









-
The shift from vegetative to reproductive growth represents a pivotal developmental checkpoint that determines plant reproductive success. To ensure optimal yield and quality, plants must initiate flowering only when environmental conditions are favorable, making flowering time a key determinant of agricultural performance[1]. This transition is orchestrated by an intricate regulatory network that integrates endogenous genetic programs with a range of external cues to fine-tune floral induction[2]. Extensive studies in long-day species such as Arabidopsis thaliana and short-day crops such as Oryza sativa have delineated five core flowering pathways: autonomous, photoperiodic, vernalization, gibberellin (GA), and age pathways[3]. Recent advances have further expanded this regulatory framework by uncovering additional mechanisms involving cytokinin (CK) signaling, temperature responsiveness, and carbohydrate metabolism[4−6].
Vernalization, the acquisition of flowering competence after prolonged exposure to low temperatures is a crucial evolutionary strategy that allows many species to flower in spring following winter cold. In Arabidopsis, vernalization accelerates flowering by epigenetically silencing FLOWERING LOCUS C (FLC), a central repressor that blocks the activation of floral pathway integrators[7]. Genes such as VERNALIZATION INSENSITIVE 3 (VIN3), VERNALIZATION 1 (VRN1), and VERNALIZATION 2 (VRN2) are essential components of this process, mediating cold-induced chromatin remodeling at the FLC locus, and ultimately enforcing its transcriptional repression[8]. In wheat, vernalization responses rely on the coordinated action of VRN1, 2, and 3, in which VRN1 and 3 are flowering activators, whereas VRN2 serves as a flowering repressor whose expression declines during cold exposure, thereby releasing VRN3 activity and promoting floral induction[9,10]. The integration of cold signals ultimately converges on flowering regulators such as FLOWERING LOCUS T (FT), and SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1), which in turn activate floral meristem identity genes, including AP1 and LEAFY (LFY), to direct the specification and establishment of floral meristems[11]. Subsequent activation of genes controlling floral organogenesis drive the development and maturation of floral structures[12].
Dendrobium, comprising roughly 1,450 species, is the third largest genus within the Orchidaceae, and is primarily distributed across tropical and subtropical ecosystems[13,14]. The genus is generally classified into two horticultural groups: phalaenopsis-type and nobile-type orchids. Phalaenopsis-type plants typically initiate flowering in autumn before the onset of cold weather, whereas nobile-type plants require exposure to low temperatures for floral induction and subsequently bloom in spring. Nobile-type Dendrobium is prized for both its ornamental value and its long-standing medicinal use[15]. However, its extended juvenile phase and irregular growth cycle impose major constraints on large-scale cultivation and industrial utilization, intensifying interest in technologies that precisely control flowering time. Artificial low-temperature treatment (often termed artificial vernalization) is widely implemented to shorten the vegetative period and synchronize flowering in Dendrobium[16]. Suboptimal temperature management can result in failure to flower or the formation of vegetative offshoots at stem nodes[17]. The application of transcriptomic approaches has greatly advanced our understanding of cold-responsive flowering pathways in Dendrobium[18−20]. Nonetheless, a temperature-independent genetic strategy for controlling flowering time in this species has yet to be established. Deciphering how ambient temperature governs both floral initiation and floral organ development is crucial for molecular breeding, and has substantial implications for the orchid industry.
To systematically characterize the transcriptional reprogramming associated with chilling-induced flowering in Dendrobium, we performed controlled temperature treatments and observed that sufficient chilling accumulation accelerated flowering by nearly 1 month. We further integrated cytological observations with transcriptome sequencing across five developmental stages, from vegetative buds to fully opened flowers (S1–S5), under both natural and artificial temperature conditions. This study provided mechanistic insights and practical guidance for artificial regulation of flowering time, while offering valuable resources to support molecular breeding in nobile-type Dendrobium.
-
Plants used in this study were obtained from cross-bred hybrids of nobile-type Dendrobium. Control plants were cultivated in a greenhouse at the Dongguan Agricultural Science Research Center, China, where temperatures naturally fluctuated between 10 and 35 °C with a relative humidity of 70%–80% (designated as the RT group). For artificial chilling treatment, plants were transferred to a climate-controlled greenhouse maintained at 20 °C during the day, and 10 °C at night under a 16 h light/8 h dark photoperiod until flowering (LT group). Five developmental stages were sampled, including vegetative growth stage (S1), floral initial differentiation stage (S2), floral organ development stage (S3), flower enlargement and development stage (S4), and blooming stage (S5), each collected in two biological replicates. After removing scale bracts, samples were immediately flash-frozen in liquid nitrogen and stored at −80 °C until RNA extraction.
