









-
Scutellaria baicalensis Georgi, a plant in the Lamiaceae family, is primarily valued for its medicinal roots[1]. This plant exhibits significant anti-cancer, antioxidant, and anti-inflammatory properties[2−5], with flavonoids identified as the primary effective compounds[6]. S. baicalensis has been shown to inhibit the replication of SARS-CoV-2 virus and play an important role in the treatment of the COVID-19[7]. Research further suggests that regulating gibberellin (GA) metabolism could influence flavonoids biosynthesis in the plant[8].
The GRAS gene family is a specific transcription factors in plants, named after the three initially discovered members: Gibberellin Insensitive (GAI), Gal Repressor (RGA), and Scarecrow (SCR)[9]. These proteins are characterized by a conserved GRAS domain at the C-terminus, which includes an α-helical cap and α/β core subdomains[10,11]. The highly variable N-terminus determines the specificity of its genetic functions[12]. GRAS proteins are essential for a variety of plant growth processes, including stress responses, plant signaling, and axillary bud and root growth[13−16]. Carboxylesterase (CXE) is a key enzyme in the hydrolytic ester bond reversal reaction, which can catalyze ester and amide compounds[17]. Structurally, CXE has a conserved core structure composed of eight β lamellar α helical and cyclic structures, categorizing it within the α/β lamellar hydrolase superfamily[18]. CXE family members play critical roles in plant growth, development and responses to biological stress. GID1, identified from CXE, is a soluble GA receptor with an extremely high affinity for biologically active GA. For example, GID1 mutants in rice exhibit severe dwarfism. Late flowering, morphological defects of floral organs, and the inhibition of seed germination were also observed in an Arabidopsis GID1 triple mutant[19].
The GRAS gene family has been identified in more than 50 species, including 60 in Oryza sativa, 34 in Arabidopsis thaliana, 62 in Hordeum vulgare, and 53 in Solanum lycopersicum[20−22]. DELLA, a well-studied subfamily of GRAS proteins, which have a novel regulatory N terminus that contains two conserved domains (DELLA and TVHYNP domains)[23]. DELLA acts as a negative regulator of the GA signaling pathway[24]. Gain-of-function mutants of the DELLA gene exhibit dwarfism and GA insensitivity, while loss-of-function mutations result in a constitutive GA-responsive phenotype[25,26]. DELLA is also involved in processes such as seed development and light signaling. In Arabidopsis, DELLA activity has been associated with larger seed size through the transcriptional activation of AINTEGUMENTA[27]. DELLA interacts with transcription factors BBX24 and ABI4 to regulate Arabidopsis morphogenesis[28−30]. DELLA regulates GA signaling through the GA-GID1-DELLA control module. In this pathway, when GA binds to GID1, the resulting GA-GID1 complex associates with the DELLA/TVHYNP motifs on the DELLA protein[31]. This interaction forms the GA-GID1-DELLA complex, which ultimately targets DELLA for degradation via the 26S proteasome[32].
Studying the GRAS and CXE families is essential because of their significant roles in physiological activities of plants and the prominent status of S. bacalensis in Chinese herbal medicine. In this study, 58 GRAS and 43 CXE families were identified from S. baicalensis and analyzed their phylogenies and tissue-specific expressions. This revealed their structural and expression diversities, providing a theoretical basis for further studies of GRAS and CXE functionality and offering new insights to interpret the gibberellin signal transduction in S. baicalensis.
-
The hidden Markov models of GRAS and CXE proteins were downloaded from the Pfam database (
http://pfam.xfam.org ), after which HMMER3.0 was used to search the GRAS and CXE genes in the S. baicalensis genome assembly of our lab with 0.01 cut-off value default parameters[33−34]. These were then verified by CD-search (www.ncbi.nlm.nih.gov/cdd ) and SMART (http://smart.embl-heidelberg.de/smart ) analysis in the NCBI domain analysis module, which identified a total of 58 SbGRASs and 43 SbCXEs. The physicochemical properties of SbGRAS and SbCXE were obtained using the ExPasy website (http://web.expasy.org/protparam ). Subcellular locations were predicted using the online website Plant-mPLoc (www.csbio.sjtu.edu.cn/bioinf/plant-multi ).Phylogenetic analyses of SbGRAS and SbCXE
-
To visualize the direct homology between S. baicalensis and A. thaliana, we used MEGA X to construct a phylogenetic tree, and then used the online website Evolview (
www.evolgenius.info/evolview ) to beautify the phylogenetic tree.Gene structure and motif composition of SbGRAS and SbCXE
-
MEME online software (
https://meme-suite.org/meme/tools/meme ) was used to identify the conserved motifs of SbGRAS and SbCXE proteins. By submitting the hitdata, MEME, and gff files of the S. baicalensis genome to Tbtools v1.089 software[35], the conserved motifs, domains, and gene structures of SbGRAS and SbCXE could be visualized. The sequence information of SbGRASs and SbCXEs can be found in the Supplementary File 1.SbGRAS and SbCXE duplication and synteny analyses
-
The chromosome positional data of SbGRAS and SbCXE was obtained from genomic data, and a chromosome location map was drawn using TBtools v1.089 software.
Analysis of protein-protein interactions in SbGRAS
-
The orthovenn2 website (
https://orthovenn2.bioinfotoolkits.net/home ) was used to search for the orthologous genes of A. thaliana and S. baicalensis, while the website String10 (http://string-db.org ) was used to analyze the interactions between SbGRAS and other proteins.Extraction of total RNA and synthesis of cDNA
-
The rhizomes and leaves of two-month-old S. baicalensis sterile seedlings were collected, and the plants were sprayed with 100 μM GA3 and 50 μM PAC (Paclobutrazol) solutions, respectively, and treated for 0, 1, 3, 6, and 12 h, respectively, to extract the RNA. Three independent biological replicates were utilized for processing.
The total RNA was isolated from cultured tissues using the Plant Total RNA Isolation Kit (Aidlab, Beijing, China). Reverse transcription of whole RNA into cDNA was done using superscript III reverse transcriptase (Invitrogen, Waltham, MA, USA).
