








-
The Nymphaeales order, including three families (Hydatellaceae, Cabombaceae, Nymphaeaceae) and 85 species (Fig. 1), are called the water lily order[1,2], partly due to the dominant character of all these species are aquatic herbs and nearly all of the species have very beautiful flowers and leaves. Hence the water lily is recognized as the common name for all species categorized in the Nymphaeales order[3−8]. Water lilies are globally distributed, spanning from tropical to cold regions[9]. They have played vital roles in the economy, ecology, and evolutionary biology.
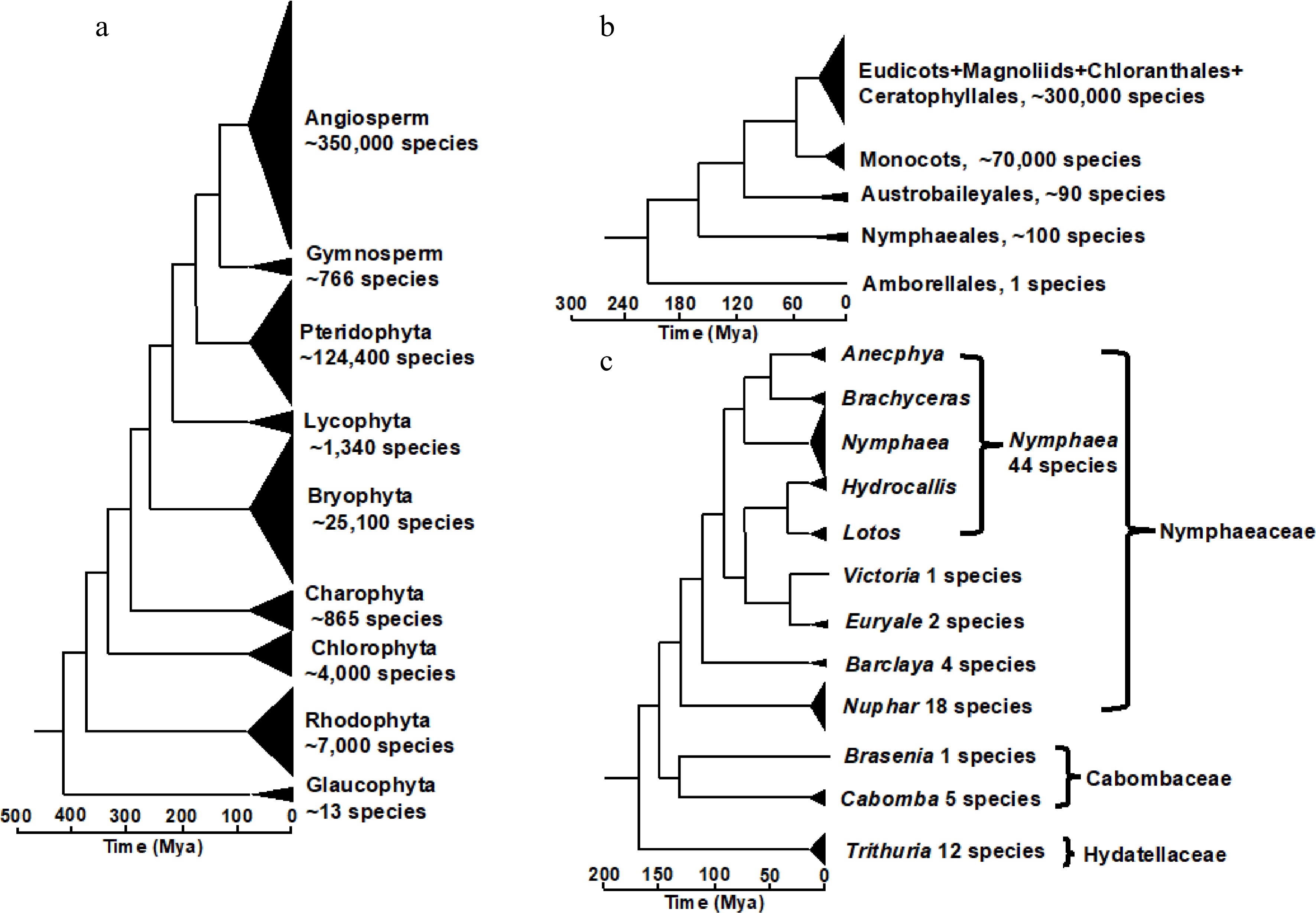
Figure 1.
The evolutionary position of water lilies. (a) Tree of green plant lineage. (b) Tree of angiosperms (flowering plants). (c) Tree of water lily order Nymphaeales. Phylogenetic relationships and timing of the nodes are based on previous phylogenomic research[9] and phylotranscriptomic research[10].