Scanning electron microscopy
-
Lateral buds were dissected by removing surrounding scale tissues and fixed in a solution containing 2% formaldehyde and 3% glutaraldehyde. Fixed tissues were dehydrated through a graded acetone series and subsequently subjected to critical-point drying using liquid carbon dioxide as the transitional fluid. Dried samples were mounted on aluminum stubs, coated with an approximately 25 nm gold layer via sputter deposition, and examined using a JSM-6360LV scanning electron microscope (JEOL, Japan).
RNA extraction, library preparation, and sequencing
-
Total RNA was extracted from floral bud tissues using a commercial kit (TaKaRa). RNA purity and concentration was assessed with a NanoDrop 2000 spectrophotometer (Thermo Scientific, USA), and RNA integrity was evaluated according to the manufacturer's guidelines. Ten high-quality RNA samples were selected for library construction. Fragmented cDNA molecules were purified using the AMPure XP system (Beckman Coulter, USA) to enrich inserts approximately 240 bp in length. After PCR amplification, library quality was confirmed using the Agilent Bioanalyzer 2100 system. The final cDNA libraries were sequenced on the Illumina HiSeq 2000 platform. Raw reads were filtered to remove adapter sequences, poly-N reads, and low-quality bases, generating high-quality clean datasets. Quality metrics, including Q20 and Q30 values, GC content, and duplication rates, were subsequently calculated. As no reference genome is available for Dendrobium, de novo transcriptome assembly was performed using Trinity to generate a comprehensive reference transcript set from the clean reads.
Differential gene expression analysis
-
Gene expression levels were normalized and quantified using the FPKM (fragments per kilobase of transcript per million mapped reads) method. Differentially expressed genes (DEGs) across pairwise sample comparisons were identified with edgeR (version 1.30.1). The resulting p-values were adjusted using the Benjamini–Hochberg procedure to control the false discovery rate. Genes exhibiting an adjusted p-value < 0.05, and |log2FC| ≥ 2 were designated as DEGs. Expression pattern clustering was performed using the Mfuzz package (
https://mfuzz.sysbiolab.edu.eu ), and heatmaps were generated with TBtools to visualize global expression dynamics.Functional annotation analysis
-
Functional annotation of assembled unigenes was performed by aligning sequences against several major protein databases, including NCBI non-redundant (Nr), Eukaryotic Clusters of Orthologous Groups (KOG), Protein families (Pfam), Clusters of Orthologous Groups of proteins (COG), Swiss-Prot, the Kyoto Encyclopedia of Genes and Genomes (KEGG), and Gene Ontology (GO), using BLASTx with an E-value threshold of 1.0E−5. To elucidate the biological functions of DEGs and identify significantly enriched pathways across developmental stages, DEGs were mapped to GO terms and KEGG pathways. Genes meeting the edgeR-derived criterion of adjusted p-value < 0.05 were classified as DEGs, and subsequently subjected to enrichment analysis.
Quantitative real-time PCR (qRT-PCR)
-
qRT-PCR was conducted to validate the expression profiles of selected genes. First-strand cDNA was synthesized from 1 μg of total RNA using Hiscript III Reverse Transcriptase (Vazyme, Nanjing, China). qRT-PCR was performed on a Bio-Rad CFX96 system using the BioSmart U+ Super Multiple Probe qPCR PreMix (Vazyme, Nanjing, China). Gene-specific primers were designed with Primer-BLAST (
https://www.ncbi.nlm.nih.gov ). DnActin was used as the internal reference gene for normalization. Relative transcript abundance was calculated using the 2−ΔΔCᴛ method. Three biological replicates were included for each sample. All primer sequences are listed in Supplementary Table S1. -
To investigate the detailed morphological characteristics associated with floral development in Dendrobium, we conducted comprehensive microscopic observations throughout the entire developmental period. The floral ontogeny of Dendrobium could be clearly divided into five consecutive and recognizable stages. Stage 1: the stalk nodes appeared noticeably flattened, and the latent buds remained tightly enveloped by the surrounding leaf sheaths. At this point, the internal growth cone was still in a completely undifferentiated state, showing no visible signs of reproductive development (Fig. 1a, Supplementary Fig. S1a). Stage 2: floral buds reached approximately 0.7 cm in height. During this early differentiation phase, the buds began to expand outward from the stalk nodes, gradually breaking through the protective leaf sheaths. The buds exhibited an elliptical shape, and pronounced bulges emerged along the flanks of the developing inflorescences. These protrusions further differentiated into multiple floral primordia, forming initial zygomorphic structures, and signaling the onset of the reproductive program (Fig. 1b, Supplementary Fig. S1b). Stage 3: the floral buds grew to about 3 cm in height, accompanied by elongation and separation of the external scale tissues. Internally, early floral structures became more distinct and continued to develop, forming clearly identifiable sepal, petal, lip, and column, which progressively expanded as floral differentiation advanced (Fig. 1c, Supplementary Fig. S1c). Stage 4: the dominant floret underwent rapid enlargement, broke through the membranous bracts, and displayed significant expansion. The floral axis elongated markedly, and pigmentation began to appear in floral organs, particularly within the petals and the labellum, indicating substantial developmental progression (Fig. 1d, Supplementary Fig. S1d). Stage 5: all floral organs continued to increase in size, reached full maturity, and ultimately proceeded to complete anthesis (Fig. 1e, Supplementary Fig. S1e).