RT-qPCR analysis
-
Using a 40-fold dilution of cDNA as a template, the 2−ΔΔCᴛ method was used to calculate the corresponding expression levels of SbGRASs and SbGID1s under the hormone treatment for different tissues[36]. Each reaction had three biological and technological replicates. The quantitative results were analyzed using GraphPad Prism 9 software. One-way ANOVA was employed to analyze the statistical significance of the data. The difference was statistically significant at < 0.05 for the p-value. The asterisk indicates a significant difference from 0 h, as determined by a one-way ANOVA Dunnett test. Data are expressed as mean ± SD, n = 3.
Subcellular localization
-
To determine the localization of SbGRAS in cells, SbGRAS1/4-GFP vectors were constructed. The onion epidermis was impregnated via Agrobacterium transformation, observed the transient expression of SbGRASs-GFP by fluorescence microscopy (Leica DM6000B, Wetzlar, Germany), and observed the nucleus with DAPI.
Transcription activation and yeast two-hybrid (Y2H) assay
-
SbGRASs and SbCXEs were constructed into pGBKT7 vectors, transferred into yeast AH109 competent cells, and transcriptionally activated using SD/-Trp-His-Ade containing X-α-gal after SD/-Trp growth.
Self-activating SbGRASs were incorporated into the pGADT7 vectors, and the BD-SbGID1s and AD-SbGRASs plasmids were transformed into yeast AH109 competent cells in equal amounts. The interactions between SbGRASs and SbGID1s were detected by SD/-Trp-His-Leu-Ade containing X-α-gal, after SD/-Trp-Leu growth. GA3 was added to the SD/-Trp-His-Leu-Ade medium at final concentrations of 10, 50, and 100 μM, respectively.
-
Based to the conserved GRAS and CXE domain and the Pfam database (PF03514/PF07859), a total of 58 GRAS sequences and 43 CXE sequences were identified in S. baicalensis, which were designated SbGRAS1-SbGRAS58 and SbCXE1-SbCXE43 (Supplementary Tables S1 and S2). The amino acid lengths varied significantly from 191 to 819 AA in SbGRASs and 71 to 579 AA in SbCXEs, with both showing considerable variation. Most of these proteins exhibited instability indices above 40, indicating they are unstable. Most of SbGRAS and SbCXE proteins were acidic and are thought to regulate the growth of S. baicalensis. The subcellular localization of 58 GRAS proteins was also predicted, finding that most were localized in the nucleus, whereas SbGRAS57 was localized in the cytoplasm. To determine the subcellular localization data of the GRAS family members, SbGRAS1-GFP and SbGRAS4-GFP fusion vectors were constructed. The transient expression of SbGRAS1 and SbGRAS4 in onion epidermal cells were studied (Fig. 1). The results indicated that the control group was localized in both the nucleus and cytoplasm, while SbGRAS1-GFP and SbGRAS4-GFP were localized in the nucleus, consistent with the predicted results.

Figure 1.
Subcellular localization analysis. SbGRAS1 and SbGRAS4 are constructed onto the pEarleyGate103 vector. Scale bars = 50 μm.
