









-
Classic breeding methods such as mutagenesis or hybridization have been the foundation for plant breeding, where spontaneous mutations and recombination are utilized for plant genetic improvement. While random mutagenesis can create many mutations across a plant species’ genome, many of the mutations are deleterious, resulting in undesirable traits. Similarly, traditional breeding through introgression of desirable traits from the donor plants is a lengthy process, in which a recurrent selection may be required for removing the undesirable traits. In many cases, this can be difficult due to limited recombination and the resulting genetic linkage drag[1]. Additionally, breeding through hybridization relies on genetic diversity, which is increasingly limited due to genetic bottlenecks through years of domestication[2]. Therefore, the intensive labor and high cost of classical breeding methods, as well as the limited supply of novel germplasm, pose challenges for incorporating specific beneficial traits via conventional breeding.
In contrast to undirected random mutagenesis methods, the clustered regularly interspaced short palindromic repeat associated endonuclease 9 (CRISPR/Cas9) system enables precise control over the mutation process[1, 3, 4]. This system takes advantage of the plant natural double-strand break (DSB) repair mechanism, consisting of homologous recombination (HR) for accurate reconstruction of the original sequences or non-homologous end-joining (NHEJ) for insertions or deletions. As a result, beneficial traits can be introduced into elite germplasm within one generation without deleterious alleles, which are frequently observed in traditional mutagenesis and introgression hybridization. Additionally, gene-edited plants without Cas9 or foreign DNA can be obtained by genetic segregation through self-crossing or backcrossing to the wild type[5]. Since the development of the CRISPR/Cas9 system, successful gene editing in multiple plant species have been reported, including agronomic, vegetable, and fruit crops as well as ornamental crops[6, 7].
The ease of programming guide RNA (s) using CRISPR/Cas9 has enabled scientists to enhance favorable characteristics in ornamental crops, including carotenoid accumulation in Ipomoea nil, flower longevity in Petunia hybrida, and flower color in Torenia fournieri[8−10]. However, important aesthetic traits such as leaf variegation remain unexplored. Ornamental crops with attractive variegated characteristics are preferred and popularly used in interior plantscapes. Hence, variegated plants hold a significant market share worldwide and comprise about one-third of the ornamental plants grown commercially[11]. While a variegated-leaf phenotype was induced by CRISPR/Cas9-targeted mutation of a Phytoene Desaturase gene responsible for an albino phenotype, the variegated phenotype was not stable and progressively disappeared over time[12−14].
In Arabidopsis, AtVAR2 or AtFtsH2 codes for a chloroplast FtsH metalloprotease, which is a protein belonging to a large class of ATPases associated with various cellular activities. In higher plants, FtsH mediates multiple functions that are critical for chloroplast biogenesis and maintenance[15, 16]. Thus, a mutation in AtVAR2 would lead to abnormal plastids and a variegated phenotype as seen in the Arabidopsis var2 mutant where white/yellow sections appear in the normally green parts of the mature plant[15, 16]. Thus, var2 would serve as a candidate gene for CRISPR/Cas9 knockout mutation to develop new variegated-leaf breeding materials in diverse species.
While CRISPR-Cas9 gene editing could overcome limitations of classical breeding methods and enable the creation of predicted phenotypes in the gene-edited plants, the successful delivery of Cas9 into plants relies on an efficient plant transformation system. Generally, in vitro transformation through either Agrobacterium infiltration or biolistic bombardment is the most common methodology for generating stable transgenic plants. However, the transformation efficiency could be very low due to the lack of a selection marker that can be easily tracked at different regeneration stages. While genes resistant to antibiotics or herbicides are commonly used as selection markers to restrict the growth of non-transformed tissues, false positive escape plants were often regenerated, and it is difficult to distinguish the transformed tissues with strong transgene expression, which is associated with high editing frequency[17]. Thus, to increase the efficiency of transformation and improve the frequency of gene edited plants, selectable markers have been utilized for monitoring the expression of the transgenes during early callus induction and shoot regeneration, and for in vitro selection of transformed plants and their progenies across generations[17]. Selective markers expressing fluorescent proteins have been used in combination with CRISPR/Cas9 to screen transformed tissues with strong expression of transgenes[18, 19]; and these fluorescent markers were convenient for eliminating seeds carrying the transgenes to select transgene-free gene edited plants[20].
Considering the effectiveness of using fluorescent proteins to non-destructively screen for transgenes in vitro as well as for Cas9-free plants, this study aimed to apply the Cas9 system with a fused neomycin phosphotransferase II and green fluorescent protein (eGFP-NPTII) under Cassava virus promoter for the knockout mutation of the Variegated gene in lettuce (Lactuca sativa L.). Thus, GFP would be monitored throughout plant regeneration from callus to seed and seedling stages, and variegated leaves would be expected in the gene-edited plants. The presence of GFP would also enable efficient removal of the Cas9 insertion via segregation. Lettuce serves as an excellent model due to its easy regeneration in vitro through indirect organogenesis, and phenotypic evaluation of variegated leaves would be evident since lettuce is a leafy vegetable and is gaining popularity in urban agriculture.
-
Our expression vector was reconstructed based on several previously published plasmids. The plasmid pDe-CAS9 containing an Arabidopsis codon-optimized Cas9 driven by parsley ubiquitin promoter and a ccdB cassette for Gateway cloning of guide RNAs was developed by Fauser et al (2014)[21]. The NptII cassette was repositioned immediately adjacent to the left T-DNA border in our previous study[17]. The BsaI site in the pDe-Cas9 backbone was replaced by a corresponding backbone fragment without BsaI site from the pYLCRSIPR/Cas9P35S-H through KasI and BsiWI-HF digestion and ligation[22]. The NptII cassette was replaced by the fused eGFP-NPT under a double-enhanced CsVMV (dCsVMV) promoter through a Hind III digestion and ligation. The dCsVMV::eGFP-NPT was PCR amplified with primer set CsVMV-nptgfp-F and CsVMV-nptgfp-R (Supplemental Table S1) from a plasmid pDEAT[23]. The ccdb cassette was then replaced by the ProAtU6-26-tRNA-2×BsaI-scaffold RNA-tRNA-2×BsaI-scaffold RNA polydT cassette through a AvrII digestion and ligation (see Supplemental Data Set 1 for sequence information, synthesized by GeneUniversal Inc, Newark, DE, USA), thus resulting in our final PHN-SpCas9 CRISPR vector with 4×BsaI restriction site (PHN-SpCas9-4×BsaI-GFP) for convenient sgRNAs cloning (Fig. 1a).