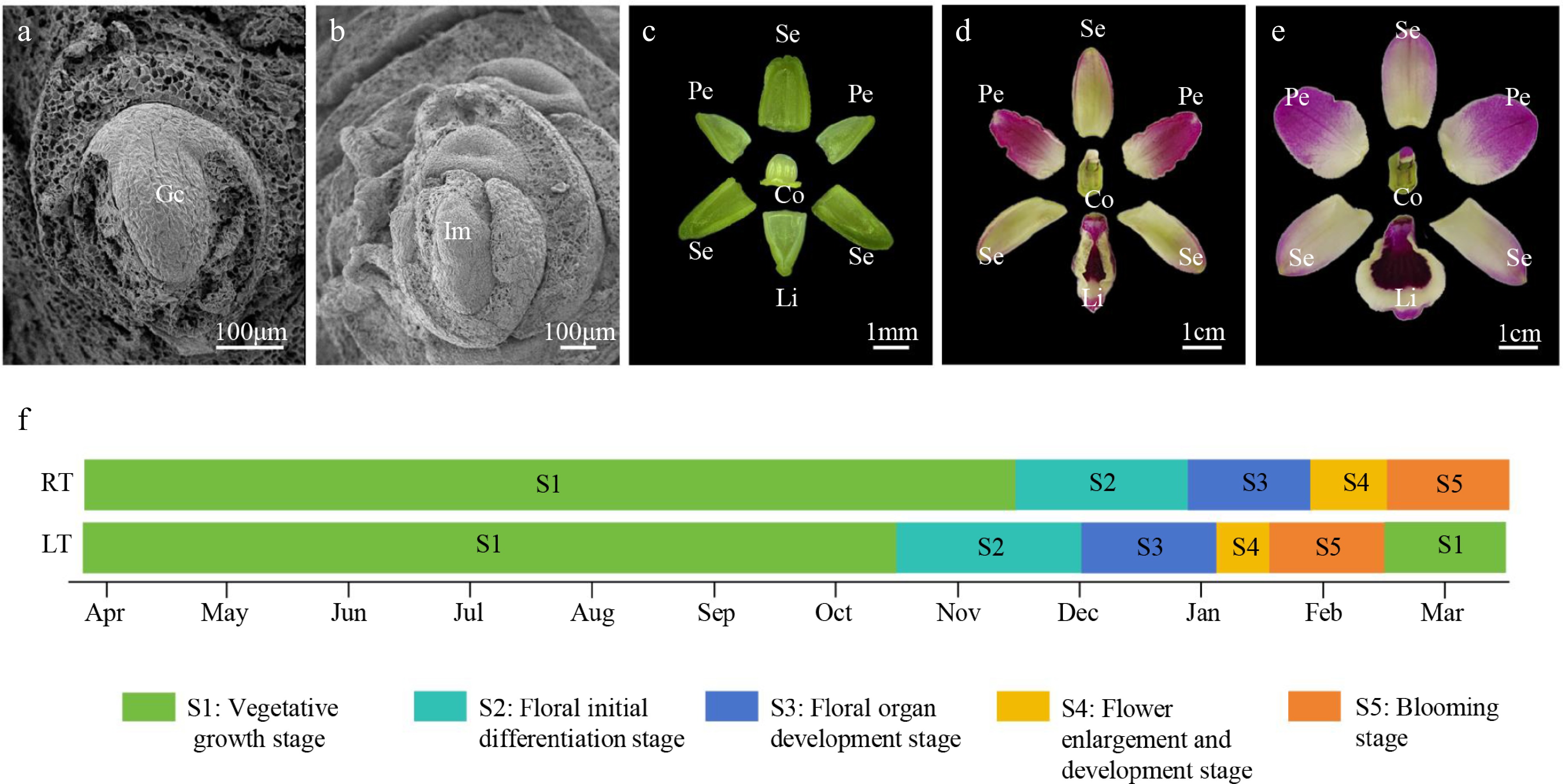
Figure 1.
A comprehensive morphological and cytological analysis of the floral developmental process in Dendrobium. (a) S1: vegetative growth stage. (b) S2: floral initial differentiation stage. (c) S3: floral organ development stage. (d) S4: flower enlargement and development stage. (e) S5: blooming stage. (f) Comparison of floral development timing between RT (natural condition) and LT (artificial chilling condition) treatments. Gc, growth cone; Im, inflorescence meristem; Se, sepal; Pe, petal; Li, lip; Co, column.