Phylogenetic analyses of SbGRAS and SbCXE
-
To investigate the evolutionary relationships of the GRAS and CXE gene families, the protein sequences of the A. thaliana GRAS and CXE families were identified and the cluster analysis was performed using MEGA X to classify SbGRAS into ten subfamilies (DELLA, SCL3, PAT1, SHR, LISCL, HAM, SCR, LAS, AT8, and SB) with reference to the classification method of AtGRAS. The PAT1 family had the highest number of members, with nine, whereas the LAS family had the fewest, with only three. Phylogenetic tree and conserved domain analyses revealed that SbGRAS1 (SbDELLA1), SbGRAS2 (SbDELLA2), SbGRAS3 (SbDELLA3), SbGRAS4 (SbDELLA4) belong to the DELLA subfamily (Fig. 2a). The CXE members were divided into five groups (Group1-5), where SbGID1A, SbGID1B, and AtGID1A, AtGID1B belong to the same evolutionary branch. The sequence agreement of the six genes was 89.22% (Fig. 2b).
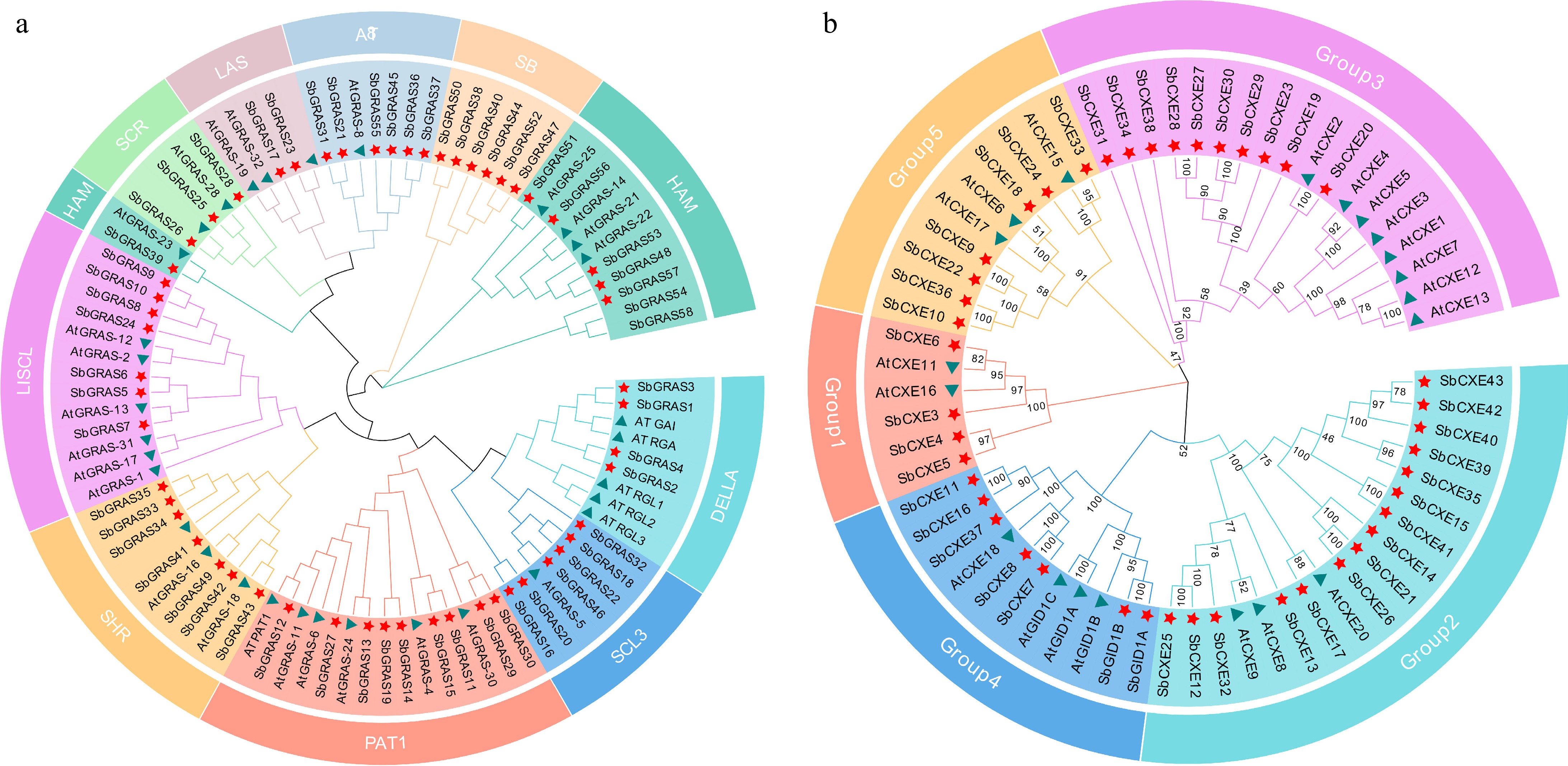
Figure 2.
Phylogenetic analysis of GRAS and CXE based on A. thaliana and S. baicalensis protein sequences. (a) GRAS proteins are divided into ten subfamilies, each of which is represented by a different color. Green triangles represent GRASs in A. thaliana, and red stars represent GRASs in S. baicalensis. (b) CXE proteins are divided into five subfamilies, each of which is represented by a different color. Green triangles represent CXE in A. thaliana, and red stars represent CXE in S. baicalensis. The phylogenetic tree was constructed by MEGA X using the neighbor-joining (NJ) method.
SbGRAS and SbCXE gene structures and motif compositions
-
To further investigate the functions of the GRAS and CXE gene families (Fig. 3), The gene structures and motifs of the SbGRAS members were analyzed and divided the 58 SbGRASs into ten subgroups. Ten conserved motifs in total were identified, noting that motif 2 was absent in the HAM subfamily, except for SbGRAS44. The remaining sequences possessed only one different motif, and these ten main motifs were widely distributed. Exon and intron analysis revealed that the number of exons in the SbGRAS family ranged from 1–14. From the structural perspective, SbGRAS1-4 had a typical DELLA domain. The 43 CXEs were divided into five subpopulations, and motif analysis revealed that Group1 and Group2 almost entirely lacked motif 10, with the exception of SbCXE13. The remaining sequences did not contain duplicate motifs. The number of exons in the SbCXE family exons ranged from 1–4, with most of the sequences containing only one exon (Fig. 3). Moreover, GRAS and CXE members within the same subfamilies have similar motif compositions. For instance, GRAS members of the SCR subfamilies lack motif 9, while CXE members of Group 1 subfamilies lack motifs 10. This implies functional conservation and diversity of the SbGRAS and SbCXE genes during evolution.
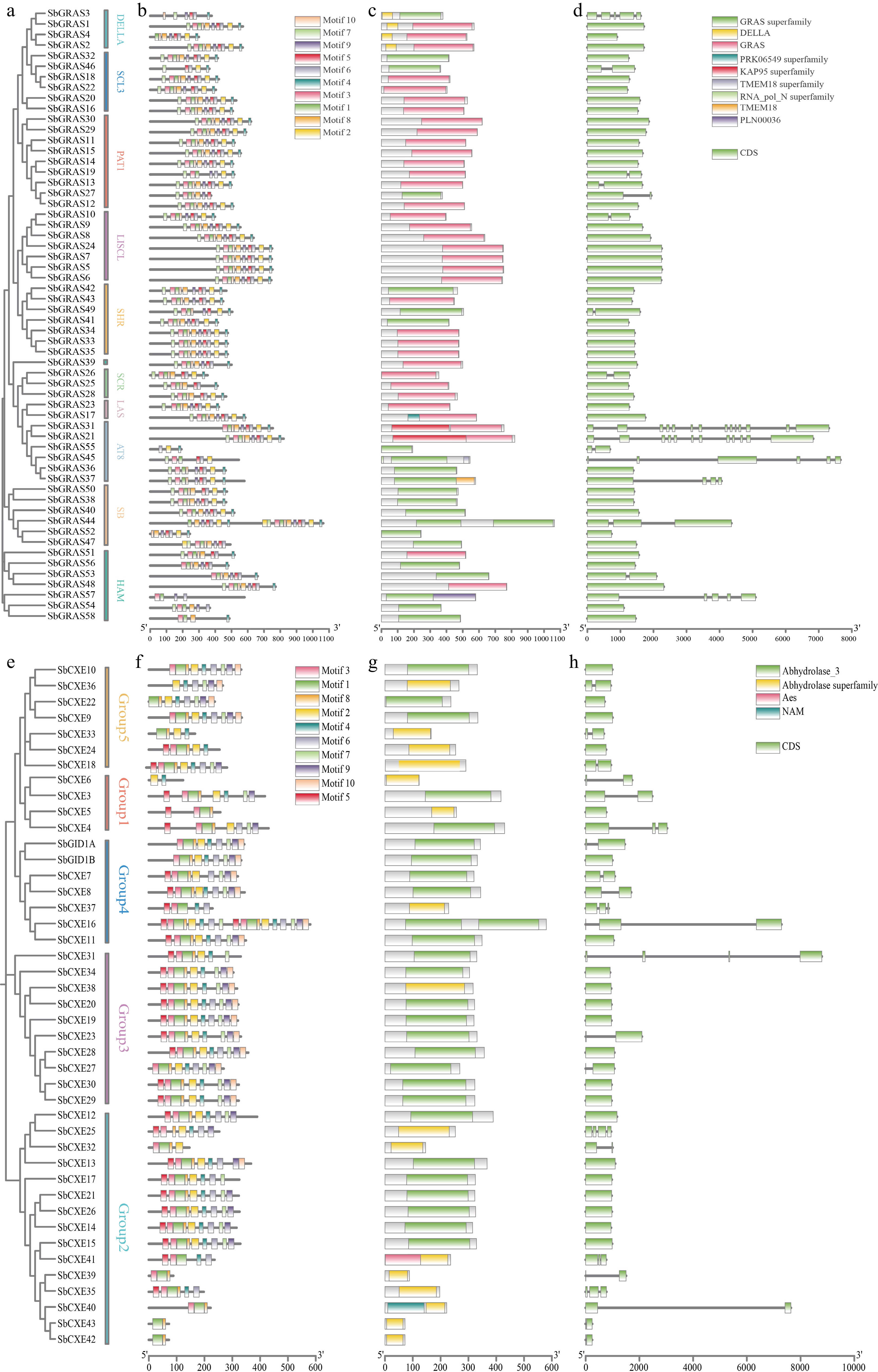
Figure 3.
