








-
Myxomycetes are present in cities. They have been recorded in urban settings in the past[1] and the information collected in previous studies has contributed to the understanding of their ecology[2]. However, due to the scarcity of studies of myxomycetes in urban environments, published data still shows an incomplete picture of their ecological dynamics. For instance, in two studies carried out in Australia and Costa Rica, Hosokawa et al.[3] and Rincón-Marín et al.[4] found that myxomycete diversity was not affected by the position of the sampling location relative to the urban centre, but the number of records was clearly affected by the location in the second study. In such investigation, more myxomycetes were recorded in areas surrounding the city than in locations closer to the urban centre.
Even though it is acknowledged that myxomycete abundance could be more affected by urbanization, such a claim is hard to make[3]. As recently shown, occurrence data based on sporocarps points in the same direction, but the myxomycete life cycle is affected by several variables and the predictability of fruiting pattern productivity requires long-term datasets for proper evaluation[5]. Long-term datasets have the advantage that assignable causes can be discovered since data are collected across a broader inferential space (i.e., more natural, and intrinsic variability is recorded in the dataset). However, a number of short-term studies carried out with comparable methodologies can also create a picture of such causes by focusing on gradient variability at levels on which long-term projects are rigid (i.e., spatially). In this manner, pattern detection based on short-term, widely distributed, and independent studies could also be a strategy to look for such support.
Based on the second approach, easier to implement in urban contexts, reliable studies in different types of city configurations could yield interesting information to evaluate the usefulness of myxomycete ecology data for monitoring applications in anthropogenic environments. Such information would also increase the knowledge on myxomycetes in non-natural biological systems as suggested by the Sustainable Development Goals (
https://sdgs.un.org/goals ). Within this context, the present study was designed with the objective of generating data on myxomycetes in three different urban landscapes within the Neotropical region. The reproductive stage of these organisms, represented by the formation of sporocarps, has the potential of generating relevant information for the management of green spaces, the evaluation of sustainability and the modelling of microorganisms in urban landscapes. -
Myxomycetes were recorded from all three studied urban landscapes. A total of 570 records were obtained, from which 289 were associated with SJO, 160 with CIX and 121 with MDI. Such differences were significant (F(2,20) = 9.3, p = 0.001). These records were assigned to a total of 38 morphospecies that were divided in 29 from SJO, 20 from MDI and 17 from CIX (Table 1). Of the 420 examined samples in moist chamber cultures, only 88 did not yield myxomycetes indicating that 79% of moist chambers were positive during the complete experiment. In CIX, 16 out of the 38 negative samples were associated with route 1. Similarly, from the 35 negative samples in MDI, 16 were associated with route 2. In SJO, eight out of the 15 negative samples came from route 3. In contrast, route 3 in CIX, route 1 in MDI and route 1 in SJO were associated with the highest number of samples yielding myxomycetes. Such results shows that sampling routes in the windward region were associated in all cases with the highest values of positive samples. In the case of SJO, route 2 in the leeward region also showed high values. Interestingly, only in CIX, the urban center showed a higher percentage of positive samples (95%) than the other sampling locations.
Table 1. Number of records of myxomycetes per species associated with the three urban landscapes studied herein.