Economic, cultural, ecological, and evolutionary values of water lilies
-
Water lilies contain various economic values. The Nymphaea spp. flowers are not only good cut flowers, but can be in food and beverages[11]. For example, some Nymphaea water lilies have starchy and edible roots[12]. The seeds of Victoria and Euryale ferox are edible[13]. Nymphaea species contain various medicinal compounds, and they have been found to function in anti-bacteria[14], anti-inflammatory[15], and anti-depressant[16] capacaties. According to data from the China Flower Association, at the end of 2019, the annual output of water lilies in China was 152 million heads, and the combined market size of the lotus and water lily industries reached 10 billion CNY (
www.cqla.cn/chinese/news/news_view.asp?id=58006 ). In Lichuan county from Hubei province, China, the 'hometown of E. ferox in China', its planting area is about 2,000 hectares, and the annual output value exceeds 200 million CNY (http://news.yuanlin.com/detail/2021128/379540.htm ). Water lilies have high cultural value and a long consumption history. In ancient Greece and Rome, together with the lotus, some Nymphaea species were regarded as the embodiment of holiness and beauty and were often used as offerings to the gods. In modern times, one of the top impressionist painters Claude Monet has painted hundreds of water lilies[17]. Many countries have issued stamps or currency with water lily flowers as the theme. Water lilies have a certain ecological value, especially in water purification and heavy metal absorption[18]. However, some Nymphaea species grow very quickly and are regarded as invasive species. For example, N. odorata and N. capensis encroach heavily on the living space of other species in the same ecosystem and destroy the ecological balance (www.invasive.org/species/aquatic.cfm andwww.issg.org/database/species/ecology.asp?si=985&lang=SC .)The water lily order occupies a critical evolutionary position. Together with Amborellales and Austrobaileyales, these three orders collectively form the ANA (Amborellales-Nymphaeales-Austrobaileyales) grade, which are the first three flowering plant branches[19]. The early diverging water lily order provides a unique window to study the origin and early evolutionary features of flowering plants. Besides, the various kinds of floral scents and colors are wonderful traits for ornamental studies. Their genes and related pathways are the focus of the molecular breeding of water lilies and even other ornamental plants.
Growing research on water lilies
-
Multi-omic tools, including genomics, epigenomics, transcriptomics, proteomics, and metabolomics, are high-throughput and effective tools for plant studies. With the development of high-throughput technologies and bioinformatics tools, various omic research continues to deepen our understanding of plant traits[20]. Through high-throughput measurement of each omics and data integration, we can not only comprehensively and systematically understand gene functions and promote molecular breeding, but also provide important data and technical approaches for the research of emerging cross disciplines such as quantitative and systems biology.
To date, there are roughly 868 publications indexed in the PubMed database (Fig. 2). The annual number grows rapidly, from eight publications in 2000 to 71 in 2021. The rapid increase reveals the boosted interest in various kinds of water lilies, from Nymphaea to others.
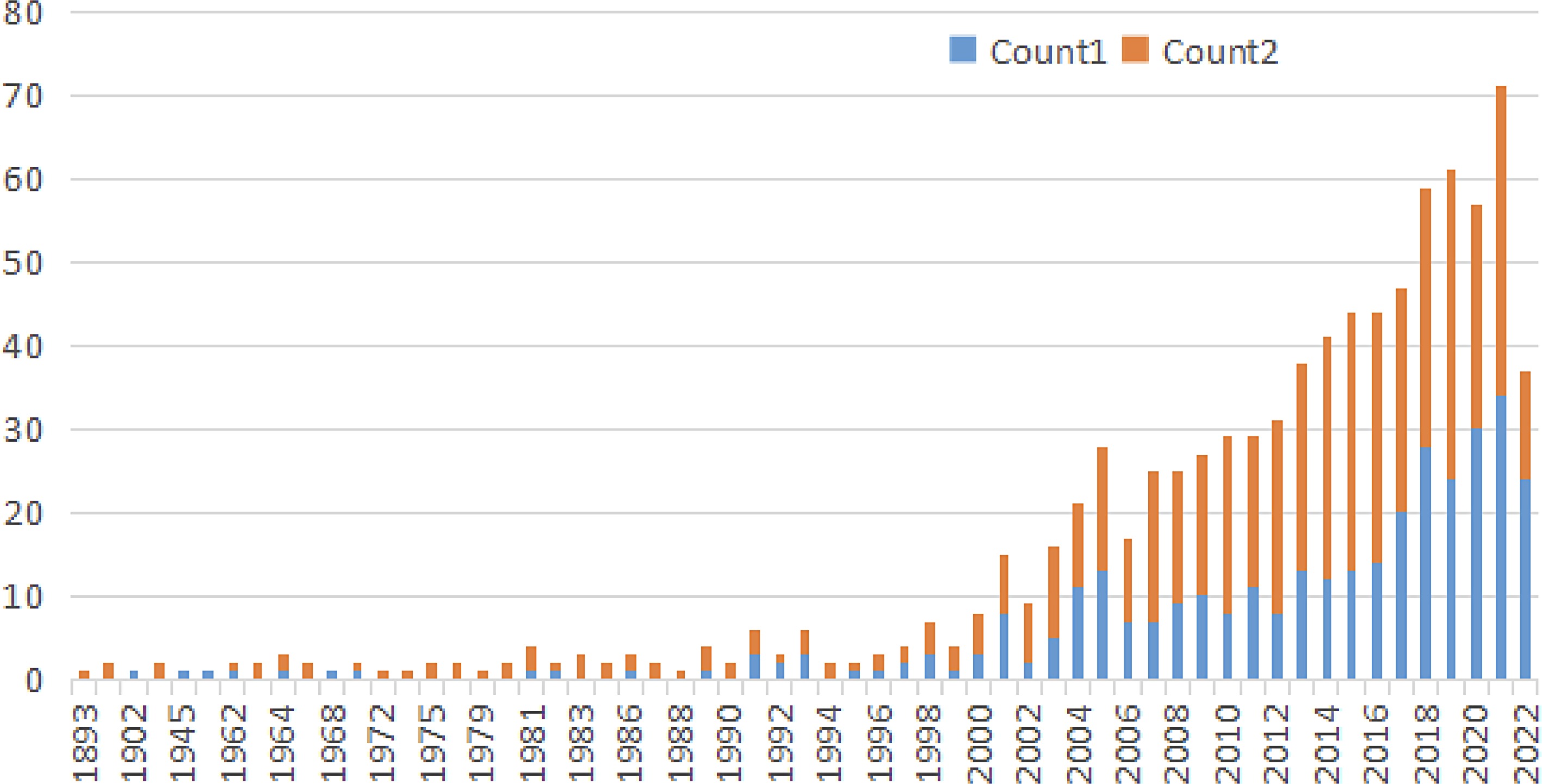
Figure 2.