Phylogeny, gene motifs, domains, and structural analyses of the S. baicalensis GRAS and CXE families. (a) Phylogenetic tree and grouping of the S. baicalensis GRAS family. (b) Gene motifs of GRAS in S. baicalensis. (c) GRAS family domains of S. baicalensis. (d) Gene structures of GRAS family in S. baicalensis. (e) Phylogenetic tree and grouping of the S. baicalensis CXE family. (f) Gene motifs of CXE in S. baicalensis. (g) CXE family domains of S. baicalensis. (h) Gene structures of CXE family in S. baicalensis.
SbGRAS duplication and synteny analysis
-
Based on chromosome annotation data, the S. baicalensis GRAS family amplification relationships were further analyzed (Fig. 4). Generally, tandem and segmental duplication were the primary gene amplification modalities[37]. A total of 35 gene pairs were identified, with tandem repeats referring to members of the same subfamily located at or near 30 bp. It was predicted that SbGRAS1 and SbGRAS3 are tandem duplication genes, whereas SbGRAS2 and SbGRAS4 were segmental duplication genes.
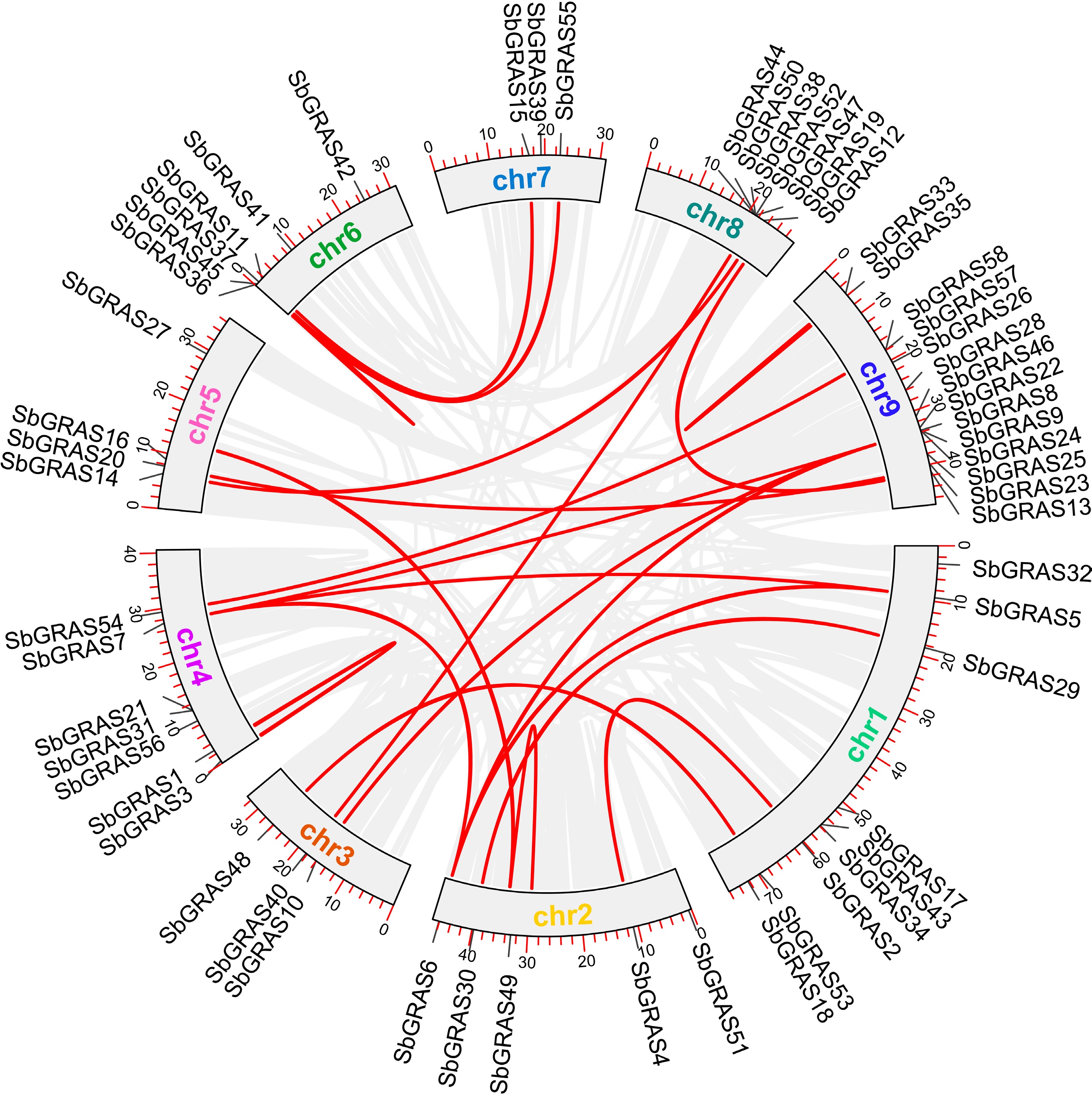
Figure 4.
