








-
Heat stress is a principal factor interfering with the development of cool-season plants, causing a myriad of negative effects on physiological and metabolic processes, including interruption of chlorophyll and amino acid metabolism that can lead to leaf senescence[1]. Heat-induced leaf senescence has been attributed to the acceleration of chlorophyll degradation due to increased activities of chlorophyll degrading enzymes, such as chlorophyllase (CHLASE), as reported in Arabidopsis (Arabidopsis thaliana)[2] and creeping bentgrass[3−5]. Leaf senescence can also be caused by the interruption of the assimilation of amino acids from inorganic nitrogen by the glutamine synthetase-glutamate synthase (GS-GOGAT) pathway, which is the key process controlling the balance between nitrogen and carbon in plant cells[6]. In the GS-GOGAT cycle, GS facilitates the entry of inorganic nitrogen into organic compounds, while GOGAT synthesizes glutamate from glutamine and
${\rm{NH}}_4^+ $ and recycles the nitrogen from glutamate for biosynthesis of other amino acids[7, 8]. In addition to the amino acids produced from remobilized nitrogen, Chl intermediates are also synthesized[9, 10]; thus, the GS-GOGAT pathway is crucial for the maintenance of Chl and amino acid metabolism during leaf senescence.Glutamate is a key component in the GS-GOGAT pathway, serving as an ammonia donor for all other amino acids and can also be catabolized into the non-protein amino acid,
$\gamma $ -aminobutyrate (GABA), which is known to accumulate under heat stress and is positively related to heat tolerance[11, 12]. Aspartate aminotransferase is a catalyst that mediates the transfer of an amino group to oxaloacetate from glutamic acid, forming aspartate, a direct precursor of methionine, threonine, isoleucine, and lysine, four amino acids in the oxaloacetate family[13, 14]. Furthermore, glutamate is a precursor to$\delta$ -aminolevulinic acid, serving as one of the first intermediates of Chl synthesis[15]. Glutamate is also involved in plant cell signaling, where it regulates the entry of Ca2+ into cells through ionotropic glutamate receptors (GLRs)[16]. Foliar application of glutamate in soybean (Glycine max L.) was shown to increase the enzymatic activity of catalase (CAT), an important antioxidant enzyme involved in the scavenging of free radicals[17]. Seed treatment of soybean with glutamate increased shoot biomass and yield under water deficit conditions[18]. Enhanced photosynthetic performance by glutamate was observed in tomato (Solanum lycopersicum) under chilling stress[19]. Only one study[20] was found related to the effects of glutamate on plant responses to heat stress, and it subjected 2-d-old maize (Zea mays L.) seedlings to 46 °C for 16 h; this study reported that seedling survival rate was increased by watering plants with glutamate solution, which could be associated with the involvement of glutamate receptor-like channel-mediated calcium signaling. Despite the reported positive effects of glutamate on plant responses to abiotic stress by a limited number of studies, the mechanisms of how glutamate may affect leaf senescence under prolonged periods of heat stress and its roles in modulating amino acid and/or Chl metabolism are not yet understood. By determining the manners in which glutamate affects heat-induced leaf senescence, it will be possible to establish how the compound may be used to control heat-induced leaf senescence in cool-season plants and may lead to the discovery of heat tolerance traits.The objectives of the current study included examining whether glutamate may be effective in suppressing heat-induced leaf senescence and to determine whether the effects of glutamate could be associated with the regulation of Chl and amino acid metabolism in creeping bentgrass. In order to examine this, glutamate was foliar-applied to creeping bentgrass plants maintained at non-stress or heat stress temperatures, and analysis of physiological characteristics associated with leaf senescence were examined, as well as the response of Chl-synthesizing and -degrading enzymes and the content of endogenous amino acids.
-
Turf quality (TQ) ratings of plants maintained under non-stress conditions remained above a score of 6.0 in plants treated with glutamate but decreased below 6.0 in untreated controls by 35 d (Fig. 1a). Through the extent of heat stress, there was significant deterioration in the TQ of untreated controls. Under optimal growth conditions, the TQ of glutamate-treated plants was evaluated to be significantly higher as compared with untreated controls at 14, 21, 28, and 35 d (12.9, 9.68, 11.67, and 13.79%, respectively). Additionally, TQ was significantly greater in glutamate-treated plants at 14, 21, 28, and 35 d (by 9.46, 14.29, 22.22, and 33.33%, respectively) of heat stress (Fig. 1b).
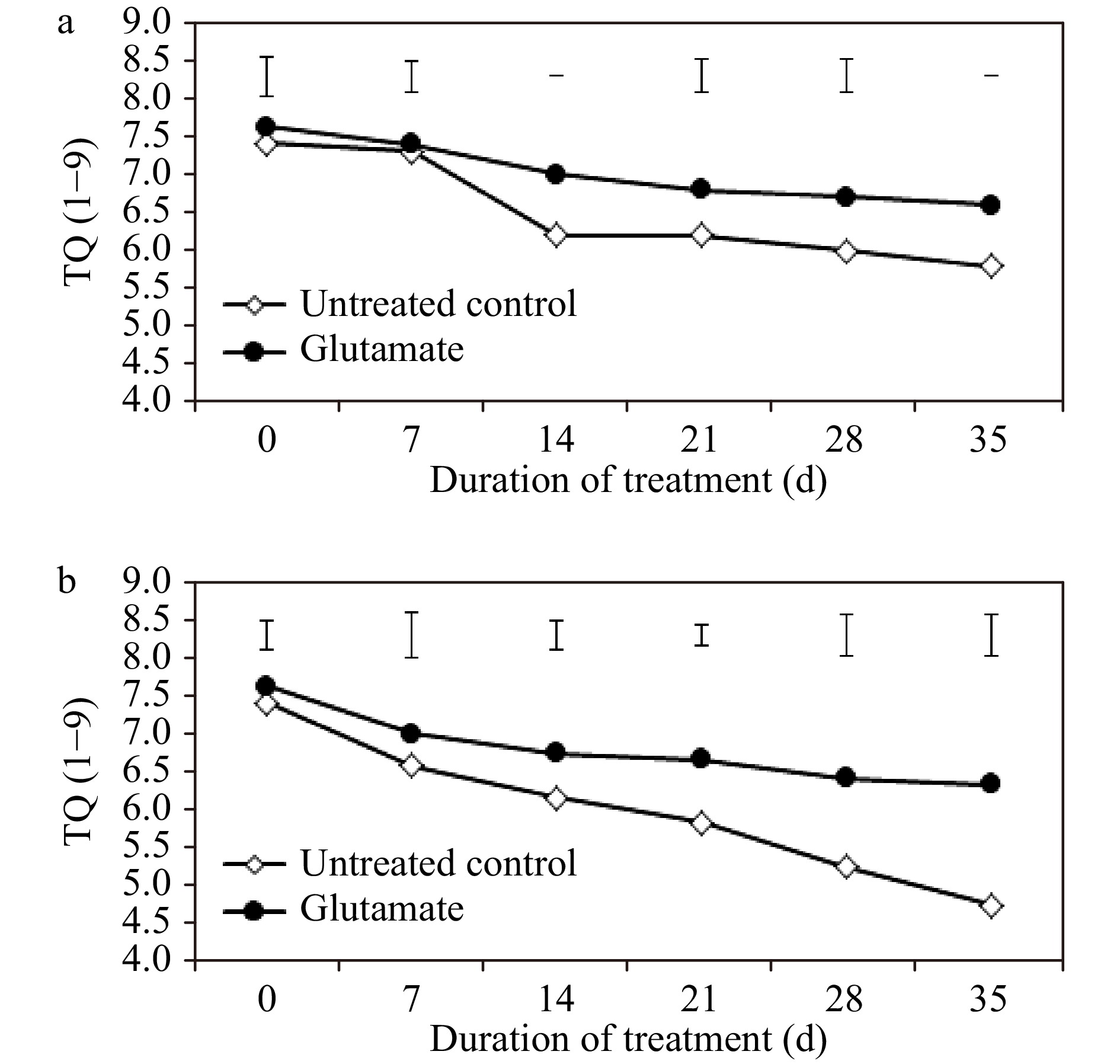
Figure 1. Numerical turf quality evaluations ranging from 1 through 9 for experimental plants treated with glutamate or untreated controls under non-stress temperature conditions (a) or heat stress (b). Error bars over each data point were used to separate means between non-stress and heat stress temperature conditions for each given date and are delineated by Fisher’s protected least significant difference (LSD) test applied at p < 0.05.