Species Urban landscapes MID SJO CIX Arcyria afroalpina 6 Arcyria cinerea 11 27 23 Arcyria denudata 1 Badhamia affinis 6 Badhamia melanospora 1 1 Comatricha elegans 3 Comatricha nigra 3 1 Comatricha pulchella 3 Didymium anellus 3 9 Didymium bahiense 18 22 16 Didymium clavus 1 6 1 Didymium difforme 42 5 Didymium dubium 2 Didymium iridis 3 8 Didymium minus 20 18 7 Didymium nigripes 2 Didymium squamulosum 10 36 Diachea leucopodia 3 Diderma hemisphaericum 4 43 Fuligo intermedia 3 Hemitrichia minor 3 Lamproderma scintillans 13 Perichaena chrysosperma 3 5 1 Perichaena depressa 13 4 12 Perichaena pedata 4 Perichaena vermicularis 2 2 Physarum atacamense 60 Physarum auriscalpium 1 Physarum bivalve 1 Physarum cinereum 9 3 5 Physarum citrinum 2 Physarum compressum 7 24 3 Physarum decipiens 1 Physarum didermoides 1 2 Physarum melleum 1 Physarum pusillum 7 5 5 Physarum stellatum 1 Stemonitis fusca 1 1 For abbreviations see Material and Methods. Significant differences were observed in the number of records between conurbated and compact urban landscapes (t = 4.2, p = 0.0004) showing that SJO, the widespread conurbated city yielded twice the number of records per sampling location (average of 41.2 vs 20.0, see Table 2) than either MID or CIX. Similarly, individual sampling locations in SJO were associated with an average value of 14.4 recorded species, followed by MDI with 7.8 and CIX with 7.7 species. This result also led to significant differences between conurbated and compact landscapes (t = 5.6, p = 0.00002). Given that both the Simpson and Shannon indices of diversity use the species richness and count values for calculations, differences were also observed in those estimators for conurbated and compact urban landscapes. No differences were recorded in either the number of records (t = 1.4, p = 0.1) or the number of species (t = 0.1, p = 0.9) per location between the two compact cities.
Table 2. Biodiversity parameters determined in the present study arranged by urban landscape and sampling location.
Urban landscape Sampling location Biodiversity parameter Number of records Species richness Simpson's
DIShannon's
DIMérida
(MDI)Urban center 19 8 0.79 1.81 1A 10 6 0.78 1.64 1B 32 14 0.88 2.39 2A 8 5 0.75 1.49 2B 18 7 0.83 1.87 3A 16 7 0.84 1.89 3B 18 8 0.78 1.79 San José
(SJO)Urban center 26 12 0.87 2.27 1A 33 17 0.91 2.65 1B 51 14 0.86 2.27 2A 44 17 0.89 2.49 2B 71 19 0.89 2.52 3A 30 12 0.87 2.25 3B 34 12 0.85 2.15 Chiclayo
(CIX)Urban center 32 11 0.85 2.13 1A 14 6 0.68 1.43 1B 16 7 0.74 1.59 2A 18 5 0.58 1.16 2B 29 11 0.78 1.9 3A 22 7 0.72 1.55 3B 29 7 0.76 1.62 The highest values for the different levels are highlighted in bold. When the number of records and species were analyzed in each urban landscape in terms of sampling routes, no clear pattern was observed (Table 3). The windward routes showed high numbers of records and species and in the case of MDI (route 1) and CIX (route 3), they were associated with the highest numbers of normalized records. In SJO, however, the leeward route (number 2) showed the highest number of records. In terms of species, the leeward routes in SJO (route 2) and CIX (route 2) were associated with the highest numbers, whereas the windward route (route 1) showed the highest number of species in MDI. Contrastingly, route 2 in MDI, route 3 in SJO and route 1 in CIX were all associated with the lowest values of normalized records and species. All these routes were located in a perpendicular position relative to the dominant winds in the studied urban landscapes.
Table 3. Biodiversity parameters determined in the present study arranged by urban landscape and established sampling route.