SbGRAS chromosomal duplication analysis. Red lines represent SbGRAS gene pairs.
To explore the origins of the GRAS genes, three dicots (A. thaliana, S. tuberosum, S. lycopersicum) and two monocots (O. sativa and Z. mays) were selected to map the homology relationships with S. baicalensis. The number of homologous gene pairs in S. tuberosum and S. lycopersicum is more than other species, and they have a closer genetic relationship with S. baicalensis. The results suggest that these genes may have originated from a common ancestor and share similar functions (Fig. 5).
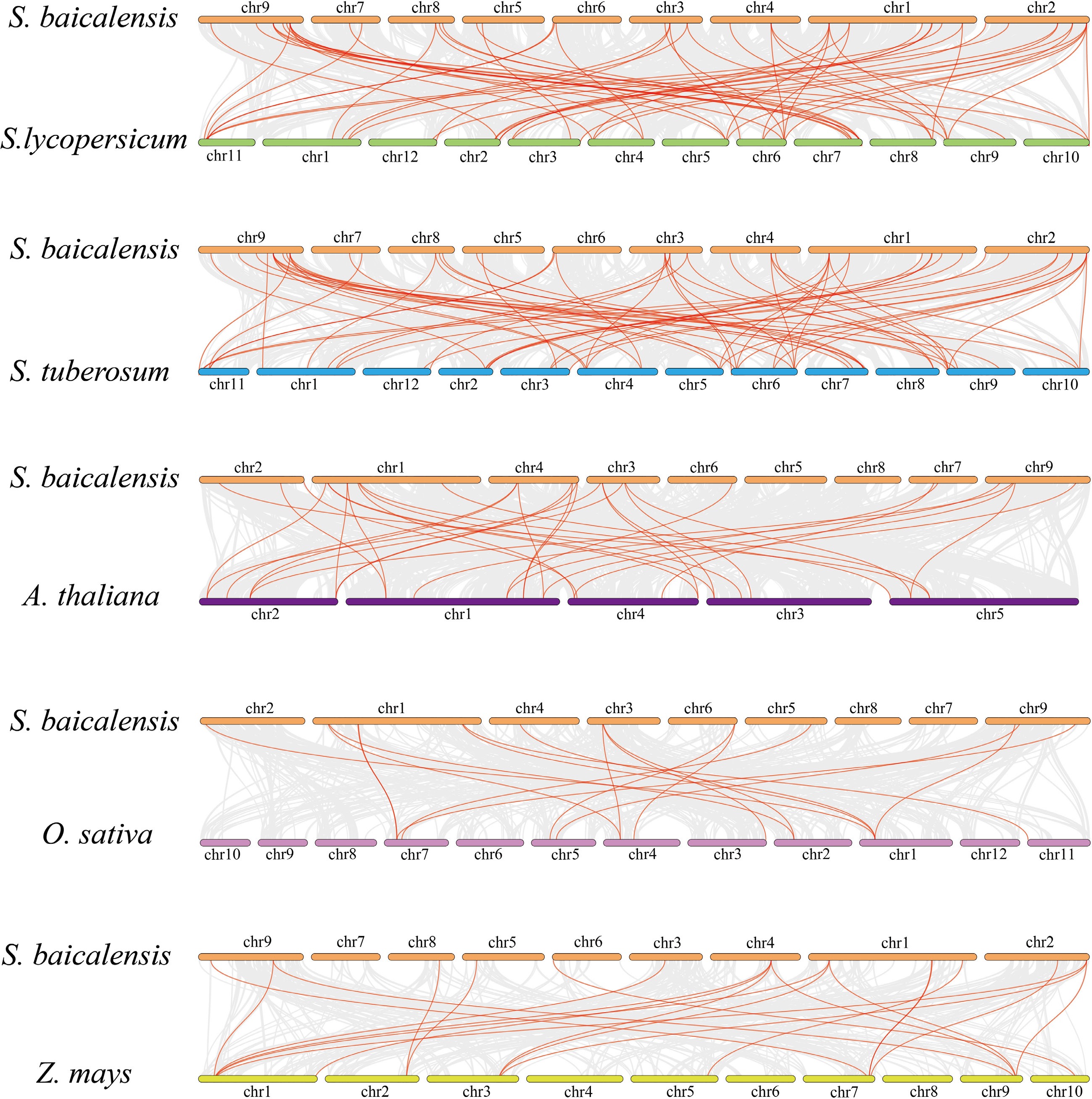
Figure 5.
Homology analysis of S. bacalensis and the remaining four plants. Gray lines indicate the homology genes of S. bacalensis and other species, and red lines indicate the GRAS of S. baicalensis and other species.
The Ka/Ks ratio represents the proportionality between the non-synonymous replacement rate (Ka) and synonymous replacement rate (Ks) of two protein-coding genes. The Ka/Ks ratios was calculated for all 58 genes and found that they were all less than 1, suggesting that the S. baicalensis GRAS family was purified and selected.
SbGRAS and SbGID1 expression patterns
-
The RNA was initially extracted from three S. baicalensis tissues: roots, stems, and leaves, and used RT-qPCR to investigate the expression patterns of SbGRAS and SbGID1 in different tissues (Fig. 6a). It was found that both SbGRAS and SbGID1 were highly expressed in the leaves.