In glutamate-treated plants, photochemical efficiency (Fv/Fm) was significantly greater at 28 and 35 d (by 0.43 and 3.91%, respectively) under non-stress conditions. Fv/Fm depreciated significantly in both treatments from 7 d through 28 d of heat stress, but to a comparatively lesser extent in plants treated with glutamate (Fig. 2a). In comparison with untreated control plants, those treated with glutamate had significantly greater Fv/Fm values at 7, 14, 21, and 28 d (3.04, 2.93, 4.51, and 3.92%, respectively) of heat stress (Fig. 2b).
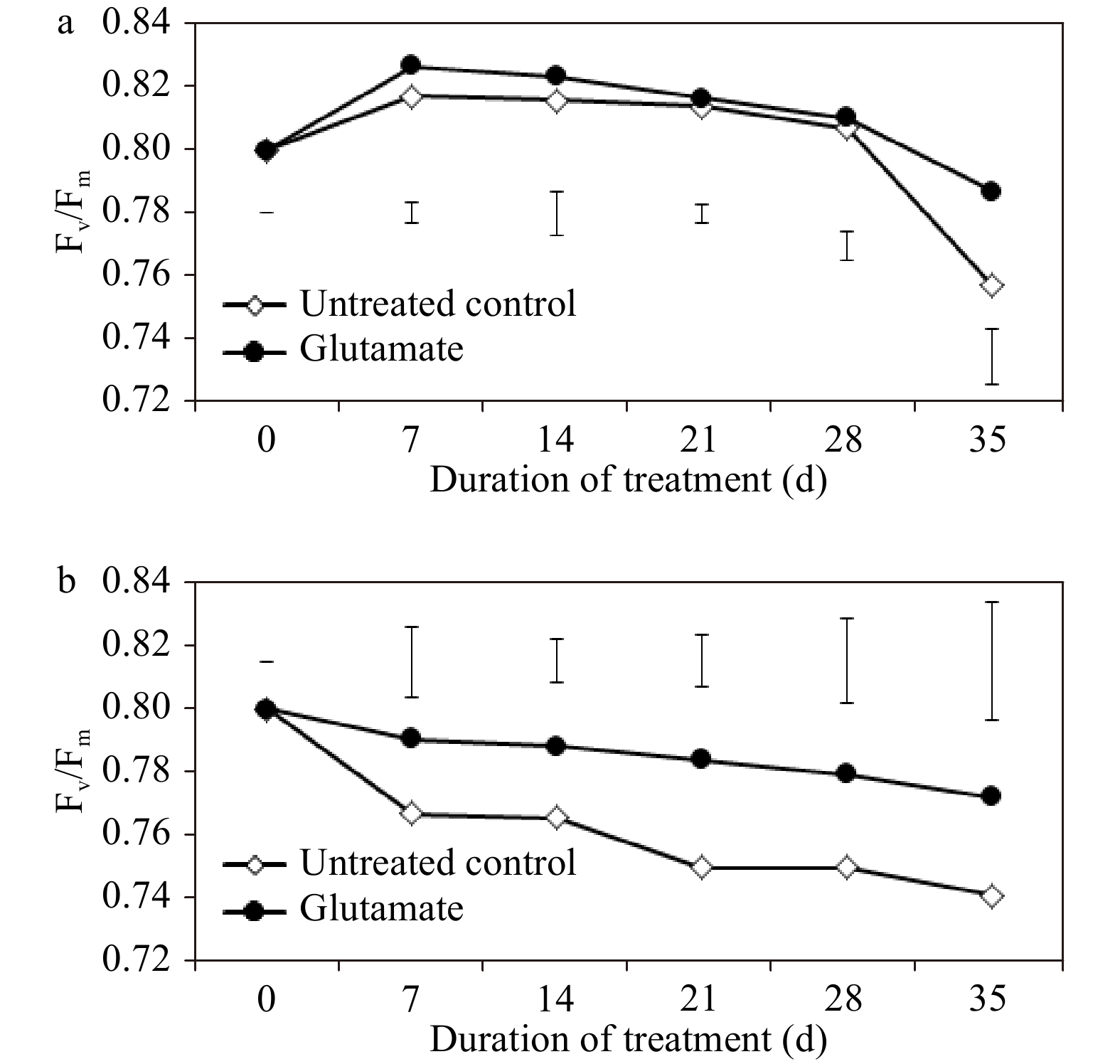
Figure 2. Photochemical efficiency for experimental plants treated with glutamate or untreated controls under non-stress temperature conditions (a) or heat stress (b). Error bars over each data point were used to separate means between non-stress and heat stress temperature conditions for each given date and are delineated by Fisher’s protected least significant difference (LSD) test applied at p < 0.05.
Through the period of non-stress conditions, leaf Chl content was significantly greater (12.6%) in plants treated with glutamate compared to untreated controls at 28 d (Fig. 3a). Chl levels significantly diminished regardless of treatment from 7 d through 35 d of heat stress, but the decline was more considerable in untreated controls. Through the duration of heat stress, glutamate significantly promoted Chl content at 7, 14, 21, 28, and 35 d (by 16.91, 23.9, 17.38, 22.32, and 40.39%, respectively) as compared to untreated control plants (Fig. 3b).
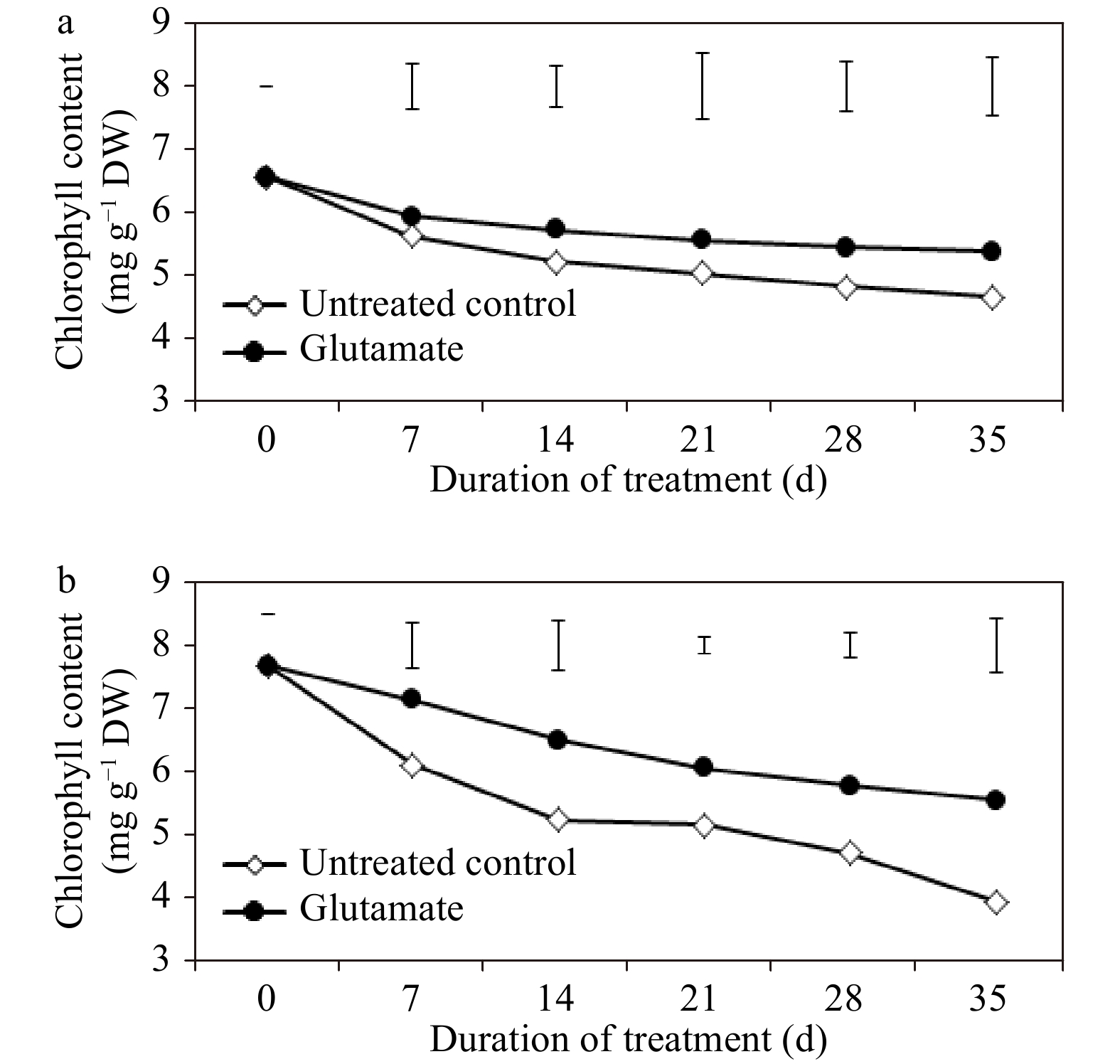
Figure 3. Chlorophyll content for experimental plants treated with glutamate or untreated controls under non-stress temperature conditions (a), or heat stress (b). Error bars over each data point were used to separate means between non-stress and heat stress temperature conditions for each given date and are delineated by Fisher’s protected least significant difference (LSD) test applied at p < 0.05.
