









-
Salt stress is a major abiotic factor limiting plant growth and development and reducing crop yield worldwide[1]. Sodium (Na+) accumulation in plants disrupts various physiological and biochemical processes, such as nutrient homeostasis, water balance, photosynthesis, hormone metabolism and balance, and antioxidant metabolism[2]. Priming of plants with various chemicals, including organic acids, has been found to be an effective approach to enhance plant tolerance to abiotic stress, including salt stress[3−5].
Acetic acid, as a small molecule that can be synthesized in plants, was not well studied or recognized in terms of its role in plant abiotic stress until a recent study reported its role as an acetylation factor in histone post-translational modification that affected plant drought tolerance[6]. The de novo acetate biosynthesis pathway branches off the main glycolytic pathway at pyruvate[6]. In the first step, pyruvate decarboxylase (PDC) catalyzes pyruvate to acetaldehyde[7]. Subsequently, acetaldehyde is converted to acetate concomitantly reducing NAD+ to NADH by aldehyde dehydrogenase (ALDH)[8]. It has been found that drought stress triggers a dynamic metabolic flux conversion from glycolysis into acetate synthesis by activating the transcription of both PDC1 and ALDH2B7[6]. In addition, exogenous acetate application or overexpression of PDC1 and ALDH2B7 significantly promoted drought tolerance in Arabidopsis[6]. In other studies, expression of PDCs and ALDHs can be induced by the phytohormone ABA[9] and response to other abiotic stresses, including cold[10], oxidative stress[10], and wounding[11].
Exogenous acetic acid treatment improved drought tolerance in Arabidopsis (Arabidopsis thaliana), maize (Zea mays), rice (Oryza sativa), rapeseed (Brassica napus L.), and wheat (Triticum aestivum L.) plants, which has been associated with promoted JA biosynthesis and signaling, but not abscisic acid (ABA)[6]. In cassava (Manihot esculenta), the results of transcriptome profiling indicated that acetic acid treatment improved drought tolerance by upregulating ABA signaling pathway genes[12]. Rahman et al. (2019) found that acetic acid treatment mitigated salt toxicity by increasing uptake of Ca2+ and Mg2+, reducing accumulation of toxic Na+, improving water use efficiency, enhancing accumulation of compatible solutes, and activating catalase activity in mung bean (Vigna radiata)[2]. It is well known that phytohormones, including ABA, JA, IAA, and CK, are crucial endogenous chemical signals regulating plant salt stress tolerance[13, 14]. Salt stress interrupts hormone metabolism[15]. For example, high salinity rapidly increases endogenous abscisic acid (ABA) content and its biosynthetic gene expression levels in plants[16]. Under higher salinity conditions, the endogenous cytokinin (CK) level was significantly decreased and its biosynthesis genes were supressed in perennial ryegrass[17]. It has been found that biosynthesis and signaling pathways of jasmonate (JA) was activated by salt stress, indicating that JA could act as an effective protector against salt stress[18]. Despite available research on the positive effects of acetate on abiotic stress tolerance, the mechanism of acetate underlying salt tolerance regulation is not well understood.
The objective of this study was to understand if acetic acid affects plant tolerance to salt stress, and if so, how. Perennial ryegrass (Lolium perenne L.) used in this study, is widely used as forage and turf grass in temperate regions worldwide but is sensitive to salt stress[19]. Results of this study revealed that salt stress induced de novo biosynthesis of acetate, and exogenous application of acetic acid significantly improved plant salt tolerance, which was associated with increased K+ uptake, suppressed ROS accumulation by activating enzymatic antioxidant systems, and altered phytohormones biosynthesis and signaling pathways. Results of this study pinpointed the novel role of acetic acid biosynthesis in plants and the application potential of this small molecule as a plant protective agent.
-
As shown in Fig. 1, salt stress significantly accelerated leaf senescence and exogenous application of acetic acid significantly alleviated this process: namely, the turf quality (TQ), photochemical efficiency (Fv/Fm) of the photosystem II, and chlorophyll (Chl) content were all significantly higher in acetic acid-treated plants than those of controls at 14, 21, and 28 d of salt stress, while the contrary was true for the degree of plasma membrane damage reflected by electrolyte leakage (EL). It was also notable that, under non-stress condition, plants treated with or without acetic acid have no significant difference in terms of these leaf senescence related parameters (Fig. 1).
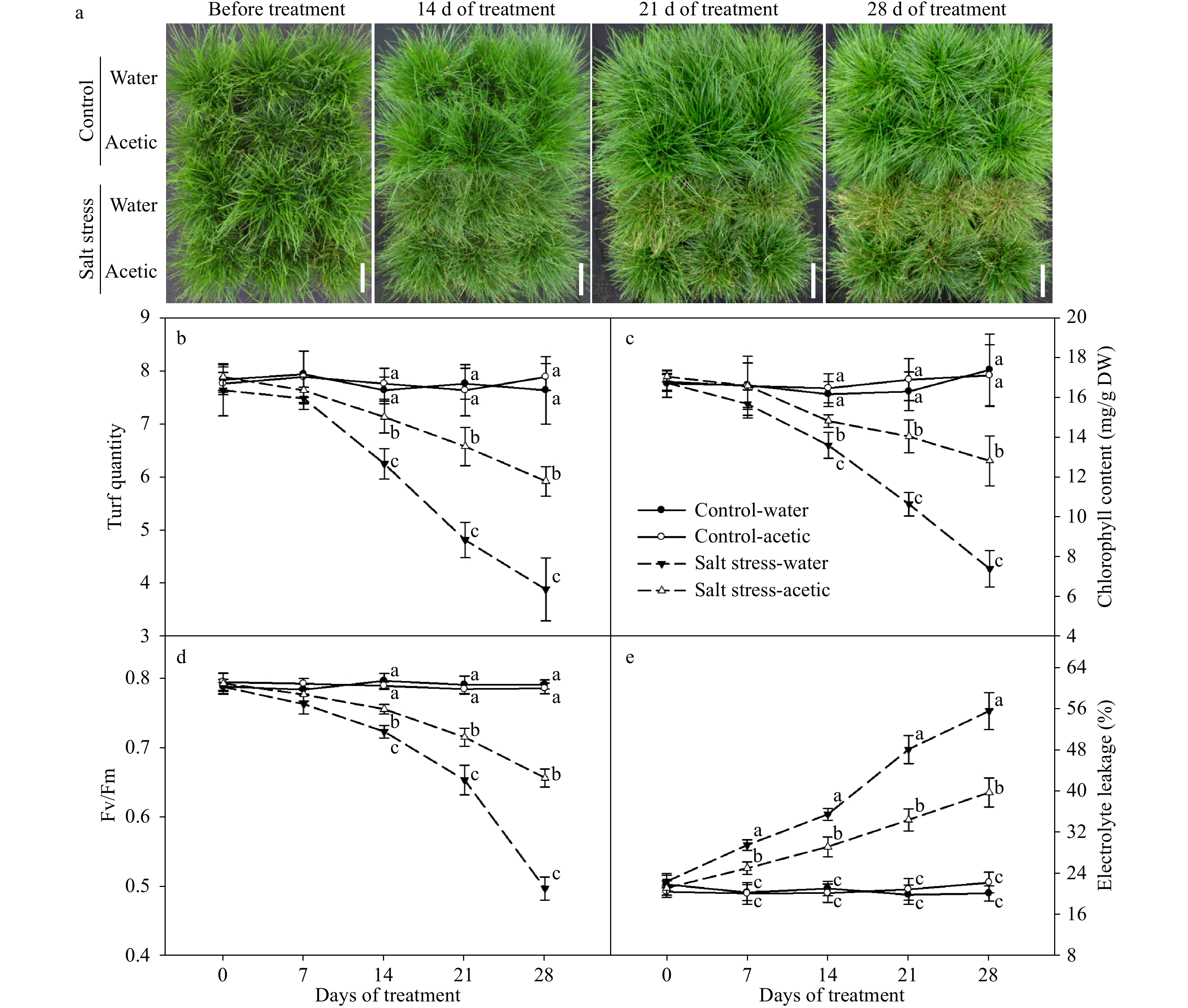
Figure 1. Effects of acetic acid on turf quality and leaf senescence of perennial ryegrass under control or salt stress conditions. (a) Phenotype; (b) turf quality; (c) Chl content; (d) photochemical efficiency; (e) electrolyte leakage. Data labeled with different letters indicate significant differences among the treatments at a given day based on Fisher’s protected LSD test at P ≤ 0.05 probability level (n = 4 in b, c, d, & e).