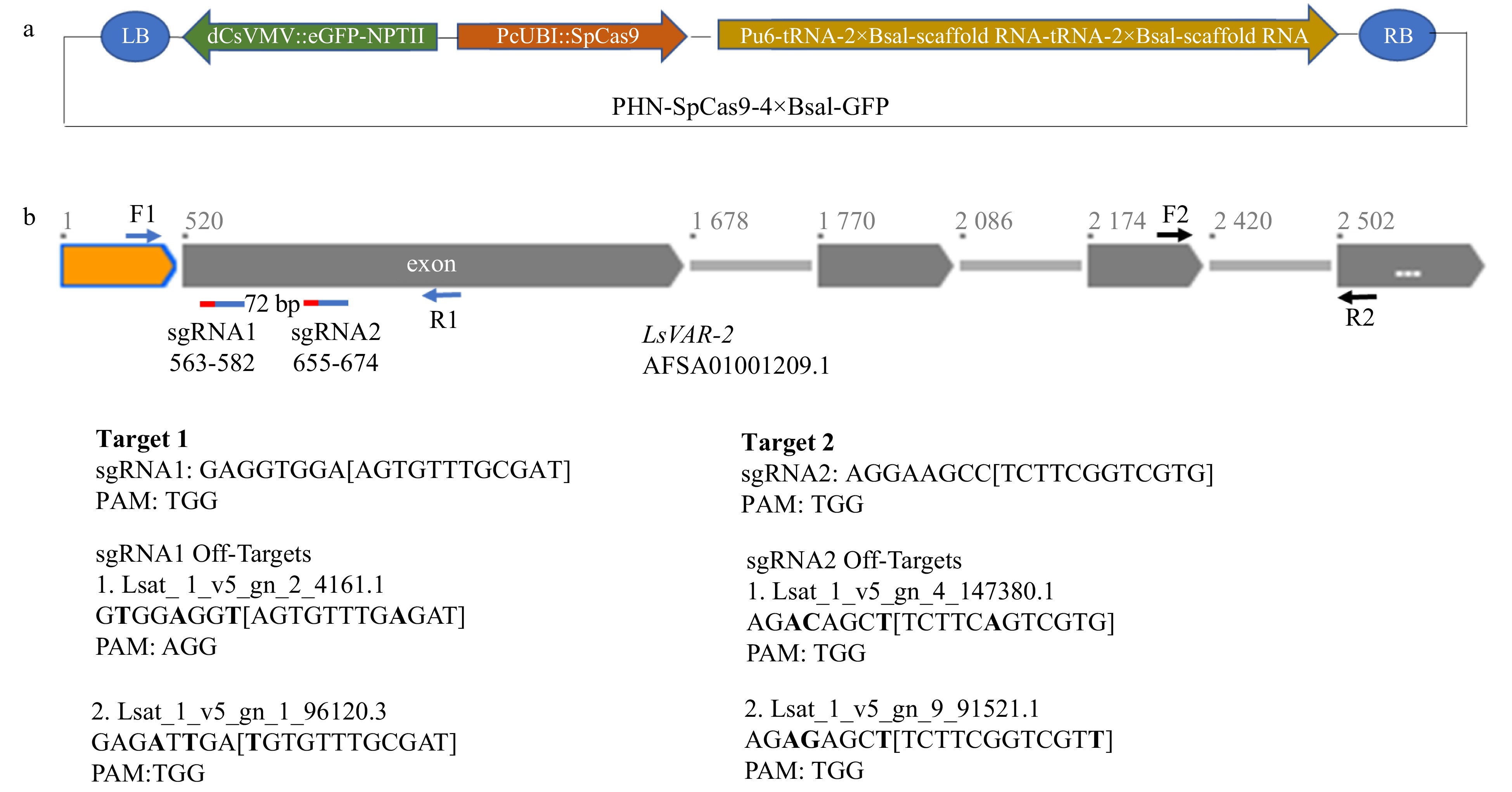
Figure 1. Schematic diagram of CRISPR/Cas9 vector design and LsVAR2 gene. (a) The PHN-SpCas9-4×BsaI-GFP vector contains a codon-optimized SpCas9 for Arabidopsis under parsley ubiquitin (PcUbi) promoter, a fused eGFP-NPTII under a dCsVMV promoter, and a sgRNA cloning cassette with two separate 2×BsaI sites, tRNAs and scaffold RNAs under the AtU6-26 promoter. (b) Schematic structure of LsVAR2 gene, two on-target sgRNAs and their potential off-targets. Orange box: 519 bp sequence upstream the first exon; gray boxes: exons; gray lines: introns. The positions of reverse complementary of two sgRNAs were indicated with two red-blue lines under the first exon with a 72 bp span between two sgRNAs. Red lines indicate PAM region of sgRNAs (blue lines), and the exact positions for both sgRNA were also shown as their numbers. The positions of primers F1/R1 for sequencing were indicated by blue arrows, and the positions of primers F2/R2 for qPCR were indicated by black arrows. Sequences of two guide RNAs and two potential off-targets were shown, and the nucleotide differences between the on-target and off-target sgRNAs were indicated in bold.