Responses of Chl-synthesizing and –degrading enzymes to glutamate
-
To ascertain whether glutamate impacts Chl synthesis or degradation in relation to heat stress-induced leaf senescence, enzymatic activity of porphobilinogen deaminase (PBGD), a vital enzyme in Chl synthesis, and three Chl-degrading enzymes, chlorophyllase (CHLASE), pheophytinase (PPH), and chlorophyll-degrading peroxidase (CHL-PRX), were investigated.
PBGD activity lessened in glutamate-treated plants as well as untreated control plants through the entirety of the experimental period, but more appreciably under heat stress (Fig. 4a and b). Glutamate application had no significant impacts on PBGD activity through most of the experimental period under heat stress and non-stress conditions.
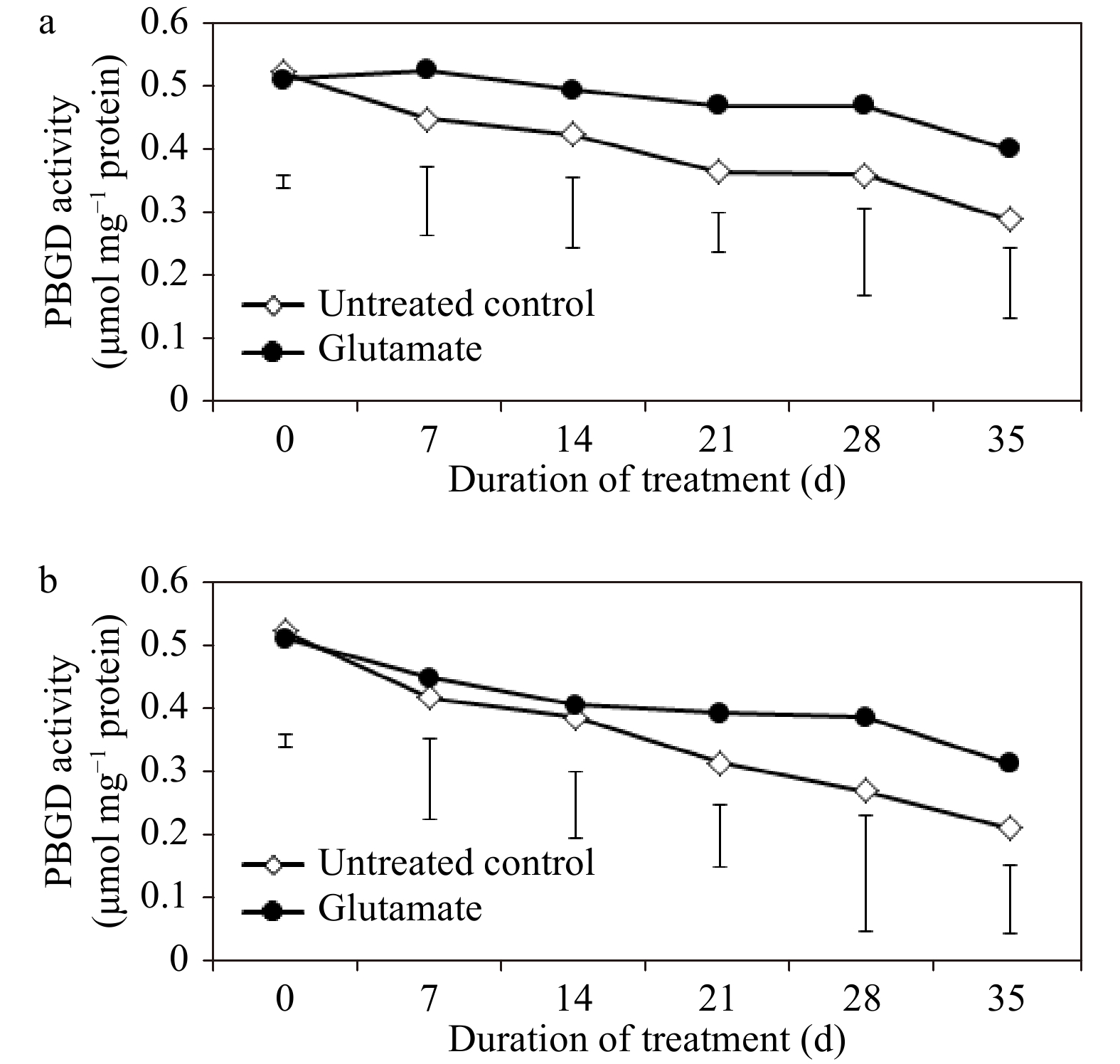
Figure 4. Porphobilinogen deaminase (PBGD) enzyme activity for experimental plants treated with glutamate or untreated controls under non-stress temperature conditions (a) or heat stress (b). Error bars over each data point were used to separate means between non-stress and heat stress temperature conditions for each given date and are delineated by Fisher’s protected least significant difference (LSD) test applied at p < 0.05.
CHLASE activity was pronounced overall through the duration of heat stress, especially in untreated control plants, when compared with non-stress controls (Fig. 5a and b). At 21 and 28 d of non-stress conditions, CHLASE activity was significantly greater in untreated control plants (by 30.11 and 36.94%, respectively) as compared to glutamate-treated plants, while there were no significant differences in PPH activity. Under heat stress, glutamate significantly reduced the activity of CHLASE at 14, 21, 28, and 35 d (by 35.9, 47.23, 46.83, and 41.64%, respectively) and that of PPH at 28 and 35 d (by 25.48 and 38.04%, respectively) (Fig. 6a and b).