A comparison of developmental timing under the two temperature regimes revealed distinct differences. Under LT treatment, Dendrobium transitioned into the reproductive phase as early as mid-October and reached full bloom by mid-January. In contrast, under natural room-temperature conditions, buds entered S2 only in November, with declining outdoor temperatures, and flowering did not occur until mid-February. These results demonstrated that exposure to low temperature significantly accelerated the flowering process by shortening the vegetative period, and advanced anthesis by approximately 1 month compared with plants grown under regular environmental conditions (Fig. 1f). Moreover, statistical analyses indicated that artificial chilling exerted only minimal effects on overall floral quality parameters in Dendrobium, suggesting that chilling treatment effectively enhanced flowering time without compromising floral traits (Supplementary Table S2).
Transcriptome analysis of Dendrobium under RT vs LT conditions
-
To examine how artificial low-temperature treatment altered the flowering regulatory mechanisms of Dendrobium in comparison with natural temperature conditions, we performed a comprehensive transcriptomic analysis using floral bud samples collected across developmental stages. After removing adaptor sequences, contaminants, and low-quality reads, a total of 140.61 Gb of high-quality, clean data was generated from 20 sequencing libraries, with Q30 values for all reads exceeding 91.37% (Supplementary Table S3). De novo assembly ultimately produced 58,955 unigenes, of which 36,403 exceeded 1 kb in length, indicating robust assembly continuity (Supplementary Fig. S2a, Supplementary Table S4). Principal component analysis (PCA) based on gene expression profiles revealed clear and consistent clustering of samples according to developmental stage, reflecting strong dataset reliability and biological reproducibility (Supplementary Fig. S4). Among the annotated unigenes with significant similarity to known plant sequences, Dendrobium catenatum accounted for 85.55% of all matches, whereas Phalaenopsis equestris represented 2.99%, highlighting the close phylogenetic proximity between Dendrobium and these orchid species (Supplementary Fig. S2b). An integrated summary of functional annotations for all assembled transcripts is provided in Supplementary Fig. S2c.
DEGs and expression trends in Dendrobium under RT vs LT conditions
-
To identify candidate DEGs involved in the regulation of flowering, pairwise comparisons between developmental stages were conducted using |log2 ratio| > 2 and p < 0.05 as significance thresholds. As floral development progressed, the number of DEGs increased progressively in both RT and LT treatment groups, with the total number of DEGs under RT exceeding those detected under LT conditions (Fig. 2a, c). Comparative analysis across developmental stages under RT conditions revealed 2,922 DEGs shared across all stages, in addition to 545, 1,608, 2,117, and 3,881 stage-specific DEGs between consecutive stages (Fig. 2b). Under LT conditions, 1,578 DEGs were shared among all stages, while 374, 1,497, 926, and 4,492 unique DEGs were identified between successive developmental transitions (Fig. 2d). In total, 13,708 DEGs were identified when directly comparing RT and LT treatments (Fig. 2e). The number of unique DEGs in the five pairwise RT–LT comparisons were 3,849, 368, 1,174, 818, and 1,549, respectively. Notably, the S1 comparison exhibited the highest DEG count, suggesting that the initial transition stage may involve more complex physiological, metabolic, or transcriptional adjustments in response to temperature changes. Conversely, the S2 group displayed the fewest DEGs, implying a relatively stable transcriptional state at this early floral differentiation phase. Moreover, nine DEGs were consistently detected across all developmental stages under both RT and LT conditions, indicating that these genes might function as core regulators of the floral developmental program in Dendrobium (Fig. 2f). To further investigate the expression trends of DEGs at identical developmental stages between the cold-treated and control groups, Mfuzz clustering was performed, revealing nine distinct expression profiles under each temperature regime (Fig. 2g, h). A substantial number of genes displayed high expression during the intermediate floral developmental period, suggesting the presence of critical transcriptional activation or repression events that likely orchestrate major developmental transitions during this stage.