Urban landscape Route Biodiversity parameter Number of records* Species richness Simpson's DI Shannon's DI Mérida (MDI) Urban center 19 8 0.84 1.99 Route 1 14 16 0.93 2.77 Route 2 8.6 9 0.89 2.21 Route 3 11.3 12 0.9 2.44 San José (SJO) Urban center 26 12 0.91 2.49 Route 1 28 20 0.91 2.68 Route 2 38.3 23 0.92 2.78 Route 3 21.3 16 0.9 2.5 Chiclayo (CIX) Urban center 32 11 0.88 2.29 Route 1 10 9 0.77 1.87 Route 2 15.6 12 0.76 1.92 Route 3 17 10 0.77 1.84 * For normalization, this value was calculated as the number of records per set of 20 samples. The highest values for the different levels are highlighted in bold. In general, more records and more species were observed in the outer zones relative to the urban centre (Table 4). However, no significant differences were recorded in these gradients (F(2,20) = 1.4, p = 0.2, and F(2,20) = 0.4, p = 0.6 for number of records and number of species, respectively). Despite that, such increasing pattern towards the outer sections of the city was clear in both MDI and SJO, the two urban landscapes surrounded by forested environments. In CIX, there were more species in the outer zone but most of the other parameters showed higher values in the urban centre. Contrastingly, in both MDI and SJO, the lowest number of species was associated with the sampling location at the urban centre. However, for both MDI and CIX, the inner impact zone showed the lowest number of records.
Table 4. Biodiversity parameters determined in the present study arranged by urban landscape and urban impact zone.
Urban landscape Urban
impact
zoneBiodiversity parameter Number of records* Species richness Simpson's DI Shannon's DI Mérida (MDI) Urban center 19 8 0.79 1.8 Zone A (inner radius) 11.3 13 0.86 2.28 Zone B (outer radius) 22.6 16 0.9 2.52 San José (SJO) Urban center 26 12 0.87 2.27 Zone A (inner radius) 35.6 23 0.91 2.7 Zone B (outer radius) 52 23 0.88 2.53 Chiclayo (CIX) Urban center 32 11 0.85 3.13 Zone A (inner radius) 18 10 0.69 1.61 Zone B (outer radius) 24.6 13 0.81 2.07 * For normalization, this value was calculated as the number of records per set of 20 samples. The highest values for the different levels are highlighted in bold. The pH values recorded in the moist chambers showed an average of 6.77 ± 0.32, 6.66 ± 0.59, and 6.48 ± 0.64 for MDI, SJO and CIX, respectively. No correlation between pH values and biodiversity parameters were observed by sampling locations, by routes or by impact zones.
-
A clear historical trend in myxomycete research has been to focus on biodiversity matters in natural environments. Anthropogenic settings such as urban systems have received little attention from researchers. Myxomycetes have clearly been documented in urbanized situations[6] and evolutionary dynamics in these non-natural systems can provide relevant data to understand the effect of global phenomena on biodiversity[7]. As such, despite the taxonomic shortcomings of studying fruiting bodies, the reproductive stage of myxomycetes provides remarkable information on the natural resilience and adaptability of these organisms to urban pressures.
In the present study, compositional differences among myxomycete assemblages recorded in each one of the three studied urban landscapes highly likely show, on a primary basis, differences in species adaptability and life zone-based distribution. For instance, Didymium anellus and Perichaena depressa were only recorded in the dryer landscapes (MDI and CIX), Arcyria afroalpina was only recorded in the premontane environment (SJO) and Physarum atacamense was only observed in the native desert system of South America[8]. Similarly, widely distributed species such as Physarum compressum, Ph. pusillum and Arcyria cinerea were recorded in all three urban landscapes studied herein. However, the extent to which urbanization influences fruiting patterns in myxomycetes remains largely undocumented and floristic examinations can only reveal individual species preferences. For that reason, a broader evaluation of patterns is more adequate to assess potential uses of the myxomycetes in urban ecology[9].