Arabidopsis VAR2 (AT2G30950) gene was used to BLAST against the whole genome and transcriptomic and genomic sequence database of L. sativa at NCBI; LsVAR2 genomic sequence (part of AFSA01001209.1) and cDNA sequence (JI576939) was used for sgRNAs selection[24]. CRISPR-P and CRISPOR were used to select two sgRNAs on the same exon using SpCas9 with Protospacer Adjacent Motif (PAM) ‘NGG’ and potential off-targeting sites for each sgRNA were also identified using CRISPR-P[25, 26] (Fig. 1b). For cloning the both sgRNAs, two sets of oligos with sticky ends that match each BsaI restriction site in PHN-SpCas9-4×BsaI were synthesized by Thermo Fisher Scientific (Waltham, MA, USA) (Supplemental Table S3). Oligos for sgRNA1 and sgRNA2 were separately mixed at equal molar amount and denatured at 98 °C on a SimpliAmpTM Thermal Cycler (Thermo Fisher Scientific, USA) for 30 min prior to being annealed at room temperature for 45−60 min. The annealed sgRNA1 and sgRNA2 were then ligated to the BsaI-linearized PHN-SpCas9-4×BsaI-GFP vector.
Plant materials and Agrobacterium transformation
-
Seeds of lettuce (L. sativa cv. Salinas) were obtained from UC-Davis and stored in sealed containers at 15 °C until used. Seeds were surface-sterilized with 70% alcohol for 2 min, followed by 1.3% hypochlorite solution for 10 min; seeds were washed with sterile double-distilled water 5−8 times prior to germinating them on MS (Murashige and Skoog) media with 3% sucrose and 0.8% agar at pH 5.8. Genetic transformation and regeneration were modified from the protocol developed by Michelmore and Marsh[27]. In brief, 12−14 days after germination, cotyledons were cut and dipped into Agrobacterium strain EHA105 harboring the plasmid PHN-SpCas9-VAR2-GFP for 10 min; the infected cotyledon was subsequently cultured on co-cultivation media (MS + 3% sucrose) in the dark for 2 days, before transferring to shooting media (MS + 2 mg/L 6-benzylaminopurine + 0.1 mg/L NAA + 100 mg/L Kanamycin and Timentin). Rooting of newly regenerated shoots was carried out on a half-strength MS medium containing 100 mg/L kanamycin but with no supplemental hormones. GFP fluorescence was detected with a fluorescent stereomicroscope (Leica, German) and images were captured using Nikon D800 digital camera attached to the microscope. After in vitro rooting of regenerated shoots, they were potted in soil and kept at constant room temperature with 16 h supplemented incandescent light for two weeks before repotting in 1-gallon pots to grow in the greenhouse. T1 seeds were collected for genotyping and phenotypic evaluation.
DNA sequencing for detection of gene editing
-
T1 seeds were germinated on filter paper moistened with distilled water and fully expanded cotyledons were used for DNA extraction using Edwards’s Buffer[28]. PCR products of primers flanking the targeted VAR2 gene and off-target sites were sent to Genewiz, Inc (South Plainfield, NJ, USA) for Sanger sequencing. Sequencing results were aligned using EMBL-EBI and Synthego Performance Analysis (ICE Analysis. 2019. v2.0. Synthego) was used for editing analysis[29]. For examining the de novo editing, we have sequenced the individual cotyledon from the same seedling with variegated phenotype (see the main text); additionally the green and albino tissues from the same leaf containing T-DNA were also included for PCR sequencing to detect the de novo editing (see the main text).
Gene expression analysis
-
For gene expression analysis, two independent gene-edited lines that were confirmed by Sanger sequencing and one WT line were used. Eight seedlings were pooled as one biological replicate for each phenotype: WT, variegated seedlings, and albino seedlings. Total RNA was extracted from cotyledons using RNAzol RT kit as described in the instruction manual (Molecular Research Center, Inc. USA)[30] and was subsequently treated with TURBO DNAase I (Thermo Fisher Scientific, USA) to remove genomic DNAs. The QuantiTect Reverse Transcription Kit was used for cDNA synthesis with 1 µg of total RNA (Qiagen, USA). Three biological replicates and two technical replicates were performed on CFX96 Real-Time System (BIO-RAD, USA) with SYBR Green dye master mix for real-time PCR assays (Thermo Fisher Scientific, USA). All primers used in the experiment are listed in Supplemental Table S3.
Phylogenetic analysis
-
Protein sequences of Arabidopsis FtsH family were retrieved from Tair (www.Arabidopsis.org) and were used to identify lettuce FtsH homologs by blastp against the lettuce protein database at NCBI[24]. A total of 22 amino acid sequences from Arabidopsis and lettuce were used to build a phylogenetic tree, and MEGA X was used for phylogenetic tree construction using the maximum-likelihood method with a 1,000 bootstrap value after sequence alignment using Clustal W[31]. iTOL v4 was used to visualize the tree generated from MEGA X[32].
Morphological and physiological measurements
-
Leaf thickness was measured by cutting the fifth leaf of 30-day old plants into a fine strip avoiding the vein, and the leaf strip image was captured using a stereomicroscope and measured with the Image J software. Ten plants of each line were sampled for each measurement. For the specific leaf weight (SLW) measurement, a puncher (diameter = 0.3 cm) was used to collect five discs from the fifth to the seventh leaves of each plant. Three different plants were used for each gene-edited line. The discs were saturated in deionized water to minimize the water loss during sample harvest and measurement, and the saturation weight was recorded. Afterward, the discs were placed in an oven at 80 °C for two days and were weighed again for dry mass.
Chlorophyll content was measured using five leaf-discs from the fifth to seventh leaves on three different plants for each gene-edited line. The discs were immersed in 15 mL of 95% ethanol in a glass tube. The tubes were shaken at room temperature at 150 rpm in the dark for five hours. Afterwards, the OD values were measured using a Nanodrop One UV-Vis Spectrophotometer (Thermo Fisher Scientific USA). The contents of chlorophyll a (Chla) and chlorophyll b (Chlb) were calculated with the equation of Arnon,
Chla = (12.72A663 −2.59A645) × v/w × 1,000
Chlb = (22.88A645 − 4.67A663) × v/w × 1,000
where 'v' is the volume of alcohol (15 mL) and 'w' is the weight of discs in each tube[33].