The growing research publications on water lilies. The searches are based on PubMed from NCBI (
https://pubmed.ncbi.nlm.nih.gov/ ) on May 24, 2022. Count 1 shows the keywords '(nymphaea) OR (waterlily) OR (water lily) OR (euryale) OR (barclaya) OR (nuphar) OR (trithuria) OR (cabomba) OR (brasenia)', while count 2 shows the keywords 'nymphaea'. -
Chen et al. selected the N. colorata, a diploid tropical species that originated from east Africa for genome sequencing due to its relatively small genome size and small plant size[21]. Relying on the PacBio long-read sequencing technology, a 409 Mb genome is assembled, falling into 14 chromosomes[9]. The genome assembly has relatively high quality with contig N50 up to 2.1 Mb. Besides, some telomeres and centromeres are very well assembled and characterized. At the same time, the genomes of its mitochondria and chloroplast are successfully assembled using the same PacBio reads[22]. Based on its genome information, we are able to precisely reveal its evolutionary place (Fig. 3) and identify the genes responsible for the floral scent, floral color[23], as well as floral morphogenesis[24]. The expansion of biotic and abiotic stress-related genes such as WRKY genes and R genes is related to the wide distribution of the Nymphaea water lilies[25]. Some of the key genes responsible for the stomata development are lost in the N. colorata genome and may explain its unique aquatic lifestyle adaptation[26]. This is the first report on the water lily genome sequence, which is a milestone showing that water lily research is entering the genomic era.
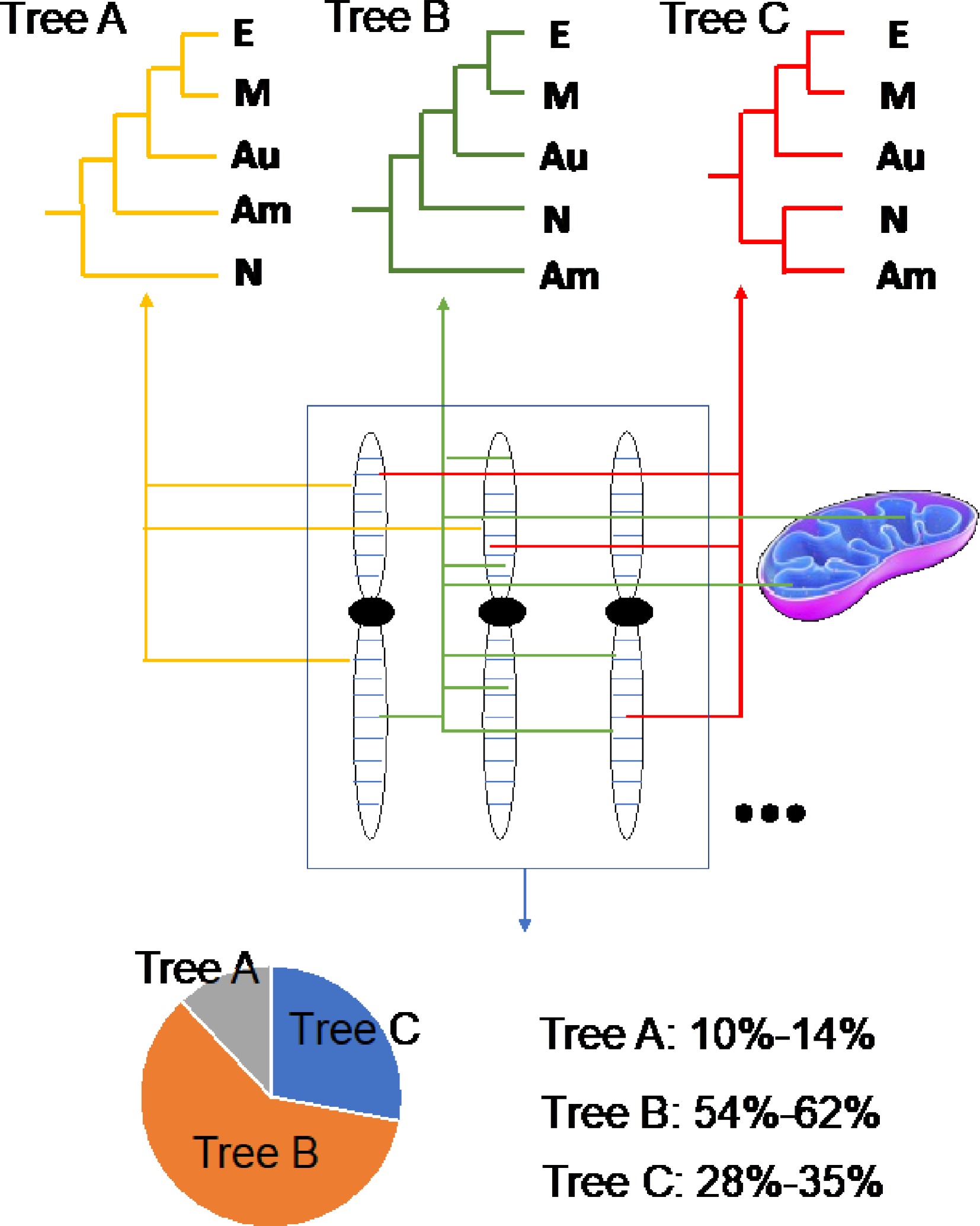
Figure 3.