It is interesting to observe, however, that few species such as Badhamia affinis, Fuligo intermedia or Ph. pusillum were recorded herein but not in the extensive work in coastal desertic areas of Peru previously carried out by Lado et al.[10]. Even more interesting to note that the last species was recorded in transitional areas further from the coast[11]. Similarly, it is interesting that Ph. auriscalpium and Ph. citrinum, recorded herein, had not been observed in the central urbanized area of Costa Rica[12]. Both situations demonstrate that the mentioned species are present in the respective regional species pool but are very elusive. This suggests that the conditions provided by the urban landscapes studied herein influenced in some way their recording, providing support for the idea that in some cases urban landscapes can facilitate the dynamics of species occurrence, via fruiting bodies[13]. Small fluctuations in temperature, humidity and rainfall are known to differ between urban and forested areas and are also known to affect the development of sporocarps of myxomycetes in the tropics[5]. The same situation likely took place in the Yucatan Peninsula, but comparisons are hard to make due to the lack of recent published data for that area.
In a broader sense, it is quite interesting that from all collected samples, the least productive sampling routes were also the ones with the lowest number of records and species. Even though this association is clearly mathematical, collected material in such routes seemed to have exhibited disadvantages for the formation of fruiting bodies in laboratory isolation conditions. Such disadvantages could be related to local aspects such as different vegetation or stage of decay associated with each of the different routes but could also be related with other aspects at the landscape level. As mentioned earlier, all these poor routes had perpendicular positions relative to the direction of the dominant winds. In contrast, all windward routes showed high values of positive samples, number of records and number of species. It is known that windward locations are moister than leeward ones due to the higher values of air moisture[14] which is retained in windward surfaces as the wind crosses the urban landscape[15]. Whether this potential difference in moisture might have influenced the results shown herein is hard to explain without local climate data, but it represents an interesting issue to examine in more detail in future studies.
Interestingly, results also showed that in the case of the conurbated city (SJO) a leeward route also displayed high values for the parameters evaluated suggesting that the non-compact nature of this urban landscape affected the results. In this case, it is possible that the larger covered area and higher availability of suitable environments in the leeward areas of the conurbated city (due to the green inter-conurbated areas) played a role. It has been documented that such green spaces within urban landscapes influence local levels of moisture, temperature, and wind[16]. Also, it is known that conurbations might attenuate the negative effect of urbanization on some organisms[17, 18], presumably by means of stabilizing habitat connectivity. If this is the case, it is likely that the same mechanisms could be taking place on myxomycete dynamics in urban landscapes. Habitat loss does affect the occurrence of sporocarps of myxomycetes[19] by decreasing the availability of substrates upon which the different species produce these structures. Such limitation of substrates is buffered when habitat connectivity is in place and species can move (i.e., spore-based colonization) across wider geographical spaces.
The remarkable observation in the present study is that both the number of records and species in the conurbated urban landscape were practically double the values observed in the compact cities. Recent literature in the Neotropical region has demonstrated that arid zones can sustain a similar or higher diversity of myxomycete morphospecies than moister environments[10, 11]. As such it is unlikely that the life zone characteristics alone might have determined such disparity and that city dynamics are involved in the complexity of results documented herein. In a recent study of airborne urban myxomycetes in Medellín, Colombia[4], differences in species richness and diversity between two urban locations were attributed to the degree and type of urbanization (i.e., industrial urbanization vs. residential urbanization), which affects convective currents and wind direction. In the present study, the green inter-conurbated spaces seemed to have been more important to explain the results.
In any case, myxomycete resilience to urban pressures is clearly a mechanism related to the production of sporocarps. Based on these structures, previous studies by Hosokawa et al.[3] and the present one, have found no statistical differences in the number of records, the number of species or species diversity between locations within the inner core of the urban landscape and the outer areas. In other words, if equivalent substrates are available, there is a high probability of recording similar assemblages of species in the functional core of the city or in the outskirts. The results presented herein suggested an increasing trend towards the outer parts of the urban landscape in MDI and SJO, but such trend could simply be related to the higher frequency of available substrates in the outer areas (less urbanization means more green space and by default more substrates for myxomycetes to grow on). Similarly, the fact that the urban centre in CIX showed high values for most of the parameters evaluated could be related to the same factor because there are more available substrates within such a city (i.e., public parks and ornamental areas) than in the desertic surroundings.