Chlorophyll a fluorescence transients were measured using the sixth leaf of 17 wild type and 17 gene-edited plants with a portable pulse-modulated fluorometer (OS30p, Optisciences, UK). Each leaf was measured twice for calculating mean values. The plants were exposed to a saturated light pulse of 3,500 µmol m−2 s−1 light intensity at pre-dawn and the resulting fluorescence measured from 0 to 1 second. Various fluorescence parameters corresponded to the OJIP test such as energy transfer efficiencies (ϕD0, ϕE0, and ψ0), overall performance index (PIABS) of photosystem II (PSII) and specific energy fluxes in PSII (DI0/RC, TR0/RC, and ET0/RC) were calculated as described by Strasser et al.[34, 35].
Statistical analysis
-
All data obtained were statistically analyzed using student’s t-test with an R package and package 'pcr' was used for real-time quantitative PCR (qPCR) analysis[36] (See Supplemental Data Set 3). Graphs were generated by both R Studio and Microsoft Excel.
-
The modified construct enabled us to track plant cells and regenerated seedlings through detecting the green fluorescence at different stages (Fig. 2). During plant regeneration, GFP was visually detected during the callus induction phase, indicating the success of Agrobacterium-mediated transformation. The stably expressed GFP could be observed in some of the calli, which were preferentially selected for further plant regeneration, significantly reducing the time and resources required (Fig. 2a, d). In addition, GFP fluorescence could also be observed in the T1seeds, which expedited the screening of the seeds containing T-DNA (Fig. 2b, e), and the seeds exhibiting weak GFP fluorescence may be further screened out by checking the germinating seedlings where GFP can be easily detected (Fig. 2c, f).

Figure 2. GFP detection after plant transformation using modified CRISPR/Cas9 vector. (a) Strong GFP fluorescence in the callus of lettuce after Agrobacterium infiltration and during plant regeneration and selection; (b) GFP fluorescence was observed in seeds from T1 plants; (c) GFP fluorescence in T1 mutant seedlings; T-DNA free seeds and seedlings segregated from the same genetic lines in (b) and (c) exhibited no GFP fluorescence. (d−f) same images as (a−c) but taken under white light.
The application of this fused eGFP-NPTII allowed us to avoid any negative positional effects on transgene expression due to T-DNA insertion, which are generally observed with sole antibiotic selection[37−39]. In this case, the plant tissues with strong expression of Cas9 and sgRNAs can be monitored and selected, and a high frequency of editing may be expected. Tagging of Cas9 with a reporter gene has been reported previously[40], and the mechanism of improving gene editing efficiency through selecting regenerated tissues with strong Cas9 expression is similar to our design, but a tagged protein may also pose a risk of reduction of Cas9 activity due to potential changes in the protein conformation. Instead, we separately expressed the eGFP-NPTII and the Cas9 genes, anticipating that both will be expressed from the successfully integrated vector (Fig. 1b). As a result, we have generated 25 transgenic plants, all of which were confirmed to contain the T-DNA. This result suggested that dual selection with GFP and antibiotics could avoid the false positive escape plants. To estimate the editing efficiency, we have performed Sanger sequencing of 8 randomly selected seeds with highly expressed GFP for each of the transgenic line. As shown in the Supplemental Data Set 2 and Supplemental Tables S1 and S2, the editing efficiency reached 40%, and mixture of mutations were identified at both sgRNAs loci. Given the limited capacity of Sanger sequencing and possible biased amplification from the pooled seeds, this editing efficiency could be underestimated, yet it is still comparable to our previous editing efficiency[17]. In addition to facilitating plant transformation and gene editing, the application of eGFP can be used to easily screen T-DNA free seeds or seedlings. For example, the seedlings with the gene edited mutations (albino cotyledons in this case) and lack of fluorescence indicated homozygous mutants without T-DNA insertion (Fig. 2c, f).
Targeted mutation of LsVAR2 by CRISPR/Cas9
-
Two guide RNAs (sgRNAs) approximately 72 base pairs apart on exon 1 of lettuce LsVAR2 were selected for knockout mutation using the CRISPR/Cas9 system (Fig. 1b). The two sgRNAs were separated by tRNA sequence for multiplex editing (Fig. 1a)[41]. T1 segregated seedlings from heterozygous plants showed distinct cotyledon coloration phenotypes: the green cotyledons without (Fig. 3c−e, i) or with white speckles/stripes (Fig. 3e, g, i, k), and the albino cotyledons without (Fig. 3e, f, i, j) or with green speckles (Fig. 3e, h, i, l). Sequence analysis suggested that there was a homozygous 53 bp deletion, a homozygous 8 bp deletion in albino seedlings segregated from the genetic lines 26-VAR-1-F_R and 25-VAR-1-F, respectively (Supplemental Fig. S1a), which were demonstrated by clean sequencing chromatogram and validated by the ICE Analysis (Supplemental Fig. S1b, c, f). By contrast, the sequencing noise from the chromatogram graph and ICE analysis revealed that heterozygous mutations existed in these variegated seedlings (Supplemental Fig. S1d, e, g). Accordingly, the 53 bp and 8 bp deletions caused large changes in amino acids and resulted in premature stop codons in both albino 1 and albino 2 (Supplemental Fig. S1a).