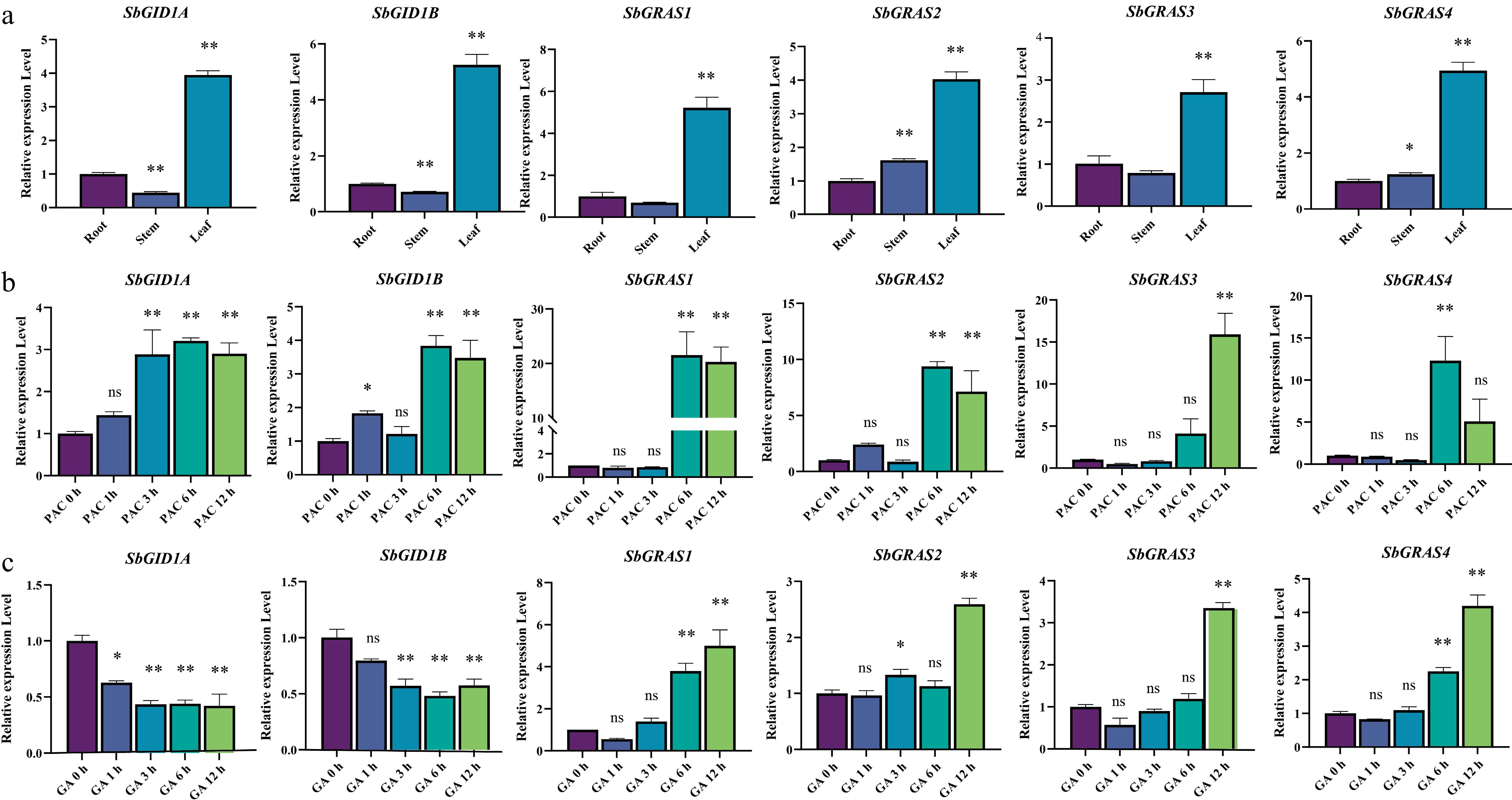
Figure 6.
Expressions of SbGRAS and SbCXE. (a) Expression levels of SbGRAS and SbCXE in tissues. (b) Relative expression levels of SbGRAS and SbCXE in S. baicalensis treated with 50 μM PAC were determined by RT-qPCR. (c) Relative expression levels of SbGRAS and SbCXE in S. baicalensis treated with 100 μM GA3 were determined by RT-qPCR. Data are expressed as mean ± SD, n = 3. Asterisks indicate a significant difference from 0 h, as determined by the one-way ANOVA Dunnett test: * p < 0.05; ** p < 0.01.
DELLA and GID1 are key proteins in the gibberellin signaling pathway and are essential for plant growth and development. In promoter analysis, it was observed that the DELLA and GID1 possess gibberellin regulatory elements (Supplementary Figs. S1 and S2). Consequently, the expression patterns of SbGRAS and SbGID1 under the GA3 and PAC (GA inhibitor) treatments were analyzed (Fig. 6b, c). The study revealed that GID1 responded positively to PAC treatment but was inhibited by GA3. Additionally, GID1 responded more quickly to GA3 treatment compared to PAC. This result can be compatible that GID1 is a gibberellin receptor. Compared to the analysis of DELLA expression patterns, it was found that DELLA responded differently to GA3 and PAC stresses. DELLA responded more slowly to both GA3 and PAC compared to GID1, but the extent of its response is greater than that of GID1. Under GA3 treatment, the expression of DELLA increased by approximately three times, contrary to the results of GID1 inhibition. Surprisingly, DELLA also responded positively to PAC treatment, with an expression level about five times higher than under GA3 treatment.
Analysis of SbGRAS protein-protein interactions
-
The homologous genes of S. baicalensis and A. thaliana was obtained using the orthovenn2 online website, and then constructed the protein interaction network of GRAS using the STRING protein interaction online website. In A. thaliana, four GIDs (GID1A, GID1B, GID1C, and GID2) and four DELLAs (RGA/GAI/RGL1/RGL2) had complex interactions with each other, specially GIDs only had effects with DELLAs and other GIDs. DELLAs, in addition to interacting with GIDs, also interact with other transcription factors to perform broader functions, the result revealed that DELLAs and GID1s proteins had complex interactions (Fig. 7).