Schematic diagram showing that the phylogenomic analyses reveal accurate phylogeny of angiosperms. Tree A represents that Nymphaeales, but not Amborellales forms the earliest extant angiosperm branch. Tree B represents Amborellales as the earliest extant angiosperm branch. Tree C represents Amborellales and Nymphaeales are sister lineage as the co-first earliest extant angiosperm branch. The proportion data in the piechart are from our previously published article[9]. E: Eudicots, M: Monocots, Au: Austrobaileyales, Am: Amborellales, N: Nymphaeales.
The genome sequence of N. thermarum
-
Povilus and colleagues from Harvard University assembled a draft genome of a dwarf tropical water lily N. thermarum[27]. This species originates from Rwanda (east Africa). Relying on Illumina short reading sequencing technology, they obtained a 368 Mb genome of N. thermarum. Analyses of this genome and phenotypes reveal the loss of genes enrolled in the vascular cambium, which might be a key innovation of its adaptation to the aquatic environment. This is the second report on the water lily genome.
The genome sequence of Euryale ferox
-
The Euryale genus only contains one species (E. ferox) which is mainly distributed in southern and eastern Asia. Their leaves and flowers are covered in sharp prickles and float on the water’s surface. Its edible seeds are usually called fox nuts and are used as food or diet therapy supplies in Asia. Relying on the long-read sequencing platform Oxford Nanopore Technology, Liu and colleagues assembled the genome sequence of E. ferox[28]. The genome assembly has a size of 725.2 Mb and 29 chromosomes. Phylogenomic studies also support that E. ferox is an early angiosperm branch, following the Amborella, and the incomplete lineage sorting may largely account for phylogenetic inconsistencies of different major lineages in angiosperms. This is the third report on the water lily genome, which provides important resources to reveal the specific evolution of seeds and leaves.
Whole-genome methylation researches
-
There has been little focus on the DNA methylation in water lily research. Bisulfite sequencing (BS-seq) on Nuphar advena did not find CHH type methylation in its genome. Comparative analyses of N. advena and other land plants’ relative levels of CG methylation are a consistent property across genes, and genic CHG methylation correlates with genome size[29]. We believe that in the future, more results will emerge as more genome sequences of water lilies become available.
-
Most Nymphaea species bloom in the morning and close their flowers in the afternoon. Only a few species bloom at night and close their flowers before the morning. This unique circadian flowering clock is very interesting and attractive. Chen et al. collected flower samples at different time points and sequenced the transcripts[30]. They uncovered eight expressional gene clusters related to floral circadia. They showed that auxin-related signaling pathway genes are linked to synchronized expression patterns with opening and closure processes.
The bloom of Nymphaea flowers usually last three to four days, which largely shortens the shelf life of cut flowers. Li et al. investigated when the senescence process is initiated and how it terminates the movement rhythm[31]. They identified multiple signaling pathways that were activated at the last stage, the floral closure, on the third day after flowering. Genes related to hydrolase are upregulated on the same day, indicating that petals enter the senescence stage at that time.