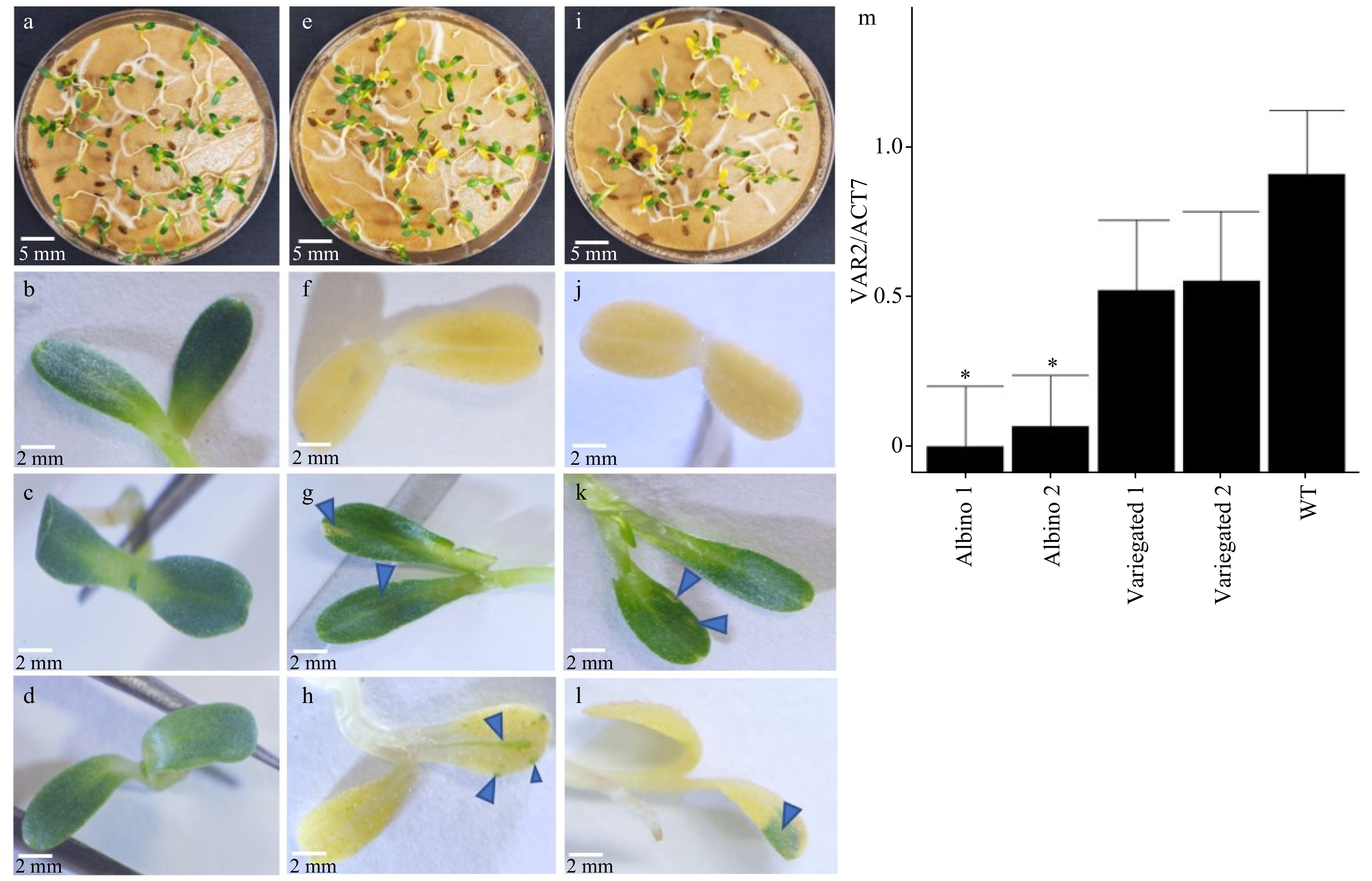
Figure 3. Different variegation phenotypes putatively created by de novo gene editing and transcription of LsVar2. (a) Germinated seedlings and cotyledon (b) of wild type lettuce cv. Salinas. (c) Green cotyledon without white speckles, (e) germinated seedlings, (f) albino cotyledon with no green speckles, (g) green cotyledon with variegated white speckles, (h) albino cotyledon with variegated green spots from heterozygous LsVAR2 gene edited T1 line 26-VAR-1-F_R. Arrows in (g) and (h) indicate variegated white speckles in the green cotyledon and green speckles in the albino cotyledon, respectively. (d) Green cotyledon without white speckles, (i) germinated seedlings, (j) albino cotyledon with no green speckles, (k) green cotyledon with variegated white speckles, (l) albino cotyledon with variegated green spots from heterozygous LsVAR2 gene edited T1 line 25-VAR-1-F. Arrows in (k) and (l) indicate variegated white speckles in the green cotyledon and green speckles in the albino cotyledon, respectively. (m) Relative gene expression in variegated and albino seedlings from 26-VAR-1-F_R (Albino 1) and 25-VAR-1-F (Albino 2) lines, and in wild type seedlings (WT). The gene expression was expressed as a normalized value to the wild type. All images were taken at 7 days after germination.
Unlike the Arabidopsis var2 mutant having a variegated phenotype, most homozygous lettuce mutants exhibited albino phenotype in the seedlings and could not survive in the absence of exogenous nutrients from MS media (Fig. 2f), however despite that, greenish speckles were observed in some albino seedlings (Fig. 4e, h). This distinct phenotype between Arabidopsis and lettuce var2 mutants could be attributed to redundant functions of Arabidopsis FtsH members. There are 12 AtFtsH family members in Arabidopsis, and homozygous mutation of AtFtsH2/AtVAR2 in Arabidopsis would lead to a variegated phenotype, but AtFtsH2/AtVAR2 and AtFtsH8 double mutants are unable to develop normal chloroplasts and caused an albino phenotype[42], suggesting the functional redundancy between AtFtsH2/AtVAR2 and AtFtsH8. The lettuce genome has 10 FtsH members, where LsVAR2 is closely related to LsFtsH6 (Supplemental Fig. S2b). However, LsVAR2 is closely related to both AtFtsH2 and AtFtsH8, while LsFtsH6 is a homolog of Arabidopsis AtFtsH6 (Supplemental Fig. S2a, b). Because the LsVAR2 is the only FtSH member that exhibits high similarity to both AtFtsH2 and AtFtsH8 in lettuce (Supplemental Fig. S2a, b), homozygous LsVAR2 mutations created by Cas9 in lettuce prevented normal development of the chloroplasts as observed in AtFtsH2 and AtFtsH8 double mutants.

Figure 4. Phenotypes of LsVAR-2 adult mutant plants. (a) Different variegated patterns on young leaves of 3 week-old mutant and WT plants. (b) Variegated phenotype in three 6 week-old mutant plants.