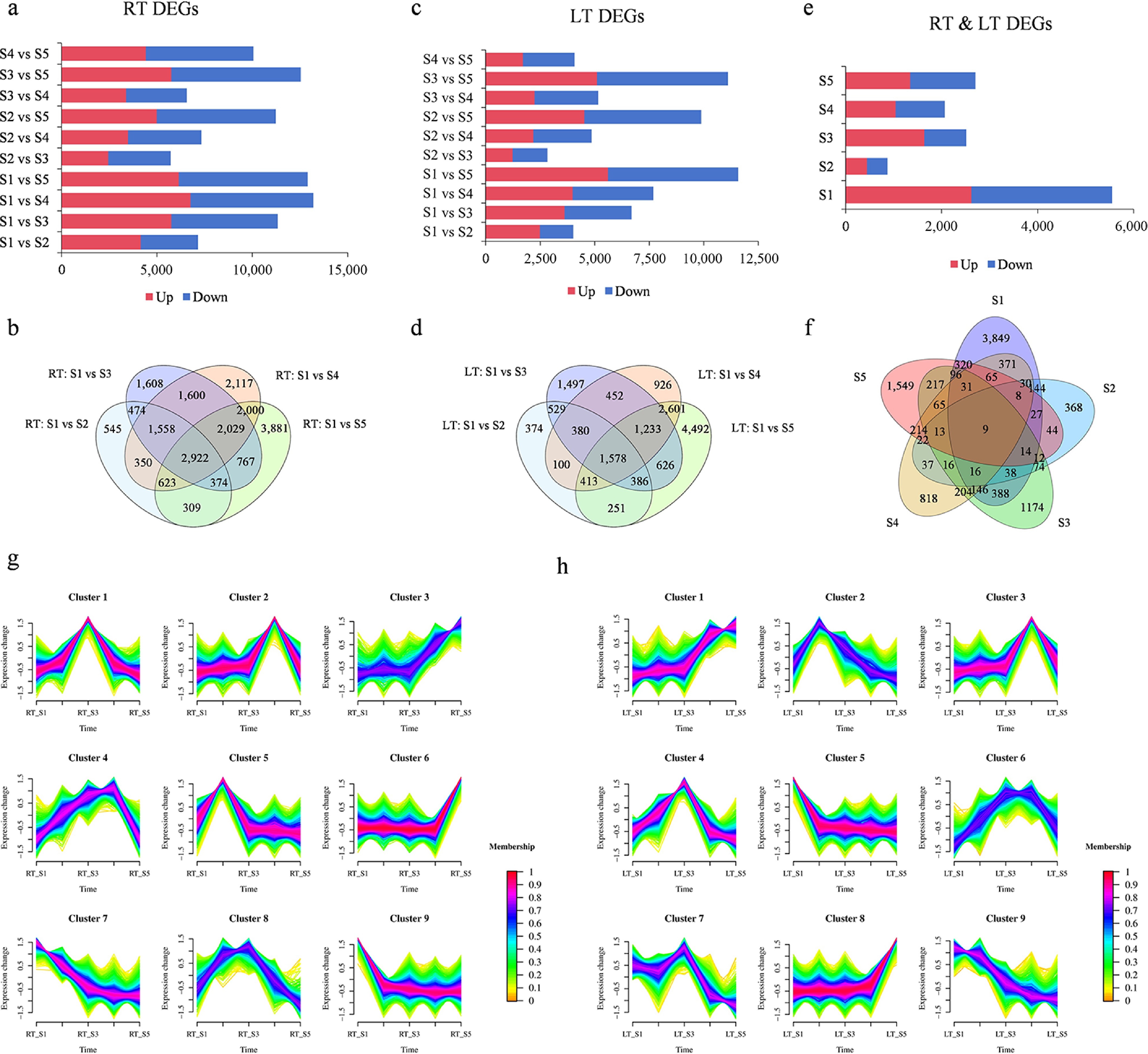
Figure 2.
Transcriptome analysis of Dendrobium under RT vs LT conditions. (a) The number of up-/downregulated DEGs in Dendrobium under RT conditions. (b) The Venn map indicating the number of coregulated and specifically regulated DEGs during Dendrobium development under RT conditions. (c) The number of up-/downregulated DEGs in Dendrobium under LT conditions. (d) The Venn map indicating the number of coregulated and specifically regulated DEGs during Dendrobium development under LT conditions. (e) The number of up-/downregulated DEGs in the two treatment comparison groups. (f) Venn diagram of all DEGs in the comparison between the two treatments. (g) Clustering analysis of time-course RNA-seq data revealing DEGs changes during Dendrobium development under RT conditions. (h) Clustering analysis of time-course RNA-seq data revealing DEGs changes during Dendrobium development under LT conditions.
Enrichment analysis of DEGs under RT vs LT conditions
-
To further elucidate the biological processes and metabolic activities affected by temperature treatments, KEGG enrichment analysis was performed for DEGs identified across the five pairwise comparisons. In the S1 comparison, the most significantly enriched pathways included DNA replication, pentose and glucuronate interconversions, and ubiquinone and other terpenoid–quinone biosynthesis (Fig. 3a). In S2, DEGs were predominantly associated with photosynthesis–antenna proteins, photosynthesis, and linoleic acid metabolism (Fig. 3b). For S3, enrichment was observed in pathways related to flavonoid biosynthesis, plant hormone signal transduction, and cutin, suberine, and wax biosynthesis (Fig. 3c). The S4 comparison was characterized by significant enrichment in plant hormone signal transduction, terpenoid backbone biosynthesis, and the circadian rhythm pathway (Fig. 3d). In S5, the most enriched pathways included photosynthesis-antenna proteins, isoflavonoid biosynthesis, and limonene and pinene degradation (Fig. 3e).