Floral fragrance
-
Unlike species from the Hydatellaceae family and Cabombaceae family, most Nymphaea flowers emit fragrant molecules, volatile organic compounds, to attract various kinds of pollinators. A wide variety of small molecules or metabolites are responsible for the diverse floral colors and scents. Identifying small molecules using gas chromatography-mass spectrometry (GC-MS) is becoming more common. For example, GC-MS-based screening on the flower petals of N. nouchali identified 16 biologically active phytochemicals[32]. GC-MS on the N. pubescens’s crude pedicel and flower also identified 45 bioactive compounds[33]. However, another study using the same method on the flower of N. pubescens identified 71 bioactive compounds[34]. A total of 22 VOCs were found in the seven species of Nymphaea subgenus Hydrocallis[35]. Gas-liquid chromatography (GLC) based extraction of N. alba leaf identified 53 bioactive compounds[36]. The ultra-performance liquid chromatography-tandem mass spectrometry (UPLC-MS/MS) screen on the flower of N. ‘Blue Bird’ identified 455 metabolites[37]. A large-scale screen on 56 water lily cultivars identified 117 volatile organic compounds (VOCs), in which alkenes and alkanes were the most abundant, followed by ketones and aldehydes[38]. The N. colorata flower releases 11 different volatile molecules, mainly terpenoids, fatty acids, and benzenoids as fragrant molecules, among which methyl decanoate is the major fragrance molecule[9] (Fig. 4).
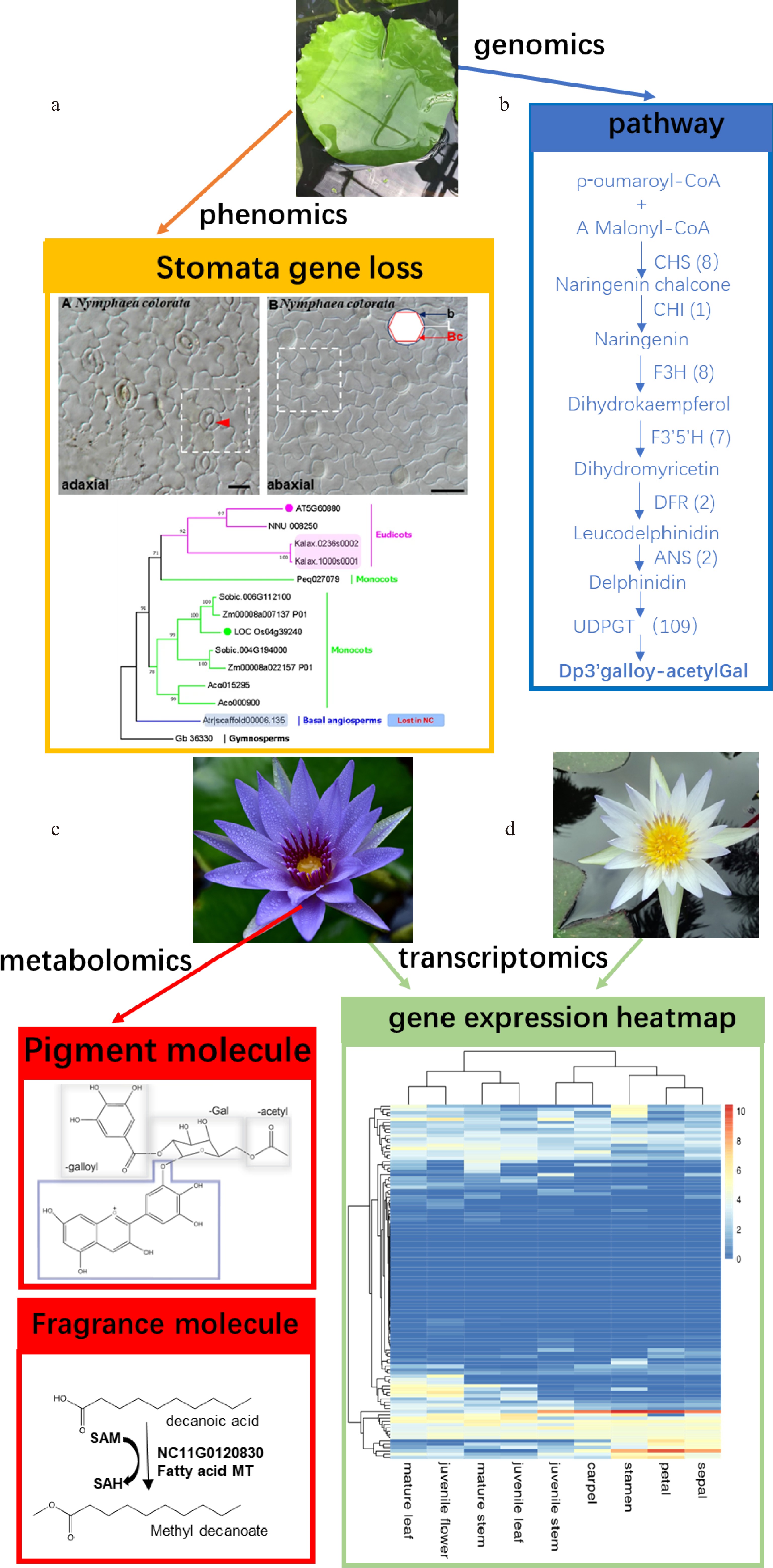
Figure 4.
(a) Phenomics, (b) genomics, (c) metabolomics, and (d) transcriptomics, guided gene screening related to blue anthocyanin in the petal of N. colorata and the stomata development. The genomic and phenomic studies identified loss of genes associated with stomatal development in the lower abaxial side leaf of N. colorata. Genomic, transcriptomic, and metabolomic studies identified floral fragrance-related genes and pathways associated with blue anthocyanin synthesis.