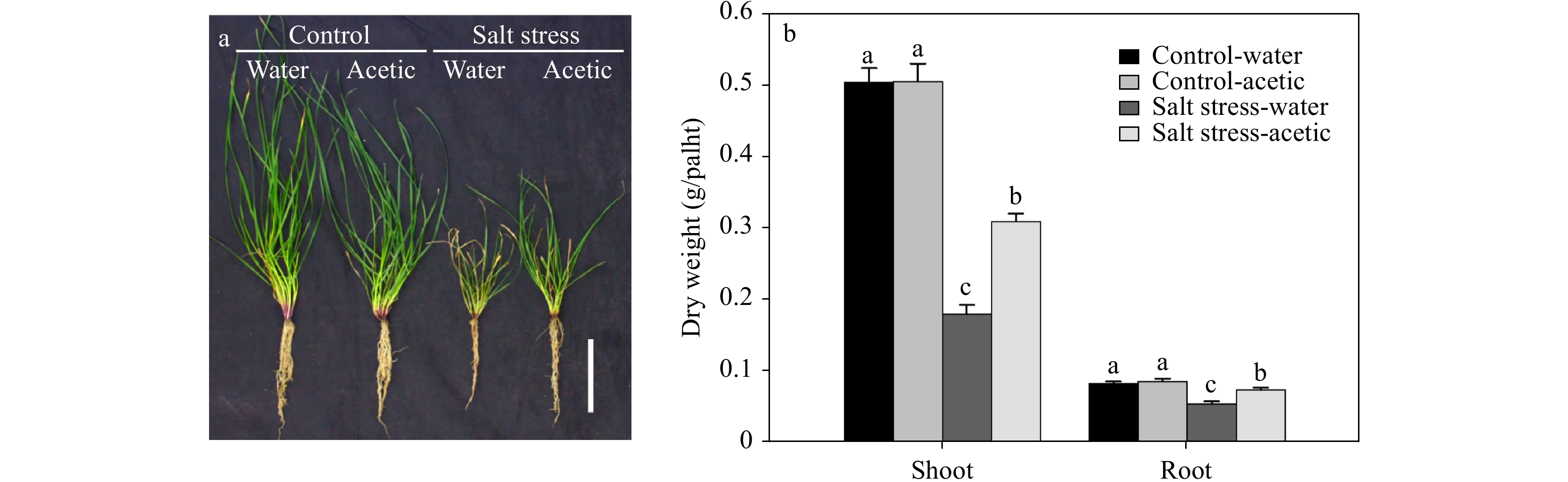
Figure 2. Effect of acetic acid on above-ground and root biomass of perennial ryegrass under control and salt stress conditions. (a) Phenotype of individual plants; (b) dry weight per plant. Photo and data were collected at 28 d of salt stress. Bars labeled with different letters indicate significant differences among the treatments at a given day based on Fisher’s protected LSD test at P ≤ 0.05 probability level (n = 4 in b).
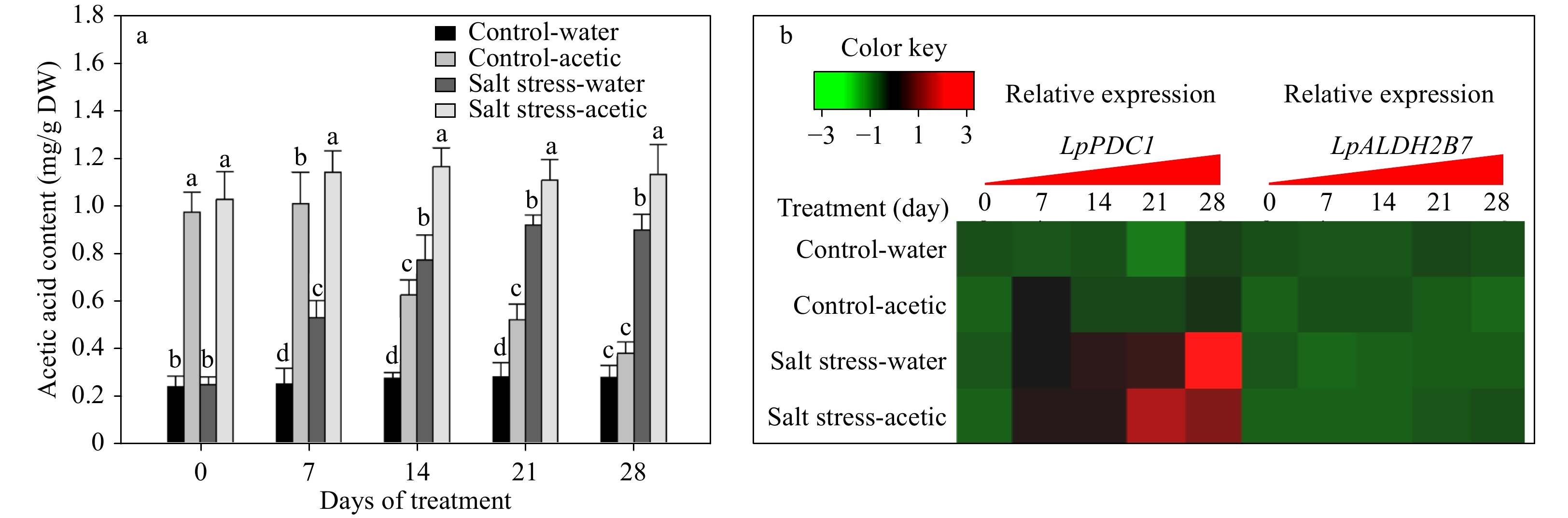
Figure 3. Effect of exogenous application of acetic acid on endogenous acetic acid levels, and the transcription of two acetic acid biosynthesis genes, LpPDC1 and LpALDH2B7, in perennial ryegrass leaves under control and salt stress conditions. Bars labeled with different letters indicate significant differences among the treatments at a given day based on Fisher’s protected LSD test at P ≤ 0.05 level (n = 4 in a & b).
