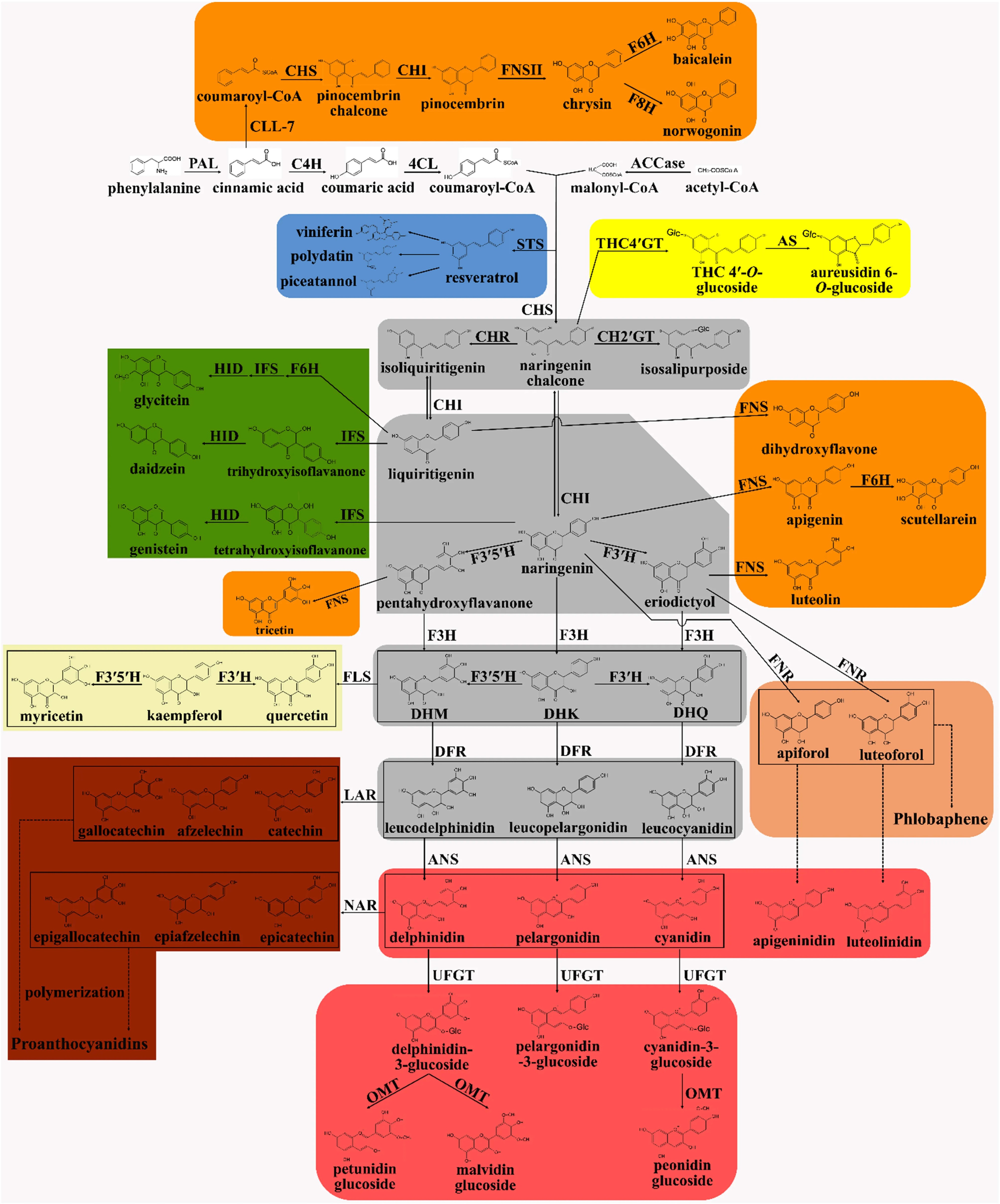
Figure 7.
Biosynthetic pathway of flavonoids[70].
Starting from the flavone hesperetin, its metabolism diverges into two principal pathways:
(1) Anthocyanin synthesis pathway: The biosynthesis of anthocyanins involves hydroxylation of flavanones by F3H, reduction by DFR to produce leucoanthocyanidins, followed by oxidation via ANS to yield pigmented anthocyanins, and subsequent modifications such as glycosylation by UFGT to enhance pigment stability[72]. However, the expression of these genes is restricted in Caryophyllales plants, explaining the absence of bluish-purple bract coloration in this order[65].