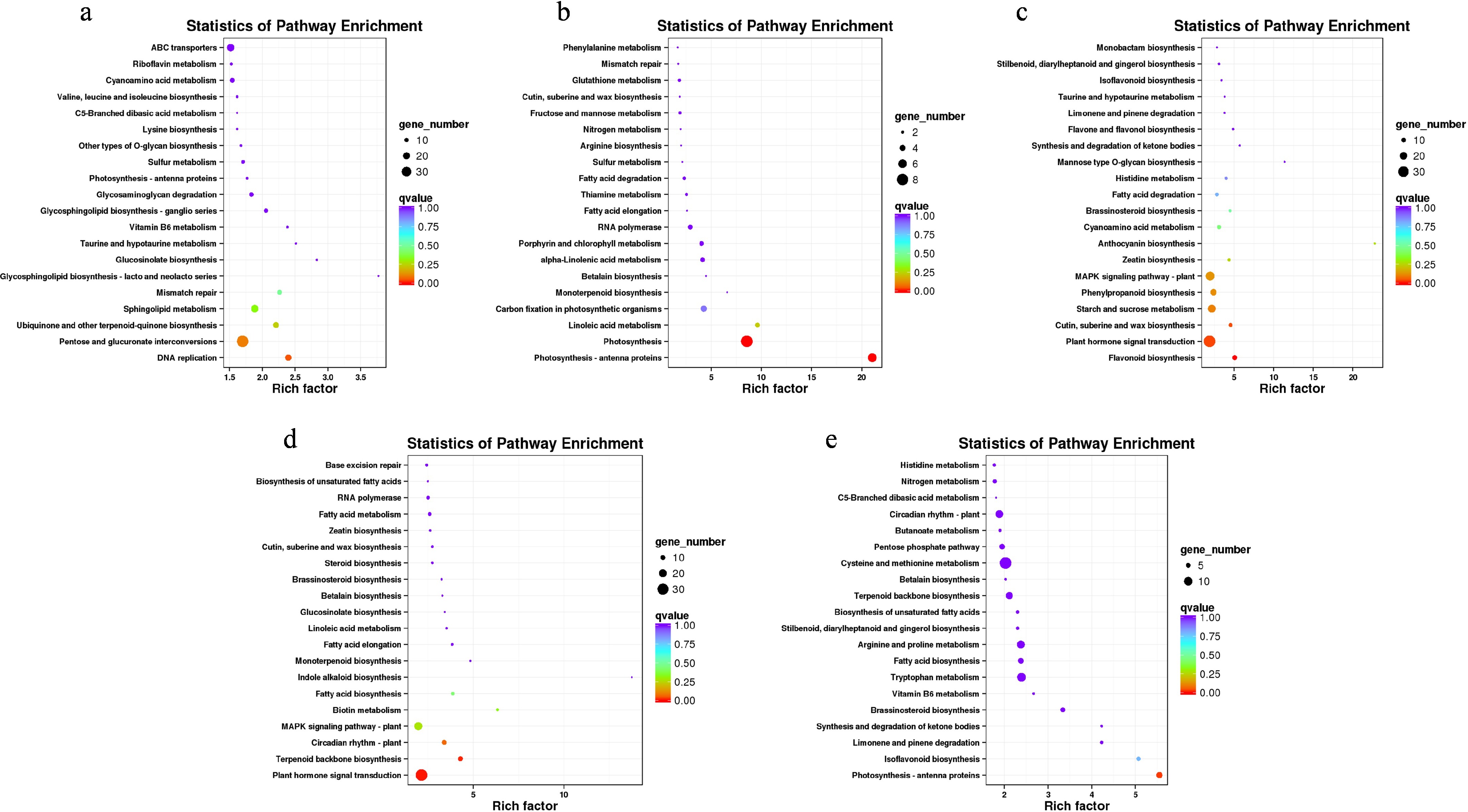
Figure 3.
Functional characterization of DEGs derived from five distinct pairwise comparisons under two experimental conditions in Dendrobium. (a) KEGG pathway enrichment profiling for DEGs identified in the S1 comparative analysis. (b) KEGG pathway enrichment profiling for DEGs identified in the S2 comparative analysis. (c) KEGG pathway enrichment profiling for DEGs identified in the S3 comparative analysis. (d) KEGG pathway enrichment profiling for DEGs identified in the S4 comparative analysis. (e) KEGG pathway enrichment profiling for DEGs identified in the S5 comparative analysis.
Importantly, plant hormone signal transduction and photosynthesis pathways were consistently enriched across all five comparisons, strongly suggesting that these two processes played central and sustained roles in regulating floral development under different temperature regimes. Additionally, GO enrichment revealed that the most significantly enriched GO term in the S3 comparison was 'regulation of transcription, DNA-templated', a result that mirrored the expression dynamics captured by the Mfuzz clustering analysis. This concordance indicated that S3 represented a developmentally critical phase marked by intensive transcriptional reprogramming (Supplementary Fig. S4).