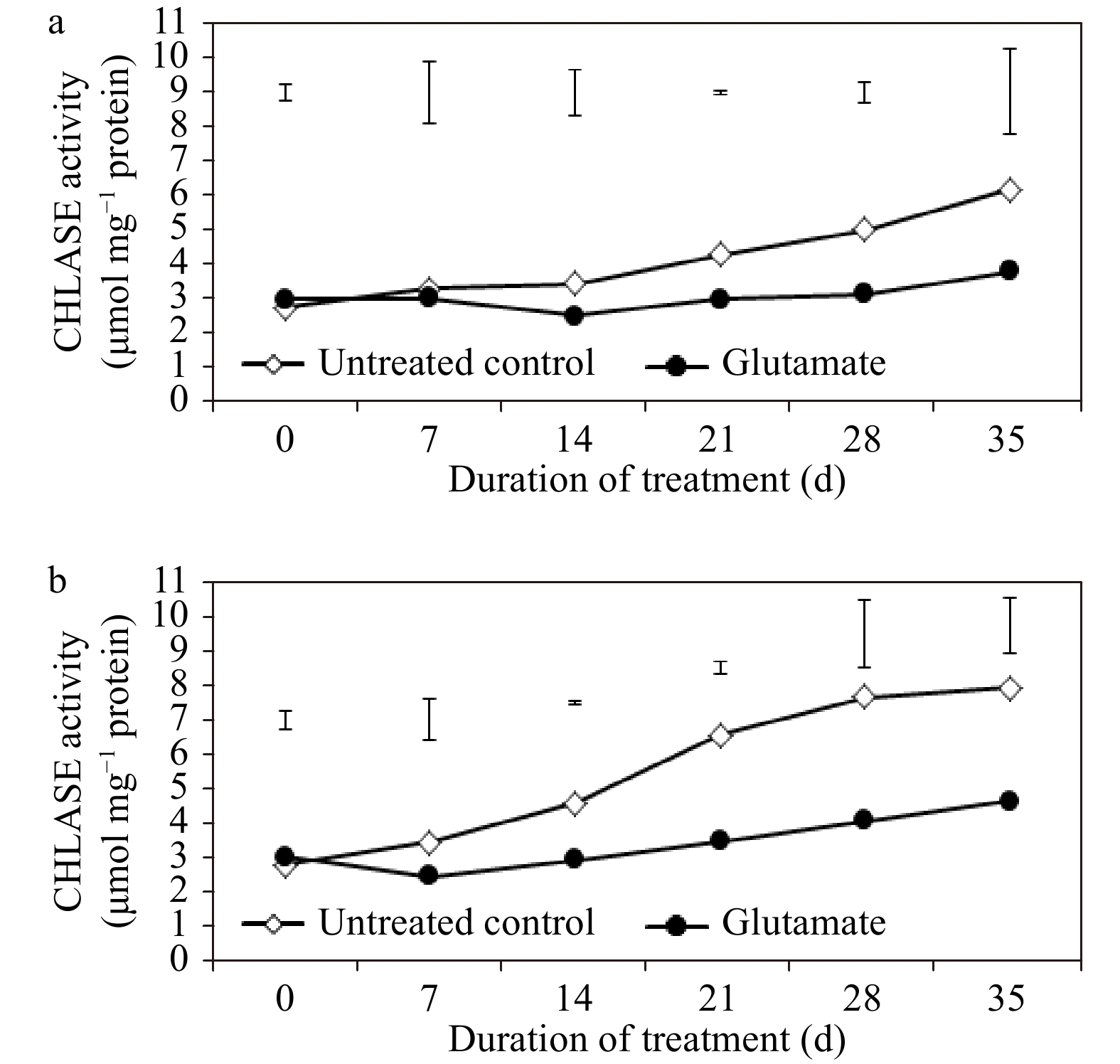
Figure 5. Chlorophyllase (CHLASE) enzyme activity for experimental plants treated with glutamate or untreated controls under non-stress temperature conditions (a) or heat stress (b). Error bars over each data point were used to separate means between non-stress and heat stress temperature conditions for each given date and are delineated by Fisher’s protected least significant difference (LSD) test applied at p < 0.05.
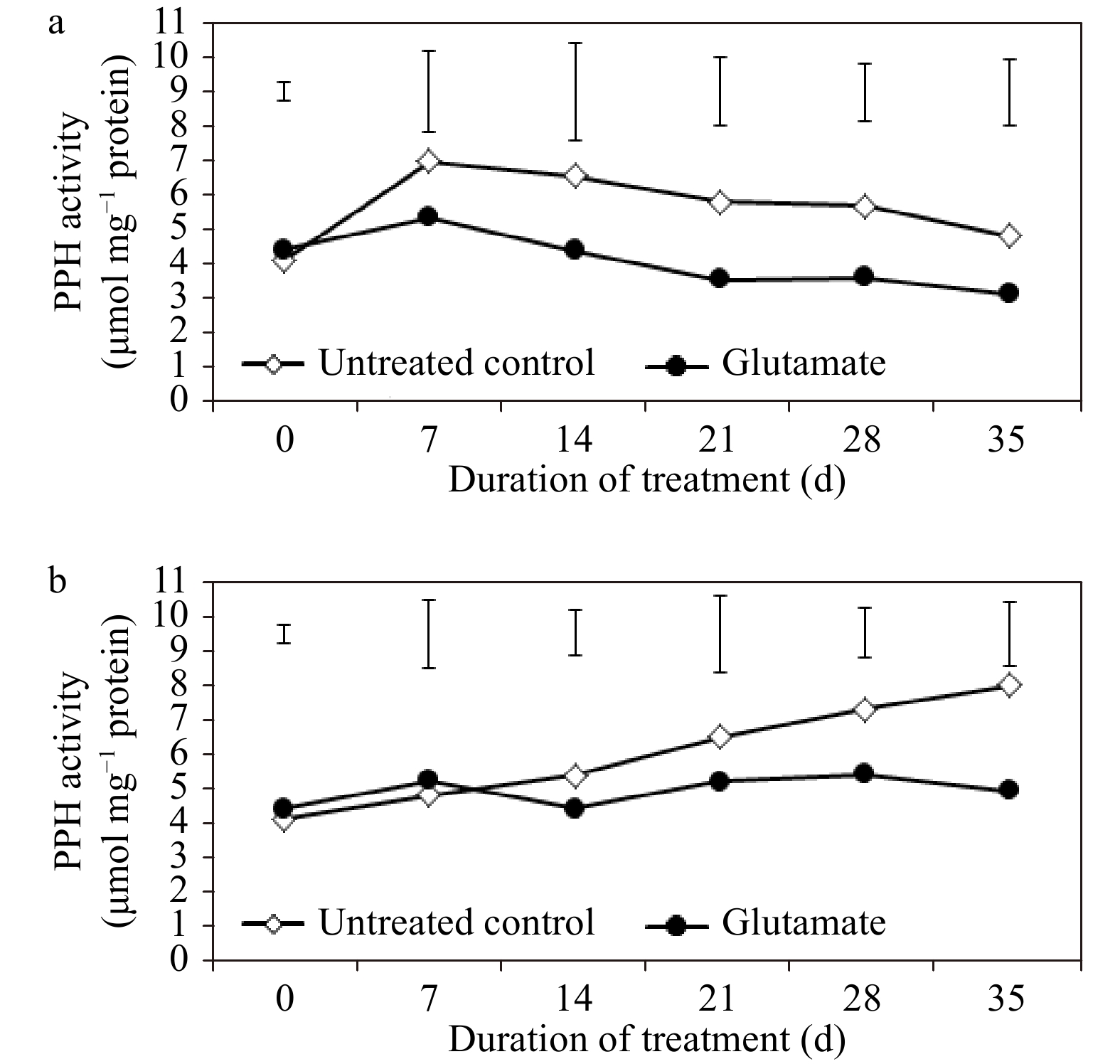
Figure 6. Pheophytinase (PPH) enzyme activity for experimental plants treated with glutamate or untreated controls under non-stress temperature conditions (a) or heat stress (b). Error bars over each data point were used to separate means between non-stress and heat stress temperature conditions for each given date and are delineated by Fisher’s protected least significant difference (LSD) test applied at p < 0.05.
CHL-PRX activity was not influenced by glutamate treatment for the majority of the experimental period under non-stress conditions (Fig. 7a). For untreated control and glutamate-treated plants, CHL-PRX enzyme activity rose through 35 d of heat stress, but only to a limited extent in those treated with glutamate (Fig. 7b). When compared with untreated controls, glutamate-treated plants had significantly less CHL-PRX activity at 14, 21, and 35 d of heat stress (by 27.84, 26.56, and 25.52%, respectively).