(2) Alternative flavonoid pathways: Flavanones are converted into flavanols via flavanone synthase (FNS) or into isoflavones via isoflavone synthase (IFS), while chalcones are transformed into aurantiamides (yellow pigments) through the action of chalcone-2'-glucosyltransferase (CH2'GT) and aurantiamide synthase (AS)[65,70]. In Bougainvillea bracts, high accumulation of flavanols, such as rutin, directly contributes to yellow bract pigmentation, regulated by the expression of CHI and flavanol synthase (FLS)[71].
Competition and interaction mechanisms between betalain and flavonoid metabolism
-
In Bougainvillea, betalain and flavonoid metabolic pathways exhibit substantial competition and interaction. These interactions are primarily mediated through substrate competition within metabolic pathways, regulation of key enzyme gene expression, and mutual exclusion of pigment accumulation. Betalains and anthocyanins, both as plant pigments, generally display mutually exclusive distribution patterns in nature. However, Bougainvillea, as a betalain-accumulating species, also accumulates detectable levels of flavonoids, including quercetin and kaempferol, suggesting potential cross-talk or shared precursor molecules between these metabolic pathways[73]. For instance, in Bougainvillea × buttiana 'Chitra', red and pale yellow bracts coexist simultaneously during peak flowering. Red bracts exhibit significantly higher betacyanin content than pale yellow bracts, whereas the latter contain markedly higher levels of flavonoids, particularly flavanols, indicating a reciprocal allocation between the two pigment pathways[74].
This interaction partly stems from common upstream nodes in the phenylpropanoid metabolic pathway. Phenylalanine ammonia-lyase (PAL) and chalcone synthase (CHS), key enzymes in flavonoid synthesis, are upregulated, which promotes flavonoid accumulation and concurrently suppresses betalain synthesis through competitive substrate utilization[75]. As shown in Fig. 8, in 'Chitra', key genes in the flavanol synthesis pathway, such as