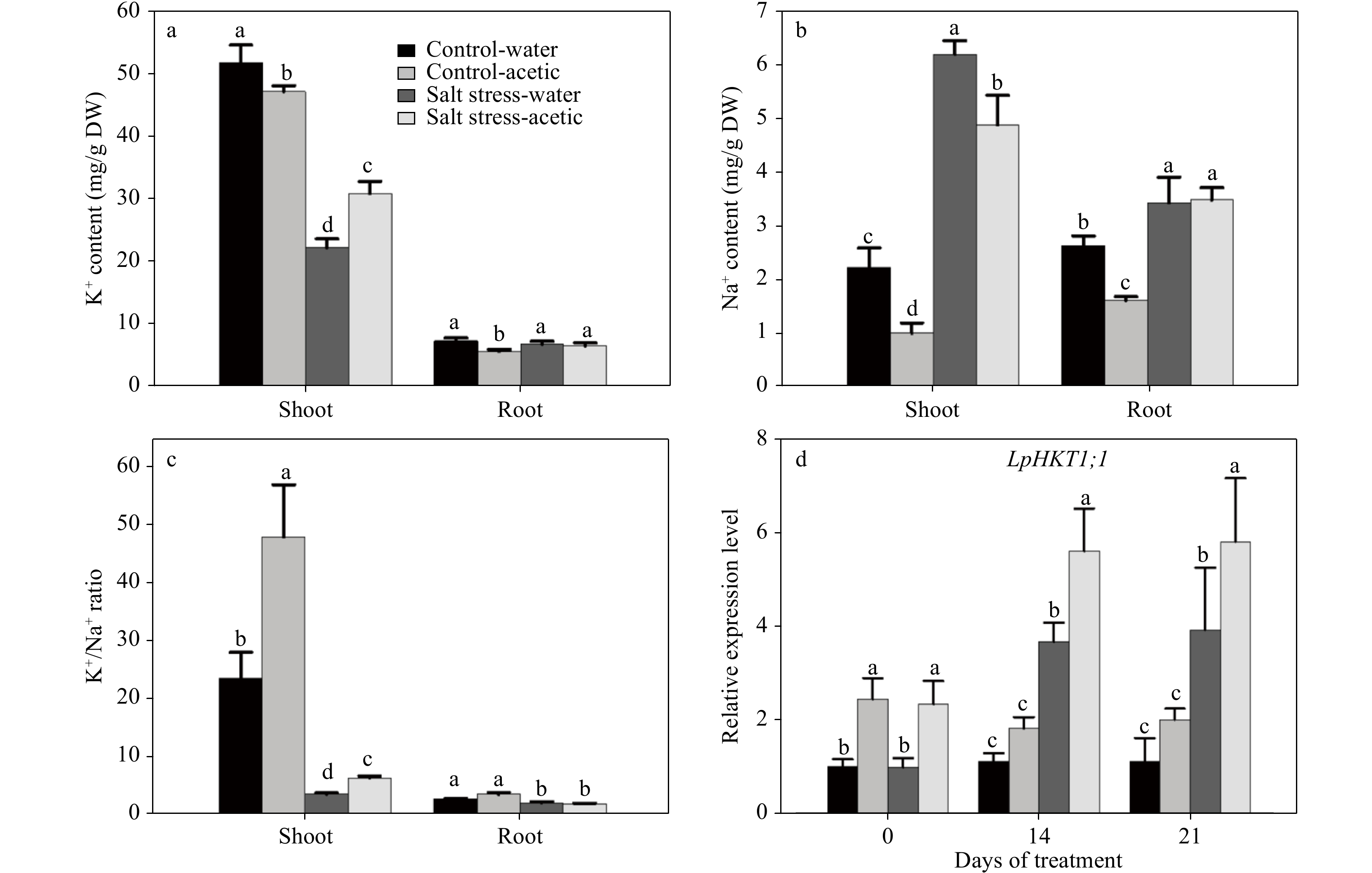
Figure 4. Effect of exogenous application of acetic acid on K+ and Na+ content, K+/Na+ ratio, and expression of LpHKT1;1 in perennial ryegrass under control and salt stress conditions. Data in (a), (b), and (c) were collected at 21 d of treatment. (d) the expression of LpHKT1;1 in leaves were analyzed at 0, 14, and 21 d. Bars labeled with different letters indicate significant differences among the treatments at a given day based on Fisher’s protected LSD test at P ≤ 0.05 level (n = 4 in a, b, & c).