Since studies of urban ecology in locations outside the developed northern hemisphere are generally scarce[20], results from the present study have the capability to contribute to global evaluations of biological data in anthropogenic systems. Urban ecology of myxomycetes has a great potential for monitoring purposes due to their resilience to urban pressures. Both results from previous studies and the present one suggest that the management of green spaces within urban landscapes is the limiting factor determining their presence. Considering the balancing role of these organisms in microbial ecosystems[21, 22], it is important to sustain them in urban landscapes.
-
Conurbations can represent important obstacles for urban sustainability but may increase biological connectivity via a higher green cover within the general urban landscape. Results and analyses from the present study suggest that myxomycete occurrence, determined by means of fruiting bodies, could benefit from the latter mechanism in tropical settings. The lack of differences at different levels of analysis in the present study may also point out that the limiting factor for myxomycete presence in cities is the availability of substrates. Regardless of its particular climate, this study showed that such patterns takes place in tropical urban landscapes. In the case of deserts, cities become shelters for species.
-
The present study was carried out between January-June 2016 in the Neotropical region. For this study, three cities with different characteristics were chosen and used as examples of urban landscapes. All three cities were located within 10 degrees of longitude and were individually separated between 12 and 15 degrees of latitude to minimize geographical distances (Fig. 1).
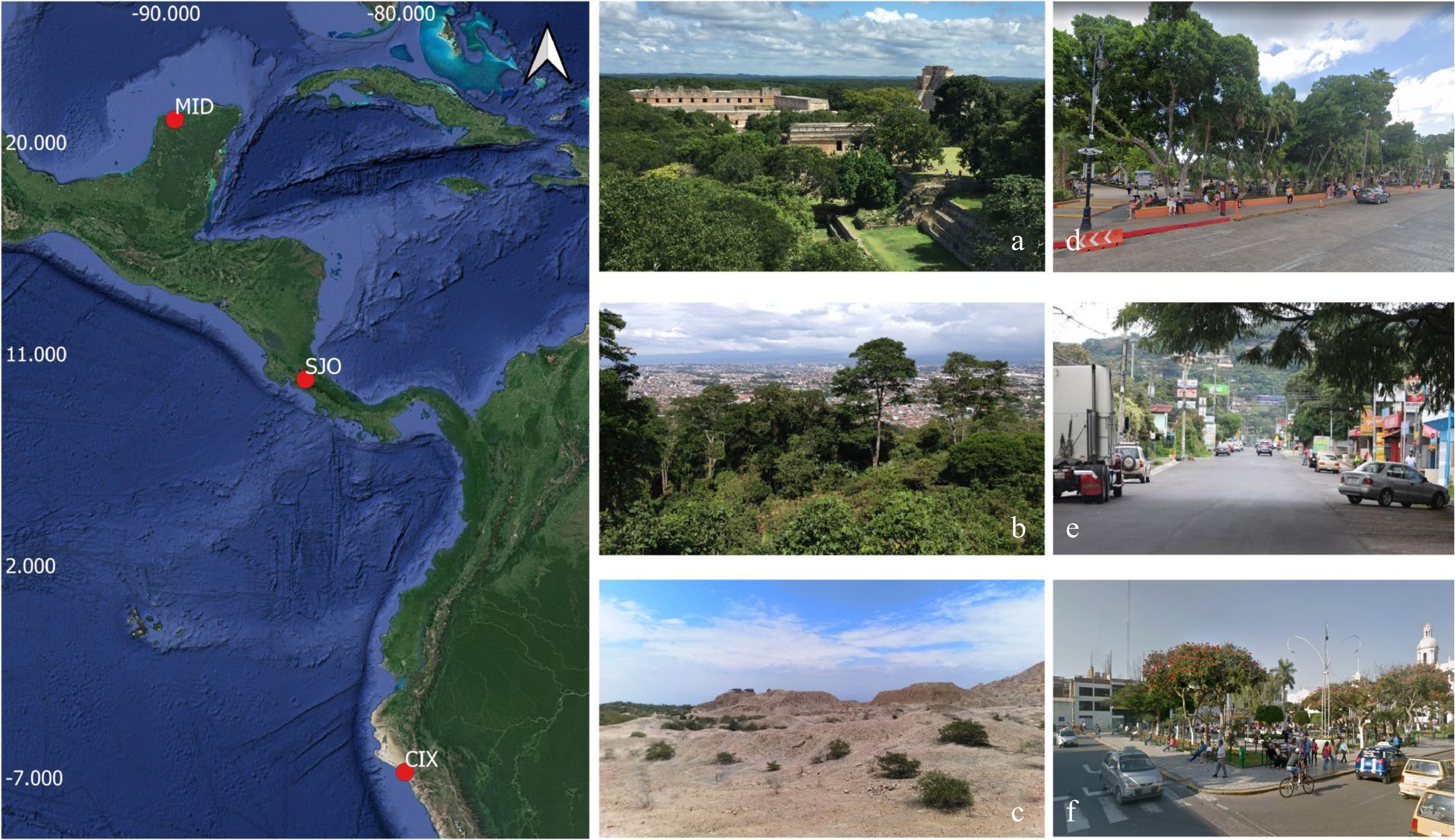
Figure 1.
Geographical location of the three urban landscapes studied in the present study (left) with images showing the characteristic vegetation and urbanization patterns in them. (a) Dry seasonal tropical forest (during rainy season) of the Yucatan Peninsula surrounding the city of Mérida, Mexico (MID). (b) Premontane moist forest of the Central Valley of Costa Rica surrounding the city of San José (SJO). (c) Desert with scattered vegetation of the Peruvian pacific coast surrounding the city of Chiclayo (CIX). (d), (e), (f) Examples of disturbance caused by urbanization and human activities in actual sampling locations in MDI (top), SJO (middle) and CIX (bottom) – the last three images are from Google Maps.