DEGs associated with hormone biosynthesis and signaling
-
Given the well-established roles of plant hormones in floral induction, dormancy release, and subsequent flower development, and considering the KEGG enrichment results described above—which strongly suggested that hormone-related pathways might contribute to the regulatory responses triggered by low-temperature exposure—we further examined transcriptional alterations in phytohormone biosynthesis and signaling pathways under the two temperature regimes. DEGs associated with CK, GA, abscisic acid (ABA), and jasmonic acid (JA) pathways were systematically identified and analyzed to clarify their roles during floral development.
Genes encoding key enzymes responsible for CK biosynthesis and catabolism included one LONELY GUY (LOG) gene, and four cytokinin oxidase (CKX) genes. Under LT conditions, LOG transcript levels remained consistently low without substantial fluctuation throughout floral development, whereas under RT conditions, LOG expression increased sharply and reached its maximum in S3. Three CKX genes involved in CK degradation showed progressive induction under LT, peaking in S3 before declining afterwards. In the CK signaling cascade, we detected one authentic histidine phosphotransferase (AHP) and three response regulators (RRs), including both type-A and type-B RRs. AHP expression was markedly elevated in S3 under LT conditions, while two distinct expression peaks were evident in the RT group. RR genes exhibited reduced expression during later developmental stages following LT treatment, particularly after S3 (Fig. 4a).
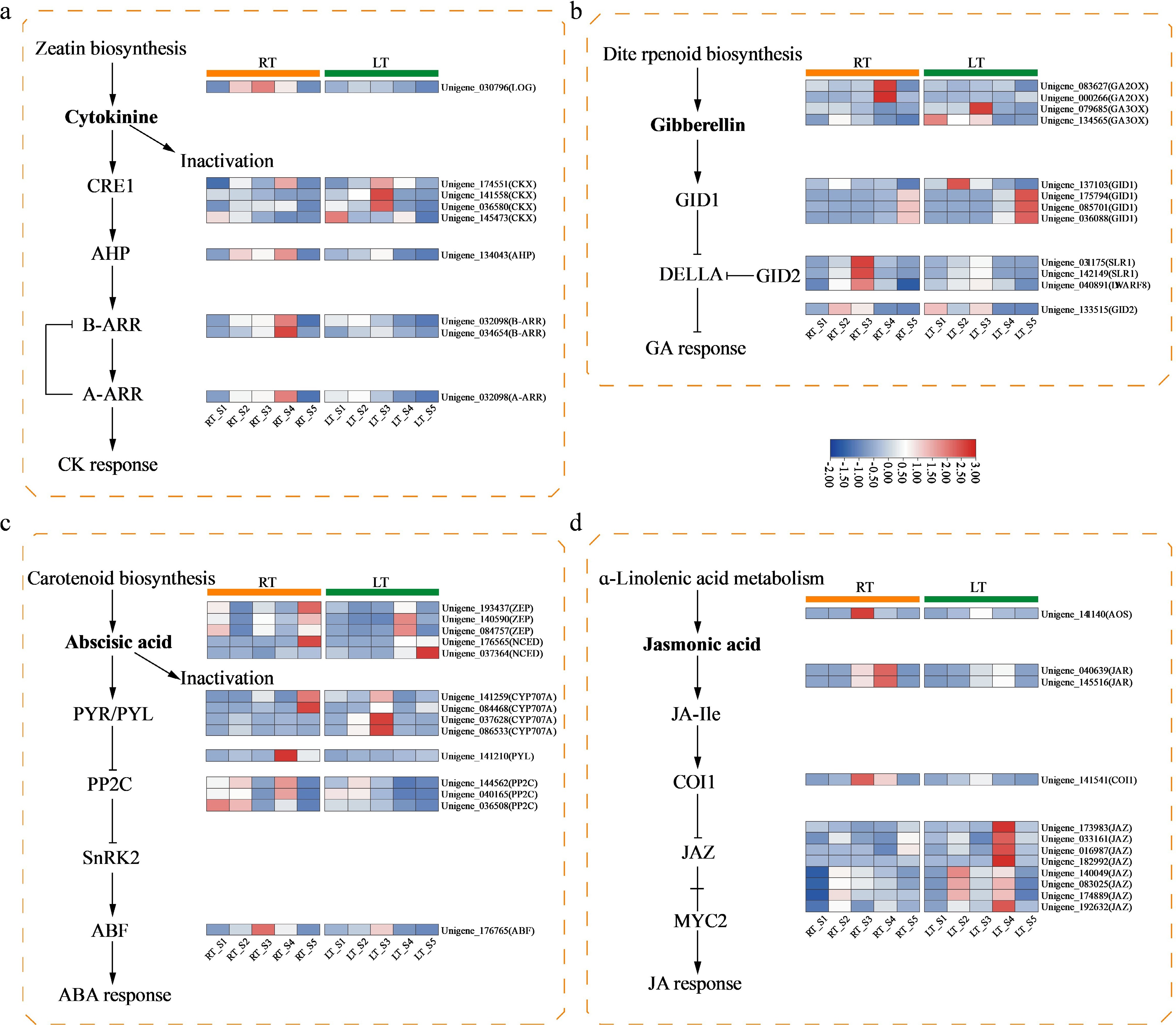
Figure 4.