BbCHS, BbCHI, BbF3H, BbFLS, and BbCYP75B1, are highly expressed in pale yellow bracts, resulting in the accumulation of flavanol compounds, including kaempferol and quercetin. Conversely, key genes in the betacyanin synthesis pathway (e.g., BbDODA) are highly expressed in red bracts, facilitating betacyanin accumulation. Both pathways share the precursor tyrosine, thereby directly competing for this shared substrate[74].
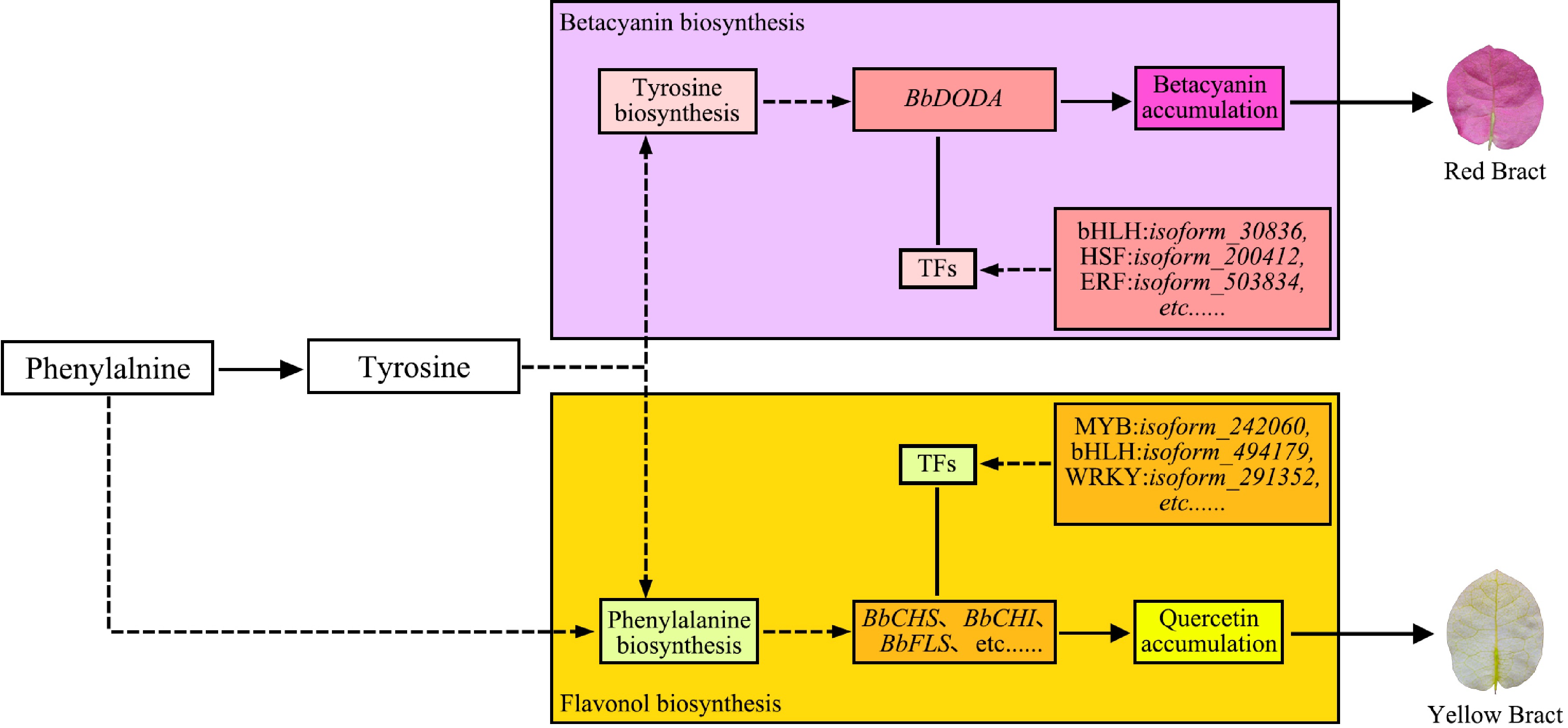
Figure 8.
Molecular mechanism model of bract colour formation in 'Chitra'[74].
As shown in Table 3, joint transcriptomic and metabolomic analysis revealed pronounced enrichment of flavonoid biosynthetic pathways in Bougainvillea bracts, including genes such as CHS, F3H, and FLS, alongside the betalain biosynthetic pathway, represented by the DODA gene. The flavonoid pathway exhibited greater enrichment relative to the betalain pathway, suggesting that flavonoid metabolism potentially suppresses betalain biosynthesis indirectly through consumption of shared precursors[76]. Expression patterns of key enzyme genes are closely associated with bract color transformation. For instance, the F3H gene exhibits low basal expression in white bracts but is significantly upregulated in purple-flowered varieties such as Bougainvillea glabra 'Alba', facilitating the conversion of dihydrokaempferol into anthocyanin precursors, thereby contributing to the purple-red coloration of bracts[77]. Similarly, the DFR gene exhibits high expression in orange bracts, such as Bougainvillea × buttiana 'Baolao Cheng', with substrate specificity favoring dihydrokaempferol (DHK). This catalyzes the formation of the colorless geraniol aglycone, resulting in the accumulation of orange pigments[78]. Concurrently, key betalain pathway genes such as cDOPA5GT display inverse expression patterns relative to flavonoid pathway genes like ANS, confirming competitive interactions between these two pigment pathways[79].
Table 3. Functional and expressive characteristics of key genes in the beet pigment and flavonoid metabolic pathways.