The first urban landscape corresponded to the Great Metropolitan Area of San José, Costa Rica (abbreviated thereafter as SJO), a region with approximately 2.5 million people. This urbanized area is characterized by numerous conurbations (defined herein as extensions of urban sprawl coming out of an urban zone in undefined patters) with a median population density of 1,800 people/km2. The second studied urban landscape was the city of Mérida, México (MID), an urbanized area with approximately 900,000 people and a median population density close to 1,100 people/km2. This urban area is compact (with a defined shape and without extensions of urban sprawl) and does not have large conurbations. The third urban landscape evaluated herein was the city of Chiclayo, Peru (CIX), with approximately 1 million people and a median population density of approximately 1,900 people/km2. This urban area has few conurbations, but it has a compact character. In this manner, SJO is an example of a widespread conurbated city, whereas MID and CIX are examples of compact ones. Since conurbations are mostly the product of unplanned urban sprawl, conurbated cities tend to have more green spaces (albeit by accident) than compact ones.
Due to their geographical location, all three cities differ in both climate and dominant natural environments surrounding the urbanized area. SJO has a tropical moist climate, with dominant winds coming from the east and premontane tropical moist forests surrounding the city. MID has a tropical dry climate, the dominant winds come from the east, and the surrounding areas are typical of lowland tropical dry forests. CIX is a city with an extremely arid climate surrounded by desert and the dominant winds come from the south. All three selected urban landscapes show similar average daily temperatures between 21−25 °C, but precipitation is quite different among them with an average yearly value of 1900 mm/year for SJO, 1,000 mm/year for MID and just 27 mm/year for CIX. Accordingly, SJO and MID are examples of landscapes with tropical forests subjected to the influence of the northeast trade winds, whereas CIX is an example of a non-forested tropical landscape with influence of the southeast trade winds.