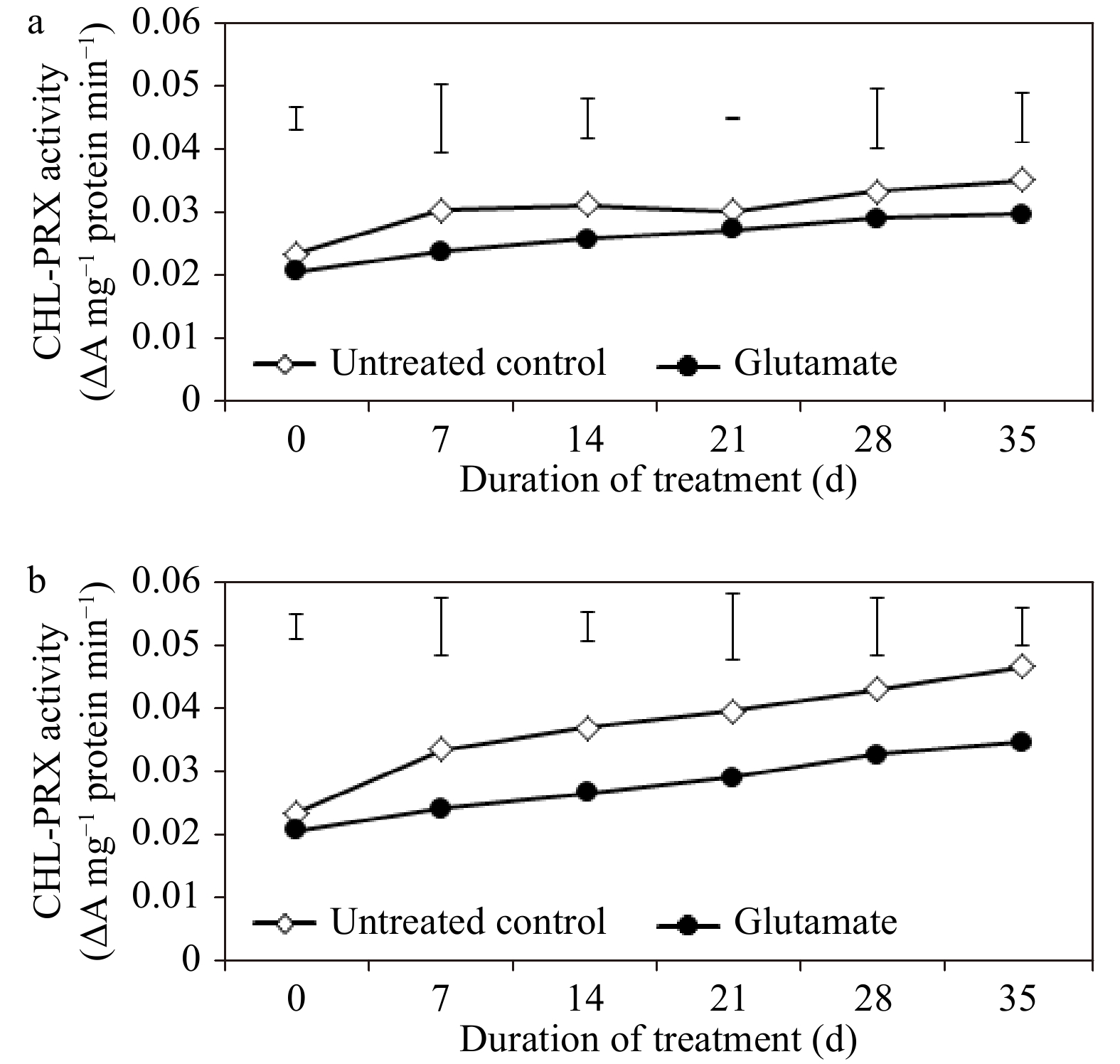
Figure 7. Chlorophyll-degrading peroxidase (CHL-PRX) enzyme activity for experimental plants treated with glutamate or untreated controls under non-stress temperature conditions (a) or heat stress (b). Error bars over each data point were used to separate means between non-stress and heat stress temperature conditions for each given date and are delineated by Fisher’s protected least significant difference (LSD) test applied at p < 0.05.
Changes in leaf amino acid content in response to glutamate
-
Under both non-stress and heat stress conditions, the majority of the amino acids examined exhibited an upward trend in their content in response to glutamate treatment compared to untreated control plants (Fig. 8); however, the magnitudes of change or level of significance in amino acid content in response to glutamate, varied according to temperature conditions and specific amino acid. Among all amino acids that were responsive to glutamate, isoleucine, histidine, asparagine, and methionine were up-regulated by glutamate only under heat stress while serine, threonine, lysine, aspartic acid, glutamic acid, and GABA exhibited significantly higher degrees of up-regulation in response to glutamate under heat stress (Fig. 9).

Figure 8. Heat map illustrating the general trend of fold-changes in the content of various amino acids inresponse to glutamate treatment in creeping bentgrass under non-stress or heat stress temperatureconditions. Each horizontal bar represents the fold-change in content for an individual amino acid inglutamate-treated plants relative to untreated plants.
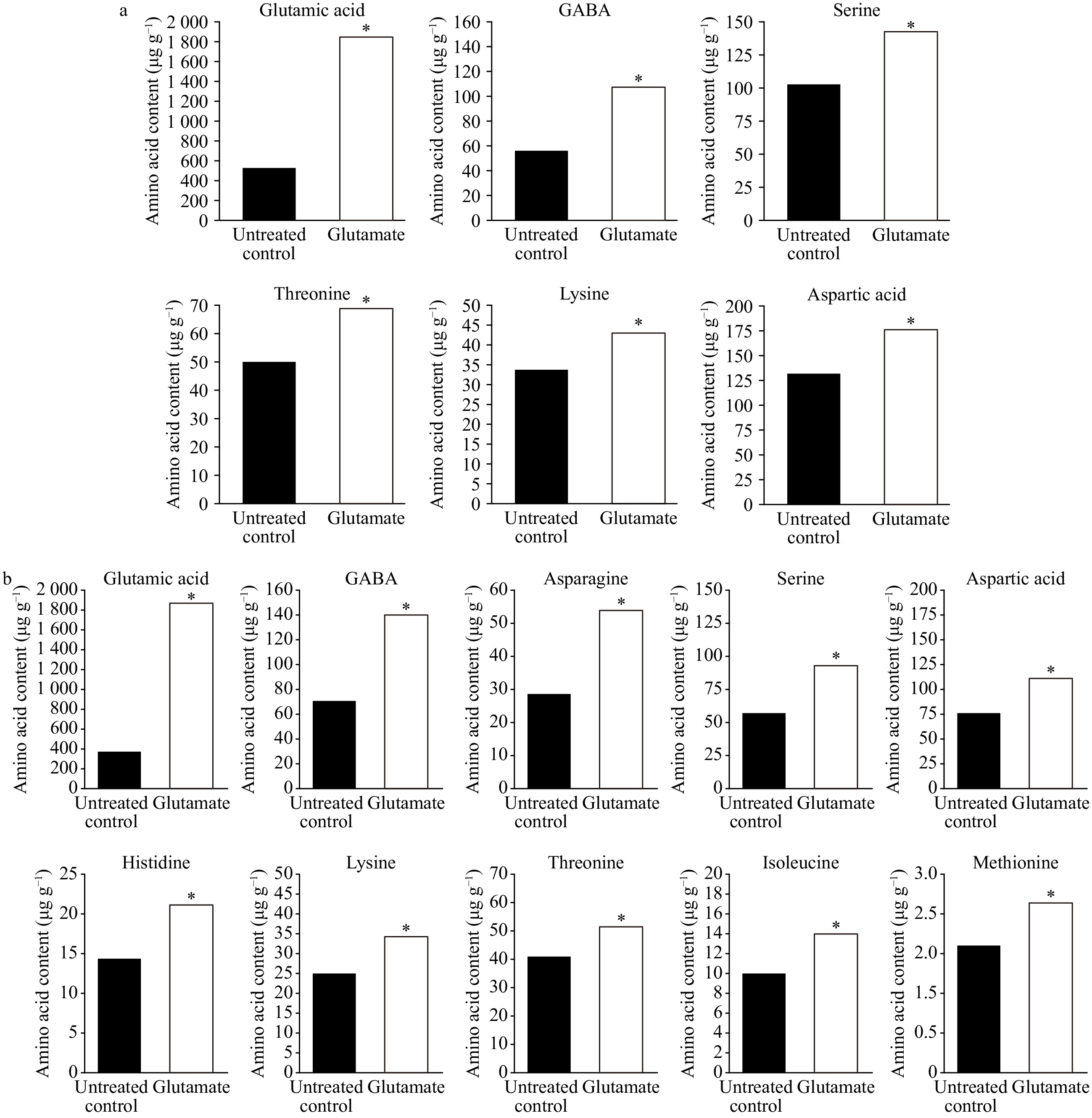
Figure 9. Content of amino acids significantly affected by glutamate for experimental plants under non-stress temperature conditions (a) or at 35 d heat stress (b). Asterisks over data bars represent significant differences between the glutamate treatment and untreated control and are delineated by Fisher’s protected least significant difference (LSD) test, applied at p < 0.05.