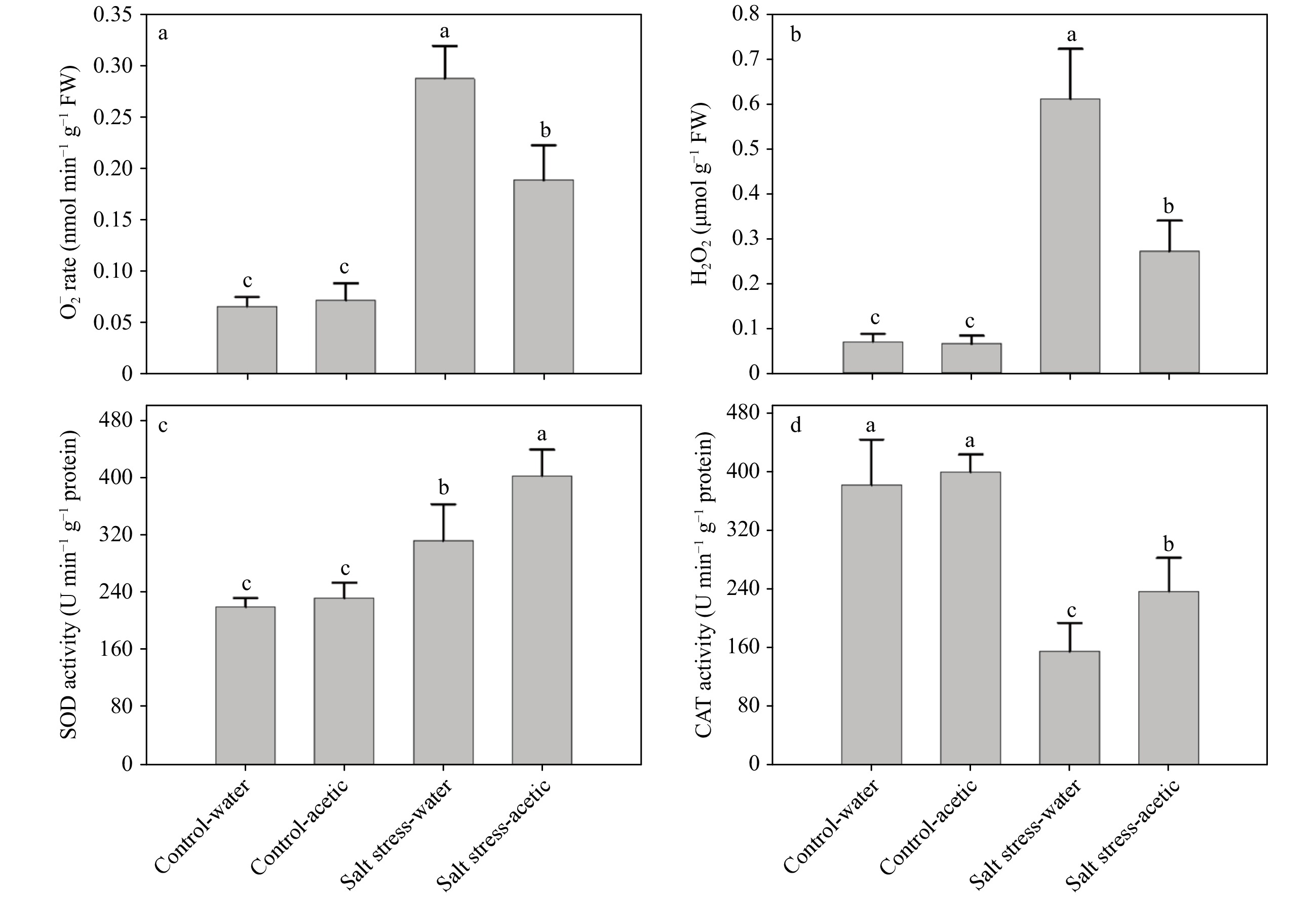
Figure 5. Effect of exogenous application of acetic acid on
${\rm O}_2^- $ production rate, and H2O2 content, and enzymatic antioxidant activity of SOD and CAT in leaves under control and salt stress conditions. Data were collected at 21 d of treatment. Bars labeled with different letters indicate significant differences among the treatments at a given day based on Fisher’s protected LSD test at P ≤ 0.05 level (n = 4).Besides the effect on accelerated leaf senescence, salt stress also inhibited the shoot and root growth of perennial ryegrass (Fig. 2). Exogenous treatment of acetic acid resulted in 72.2% and 34.5% increases in shoot and root dry weight, respectively, compared to that of water control at 28 d of salt stress (Fig. 2b). While, under the non-stress condition, acetic acid had no significant effects on the growth of ryegrass after 28 d of treatment (Fig. 2).
Both salt stress and application of exogenous acetic acid lead to increased endogenous acetic acid content and upregulated expression of acetic acid biosynthetic genes
-
Without exogenous acetic acid or salt stress treatments, the endogenous acetic acid content in ryegrass leaves was maintained at a constant level of ~260 μg/g dry weight (DW) during the 28 d of the experiment (Fig. 3a). Under salt stress, the endogenous acetic acid level steadily increased to 901.4 μg/g·DW by 28 d of treatment (Fig. 3a). Exogenous application of acetic acid led to significant increases in endogenous content of acetic acid content prior to plants exposure and during salt stress. The endogenous acetic acid level gradually reduced to the level of 383.3 μg/g·DW by 28 d of exogenous acetic acid treatment under non-stress condition, while the acetic acid-treated plants under salt stress condition maintained a high level of acetic acid content of ~1150 μg/g·DW (Fig. 3a).
Relative expression levels of two step-limiting acetic acid biosynthesis genes, LpPDC1 and LpALDH2B7, were further analyzed. As shown in Fig. 3b, expression of LpPDC1, but not LpALDH2B7, was significantly affected by salt stress and acetic acid treatment. Specifically, under the control condition, acetic acid treatment caused a transiently increased expression of LpPDC1 at 7 d. Under salt stress conditions, the transcription of LpPDC1 steadily increased, and those treated with acetic acid increased to its peak level at 21 d that was earlier than those with water control (Fig. 3b).
Acetic acid affected K+ content, and K+/Na+ ratio in shoot but not in root of ryegrass under salt stress
-
Without acetic acid treatment, salt stress caused 2.8 and 1.3-fold increases of Na+ content in ryegrass leaves and roots, respectively, but resulted in a 56.9% decrease in K+ content in leaves but no change in roots (Fig. 4). Acetic acid treatment alleviated the effect of salt stress on the plants Na+/K+ content. Specifically, under salt stress condition, acetic acid treated plants had significantly higher K+ but lower Na+ content in leaves than those treated with water, and thereby a significantly higher K+/Na+ ratio (Fig. 4).
At the gene expression level, pretreatment with acetic acid caused an increased expression level of LpHKT1;1, a Na+ specific transporter or Na+/K+ co-transporter gene. Salt stress also increased the expression of LpHKT1;1 at 14 and 21 d after the stress. Yet, plants treated with acetic acid exhibited 52.7% and 48.4% higher expression levels of LpHKT1;1 than those control plants under salt stress alone at 14 and 21 d, respectively (Fig. 4d).
Acetic acid alleviated salt-induced ROS accumulation and oxidative damage
-
Without acetic acid pre-treatment, salt stress caused increased production of
${\rm O}_2^- $ by 3.32-fold and the H2O2 content by 7.43-fold in leaves after 21 d of treatment (Fig. 5a and b). Comparatively, acetic acid treated plants displayed 1.81-fold decreased${\rm O}_2^- $ production rate, and 1.64-fold decreased H2O2 content (Fig. 5a and b). Nevertheless, the${\rm O}_2^- $ production rate and the H2O2 content of plants treated with and without acetic acid were at similar levels when grown under non-salt conditions (Fig. 5a and b).In order to evaluate whether acetic acid induced reduction of ROS accumulation was associated with the activation of ROS scavenging enzymes, the enzymatic antioxidant activity were further examined. Regardless of pre-treatment with or without acetic acid, salt stress caused increased SOD, but decreased CAT activities in ryegrass (Fig. 5c and d). Yet, it was notable that pre-treatment with acetic acid significantly increased the activities of both SOD and CAT by 1.29- and 1.53-fold than that of the water control under salt stress conditions; while no difference was found between them when grown under non-salt conditions (Fig. 5c and d).
Acetic acid and salt stress affected contents of JA, ABA, IAA and CKs in ryegrass
-
To examine whether the effect of acetic acid on regulation of salt-induced leaf senescence was associated with its effects on hormone metabolism, we measured the phytohormone content of ryegrass leaves at 21 d after salt stress, including jasmonic acid (JA, JA-Ile, cis-OPDA), ABA, auxin (IAA), salicylic acid (SA), ethylene biosynthesis precursor (ACC), and five forms of CKs (tZ, cZ, tZR, cZR, and iP). As shown in Fig. 6, salt stress resulted in significantly increased content of JA, JA-Ile, ABA, and ACC, but decreased IAA, SA, tZ, cZ, cZR, and iP content compared to those under control conditions. Under non-stress conditions, acetic acid treatment only caused significantly higher levels of JA and JA-Ile by 155.35% and 270.76%, respectively, than those treated with water. Under salt stress conditions, acetic acid-treated plants showed significantly higher content of JA, JA-ILe, cis-OPDA, IAA, tZ, cZR, and iP, but significantly lower ABA content than those of the control (Fig. 6).