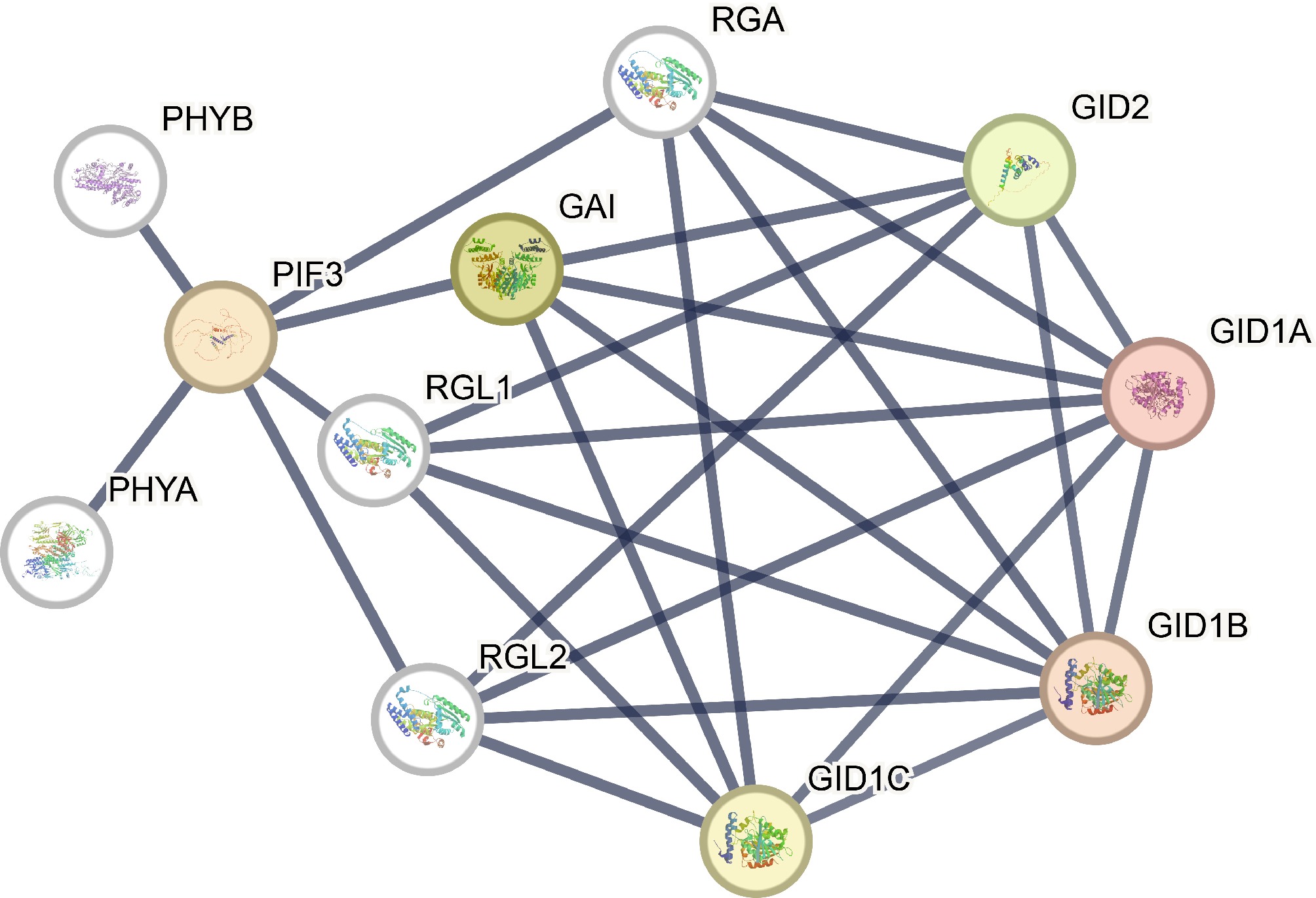
Figure 7.
Protein-protein interaction networks of GRAS proteins. Line thicknesses reflect the strengths of the interactions between proteins.
SbGRASs interacts with SbGID1s
-
The results of protein interaction predictions showed that there be interactions between DELLAs and GID1s. The BD-SbGRASs and BD-SbGID1s fusion vectors were constructed with unloaded pGBKT7 as a negative control, and it was found that SbGRAS1 and SbGRAS2 engaged in self-activating activities (Fig. 8a).

Figure 8.
SbGID1s interaction with SbDELLAs. (a) Transcriptional activity of SbGID1s and SbDELLAs. The negative control (pGBKT7). (b) Interaction between SbGID1s and SbDELLAs. The negative control (pGBKT7 + pGADT7). (c) Facilitated the interaction between SbDELLAs and SbGID1s by adding 10 μM GA3/50 μM GA3/100 μM GA3 to SD/-THLA medium. Negative control (pGBKT7 + pGADT7).
GA signal transduction depends on the interactions between SbGRASs and SbGID1s. In order to verify the interactions, SbGID1s were constructed into the pGBKT7 vector and SbGRASs were constructed into the pGADT7 vector. The results suggested that in the absence of GA3, BD-SbGID1A and AD-SbGRAS1, AD-SbGRAS2; BD-SbGID1B and AD-SbGRAS1, AD-SbGRAS2 turned blue on SD/-Trp-His-Leu-Ade medium, indicating that there was interaction between SbGID1A/B with SbGRAS1/2. AD-SbGRAS4 and BD-SbGID1A and BD-SbGID1B did not turn blue, which implied that there were no interaction between SbGID1A/B with SbGRAS4 (Fig. 8b), meaning that DELLA also has other functions which is consistent with the above prediction. When 10, 50, 100 μM GA3 was added to the medium progressively, the interactions between SbGID1A/B with SbGRAS1/2 were gradually becoming stronger, with the yeast turning blue more rapdily as the GA3 concentration increased (Fig. 8c). The results revealed that the GA3 promoted the interactions between SbDELLAs and SbGID1s.