For untreated control plants exposed to non-stress conditions, application of glutamate significantly up-regulated the levels of glutamic acid, GABA, serine, threonine, lysine, and aspartic acid, which were higher by 254.16, 93.41, 38.73, 36.96, 28.06, and 35.62%, respectively, compared to untreated controls (Fig. 9a). In plants treated with glutamate under heat stress, content of glutamic acid, GABA, asparagine, serine, aspartic acid, histidine, isoleucine, lysine, methionine, and threonine was significantly enhanced by 391.4, 98.64, 88.98, 64.75, 48.4, 48.23, 41.3, 37.7, 32.59, 27.0%, respectively, compared to untreated controls (Fig. 9b).
-
Physiological analysis demonstrating the deterioration of TQ, Chl content, and photochemical efficiency under extended periods of heat stress indicated that high temperatures activated or expedited leaf senescence in creeping bentgrass, as reported in previous studies[3−5]. The ability of glutamate to alleviate the decline in all of those parameters is inextricably associated with leaf senescence, as glutamate-treated plants exhibited significant elevation of TQ, Chl content, and Fv/Fm, suggesting that glutamate could act as a repressor of heat-induced leaf senescence. Furthermore, this study revealed that glutamate-mediated regulation of leaf senescence for plants exposed to heat stress could be attributed to its effects on both amino acid and Chl metabolism, as discussed below.
Leaf Chl content, the hallmark indicator of leaf senescence, is regulated by a balance between synthesis and degradation processes; however, it was unclear prior to this study whether glutamate may control Chl decline via Chl synthesis or degradation under heat stress during leaf senescence. This study discovered that glutamate treatment led to a reduction in the activities of all three Chl-degrading enzymes and had more distinct effects on CHLASE and CHL-PRX but did not alter activity of the Chl-synthesizing enzyme, PBGD, in creeping bentgrass subjected to heat stress (Fig. 10). In a prior study, both CHLASE and CHL-PRX activity was heightened in creeping bentgrass exposed to heat stress, whereas activity of PPH did not increase significantly; furthermore, heat stress did not significantly modify activity of the Chl synthesis enzyme, PBGD[5]. The current findings imply that enhancement of Chl degradation, rather than suppression of synthesis, may contribute to loss of Chl in creeping bentgrass during heat-induced leaf senescence. One interesting finding in this study is that glutamate can control Chl degradation by inhibiting CHLASE and CHL-PRX but does not affect Chl synthesis through regulation of the Chl-synthesizing enzyme (PBGD). The repression of Chl degradation by glutamate could represent a metabolic pathway contributing to the preservation of leaf Chl and photochemical efficiency, in turn attenuating heat-induced leaf senescence in creeping bentgrass (Fig. 10).
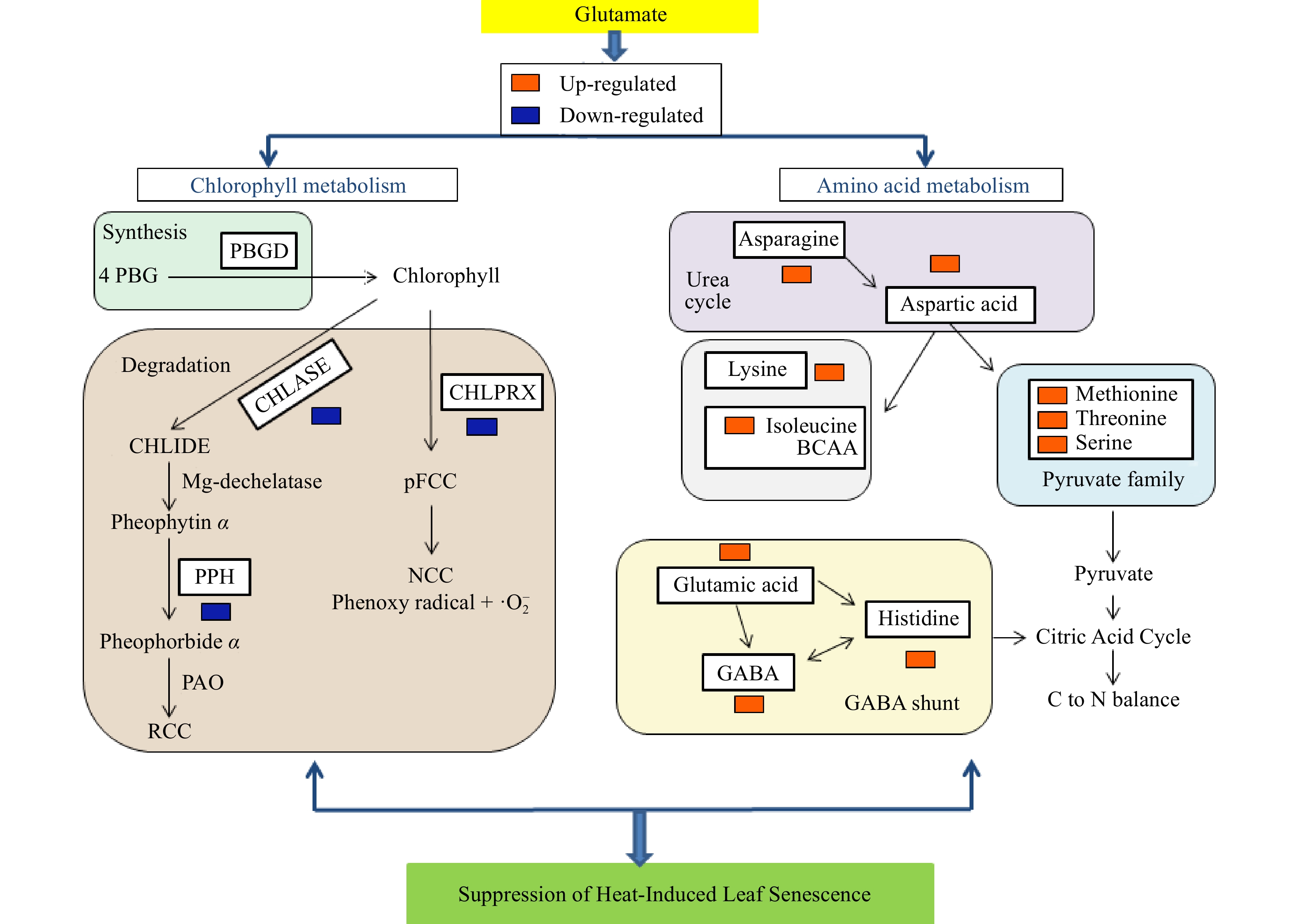
Figure 10. Visual pathway illustrating glutamate-mediated regulation of Chl synthesis and degradation and amino acid metabolic pathways for the abatement of heat-induced leaf senescence in creeping bentgrass.
Amino acid metabolism is another metabolic pathway that may be connected with glutamate-mediated modulation of heat-induced leaf senescence (Fig. 10). Among all amino acids detected in this study, four amino acids (methionine, asparagine, isoleucine, and histidine) were significantly up-regulated by glutamate only under heat stress and six amino acids (glutamate, GABA, serine, threonine, lysine, and aspartic acid) were up-regulated to a significantly greater level due to glutamate treatment under heat stress compared with those under non-stress conditions. The regulation of those differentially-responsive amino acids, including endogenous glutamate, by exogenous glutamate treatment could be associated with its roles in suppressing heat-induced leaf senescence (Fig. 10).