Gene name Functional description Mode of expression Associated traits PAL Catalyzing the conversion of phenylalanine to cinnamic acid. Highly expressed in magenta bracts and less so in pale yellow bracts. Indirect regulation of red and yellow phenotypes in bracts. CHS Catalyzing the formation of cinnamoyl-CoA from chalcone. Significantly highly expressed in pale yellow bracts, with lower expression in red bracts. Regulating the formation of pale yellow bracts. F3H Catalyzing the conversion of naringenin to dihydroflavonol. Expression is higher in pale yellow bracts and lower in red bracts. Promote the accumulation of yellow pigment. DFR Reduction of dihydroflavonol to form colorless anthocyanins. Highly expressed in orange or purple bracts, with lower expression in red bracts, which are dominated by betacyanins. Promote the formation of orange or purple bracts. ANS Catalyzing the formation of anthocyanins from colorless anthocyanidins. In beetroot pigment plants, it is typically expressed at low levels or not expressed at all. Anthocyanins are absent in Bougainvillea. TYR Catalyzing the hydroxylation of tyrosine to produce L-DOPA. Highly expressed in red bracts and less so in pale yellow bracts. Promoting the formation of red bracts. DOPA Catalyzing the conversion of L-DOPA into betacyanin precursors. Significantly highly expressed in red bracts and lowly expressed in pale yellow bracts. Promoting the formation of red bracts. CYP76AD Participation in L-DOPA hydroxylation. Highly expressed in red bracts and negatively correlated with the expression of flavonoid pathway genes. Influencing the colour ratio of red and yellow bracts. Interactions between genes, hormones and environmental factors
-
During the formation of Bougainvillea bracts, genes, hormones, and environmental factors interact through multi-tiered transcriptional regulatory networks, precisely modulating the balance and competitive dynamics between betalain and flavonoid metabolism[80,81]. As shown in Fig. 9, at the hormonal signaling level, auxin modulates ARF transcription factor activity via degradation pathways mediated by the AUX/IAA gene family, thereby controlling the expression of major pigment-related transcription factors such as MYB and bHLH. This, in turn, influences the transcription of downstream structural genes, including CYP76AD1 and F3H[82,83]. Abscisic acid (ABA) regulates endogenous ABA levels via the LbNCED1 gene within the NCED family, activating the MYB-bHLH-WD40 transcription factor complex to directly upregulate flavonoid synthesis genes such as CHS and F3H[84]. Jasmonic acid (JA) may positively regulate the expression of betalain pathway genes, including DODA and CYP76AD1, by releasing MYC2 transcription factors through JAZ protein degradation[85], whereas gibberellin (GA) inhibits JA signaling via DELLA proteins, establishing antagonistic hormonal crosstalk[86,87]. At the environmental level, light signals are perceived via phytochromes in response to red and far-red light, modulating PIF transcription factor activity, which in turn regulates pigment gene promoter binding. Temperature fluctuations induce heat shock