-
The root of S. baicalensis could be used for lung and liver diseases treatment as a widely used traditional Chinese medicine[38,39]. The gene families GRAS and CXE play crucial roles in plant growth and development. The GRAS gene family has been reported in crops such as A. thaliana[13], S. lycopersicum[15], and Z. mays[40], as well as in Chinese medicinal materials such as Dendrobium chrysotoxum[41], Panax ginseng[42]. The CXE gene family has also been identified in A. thaliana[30], Prunus persica[43] and Salvia miltiorrhiza[44]. However, GRAS and CXE gene families have not been reported in S. baicalensis. Here, 58 SbGRAS and 43 SbCXE genes in S. baicalensis were identified. The sequences within these two families exhibited significant divergence, suggesting that the GRAS and CXE gene families have undergone repeated expansion events throughout evolution. Based on the classification of A. thaliana, the identified SbGRAS and SbCXE genes were classified, and the sequences within each cluster exhibited high similarity. Interestingly, there were six SbGRAS that were not clustered with A. thaliana, while PtGRAS20 was reported to be endemic in poplar[45]. The uniqueness of these six SbGRAS genes to S. baicalensis requires further verification.
DELLA is a negative regulator of the GA signaling pathway and interacts with the GID1 to recognise GA signals[46]. In this study, the SbDELLAs and SbG1D1s were found to be significantly expressed following PAC and GA3 treatment, suggesting that two genes may play the important roles in the GA signaling pathway in S. baicalensis. Meanwhile, the expression of SbDELLAs was upregulated under GA3 treatment, a phenomenon similar to that observed in S. miltiorrhiza[43], indicating that four DELLAs played a synergistic role in GA signal transduction. SbGID1s expression significantly decreased under GA3 treatment, this result was consistent with A. thaliana[47], but inconsistent with S. miltiorrhiza. This discrepancy suggests that GID1s may play different roles in different species, likely due to species-specific differences. This study also found that SbGID1s responded to GA3 treatment significantly more rapidly than SbDELLAs, which was consistent with the characteristics of GID1s as GA receptor genes. PAC, a GA inhibitor, led to a notable increase in both SbGID1s and SbDELLAs expression when compared to GA3 treatment. However, the expression of SbDELLAs should be inhibited under PAC treatment, and SbDELLAs responded to PAC treatment much more swiftly than to GA3 treatment. Therefore, it was speculated that GID1s may be relatively conserved evolutionarily, while DELLAs have acquired additional functions through evolutionary processes.
The protein-protein interaction network revealed a strong association between DELLAs and GID1s. Specifically, SbGRAS1 and SbGRAS2 interacted with SbGID1s, and SbGRAS4 did not interact with SbGID1s in S. baicalensis. Upon the addition of GA3, SbGRAS4 was unable to bind to SbGID1s, however, it facilitated the binding between SbGID1s and SbGRAS1 and SbGRAS2. RT-qPCR results suggest that SbGRAS4 may have distinct functions in S. baicalensis. Furthermore, the interaction between SbDELLA and SbGID1 in S. baicalensis belongs to GA-independent interactions[48]. In summary, GA stimulated the formation of the GA-GID1-DELLA complex[49] and enhanced the interactions of SbDELLAs and SbGID1s.
-
This study performed the first genomic analysis of GRAS and CXE families of S. baicalensis. A total of 58 SbGRAS members and 43 SbCXE members were identified and four DELLAs (SbGRAS1-SbGRAS4) and two GA receptors (SbGID1A and SbGID1B) were identified, showing significantly responsive to GA3 and PAC treatments. The study investigated direct interactions between DELLAs and GID1s, and the effects of gibberellin on these interactions were investigated. This study provides a foundational understanding of the molecular mechanisms of GRAS and CXE genes under plant abiotic stress, and offer insights into the gibberellin signaling pathway in S. baicalensis.
This research was supported by the Project of Shaanxi Science and Technology Department (Grant No. 2020JM-628), the Fundamental Research Program of Shanxi Province (Grant No. 202203021212013), and the Science and Technology Plan Project of Yulin City (Grant No. 2024-CXY-157).
-
The authors confirm their contributions to the paper as follows: experiments conception and design: Wang Z, Wang D, Pang Q; vectors construction, data analyses: Pang Q, Xie Z, Han W; vectors sequencing: Niu J, Wang S, Zhou W, Li L, Pang Q; draft manuscript preparation: Pang Q. All authors reviewed the results and approved the final version of the manuscript.
-
The authors confirm that all data supporting the findings of this study are available within the article.
-
The authors declare that they have no conflict of interest.
- Supplementary File 1 cDNA sequences.
- Supplementary Table S1 Physicochemical properties of the GRAS family of S. baicalensis.
- Supplementary Table S2 Physicochemical properties of the CXE family of S. baicalensis.
- Supplementary Fig. S1 Distribution of cis-elements in the 2000 bp promoter region upstream of SbCXEs. Different cis-elements are denoted by different colors.
- Supplementary Fig. S2 Distribution of cis-elements in the 2000 bp promoter region upstream of the SbGRASs. Different cis-elements are denoted by different colors.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Pang Q, Xie Z, Han W, Niu J, Wang S, et al. 2025. Genome-wide analysis of GRAS and CXE and interactions between SbDELLAs and SbGID1s in Scutellaria baicalensis Georgi. Medicinal Plant Biology 4: e031 doi: 10.48130/mpb-0025-0026 

Genome-wide analysis of GRAS and CXE and interactions between SbDELLAs and SbGID1s in Scutellaria baicalensis Georgi
- Received: 27 March 2025
- Revised: 06 June 2025
- Accepted: 09 June 2025
- Published online: 29 September 2025
Abstract: A total of 58 SbGRAS and 43 SbCXE family members were identified using phylogenetic analysis, based on the S. baicalensis genome database. And four DELLA subfamily members (SbGRAS1-4) and two gibberellin receptors (SbGID1A and SbGID1B) were identified. This study found that SbDELLAs and SbGID1s significantly responded to GA3 and PAC treatments. GA signal transduction depends on the interactions between SbDELLAs and SbGID1s, SbGRAS1 and SbGRAS2 can interact with GID1s regardless of the presence of GA3, and this interaction is accelerated when GA3 is added. This suggests that SbGRAS1 and SbGRAS2 play a more critical role in gibberellin signaling. This study provides the systematic analysis and investigation of two gene families, and offers guidance for regulating flavonoid metabolism in the gibberellin signaling pathway in S. baicalensis.
-
Key words:
- Scutellaria baicalensis Georgi /
- Gene family /
- SbGRAS /
- SbCXE /
- Gibberellin (GA)