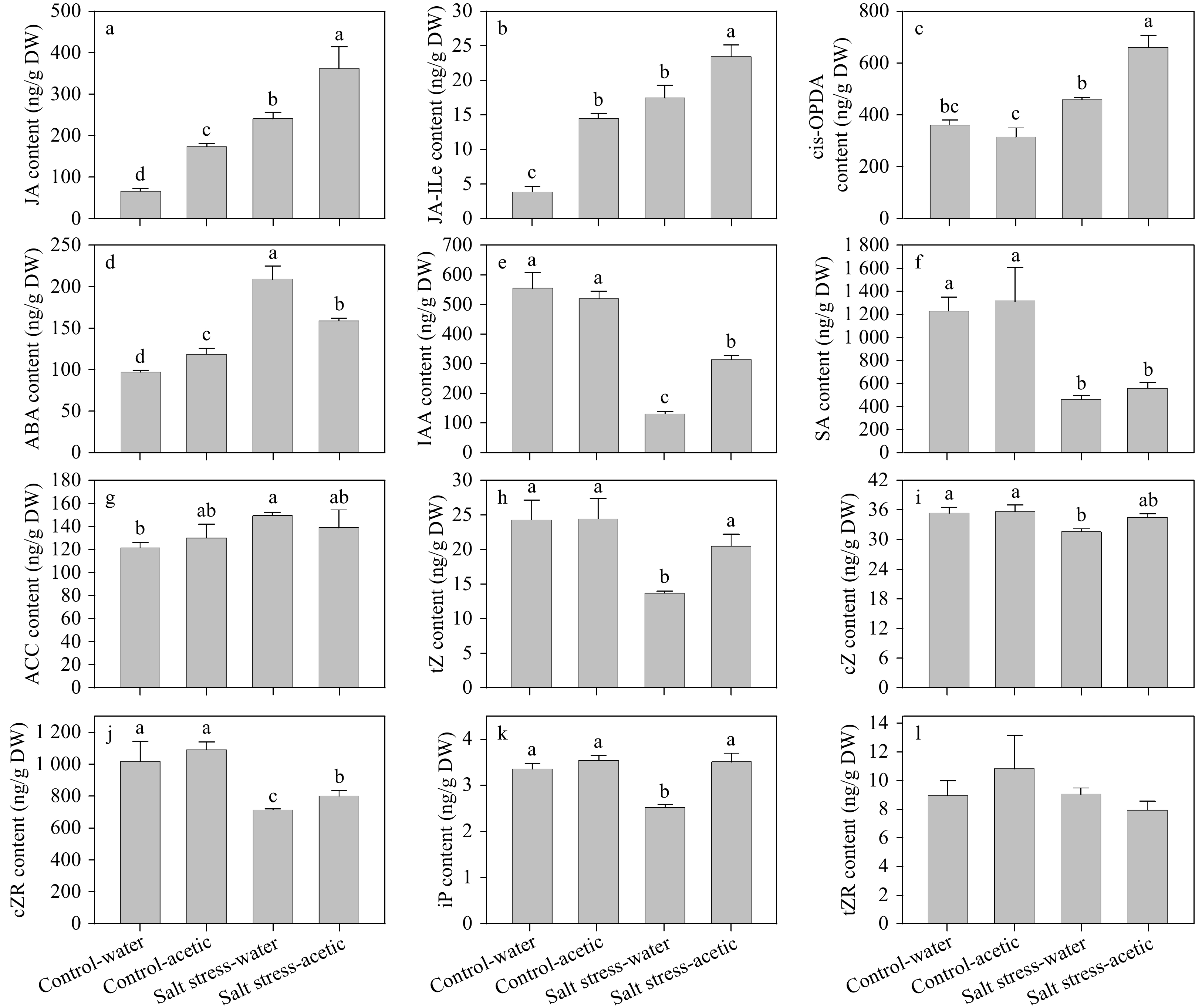
Figure 6. Effect of exogenous application of acetic acid on endogenous phytohormones, including three forms of jasmonate (JA, JA-ILe, and cis-OPDA), ABA, IAA, SA, ACC (an ethylene precursor), and five forms of cytokinins (tZ, cZ, tZR, cZR, and iP) in perennial ryegrass leaves under control and salt stress conditions. Bars labeled with different letters indicate significant differences among the treatments based on Fisher’s protected LSD test at P ≤ 0.05 level (n = 4).
The effects of salt stress and acetic acid and their interaction on endogenous acetic acid, iron, and hormone contents were further analyzed. As shown in Supplementary Table 1, the effects of salt stress and acetic acid reached significant levels (P ≤ 0.05) on the content of endogenous acetic acid, Na+ and K+, and several hormones, including JA, JA-Ile, cis-OPDA, ABA, IAA, iP, and cZR. Furthermore, content of endogenous acetic acid was found to correlate with the content of K+ and several hormones (e.g. JA, JA-ILe, cis-OPDA, IAA, tZ, cZR, and iP) in a positive way, but correlated with Na+ and ABA in a negative way (Supplementary Table 2) at 21 d of salt stress treatment. Na+ content was negatively correlated with JA, JA-ILe, cis-OPDA, IAA, tZ, cZR, and iP, but positively correlated with ABA. Unlike Na+, K+ content was found positively correlated with content of JA, JA-ILe, cis-OPDA, IAA, tZ, cZR, and iP, but negatively correlated with ABA (Supplementary Table 2).
Acetic acid and salt stress altered the expression of key genes involved in metabolism and signaling pathways of JA, CK, IAA and ABA
-
Relative expression of several key genes involved in metabolism and signaling of JA, CK, ABA, and IAA were further analyzed. As shown in Fig. 7, the transcriptions of JAR, a JA biosynthesis gene, and MYC2, a transcription factor in the JA signaling pathway, were both significantly down-regulated by salt stress. Furthermore, the relative expression levels of JAR and MYC2 were both up-regulated by acetic acid treatment under both salt stress and non-stress conditions (Fig. 7). Expression of CK biosynthesis genes, including IPT2 and LOG1, were significantly up-regulated by acetic acid under salt stress conditions, but were not affected by acetic acid under the optimum control conditions (Fig. 7). Expression of ARR1 and ARR10, two CK signaling genes, were also up-regulated by acetic acid treatment under salt stress conditions (Fig. 7).

Figure 7. Heatmap of the transcription of metabolic and signaling genes involved in JA, CK, IAA, and ABA in perennial ryegrass under control and salt stress conditions.