protein expression through HSF transcription factors, affecting cytochrome P450 metabolic enzymes such as members of the CYP450 family. Water stress may further amplify pigment metabolic responses via ABA signaling[88−90]. Collectively, these inputs integrate into a core regulatory network in which transcription factor complexes (e.g., MYB-bHLH-WD40) respond to hormonal and environmental cues, modulating structural gene expression via cis-acting elements and ultimately generating bract color diversity by modulating the relative accumulation of betalains and flavonoids.
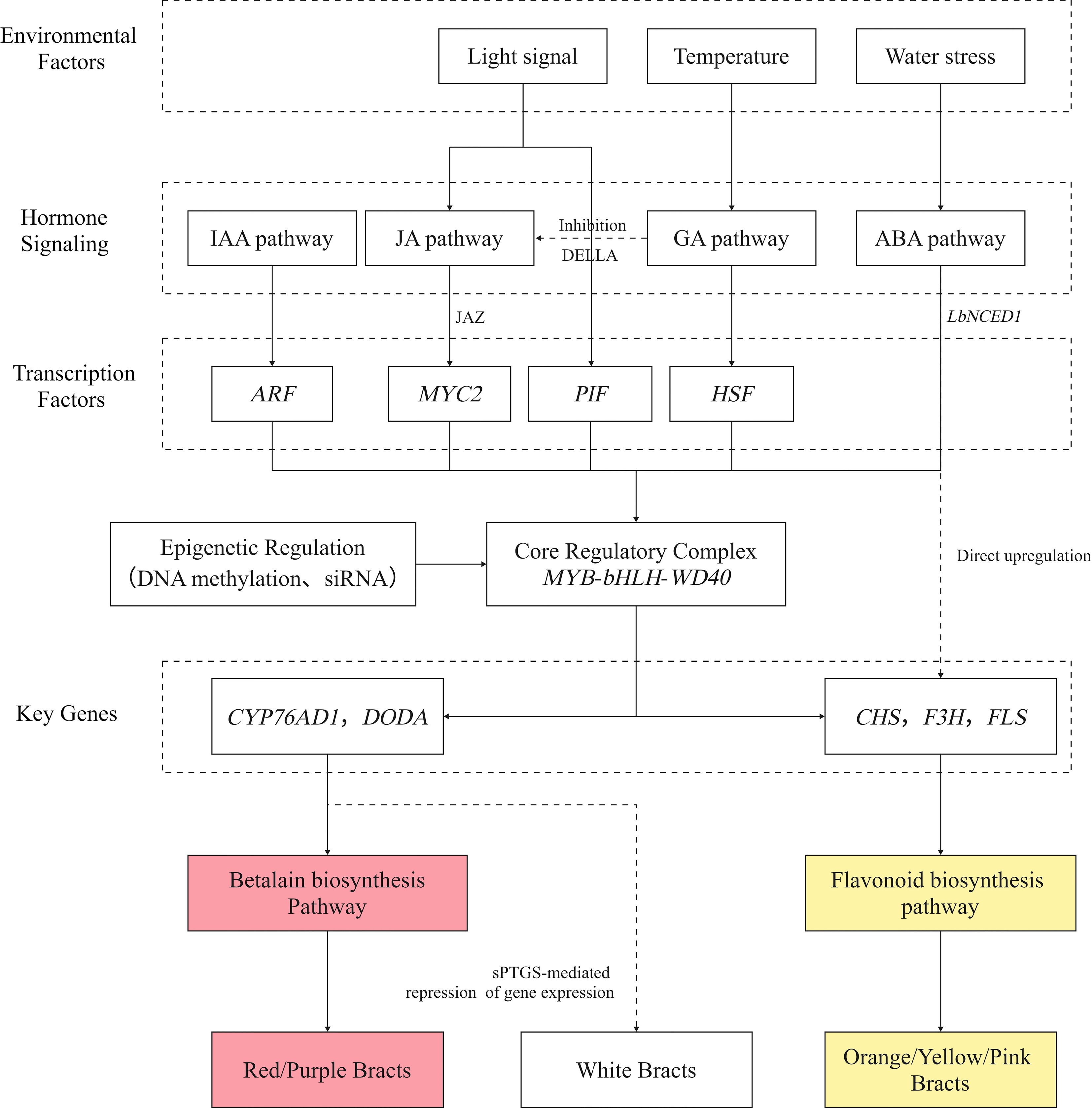
Figure 9.
Regulatory diagram illustrating how genes, hormones, and environmental factors influence the formation of Bougainvillea bract coloration.
Gene expression patterns are closely associated with bract color transitions in Bougainvillea. For instance, F3H gene expression is upregulated during the white-to-pink bract development stage, leading to the accumulation of flavanols, such as quercetin, contributing to yellow pigmentation. Conversely, CYP76AD1 is highly expressed in red bracts, promoting betalain synthesis via tyrosine hydroxylation. Additionally, post-transcriptional gene silencing (PTGS) of CYP76AD1 results in a white phenotype[74,75,91]. This dynamic interactive network confers bract color plasticity and facilitates long-term adaptive responses through epigenetic mechanisms, including DNA methylation and siRNA-mediated gene silencing.
-
As a tropical and subtropical woody plant of exceptional ornamental value, Bougainvillea has long attracted attention in horticultural and plant biology research owing to its unique bract morphogenesis, extensive floral color diversity, and varied floral traits. In recent years, substantial progress has been achieved across several research domains. Regarding bract development, studies have shown that the morphogenesis of bracts, as specialized leaves, is regulated by complex interactions among hormonal pathways (e.g., gibberellins, cytokinins) and critical gene families (e.g., YABBY, KNOX, ARP). The delicate balance between cell division and expansion ultimately determines bract size and morphology. Regarding floral color formation, studies have established that Bougainvillea's coloration is predominantly determined by two metabolic pathways: betalain and flavonoid biosynthesis. These pathways compete and interact at the levels of substrate availability, key enzyme gene expression, and transcriptional regulation. The coordinated allocation and balance of metabolic fluxes underpin the rich spectrum of bract colors, including red, yellow, orange, purple, white, and multicolored patterns. Regarding floral trait variation, particularly the formation of double-flowered forms, functional variation or altered expression of Class C and Class B MADS-box genes acts as a principal determinant. These changes induce transformations in floral organ attributes, such as stamen petalization, thereby generating double-flowered structures enveloped by multiple layers of bracts. Collectively, these findings signify a new era in the molecular study of Bougainvillea.