In all three urban landscapes, a location determining the urban center was defined using a geographic centroid approach based on the area covered by the urban patch. The average distance between such center and the urban edge in different directions was used to create two areas of urban impact at approximately 2.5 and 5 times such value. These areas defined an 'internal' (abbreviated as A) and an 'external' (B) impact zones relative to the urban center. Three routes, labeled 1, 2, and 3, at an equivalent elevation (± 100 m) to the urban center were established for the selection of two locations in each route. All these six locations were positioned at the edge of the predefined urban impact zones and were abbreviated using a combination of route and urban impact area in all cases (i.e., 1A for location on route 1 within the internal urban impact zone). All these processes were carried out using an a priori spatial analysis carried out in QGIS to minimize bias (Fig. 2).
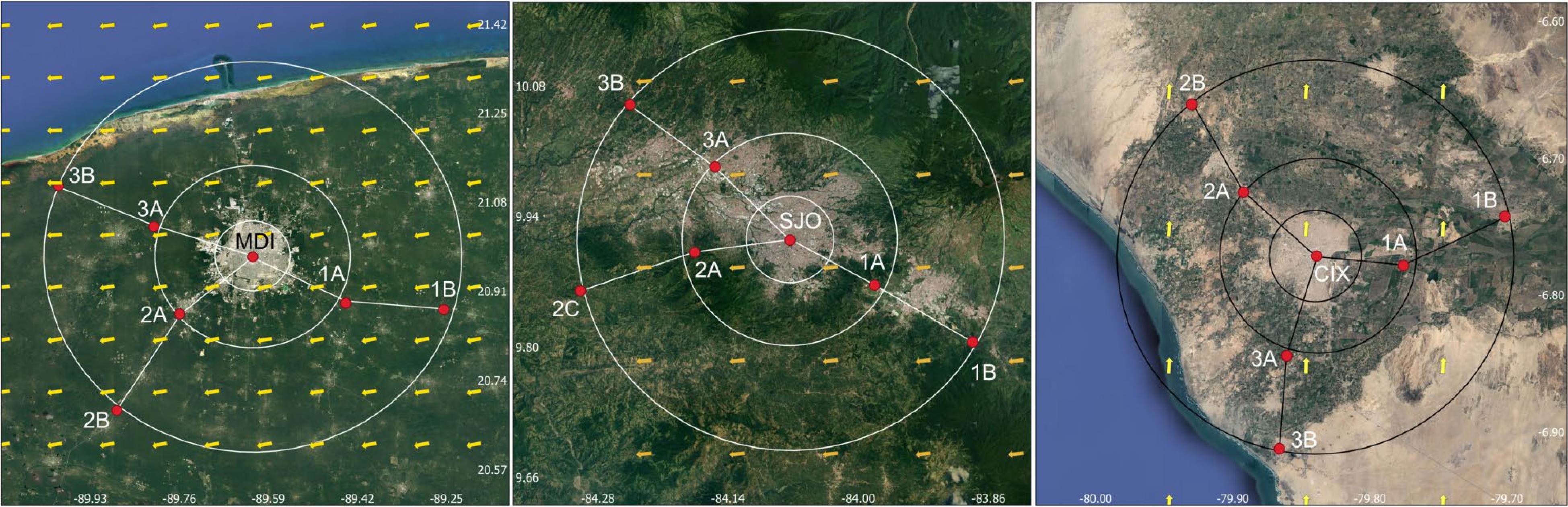
Figure 2.
General aspect of the three urban landscapes studied herein in relation with the sampling strategy of one central location, three sampling locations in an inner (A) ring and three more locations in an outer (B) ring. From left to right, Mérida, Mexico (MDI), San José, Costa Rica (SJO) and Chiclayo, Peru (CIX).
In all six established locations moving out of the urban center as well as in the original central location in all three urban landscapes, a series of collections of material for isolation was carried out. Each consisted of 20 samples of ground litter collected at the edge (maximum 2 m away) of public roads for a total of 140 samples in the case of each city. As such, all sampled material was collected in a 'heavily disturbed' ecological setting. For reference, the average monthly precipitation during the period of field sampling was 10.2, 69.4 and 4.0 mm for SJO, MID and CIX, respectively, demonstrating that all urban landscapes were visited during dry periods.