Glutamate is the key precursor for the synthesis of GABA, catalyzed by glutamate dehydrogenase in the GABA shunt pathway[21]. GABA accumulation in plants has been positively associated with thermotolerance in a multitude of plant species, including creeping bentgrass[22−25]. Glutamate can also be metabolized to produce histidine by pyridoxal-phosphate-dependent IAP aminotransferase leading to the generation of α-ketoglutarate and L-histidinol phosphate, which is further converted to L-histidine via reactions catalyzed by histidinol phosphate phosphatase and L-histidinol dehydrogenase[26]. GABA and histidine interacts to produce homocarnosine, serving as an endogenous antioxidant in mammalian cells[27], although the biological functions of synergistic or interactive effects of GABA and histidine in plants are unknown. In the current study, the content of GABA and histidine was significantly elevated at 35 d of heat stress in plants treated with glutamate, indicating that glutamate could enhance GABA shunt and histidine metabolism, which could synergistically protect plants from heat damage by activating the antioxidant defense system.
Pyruvate metabolism through glycolysis is the major source of intermediate metabolites for the synthesis of amino acids[28]. Of the pyruvate-generating amino acids, methionine, serine, and threonine were elevated by glutamate treatment under heat stress. Increases in the levels of methionine and threonine have previously been linked to the improvement of heat tolerance in hard fescue (Festuca trachyphylla)[29] and serine accumulation has been positively related to heat tolerance in creeping bentgrass[22]. Arabidopsis mutants with loss of function for serine hydroxymethyltransferase for serine synthesis were sensitive to salinity stress and high light stress[30]. The 3-phosphoglycerate derived from glycolysis is oxidized to become 3-phosphohydroxypyruvate, which is transaminated by glutamic acid in a reaction with phosphoserine aminotransferase, yielding phosphoserine, a precursor of serine[31, 32]. Additionally, serine generated from 3-phosphoglycerate during glycolysis can be broken down into glycine or cysteine[33]. Glutamic acid is also a source of lysine under normal environmental conditions[34]. Lysine and isoleucine are branched-chain amino acids metabolized to yield acetyl-CoA, the key substrate for respiration in mitochondria[35]. In addition to their roles as electron donors in the mitochondrial electron transport chain, there is evidence that isoleucine and lysine are also catabolized into isovaleryl-CoA and donate electrons in the electron-transfer flavoprotein complex, which feeds directly into the citric acid cycle (CAC) in the respiration process for ATP production[36, 37]. Isoleucine and lysine were shown to accumulate endogenously in a heat-tolerant bermudagrass hybrid (Cynodon transvaalensis x Cynodon dactylon ‘Tifdwarf’) subjected to heat stress[38]. In the current study, levels of isoleucine and lysine were significantly increased under heat stress due to glutamate treatment. These findings indicate that treating plants with glutamate may mitigate heat stress by promoting the accumulation of amino acids that generate substrates necessary for respiratory energy metabolism, involving both glycolysis and CAC metabolic processes.
The CAC is the primary origin of constituents involved in the biosynthesis of amino acids in addition to its function in generating ATP molecules, including aspartate and asparagine, two important intermediates in the urea cycle. Aspartate is produced from the transamination of oxaloacetate in the CAC pathway and enters the urea cycle, converting ammonia to urea using bicarbonate and its amine group, which can remove toxic ammonia and maintain cellular balance of nitrogen[39, 40]. Aspartate acts similarly to glutamate as it funnels ammonia into the urea cycle and serves as a scavenger of ammonia[40]. In addition, aspartate can be converted into isoleucine, lysine, methionine, and threonine, four essential amino acids, while aspartate and asparagine can be reversibly formed by the enzyme asparaginase or asparagine synthetase[41]. Aspartate and asparagine play roles in balancing nitrogen under environmental stress conditions, including heat stress, when nitrogen compounds, such as amino acids or proteins, are the primary sources of energy[1]. In this study, glutamate-treated plants had elevated levels of aspartic acid and asparagine, suggesting that exogenous treatment with glutamate could promote the urea cycle metabolic pathway for nitrogen balance and scavenging of ammonia under heat stress.
In summary, the current study aimed to determine the mechanisms by which glutamate may regulate Chl and amino acid metabolism for the suppression of heat-induced leaf senescence. Glutamate protected the photosynthetic apparatus of plants exposed to prolonged heat stress, as indicated by elevated photochemical efficiency and Chl content, and manifested in improved turf quality. The suppression of heat-induced leaf senescence by glutamate was associated with the down-regulation of Chl degradation and up-regulation of amino acid metabolism for energy production, antioxidant defense, and nitrogen balance. The direct evidence for the regulatory mechanisms of those metabolic processes by glutamate as a repressor of leaf senescence deserves further research, which may be accomplished by genetic modification of target or down-stream genes of glutamate as a metabolic precursor or signaling molecule.
-
Sod plugs of creeping bentgrass (cv. ‘Penncross’) were extracted from the Rutgers University Horticultural Research Farm #2 in North Brunswick, NJ and transplanted into polyvinyl chloride pipes (40 cm depth × 10 cm diameter), each filled with sterilized fritted clay (Profile Products, Deerfield, IL). Through an acclimation period of 28 d, plants were maintained in a greenhouse at average day and night temperatures of 22 and 17 °C, respectively. Lighting was controlled at approximately 700 µmol m−2 s−1 photosynthetically active radiation, administered by sunlight and sodium gas-discharge lamps, for a photoperiod of 16 h. Plants were irrigated manually on a daily basis, trimmed twice per week to a height of 1.9 cm, and fertilized once a week with half-strength Hoagland’s nutrient solution[42] before being transferred to climate-controlled growth chambers (Environmental Growth Chambers, Chagrin Falls, OH), set to a temperature of 22/18 °C (day/night), 60% relative humidity, 750 µmol m−2 s−1 photosynthetically active radiation, and a photoperiod of 14 h. During the 7-d acclimation period, plants were manually irrigated with 500 ml water on a daily basis and fertilized once per week with half-strength Hoagland’s nutrient solution.
Experimental design and treatments
-
The current study was carried out in a split-plot design, designating the temperature treatment (heat stress and non-stress) as the main plot and chemical treatments as the sub-plots. All chemical treatments consisted of four replicates, which were randomly arranged among four growth chambers on a biweekly basis in order to curtail the effects of chamber conditions on results. Upon initiation of heat stress and at 7 d intervals, plants were foliar-sprayed with 60 mM glutamate or 50 ml water (untreated control) until the canopy was fully saturated. The concentration of glutamate was selected as the optimal dose by screening multiple concentrations in a preliminary experiment. Heat conditions were imposed at temperatures of 35/30 °C (day/night) for a duration of 35 d, while plants under non-stress conditions were maintained at 22/18 °C (day/night).
Physiological measurements
-
To measure overall plant health, turf quality (TQ) was visually evaluated on a numerical scale of 1 through 9, dependent on qualities such as green color, texture, uniformity, and canopy density according to Beard[43]. On this scale, a rating of 1 signifies turf that is dry, brown in color, and unhealthy or dead, while a 9 is assigned to an evenly-textured, green canopy that is not at all damaged, and the minimal rating at which TQ is considered adequate is a 6. Leaf photochemical efficiency was quantified using an FIM 1500 fluorometer (ADC BioScientific Ltd., Herts, UK) by measuring maximum fluorescence (Fm) and variable fluorescence (Fv) in dark-acclimated leaves and was expressed as a ratio (Fv/Fm). Leaf Chl content was measured by excising fresh leaf tissue (0.1 g) from each plant and submerging the tissue in 10.0 ml dimethyl sulfoxide (DMSO) in complete darkness for 3 d. Following incubation, absorbance values of the extracted samples were read at 663 and 645 nm using a spectrophotometer (Thermo Fisher Scientific, Inc. Madison, WI), leaves were oven-dried for 72 h, weighed on a balance, and the equation by Arnon[44] was used to quantify total Chl content.
Evaluation of the activities of Chl-synthesizing and –degrading enzymes
-
Leaves harvested from plants at 35 d of heat stress were frozen in liquid nitrogen and stored at a temperature of −80 °C, prior to measurement of enzymatic activities. The activities of PBGD, the rate-determining enzyme for Chl synthesis, and three important Chl-degrading enzymes, CHLASE, PPH, and CHL-PRX, were quantified.