Previous studies have indicated that CRISPR/Cas nucleases can cut DNA at off-target sites that share similarity with the targeted region[43, 44]. To check whether there are any off-target edits in our T1 plants, we picked two off-target sequences with the highest similarity to each of our on-target sgRNA sequences for sequencing (Fig. 1b). Our PCR sequencing results showed that no mutation was detected at the off-target sites in the gene-edited plants in comparison to the wild type (Supplemental Fig. S3a−d). The results are consistent with recent reports showing high precision of CRISPR/Cas9 when applied in plants[45, 46].
Next, we examined whether the transcription of LsVAR2 was altered by these mutations. As expected, the transcript level of LsVAR2 in the homozygous albino seedlings was significantly lower than in the wild type and heterozygous mutants, suggesting that LsVAR2 transcription was indeed affected in the mutants (Fig. 3m). The remaining transcription in albino seedlings could be caused by the greenish speckles observed in some pooled albino seedlings (Fig. 3h, l). Surprisingly, a comparable level of LsVAR2 transcript were also detected in variegated seedlings and wild type of seedlings. The major parts of variegated seedlings are green with tiny white speckles (Fig. 3g, k), indicating LsVAR2 exhibited normal transcription in the majority of these cells. The normal transcription of LsVAR2 in these cells was probably caused by an enhanced transcription of the functional alleles to compensate the loss of function in the mutated allele.
De novo editing by CRISPR/Cas9 in variegated plants
-
Interestingly, lettuce plants heterozygous for the edited var2 mutant showed a variegated phenotype that exhibited great variation from a few white speckles to white stripes on the cotyledons of germinating seedlings (Fig. 3g, k) and this phenotype intensified as the plant matured (Fig. 4a, b). The variegation of the heterozygous plant leaves may be derived from continuous de novo editing by CRISPR/Cas9 that is stably expressed in transgenic T1 plants. Indeed, evidence of the CRISPR/Cas9 system re-editing events from T0 to T1 have been recorded in lettuce and watermelon, and the de novo edited mutations by CRISPR/Cas9 also happened during meristem induction in multiple plant species[17, 47, 48]. Our sequencing results revealed that different mutations existed between variegated and normal green cotyledons from the same seedling (Fig. 5a−c). The green cotyledon had an editing efficiency of 16% while the editing efficiency in the variegated cotyledon increased to 42% (Fig. 5b). By contrast, the wild type sequence accounts for 20% of the PCR sequencing detection in the green cotyledon, while the predominant (25%) edited mutations in the variegated cotyledons are 1bp deletion at the sgRNA2, and only 5% contribution came from the wild type sequence (Fig. 5b, c). Additionally, we have sequenced the albino/variegated tissue and green tissue from the same leaf (Supplemental Fig. S4a), and the sequencing results indicated that green leaf tissues contained different mutations compared to the albino/variegated tissues (Supplemental Fig. S4b). The wild type sequence accounted for 40% sequencing contribution in the green tissues, compared to 32% wild type sequence contribution in the albino/variegated tissues (Supplemental Fig. S4b). In addition, an 11% of 1bp deletion contribution at sgRNA2 only occurred in the variegated tissue (Supplemental Fig. S4b). All sequencing results indicated that active de novo editing existed in the seedlings containing T-DNA. Thus, the white speckles or striped sections were likely caused by de novo homozygous mutation in those cells. The variegation phenotype intensified as the lettuce matured in the greenhouse, which may be due to the increase in temperature that positively affects Cas9 efficiency[49−51]. Therefore, it is possible that the continuous editing of Cas9 at room temperature during the cotyledon developmental stage was not as strong as in the matured plant in the greenhouse.
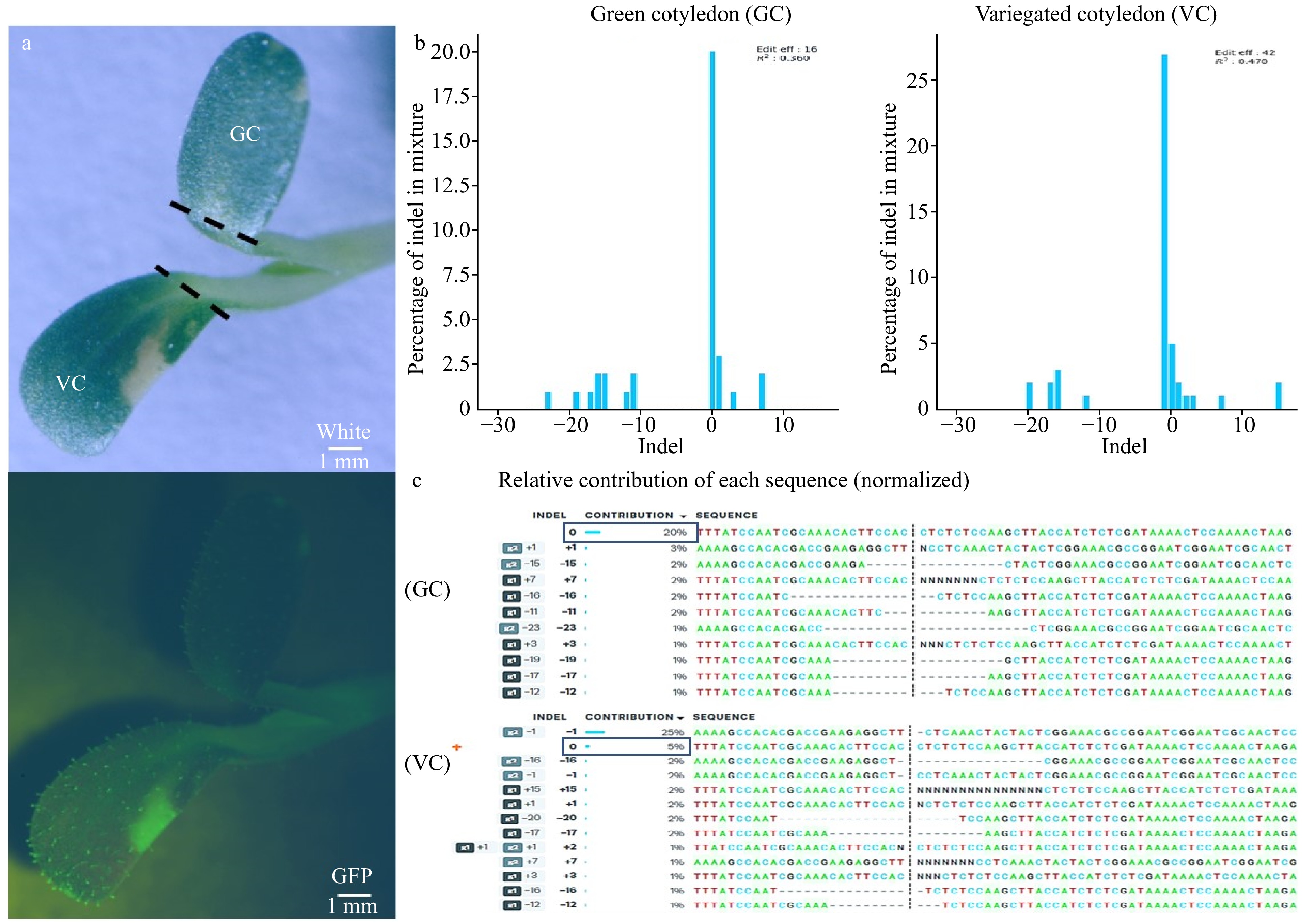
Figure 5. Phenotypic difference caused by de novo gene editing in cotyledon containing Cas9. (a) Green cotyledon (GC) and variegated cotyledon (VC) cut (---) from the same seedling under white light (top), GFP (bottom). (b) The percentage of indel mixture indicated that green cotyledon only had an editing efficiency of 16% while the editing efficiency in variegated cotyledon increased to 42%. (c) Relative contribution of each sequence showed green cotyledon had 20% of WT sequence contribution while the variegated cotyledon WT contribution decreased to 5%. g1: sgRNA1, g2: sgRNA2.