The relative expression of TAA1, an IAA biosynthesis gene, was increased by acetic acid treatment under salt stress conditions, as well as ARF1 and ARF2, two IAA signaling pathways genes (Fig. 7). The expression of ABA biosynthesis genes ZEP was activated by salt stress, but was suppressed by acetic acid under both salt stress and control conditions (Fig. 7). Salt stress also activated the transcription of two ABA signaling pathway genes, ABI3 and ABI5. While, exogenous application of acetic acid decreased their expression levels under both salt and non-stress conditions at 21 and 28 d (Fig. 7).
-
Exogenous application of acetic acid led to increased endogenous content of this organic acid in perennial ryegrass, which was associated with increased expression level of LpPDC1. The elevated acetic acid content and up-regulation of the synthesis gene could be attributed to the positive effects of promoting perennial ryegrass tolerance to salt stress via regulation of several processes, including K+ and Na+ homeostasis, hormone metabolism, and antioxidant metabolism, as discussed below and illustrated in Fig. 8.
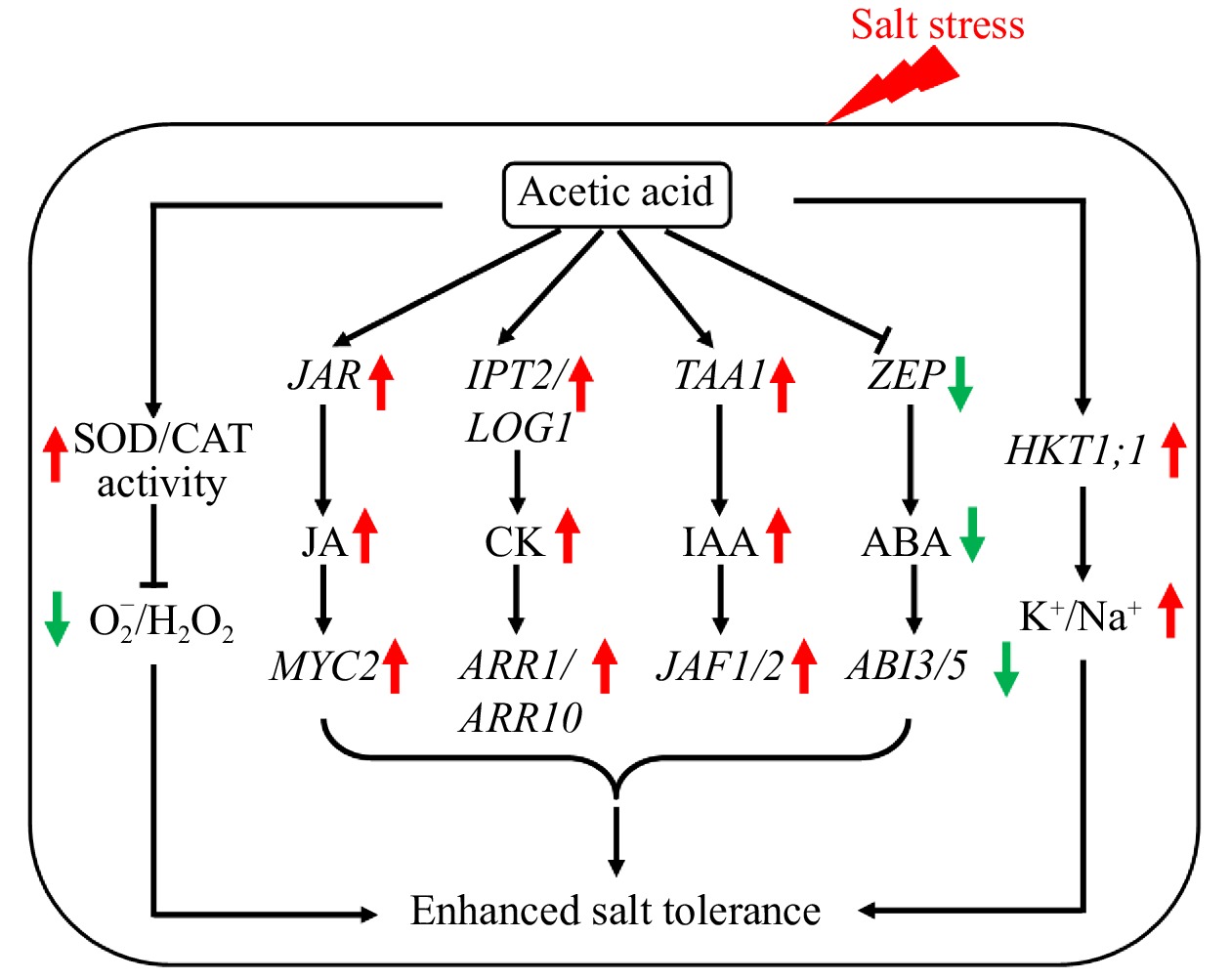
Figure 8. Proposed model for the regulation of salt tolerance by acetic acid in perennial ryegrass. Acetic acid affected multifaceted pathways to improve plant salt tolerance involving ionic transportation, ROS balance, and JA, CK, IAA, and ABA biosynthesis and signal transduction.
Firstly, it is important to note that both exogenous application of acetic acid and salt stress caused increased endogenous acetic acid content in ryegrass (Fig. 3). Since acetic acid is a water-soluble small molecule, it should be up-taken by roots and transported to leaves. When ryegrass was under control conditions, the endogenous content of acetic acid gradually declined. However, for those under the salt stress conditions, the endogenous acetic acid content gradually increased along with increased expression levels of the key biosynthetic gene LpPDC1 in ryegrass. This data suggested that increased biosynthesis of acetic acid was a natural endogenous response to salt stress in ryegrass which may be important for the plants’ adjustment to salt stress.
Secondly, acetic acid application affected the maintenance of K+ and Na+ homeostasis in leaves, one crucial mechanism for plant salt tolerance[16]. Ion homeostasis during salt stress requires the maintenance of constant K+ acquisition and distribution to balance the toxic effects of Na+ accumulation[20]. Over-accumulation of Na+ could disrupt K+ uptake by root cells[21]. Therefore, increasing innate K+ content or the K+/Na+ ratio was a common strategy to alleviate salt stress. Application of some organic molecules (e.g. choline) can increase K+ content and the K+/Na+ ratio in plants under salt stress[22]. In the current study, we found that salt-stressed plants treated with acetic acid maintained higher K+ but lower Na+ contents in leaves and thus a better status of Na+/K+ homeostasis than the water-treated perennial ryegrass plants. It had been reported that HKT, serves as a Na+ specific transporter or Na+/K+ cotransporter, and plays an important role in plant tolerance to salt stress[23]. Rice oshkt1;1 mutant accumulated higher concentration Na+ in leaf blades and exhibited hypersensitivity to salt stress[24]. Overexpressing HKT in durum wheat increased the grain yield by 25% compared with the wild type when grown in saline soil[25]. Our study found that the transcription of K+/Na+ transporters, HKT1;1, was up-regulated by acetic acid under salt stress conditions in leaves of perennial ryegrass. We also noted that acetic acid treatment has little effect on the K+/Na+ homeostasis in roots. We hypothesized that acetic acid affected K+/Na+ transporter gene expression or protein acetylation in a tissue-specific manner. Yet, the exact reason underlying this phenomenon requires further research.