The Victoria genus only includes species V. amazonica, V. cruziana, and the recently identified species V. boliviana[39]. Exploration of floral scent composition using GC-MS could provide a large array of floral VOCs in a series of water lilies. In Victoria cruziana, four VOCs were identified from its flower, in which the VOC composition and emission time are also identified[40].
At the genetic level, transcriptome analyses identified a SABATH gene family member, NC11G0120830, and a terpene synthase gene family member, NC11G0123420, as highly and uniquely expressed in petals and involved in the metabolism of fragrant molecules[9].
Floral colors
-
Nymphaea species display a variety of floral petal colors, including white, red, purple, yellow, blue, etc. A few Nymphaea water lilies, such as N. colorata, N. caerulea, N. 'King of Siam'
, etc., exhibit beautiful blue petals. Flavonoids are the key elements of floral color. Nymphaea species contain a wide variety of flavonoids. For example, a broad sampling of 35 tropical Nymphaea cultivars identified 34 flavonoids[41]. Transcriptome sequencing was conducted on the colorless stage and the fully colored stage of the blue flower cultivar N. 'King of Siam'. Although no reference genome was available for this species, Wu et al.[42] still identified 26,206 unigenes, among which 1,581 genes have differential expression values between the two stages. An in-depth survey revealed that 33 genes with significant expressional levels are responsible for color formation. Finally, the authors validated the expressional changes of seven ANTHOCYANIDIN 3-O GLUCOSYLTRANSFERASE (UA3GT) genes using the qRT-PCR method[42]. N. colorata has very attractive blue petals. Zhang et al.[9] have identified the Dp3′galloylacetylGal as the main anthocyanin for blue coloration in the petals (Fig. 4). The Victoria species display large flowers, showing white petals on the first day, turning pink or red on the second day. Relying on the combination method of high-performance liquid chromatography with photodiode array detection (HPLC-DAD) and ultra-performance liquid chromatography coupled with tandem mass spectrometry (UPLC-MS/MS), Wu et al. identified 14 flavonoids, including 10 flavonols and four anthocyanins, in the petals of two Victoria species[43].
-
The large and starchy seeds of E. ferox are both a nutritious food and a medicine in East Asia. By transcriptomic analyses of different seed stages, Liu et al. found increased expression of P450 and PAL genes in phenylpropanoid metabolism, which might be involved in the maturation of its seeds in aspects of seed size, color, harness, and accumulation of secondary metabolites[13]. Specifically, the authors found the changed expressional pattern of flavonoids during the seed development, involving ABA and SA phytohormones. Candidate genes in regulating the two phytohormones in regulating the flavonoids are identified[44].
-
Some Nymphaea species are leaf vivipary by producing seedlings on the leaves. As an interesting horticultural trait, the leaf vivipary is not only a supplement to breeding programs, but also an opportunity to study stem cell initiation in mature organs. By comparing the transcriptomes of four stages of leaf development between the non-vivipary N. colorata and the vivipary N. micrantha, Su et al. found four potential regulators, including the transcription factors ERF1B, ERF105, RAP2-3, and WRKY22[45]. In the future, through functional studies of genes, such as CRISPR-Cas9 mediated gene knockout and transgenic technologies, it is possible to gain more knowledge of the origin and evolution of vivipary in plants.
-
Nymphaea prolifera is the water lily currently known to produce floral vivipary, an amazing trait called branching flowers. Instead of fertile flowers, N. prolifera usually produces tuberiferous and sterile flowers that act as vegetative organs. N. prolifera can produce two levels and sometimes even three levels of branching flowers, giving rise to dozens and often more than 100 vegetative propagules[46]. This unique trait is, however, still under researched. In the future, its potential in general biology will perhaps show its significance, at least in breeding or maintaining important mutations.
-
A large number of angiosperm crops, including the Allium, Iris, Tulipa, and Begonia, have bulbs. All Nymphaeaceae species contain starchy tuber bulbs, although ranging dramatically in size from one centimeter to a foot. It seems that the bulbs have evolved independently many times during evolution. Nymphaea water lilies develop four major types of bulbs, including Marliac-type rhizome, pineapple-type rhizome, ordorata-type rhizome, and tuberosa-type rhizome. These bulbs are rich in starch and nutrients, making them a good alternative food in some tropical regions. At the genetic level, the FLOWERING LOCUS T (FT) genes are found to be involved in bulb formation in onions[47]. Zhang et al. found a total of five FT genes in the genome of Nymphaea colorata[9]. The five FT genes are clustered into two groups, which are produced by Pi-whole genome duplication[48]. This suggests that the two groups may have different functions, with one group controlling the flowering time and the other controlling the bulb formation. However, due to the very limited research on water lilies, we still do not know which FT gene is responsible for its bulb initiation and development.
-
A few water lily species produce stems that grow flat along the ground called runners. It will root at the node and form a new plant, thereby spread rapidly. The Nymphaea mexicana and the Brasenia schrebri produce runners and become an invasive species in many aquatic environments. The runner is also common in monocot plants such as wild rice and Iris spp., and eudicots such as strawberry and fish mint. Runners could be utilized for plant propagation. Although we now know that the phytohormone GA and the DELLA gene are involved in runner formation[49], the genetics of the runner is completely unknown in water lilies.