Nevertheless, current research exhibits limitations, with several avenues meriting further exploration:
(1) Current analyses of traits, including bract development and flower color formation, predominantly rely on association studies based on omics datasets. Future studies should focus on integrating genomic, transcriptomic, proteomic, metabolomic, and epigenomic datasets to construct more refined and dynamic regulatory networks. Concurrently, the establishment of efficient genetic transformation and gene-editing systems is imperative for the functional validation of candidate regulatory genes, facilitating the transition from correlational analyses to causal investigations[92,93].
(2) Beyond floral color and morphology, research on other key traits of Bougainvillea remains relatively limited. For instance, climbing habit is associated with hormone signaling and mechanoperception; flowering time is regulated via photoperiod and vernalization pathways; and distinctive fragrances derive from the biosynthesis of volatile organic compounds. These areas remain largely unexplored and merit systematic investigation[94,95].
(3) In terms of breeding applications, future studies should emphasize the extensive application of genomics-assisted breeding strategies in ornamental plants. Specifically, molecular markers associated with sterility, flower color, and stress tolerance can be employed for marker-assisted selection. When combined with precise gene-editing technologies, this approach can accelerate the development of novel cultivars featuring unique flower colors, improved stress tolerance, and suitability for container cultivation[96,97].
(4) Bougainvillea is rich in secondary metabolites, including flavonoids, phenolics, and betalains, and exhibits significant medicinal potential. Multi-omics approaches, including genomics, transcriptomics, and proteomics, facilitate systematic elucidation of the biosynthetic pathways and regulatory networks governing these bioactive compounds, providing valuable genetic resources and a theoretical foundation for the development of plant-derived pharmaceuticals via synthetic biology strategies[2,98].
In summary, Bougainvillea, owing to its distinctive biological traits and considerable economic significance, is increasingly recognized as a focal point in both fundamental and applied research on ornamental plants. With the continued advancement of multi-omics technologies and biotechnological approaches, our comprehension of Bougainvillea is poised to deepen, ultimately facilitating a shift from merely 'understanding nature' to actively 'designing nature.' Such progress will enable the targeted development of novel, high-quality Bougainvillea cultivars that are better aligned with future market demands.
Sincere thanks to all the authors who participated in this paper.
-
The authors confirm contribution to the paper as follows: study conception and design: Lin H, Du X, Dai S; literature search, collection, and organization, draft manuscript preparation: Lin H, Du X, Ye Z; Wen Z; literature analysis and synthesis: Lin H, Du X, Ye Z, Wang F; manuscript review and editing: Dai S, Wen Z, Wang F. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article, as no new datasets were generated or analyzed during this study
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lin H, Du X, Ye Z, Wen Z, Dai S, et al. 2026. Bract development, flower color diversity, and floral trait variation in Bougainvillea: a comprehensive review. Ornamental Plant Research 6: e025 doi: 10.48130/opr-0026-0017 

Bract development, flower color diversity, and floral trait variation in Bougainvillea: a comprehensive review
- Received: 08 December 2025
- Revised: 02 April 2026
- Accepted: 20 April 2026
- Published online: 14 July 2026
Abstract: Bougainvillea spp., a tropical and subtropical woody plant of the family Nyctaginaceae, is highly valued for its ornamental traits, particularly its vibrant and diverse bracts. Despite its widespread horticultural applications, the molecular mechanisms governing bract development, flower color variation, and other floral traits remain largely unresolved. Recent advances in genome sequencing and multi-omics technologies have significantly advanced biological research on Bougainvillea. This review provides a systematic summary of recent progress, focusing on three aspects: (1) the hormonal regulatory network involved in bract development, including gibberellin and cytokinin signaling pathways and their interactions with key genes such as YABBY, KNOX, and ARP; (2) the formation of floral color, emphasizing the independent roles of betalain and flavonoid metabolic pathways and their interactions through substrate competition and transcriptional regulation; and (3) floral trait variation, integrating classical physiological and biochemical studies to highlight the functional diversification of MADS-box genes in the development of double-flowered forms. Collectively, this review establishes a comprehensive framework from phenotype to molecular mechanisms, providing a theoretical foundation for the molecular breeding and genetic improvement of Bougainvillea.