Physiological and growth defects caused by CRISPR/Cas9 induced mutations of LsVAR2
-
Given that LsVAR2 functions in the regulation of chloroplast development, we measured how the dysfunctional var2 mutants affect the photochemical quantum yield of PSII photochemistry and the electron transport activity using OJIP test parameters. The OJIP tests are used for quantitative analysis of the fluorescence transients, and are often used to explain the stepwise flow of energy through PSII at the reaction center (RC) and PSII cross section (CS) levels[34]. Our results revealed a notable increase in the PSII energy dissipation flux (DI0) at both the reaction center and cross section levels in the variegated plants (Fig. 6), resulting in a higher value of DI0/RC. The higher value of DI0/RC indicated low efficiency in trapping energy at PSII reaction centers, suggesting that energy cannot be efficiently transferred upstream of PSII due to the damage in the chloroplast in the lettuce var2 mutants[52]. Consequently, the performance index for energy conservation from photons absorbed by PSII antenna to the reduction of the secondary quinone (QB) and to the reduction of PSI acceptors (PIABS and PITotal) decreased by 30% and 35% respectively in the variegated plants[34]. The alternation in energy transfer and conservation due to the dysfunction in VAR2 may cause lower photosynthetic efficiency, which further explains the reduction in plant development of lettuce var2 mutants (Fig. 7a, d).

Figure 6. OJIP test parameters of lettuce mutant (var2) induced by CRISPR/Cas9 and wild type. PSII energy dissipation flux increased while the performance index for energy conservation decreased in the mutant when compared to the wild type.
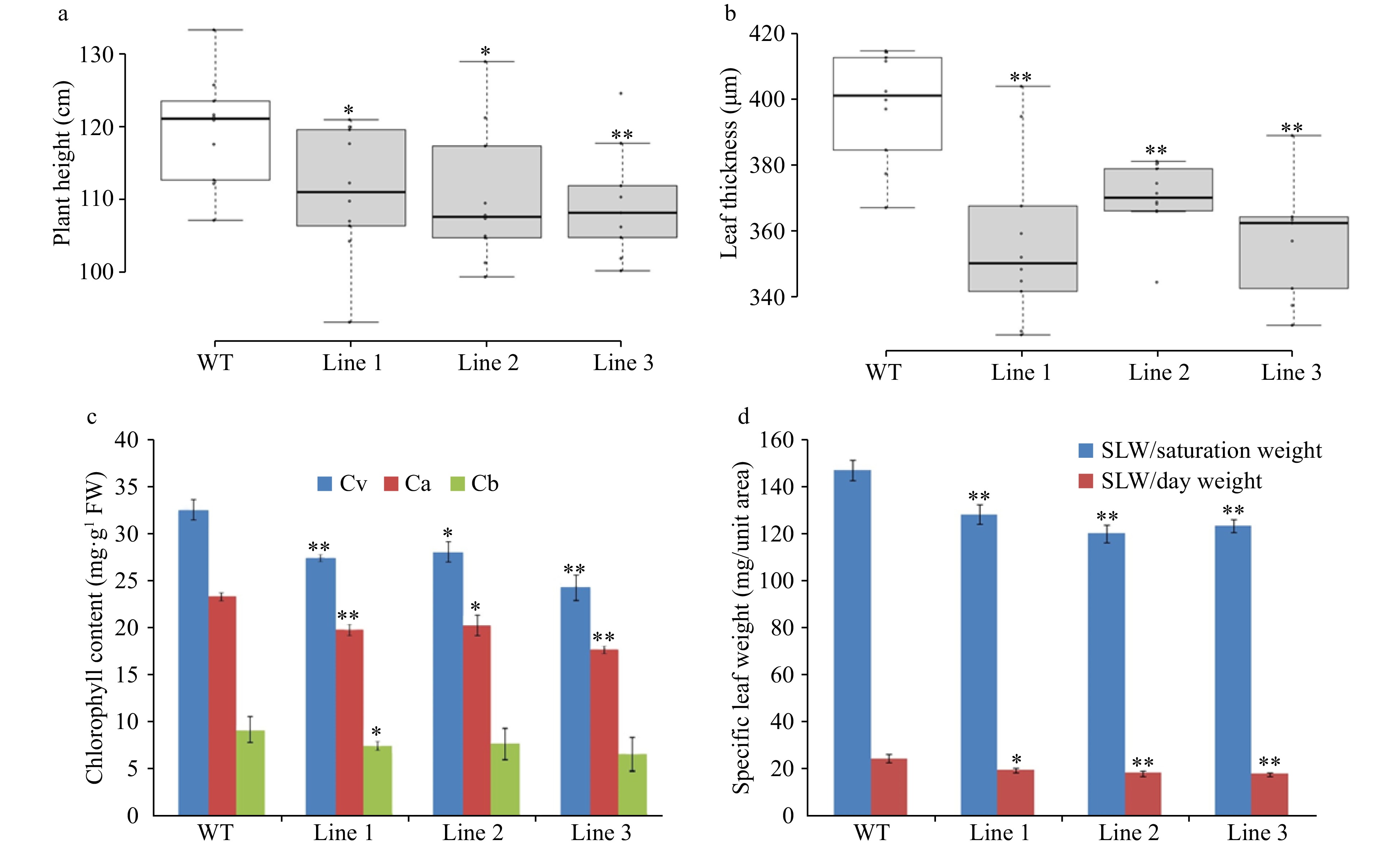
Figure 7. Morphological and physiological measurements of LsVAR2 mutants by CRISPR/Cas9. (a) Plant height; (b) Leaf thickness; (c) Chlorophyll content; (d) Specific leaf weight (SLW). The asterisks denote a statistically significant difference at p < 0.05 between the mutants and wild type.