A previous report on the effect of acetate on plant drought tolerance has emphasized the key role of JA that drought triggered metabolic flux conversion from glycolysis into acetate synthesis to stimulate the JA biosynthesis and signaling pathway, and the activated JA signaling pathway further improved plant drought tolerance[6]. Kim et al. (2017) found exogenous acetic acid promotes de novo JA, but not ABA synthesis[6]. While, Hossain et al. (2020) found that acetic acid treatment enhanced drought tolerance by upregulating ABA signaling pathway-related genes in cassava[12]. In the current study, we also found that, upon acetic acid treatment, the content of JAs, including JA, JA-ILe, and cis-OPDA, was significantly increased compared to water control under salt stress conditions. Our results indicated that acetic acid alleviated the salt caused damage associated with ABA metabolism, which was consistent with the results of Hossain et al. (2020)[12], while inconsistent with that of Kim et al. (2017)[6]. Auxin such as IAA can promote root initiation and also delay stress-induced senescence[26]. At the root tip, salt decreases meristem size and primary root elongation, partly through a reduction in local auxin levels[27]. In our study, acetic acid treatment increased IAA content under salt stress conditions. Additionally, the relative expression levels of an auxin biosynthesis gene, TAA1, and two auxin signaling pathway genes, ARF1 and ARF2, were increased by acetic acid treatment under salt stress conditions. These results indicate that acetic acid conferred salt treatment was associated with IAA metabolic pathways in perennial ryegrass. Cytokinins also play a role in mitigating salt stress. For example, transgenic creeping bentgrass and apple plants with higher endogenous CK level by overexpressing a key CK synthesis gene ipt also had significantly increased salt tolerance[28]. Exogenous CK (6-BA) treatment also delayed salt-induced leaf senescence in perennial ryegrass[17]. In the current study, content of three forms of CKs, including tZ, cZR, and iP, with the exception of cZ and tZR, were significant increased by acetic acid treatment under salt stress conditions. In addition to the above-mentioned hormones, ethylene has long been regarded as a stress-related hormone. Ethylene was found to increase salt tolerance in Arabidopsis by maintaining Na+/K+ homeostasis in shoots[29]. Transgenic plants with over-expression of a constitutively ethylene signaling activated ctr1-1 or EIN3 gene exhibited increased salt tolerance[30]. Our results found that salt stress significantly increased the content of ACC, an ethylene precursor, while, acetic acid treatment has no effects on its level under both salt stress and non-stress conditions, indicating acetic acid in alleviation of salt stress was not associated with ethylene biosynthesis in perennial ryegrass.
Salt stress damages cell membrane integrity, as indicated by the increased levels of EL and MDA, and could be as a result of excessive accumulation of
${\rm O}_2^- $ and H2O2[2]. Strategies which suppress over-accumulation of ROS may be effective in suppressing salinity-caused membrane damage. Several recent studies reported that exogenous application of acetic acid suppressed ROS accumulations and protected membrane integrity under salt and drought stress conditions[2, 31]. In the present study, we also found acetic acid treatment significantly decreased the endogenous${\rm O}_2^- $ production rate and H2O2 level during the salt stress treatment in perennial ryegrass. Previous studies suggested that acetic acid may reduce stress-induced oxidative damage by activating ROS scavenging systems. For example, Rahman et al. (2020) found that exogenous acetic acid treatment reduced drought-induced ROS accumulation due to the enhancement of antioxidant activities, such as SOD and CAT[31]. Results of Rahman et al. (2019) indicated that acetic acid treatment suppressed salt-induced accumulation of ROS, which was attributed to the activation of CAT activity, but not SOD[2]. The results in our study demonstrated that acetic acid treatment significantly increased the activity of both SOD and CAT under salt stress conditions in perennial ryegrass. Our results strongly suggest that acetic acid suppressed salt-induced oxidative damage may by regulating the SOD-CAT antioxidant pathway in perennial grass species.From the perspective of application, improved grass salt tolerance is the key for turf establishment in saline soil even if it is a temporary effect by watering or spraying with an exogenous substance. Application of acetic acid or acetic salt at low concentrations is an affordable solution, therefore further research should be undertaken to perform field studies on turf establishment and management in saline soils.
-
This study found that acetic acid mitigated salt stress, or promoted plant tolerance to salt stress, involving alleviation of ionic and ROS toxicity and regulation of synthesis and signaling of auxin, CK, ABA, and JA in perennial ryegrass (Fig. 8). Further studies are required to understand the direct targets of acetic acid in plants, and how crosstalk occurs between acetic acid and phytohormones at levels of metabolism and signaling pathways. Elevating acetic acid content through chemical priming or genetic modification of acetic acid synthesis genes is a promising approach to improve tolerance to salt stress.
-
Seeds of perennial ryegrass (cv. ‘Pinnacle’) were sown in plastic pots (14.5 cm in diameter and 13 cm in height) filled with fritted clay and maintained in a growth chamber for 45 d. The growth chamber was controlled at photoperiod of 16 h (25/20 °C, day/night), 70% relative humidity, and photosynthetic active radiation (PAR) of 750 µmol m−2 s−1. Prior to treatment, plants in each pot were trimmed weekly to maintain a height of 12 cm and fertilized once a week with Hoagland's nutrient solution[32].
The method for actic acid treatment was selected according to a study reported by Kim et al. (2017)[6]. In detail, 100 ml acetic acid (20 mM) was irrigated to the soil in each pot daily for three days. The same amount of water with its pH value adjusted with hydrochloric acid (HCl) to the same with acetic acid solution was used as the control. After removing the treatment solutions by capillarity using a paper towel at the bottom of each pot, plants were maintained in a growth chamber for another five days and then subjected to salt stress treatment. For salt stress treatment, plants were watered with 200 ml of 80, 160, and 240 mM NaCl for each pot for the first three days, respectively, and then irrigated with 200 ml of 250 mM NaCl every two days for 25 d. The control for salt stress treatment was irrigated with the same volume of water instead of salt solution.
Physiological parameters for the evaluation of salt tolerance
-
Stress indicators, including Fv/Fm, TQ, EL, and Chl content were measured at 0, 7, 14, 21, and 28 d after salt stress. TQ was visually rated using a 1-9 scale to evaluate the overall plant performance based on shoot density, plant color, and uniformity[33]. For quantification of Chl content, about 0.1 g of leaves were soaked in 15 ml dimethyl sulfoxide (DMSO) and kept under dark conditions for at least 72 h, subsequently the absorbance of the extraction solution was measured using a spectrophotometer (Biochrom Ltd, Cambridge, England) at wavelengths of 645 nm and 663 nm[34]. Leaf Fv/Fm was measured on leaves after 30 min dark-adaption using a fluorescence meter reported previously[13].