-
Hybrid breeding within and across various Nymphaea species is relatively easy and popular. To date, there are hundreds of Nymphaea cultivars available. Different Nymphaea cultivars display various floral colors, floral scents, flowering times, leaf shapes, and bulb sizes. Various traits of water lily populations have been collected for a long time and many of them have been well described and statistically analyzed. Dąbrowska et al. studied nine traits, including leaf length, length of outer petals, and maximum leaf width, of four Nymphaea species[50]. Pan et al. collected 86 Nymphaea cultivars and 45 traits. These traits include floral colors, length of the leafstalk, leaf size, bud shape, and bulb shape. Results show that extensive variations occur in this population, in which floral color is the most highly diverse trait[51].
Xu et al. find that Nymphaea species have an asymmetrical distribution of stomata on the leaf, with normal stomata on the adaxial side and abnormal ones on the abaxial side. Relying on the reference genome sequence of N. colorata, a series of core stomata regulators, including EPF2, AP2C3, MPK6, as well as stomatal polarity regulators BASL and POLAR genes, are absent in its genome, indicating that gene loss is a major innovation in its aquatic adaptation. This study provides an example that gene linkage phenotype analyses could be a trend in future studies[26].
-
Water lilies have some advantageous characteristics as potential model plants for genetic and genomic studies (Fig. 5). All water lilies grow into aquatic herbs, and most Nymphaea water lilies have small sizes, with some species even stretching up to 0.5 m in diameter. Some Nymphaea water lilies (i) have rapid life (around three to four months from seed to seed), and bloom all the year in tropical regions, (ii) have lots of flowers in a single plant and each flower contains ~3,000 fertile seeds. These characteristics make them ideal candidates for model plants. In addition, the release of the genome sequence of N. colorata, together with the genetic transformation pipeline of Nymphaea species[4], all allow Nymphaea water lilies to be ideal model plants for studying the evolution of angiosperms and floral characteristics.

Figure 5.
The life cycle and characteristics of Nymphaea water lily make it a suitable model plant. Genetic transformation through its flowers is available in water lilies, similar to that in Arabidopsis. The numerous seeds in one Nymphaea fruit pod provide a good opportunity for the construction of a mutant library. By laboratory culture of seeds or organs, water lily could be ideal for lab research. Batch culture of water lilies is relatively easy as shown in the illustration.
Cabomba plants also have some excellent traits and growth characteristics, suitable as model plants for studying the origin of angiosperms. For example, C. aquatica and C. caroliniana can be efficiently cultivated in the laboratory[52]. Their flowering can be induced by light, and their small size as an aquatic herb with a smaller than average genome of the ANA grade, all make Cabomba a suitable model plant[52].
Trithuria, a small genus with 12 species from Australia, New Zealand, and India, grows to very a small size (~2 cm in height) and flowers very quickly in the laboratory[53]. Its seeds can be stored conveniently[53]. In summary, Trithuria spp. may provide a good opportunity for both reverse and forward genetic screening.
-
The water lily order contains about 100 species with a relatively wide geographical distribution, and different species possess a large number of different metabolites. In the future, multi-omic tools will be combined in water lily research. Eventually, all water lily genome sequences will be deciphered by relying on long-length or even ultra-long read-length sequencing technology with high accuracy. Based on this, the micro-evolution of all water lilies will be deeply revealed, such as more phenotypically related genes can be mined using genome-wide association studies (GWAS) technology. Using epigenomic technology, we can deepen our understanding of gene regulation. More metabolites can be mined using metabolomics, while the association between genes and phenotypes can be studied by phenomics. In conclusion, more and more technologies will contribute significantly to the understanding and application of water lilies.
From genetics to breeding and industry
-
In the near future, we are confident that all water lily genomes will be decoded (Fig. 6). The pan-genome of water lilies will be established. Genome-wide association studies and genome editing will accelerate the genetics of water lilies. The various genes and pathways responsible for important traits will be uncovered. Afterward, these genes will be employed for intelligent molecular design breeding. Excellent varieties of water lilies will emerge, coupled with large-scale cultivation for application in the water lily industry.