In addition to the measurement of chlorophyll fluorescence transients, chlorophyll was extracted from variegated plants in three different gene-edited lines. All three lines showed a significant decrease in total chlorophyll content, most notably in chlorophyll a content (Fig. 7c). The result aligned with OJIP parameters, where the productivity of photosynthesis was reduced due to the deficiency of chloroplasts from LsVAR2 mutations. The reduction of photosynthesis was further validated through specific leaf weight measurement where the specific leaf weights of gene edited lines were reduced by 22%, 27%, and 24% in three independent lines, respectively (Fig. 7d). Similar to results for specific leaf weight measurements, there were significant decreases in plant height and leaf thickness when compared to the wild type (Fig. 7a, b). These results indicate dysfunctional LsVAR2 induced by CRISPR/Cas9 caused significant developmental defects in lettuce.
-
In this study, we demonstrated the improved efficiency of a modified CRISPR/Cas9 construct through minimizing T-DNA positional effects and simplifying the process of transgene selection. Using this modified construct, we have efficiently edited LsVAR2 in lettuce, resulting in an albino phenotype in the homozygous plants. Variegated phenotypes were observed in plants with stably expressed Cas9, and sequencing results revealed de novo editing of CRISPR/Cas9 in different parts of the leaves causing disparate albino sections and an overall variegated appearance. The results in this study provided further insight into the broad applicability of CRISPR/Cas9 for creating plants with novel phenotypes for faster plant production or marketing.
Availability of data and material
-
All data generated or analyzed during this study are included in this published article (and its supplementary information files).
Code availability
-
R codes are in supplementary data.
Key Message
-
A modified CIRSPR/Cas9 vector increased efficiency in selection of transgene and Cas9-free progenies of lsvar2 mutants, which demonstrated a new avenue to create novel breeding materials for horticultural plants.
- This project has been sponsored by the USDA-NIFA GRANT12683186
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Sequence analysis of lsvar2 from two mutants created by CRISPR/Cas9.
- Supplemental Fig. S2 Phylogenetic analysis of Arabidopsis and lettuce FtsH genes.
- Supplemental Fig. S3 Sequencing results for detecting off-targeting induced by CRISPR/Cas9.
- Supplemental Fig. S4 De novo gene editing in lettuce leaf.
- Supplemental Table S1 Gene edited percentage based on T1 lines with highly expressed GFP.
- Supplemental Table S2 Summary of 25 independent gene edited lines.
- Supplemental Table S3 Primers used in this study.
- Supplemental Data Set 1 Sequence of AtProAtU6-26-tRNA-2×BsaI-scaffold RNA-tRNA-2×BsaI-scaffold RNA polydT.
- Supplemental Data Set 2 Sequencing data of 25 independent transgenic lines.
- Supplemental Data Set 3 Code used for Real-time PCR analysis .
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Nguyen CD, Li J, Mou B, Gong H, Huo H. 2021. A case study of using an efficient CRISPR/Cas9 system to develop variegated lettuce. Vegetable Research 1: 4 doi: 10.48130/VR-2021-0004

A case study of using an efficient CRISPR/Cas9 system to develop variegated lettuce
- Received: 22 December 2020
- Accepted: 10 May 2021
- Published online: 25 May 2021
Abstract: The clustered, regularly interspaced, short palindromic repeat associated endonuclease 9 (CRISPR/Cas9) system has emerged as a powerful approach for precision breeding to create plants with desirable traits. However, the CRISPR/Cas9 system relies heavily on an efficient plant transformation system that is usually time-consuming and costly. Here, we have constructed a CRISPR-Cas9 vector with neomycin phosphotransferase II and green fluorescent protein (eGFP-NPTII), where the high expression of GFP during plant regeneration allowed us to minimize the positional effect on T-DNA expression and facilitate screening T-DNA-free mutants. Successful gene editing using CRISPR/Cas9 has been illustrated in different plant species, but an important aesthetic characteristic of leaf variegation remained unexplored. With the newly designed construct, we have targeted the variegation gene LsVAR2 in lettuce. Our results indicated that LsVAR2 is closely related to both AtFtsH2 and AtFtsH8, in which homozygous mutations lead to an albino phenotype while a variegated phenotype was induced by CRISPR/Cas9 de novo gene editing. In conclusion, the unique design of our CRISPR/Cas9 construct could efficiently edit the target gene and ease the screening of non-TDNA mutants through detecting GFP signals during plant regeneration and progeny segregation. Additionally, the success of gene-editing of LsVAR2 in lettuce demonstrates proof in this method to develop novel plant breeding materials for valuable horticultural plant species.
-
Key words:
- CRISPR/Cas9 /
- Gene Editing /
- Lettuce /
- Leaf Variegation /
- eGFP-NPT