Relative EL was calculated as (Ci/Cmax) ×100 described previously by Blum and Ebercon (1981)[35]. The initial conductivity (Ci) and the maximum conductivity (Cmax) of leaves were measured using a conductivity meter (Thermo Scientific, Baverly, USA). After 28 d of treatment, roots and shoots were harvested and dried in an oven at 60 °C, the shoot and root dry weight were then measured separately.
Analysis of Na+ and K+ content,
${\bf O}_{\bf 2}^{ \large{\bf{–}}}$ and H2O2 level, and enzymatic antioxidant activity -
Content of Na+ and K+ in shoot and root was measured using an inductively coupled plasma atomic emission spectrometer according to the method described by Gao et al. (2020)[22]. The
${\rm O}_2^- $ production rate was quantified using the method reported by Elstner and Heupel (1976)[36]. H2O2 content was measured according to the method described by Velikova et al. (2000)[37]. The activity of SOD and CAT were quantified following the methods of Meloni et al. (2003)[38] and Chance and Maehly (1955)[39], respectively. The details for the measurement of these four traits in perennial ryegrass have been published in our previous study[40].Quantification of endogenous acetic acid content
-
Acetic acid content was quantified using gas chromatography–mass spectrometry (GC–MS) according to the method reported by Kim et al. (2017)[6]. In brief, about 0.5 g of leaves were sampled after 0, 7, 14, 21, and 28 d of treatment, ground into powder in liquid nitrogen, homogenized in 5 ml of cold distilled water, and centrifuged at 15,000 g for 1 min at 4 °C. The supernatant was then collected and diluted with an equal amount of acetone. Acetic acid in sample extracts were further analyzed using GC–MS system.
Quantification of phytohormone content
-
After 21 d of salt stress, phytohormone content was quantified in leaves according to the method reported by Redestig et al. (2009)[41] with minor modifications: Approximately 0.1 g leaf was collected, ground into fine powder in liquid nitrogen, homogenized in 1 ml cold acetonitrile solution in a 2 ml tube and maintained in the dark, 4 °C for 12 h. The extraction solution was then centrifuged at 14,000 g for 10 min and 800 μl supernatants were collected to a new 1.5 ml tube and dried with nitrogen gas. The dried extractions were dissolved using 100 μl 50% acetonitrile solution (v/v), and the supernatants were used for further analysis after centrifuge at 14,000 g for 20 m. Hormone content in the extraction solution were determined using an ultra-high-performance liquid chromatography (UPLC) system (WatersI-ClassLC, Waters, Milford, USA) using an ODS column (ACQUITYUPLCBEHC 18, 1.7 μm, 2.1 mm ×100 mm; Waters) based on the retention time (RT) and peak area compared to the internal control of each known hormone.
Gene relative expression level analysis
-
The total mRNA of leaves were extracted using Plant RNA Kit from OMEGA. First-strand cDNA Reverse Transcription Kit from Takara was used for the synthesis of cDNA. The qRT-PCR reaction was performed in a 20 μl reaction with power SYBR® Green PCR Master mix (Applied Biosystems) using the Roche LightCycler® 480 II Real-Time PCR System. All PCR reactions were performed with four biological replicates. LpelF4a was used as reference gene and the gene relative expression level was analyzed using the delta-delta-CT method[42]. The gene bank number and sequence of primers used in this study were listed in Supplementary Table 1.
Statistical analysis
-
Data from different treatments were analyzed by Fisher’s protected LSD test at P ≤ 0.05 level using SAS 9.3 software (SAS Institute Inc., Cary, NC, USA). The variance for the effects of salt and acetic acid treatment, and their interaction on iron and plant hormone contents, as shown in Supplementary Table 2, was performed using the software SPSS for Windows (Version 12, SPSS Inc., Chicago, IL). Pearson correlation analysis among the Na+ and K+ level and plant hormone content, as shown in Supplementary Table 3, was performed using the Bivariate Correlations program in software SPSS for Windows. The heatmap of the gene relative expression data in Fig. 3 and Fig. 7 were performed using R statistical software (R3.5.2 by R Development Core Team). Data are expressed as means ± standard error (SE) in Figs 1−6.
-
This study was supported by the Natural Science Foundation of Jiangsu Province (Grant No. BK20180546), by the discipline construction fund project of Gansu Agricultural University (Grant No. GAU-XKJS-2018-002), and by National Natural Science Foundation of China (Grant No. 31760701).
- The authors declare that they have no conflict of interest.
- Supplemental Table 1 Primers used in this study.
- Supplemental Table 2 Pearson correlation coefficients analysis among the iron and plant hormone contents with the data collected at 21 d under salt stress conditions.
- Supplemental Table 3 Summary of variance for the effects of salt treatment, acetic acid treatment, and the interaction between salt and acetic acid treatment on iron and plant hormone contents with the data collected at 21 d of treatment .
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang J, Zhang Q, Xing J, Li H, Miao J, et al. 2021. Acetic acid mitigated salt stress by alleviating ionic and oxidative damages and regulating hormone metabolism in perennial ryegrass ( Lolium perenne L.). Grass Research 1: 3 doi: 10.48130/GR-2021-0003

Acetic acid mitigated salt stress by alleviating ionic and oxidative damages and regulating hormone metabolism in perennial ryegrass (Lolium perenne L.)
- Received: 22 January 2021
- Accepted: 17 February 2021
- Published online: 11 March 2021
Abstract: Organic chemical priming is an effective strategy in mitigating salt stress to plants. The objective of this study was to determine effects and associated mechanisms of acetic acid regulating plant tolerance to salt stress. Perennial ryegrass plants were pre-treated with 20 mM acetic acid and subsequently subjected to salt stress for 28 days. Salt stress caused increased endogenous acetic acid content with up-regulated expression of its key biosynthetic gene LpPDC1. Application of acetic acid effectively alleviated salt caused damage in perennial ryegrass. Acetic acid treatment increased K+ content and suppressed Na+ accumulation to maintain a higher K+/Na+ ratio in leaves exposed to salt stress. Plants treated with acetic acid also had significantly lower levels of ${\rm O}_2^- $ and H2O2, but higher SOD and CAT activities than those of the control after 21 days of salt stress. Acetic acid treatment also altered the plants endogenous phytohormone content with higher content of jasmonate (including JA, JA-ILe, and cis-OPDA), auxin (IAA), and cytokinins (CK, such as tZ, cZR, and iP), but lower content of abscisic acid (ABA) under salt stress conditions. Furthermore, expression of genes involved in JA, IAA, and CK biosynthesis and signaling pathways were up-regulated, while those involved in ABA were down-regulated by acetic acid treatment under salt stress. The results demonstrate that acetic acid could mitigate salt stress in perennial ryegrass by regulating K+ and Na+ balance, promoting ROS scavenging, and activating stress-protection hormone synthesis and signaling.
-
Key words:
- Perennial ryegrass /
- salt stress /
- acetic acid /
- ionic balance /
- plant hormone