Upon field collection, all the material was taken to the same laboratory for isolation of myxomycetes using the moist chamber technique. With this technique, the material was placed on a piece of filter paper within a standard petri dish and was soaked with distilled water for 24 h. After this period, the pH value of the microcosm was determined, the water in excess was decanted and the moist chamber was placed in a stack corresponding with its origin. All stacks were kept in the laboratory next to each other, and the petri dishes were rotated every week for the moist chamber to have a different relative position during the three months that they were examined. During the entire period of examination, moist chambers were studied with a stereoscope looking for the presence of myxomycete activity. When fruiting bodies were detected, they were extracted from the moist chamber, glued to pasteboard boxes, and dried out in normal room conditions, after which they were deposited in the Myxogastrid Repository of the Engineering Research Institute of the University of Costa Rica.
Statistical analyses
-
For analysis, the number of records and the species richness associated with each sampling point were determined. In all cases, both the Simpson (1-D) and the Shannon indices of diversity were also calculated. All four estimations were used as response variables to test the hypothesis of urban effect on myxomycetes, and in the case of the number of records, estimations were also recalculated per set of 20 samples. In this case, the total number of records for both Zone A (internal radius) and Zone B (external radius) was divided by a factor of 3, which corresponded to the number of sampling locations at each radius. Normality in the obtained data was evaluated using the Shapiro Wilk test, and differences in the calculated estimations were evaluated using two-tailed t-tests or one-way ANOVAs with an α-value of 0.05 depending on the number of evaluated groups. For ANOVAs, homoscedasticity was evaluated using Levene's test and in the case of differences, a post-hoc Tukey test was used to determine which data group was associated with them. For normalization of effort in some comparisons, the number of records was also expressed as a value relative to every 20 evaluated samples. All calculations were carried out in Past v. 4.05.
This work was financed by the University of Costa Rica (Vicerrectoría de Investigación 731-B7-721). Gratitude is extended to Magdalena Pavlich, Bertha Leiva Berríos and Robin G. Doss for support with the logistics of the project and help with field sampling.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Rojas C, Valverde R, Rojas PA. 2022. Myxomycetes associated with three different types of Neotropical urban landscapes. Studies in Fungi 7:14 doi: 10.48130/SIF-2022-0014 

Myxomycetes associated with three different types of Neotropical urban landscapes
- Received: 01 December 2021
- Accepted: 21 November 2022
- Published online: 05 December 2022
Abstract: Urban ecology is a research field gaining momentum due to contextual pressures to study macroecological effects on biodiversity. Herein, we evaluated the occurrence of myxomycetes in three urban Neotropical environments representing different levels of conurbation within different life zones. We used the moist chamber technique set up with ground litter collected by public roads for such evaluation. Results showed that 79% of the 420 moist chambers yielded myxomycetes and the same number of samples generated 570 records. The highest number of records and species were found in the cornurbated widespread city of the premontane moist forest life zone; and fewer records and species were recorded in the two non-conurbated compact cities. The lowest number of records was associated with the lowland dry forest life zone, whereas the lowest number of species was recorded in the desertic context. In general, more records and more species were observed in the outer zones relative to the urban centre but no significant differences were observed in either case. It is unlikely that the life zone characteristics could have primarily determined the results and highly likely played a secondary role. Also, previous geographical analyses in cities have reported no differences in myxomycete abundance or species richness in relation to the central parts of the urban landscape. The results presented herein support the latter idea, but also suggest that the presence of inter-conurbated green spaces and availability of substrates might play a role in the occurrence of sporocarps of myxomycetes in urban environments.
-
Key words:
- Cities /
- Sustainability /
- Planning /
- Ecology