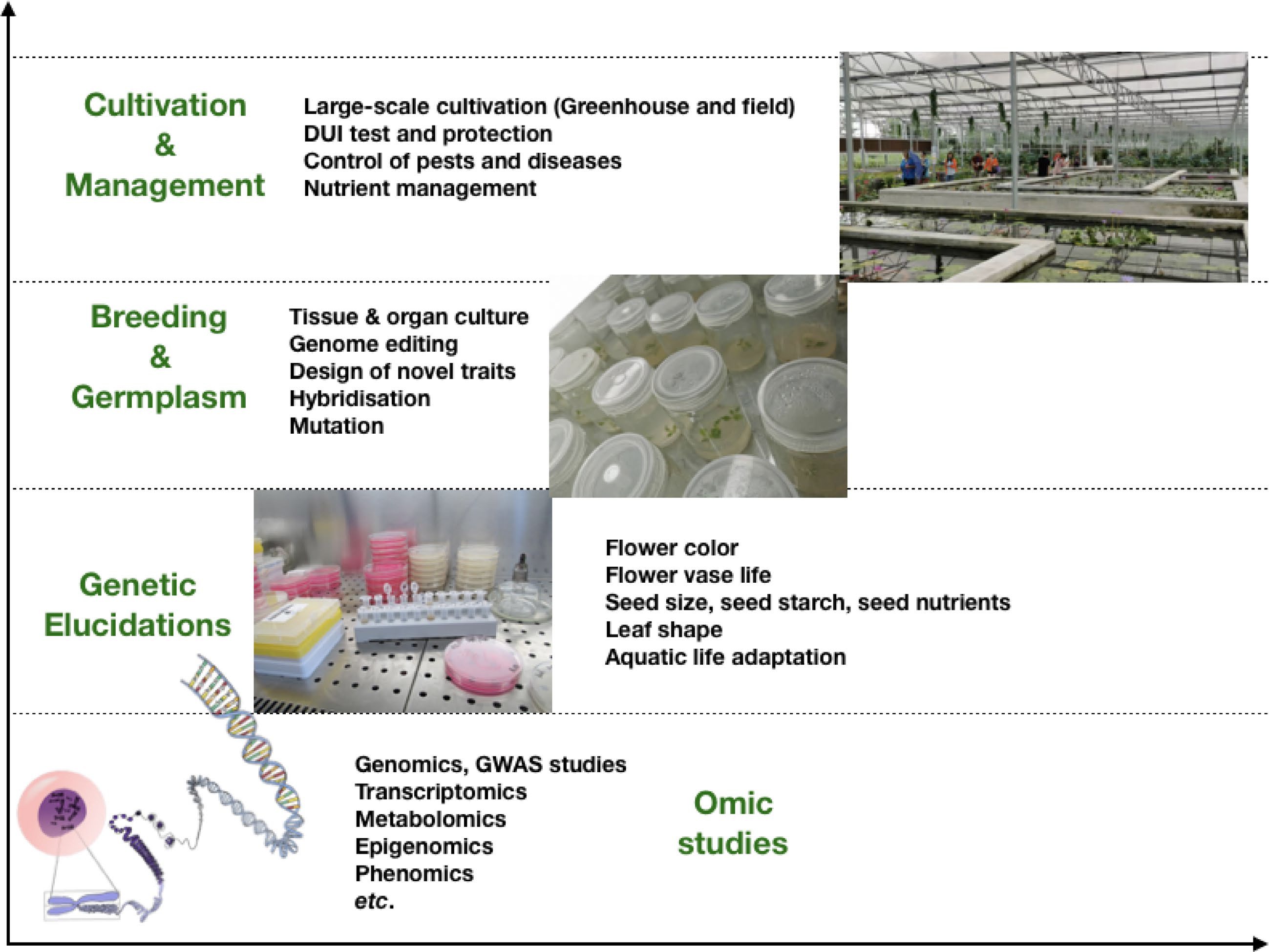
Figure 6.
Future roadmap of water lily research.
The authors acknowledge the funding from the National Natural Science Foundation of China (32172614) and a startup fund from Hainan University.
-
Fei Chen, Zhiqiang Xia, and Liangsheng Zhang are the Editorial Board members of Journal Tropical Plants. They were blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of these Editorial Board members and their research groups.
-
Received 20 June 2022; Accepted 13 December 2022; Published online 30 January 2023
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xiong X, Zhang J, Yang Y, Chen Y, Su Q, et al. 2023. Water lily research: Past, present, and future. Tropical Plants 2:1 doi: 10.48130/TP-2023-0001 

Water lily research: Past, present, and future
- Received: 20 June 2022
- Accepted: 13 December 2022
- Published online: 30 January 2023
Abstract: The water lily order Nymphaeales includes ~100 species and all grow into aquatic herbs. Most of them are not only economic crops but have, for thousands of years, been regarded as cultural symbols, especially in Buddhism and Hinduism. Water lily order forms an early angiosperm branch, occupying vital roles in evolutionary biology. With the advent and rapid development of omic techniques and bioinformatic software, the research on water lily biology has achieved unprecedented success. In this review, we summarized a series of recent and important findings in water lily research. Genomic analyses of water lilies provide insights into details of their evolutionary history and ecological adaptation. The genomes also provide rich resources for genetic studies and molecular breeding. The opening and closing rhythm of flowering is controlled by auxin and the candidate genes are proposed. Part of the genes responsible for floral scents and floral colors in Nymphaea colorata have been studied in-depth for their functions. Metabolomic profiling reveals the anthocyanins responsible for the floral color formation and the volatile organic compounds as floral scent molecules. Phenotypic studies surveyed the diverse traits from flowers to leaves and bulb roots, and some of these studies reveal the link between genes and phenotypes. However, genes responsible for some critical traits such as the growth of water lily plants, such as the stem, bulb formation, and the initiation of vivipary are still unknown. Finally, we propose potential future research of water lilies, including genetics, breeding, and industrialization.
-
Key words:
- Water lily /
- Genomics /
- Transcriptomics /
- Metabolomics /
- Omics /
- Molecular breeding