Chl was extracted and purified from leaf samples harvested under non-stress and heat stress conditions, and the tissue was used in a series of enzymatic reactions. Extraction of Chl was performed by grinding 10 g of flash-frozen leaves into a fine powder using liquid nitrogen, adding 50 ml of chilled propan-2-one, and incubating the sample under dark conditions for 2 h at 4 °C. Purification of Chl from the extract was performed by directly implementing the methods provided by Iriyama et al.[45]. The pellet resulting from the purification was solubilized in 50 ml propan-2-one to a concentration of 500 µg ml−1 Chl.
A solution containing 1.0% Triton X-100, 0.5 M KH2PO4 buffer (pH 7.0), 0.1 M phenylmethanesulfonyl fluoride, and deionized water, was concocted for the purpose of measuring the activities of Chl enzymes in a series of bioassays and mixed with 400 mg of ground, frozen leaf tissue, which supplied the necessary enzymes. The slurry was subsequently ground with 3.0% 1-ethenylpyrrolidin-2-one and centrifuged at 9,000 g
at 4 °C for 20 min. The supernatant derived from the ground slurry was utilized in the CHLASE, PPH, CHL-PRX, and PBGD enzyme reactions as well as for analysis of the soluble protein content in individual samples. To extract protein, crude enzyme extract was combined with 20% trichloroethanoic acid and incubated for 40 min at 4 °C until the protein precipitated and could be seen with the naked eye. Once the sample was centrifuged for 15 min at 11,500 g, the supernatant was discarded, and the pellet was dried by exposure to air and then resuspended with 1.0 M NaOH. A standard curve analysis was performed to measure soluble protein, using bovine serum albumin (BSA) as a reference, where BSA was mixed with acidic Coomassie G-250 dye reagent (Bio-Rad Laboratories, Inc.) in a sequence of serial dilutions, and the absorbance of the solution for each dilution factor was measured at a wavelength of 595 nm via a spectrophotometer. The soluble protein contents quantified using the BSA standard curve were finally implemented into calculations for determining Chl enzyme activities following the execution of the enzymatic reactions, each detailed below. Enzymatic activity of PBGD was quantified according to the methods provided by Jones and Jordan[46], with minor alterations to the reagents used. Enzyme extract (100 µl) of each sample was added to a reaction tube containing 2.5 mM diaminoethane-tetraacetic acid (EDTA), 2.0 mM porphobilinogen, 0.1 M tromethane hydrochloride (pH 7.5), 1.0 mg/ml BSA, and 0.015 M MgCl2, and the mixture was incubated for at 37 °C for 1 h. To halt the reaction, 5.0 M HCl and 0.1% para-quinone were added, and absorbance was immediately read via spectrophotometer at a wavelength of 405 nm.
The protocol of Fang et al.[47] was used to determine CHLASE activity, with modifications made to the reagents comprising the reaction solution. The enzyme extract (100 µl) from each sample was combined with 200 µl purified Chl, and the mixture was added to 300 µl propan-2-one, 20 µl 100 mM ascorbic acid, and 700 µl 50 mM tromethane hydrochloride buffer (pH 7.0), and the solution was incubated for 1 h at 35 °C. To stop the action of CHLASE, 2.0 ml hexane, 1.0 ml, propan-2-one, and 1.0 ml 500 mM tromethane hydrochloride buffer (pH 9.0) were added to each reaction tube, and the solution was shaken to separate the chlorophyllide produced in the reaction from the other intermediates. Once reaction tubes were centrifuged at 10,000 g for 15 s to create a stronger distinction between the two phases, the activity of CHLASE was quantified by reading the absorbance of chlorophyllide from the lower aqueous phase at 665 nm on a spectrophotometer.
Following the procedure outlined by Kaewsuksaeng et al.[48] with modifications to reagents and incubation conditions, PPH activity was quantified. The pheophytin used in the subsequent enzymatic reaction was generated by adding 60 µl 0.1 M HCl per 1.0 ml Chl and incubating for 5 min at room temperature. Pheophytin (100 µl) was mixed with 200 µl enzyme extract, 600 µl 20 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) buffer (pH 7.75) and 100 µl 1% Triton X-100, and the solution was incubated in the dark at 30 °C for 30 min. To stop the conversion of pheophytin to pheophorbide, 1.0 ml propan-2-one, 2.0 ml hexane, 100 µl 0.5 M Tromethane hydrochloride buffer (pH 9.0), and 1.0 ml deionized water was added. After samples were shaken to separate pheophytin and pheophorbide to the upper and lower phases, respectively, the absorbance of pheophorbide generated was quantified at 665 nm on a spectrophotometer.
Leaf CHL-PRX activity was quantified using the procedure provided by Aiamla-or et al.[49], with amendments made to the reagents comprising the reaction solution. Enzyme extract (50 µl) derived from each sample was added to a solution containing 100 µl Chl extract, 1.0 ml 0.1 M KH2PO4 buffer (pH 7.0), 100 µl 25 mM p-coumaric acid, and 100 µl 1.0% Triton X-100. To initiate the degradation of Chl by CHL-PRX, 100 µl 1.0% H2O2 was added to the solution to initiate CHL-PRX activity, and the rate at which Chl decreased was quantified at 668 nm via spectrophotometer at 20 s intervals for a duration of 10 min. A decrease in absorbance of 0.1 mg−1 protein min−1 defined one unit of CHL-PRX activity.
Quantification of free amino acids
-
To determine free amino acid content, ultra-high-performance (UHPLC) hydrophilic liquid chromatography (HILIC)-tandem mass spectrometry (MS/MS) was performed using the instruments, preparatory specifications, and procedural methods outlined in Yuan et al.[50] with no modifications or amendments.
Statistical analysis
-
Using SAS version 9.2[51], the effects of temperature and chemical treatments were analyzed using a two-way analysis of variance (ANOVA) test according to the general linear model procedure. Fisher’s least significant difference (LSD) test was utilized at a probability level of p < 0.05 to separate the differences between chemical and temperature treatments.
-
The authors sincerely acknowledge the Rutgers Center for Turfgrass Science and New Jersey Agricultural Experiment Station for their support in funding this research.
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Rossi S, Chapman C, Yuan B, Huang B. 2021. Glutamate acts as a repressor for heat-induced leaf senescence involving chlorophyll degradation and amino acid metabolism in creeping bentgrass. Grass Research 1: 4 doi: 10.48130/GR-2021-0004

Glutamate acts as a repressor for heat-induced leaf senescence involving chlorophyll degradation and amino acid metabolism in creeping bentgrass
- Received: 06 February 2021
- Accepted: 23 March 2021
- Published online: 09 April 2021
Abstract: Leaf senescence is accelerated by prolonged exposure to high temperatures in cool-season plants and is characterized by loss of chlorophyll and adjustment in amino acid metabolism. The objectives of this study included ascertaining how glutamate may affect the tolerance of cool-season grass species to heat stress in relation to leaf senescence through the regulation of leaf chlorophyll (Chl) and amino acid metabolism. Creeping bentgrass (Agrostis stolonifera L.) plants were maintained for 35d under optimal growth temperature (22/18 °C, day/night, non-stress control) or heat stress (35/30 °C, day/night) in growth chambers, and glutamate (60 mM) or water (untreated control) was applied weekly as a foliar treatment. Under heat stress, turf quality (TQ), photochemical efficiency (Fv/Fm), and Chl content were significantly higher in plants treated with glutamate at 14-35 d, 7-28 d, and 7-35 d, respectively, compared to untreated controls. Glutamate application significantly reduced the activities of chlorophyll-degrading peroxidase and chlorophyllase, two Chl-degrading enzymes, at 14, 21, and 35 d of heat stress, whereas activity of the rate-limiting Chl synthesis enzyme, porphobilinogen deaminase, was not significantly affected. Glutamate treatment significantly increased the endogenous content of serine, threonine, lysine, aspartate, glutamate, and GABA under heat stress, while methionine, asparagine, isoleucine, and histidine were significantly up-regulated exclusively under heat stress. These findings suggest that glutamate could serve as a repressor for heat-accelerated leaf senescence by suppressing Chl degradation and activating amino acid metabolism involved in energy production, antioxidant defense, and nitrogen balance.