Heatmap displaying DEGs associated with the biosynthesis and signal transduction pathways of plant hormones under two treatment conditions in Dendrobium. (a) DEGs involved in CK biosynthesis and signaling; (b) DEGs involved in GA biosynthesis and signaling; (c) DEGs involved in ABA biosynthesis and signaling; (d) DEGs involved in JA biosynthesis and signaling. Expression levels of DEGs were normalized to log2 counts based on FPKM values.
Within the GA biosynthesis pathway, two GA 2-oxidase (GA2ox) genes and two GA 3-oxidase (GA3ox) genes were identified. GA2ox, which catalyzes GA deactivation, showed minimal changes in expression under LT conditions. By contrast, GA3ox, an essential enzyme for generating bioactive GA, displayed enhanced transcriptional activity during early developmental stages under LT treatment. In the GA signaling pathway, four GIBBERELLIN INSENSITIVE DWARF1 (GID1) receptors, three DELLA proteins (DELLA), and one GIBBERELLIN INSENSITIVE DWARF2 (GID2) gene were detected. GID1 genes exhibited comparable expression dynamics under both temperature treatments. However, DELLA transcripts accumulated to higher levels in S3 under RT, suggesting more pronounced GA signaling repression in the absence of chilling. GID2 showed a dynamic pattern under LT: downregulated in S2, upregulated in S3, and subsequently reduced again from S3 through S5 (Fig. 4b).
In the ABA biosynthesis and catabolism pathways, we detected three zeaxanthin epoxidase (ZEP), two 9-cis-epoxy carotenoid dioxygenase (NCED), and four abscisic acid 8'-hydroxylase (CYP707A) genes. ZEP expression exhibited striking differences between treatments: ZEP genes were strongly induced in S4 under LT, whereas RT-grown plants showed three separate expression peaks in S1, S3, and S5. NCED genes, which mediate a key rate-limiting step in ABA biosynthesis, displayed similar increasing trends in both treatments during later stages. CYP707A genes, responsible for ABA hydroxylation and degradation, reached their highest expression in S3 under LT. Transcript analysis of ABA signaling components, one Pyrabactin resistance 1-like (PYL), three protein phosphatase 2C (PP2C), and one ABRE-BINDING FACTOR (ABF), revealed that these genes maintained higher expression levels after S3 under RT than LT (Fig. 4c).
For the JA biosynthesis pathway, one allene oxide synthase (AOS) and two JASMONATE RESISTANT1 (JAR1) genes were identified. AOS expression peaked prominently in S3 under RT conditions. JAR1, which catalyzes the formation of the bioactive isomer (+)-iso-JA-L-Ile, showed elevated expression from S3 to S4 under RT. In JA signaling, one CORONATINE INSENSITIVE1 (COI1) and eight JASMONATE ZIM DOMAIN (JAZ) genes were detected. JA signaling is initiated when the SCFCOI1 complex mediates the degradation of JAZ repressors, thereby releasing downstream transcription factors. Under LT conditions, COI1 expression was downregulated, while numerous JAZ genes were upregulated and accumulated to high levels during later developmental stages. (Fig. 4d).
DEGs associated with vernalization and cold-responsive genes
-
CBF/DREB–COR/LEA modules are central components of the plant cold-response network and are rapidly induced by low temperatures to enhance freezing tolerance. In this study, we identified two C-repeat binding factor (CBF) genes, five dehydration-responsive element-binding (DREB) genes, one Late Embryogenesis Abundant (LEA) gene, and two Dehydrin (DHN) genes. Among these, two CBFs, one DREB and one DHN gene displayed extremely high transcript abundance, specifically in S4 under RT conditions suggesting that normal-grown plants might activate alternative cold-independent stress pathways at this stage. By contrast, the remaining cold-responsive genes were strongly induced in S3 following LT treatment, consistent with canonical chilling-triggered transcriptional activation.
We also examined FRI and VIN3-like 2 (VIL2) homologs, which are known to participate in the vernalization pathway. Under LT conditions, FRI expression decreased sharply after S1 and remained at minimal levels during all subsequent stages, suggesting a potential release of floral repression analogous to the Arabidopsis vernalization mechanism. One VIL2 gene was markedly upregulated in response to chilling in the LT treatment, whereas two additional VIL2 homologs were induced during floral development under both temperature regimes, with peak expression observed in S3 (Fig. 5). These results collectively indicated that chilling activated multiple layers of the cold-response machinery while simultaneously modulating vernalization-associated components in Dendrobium.
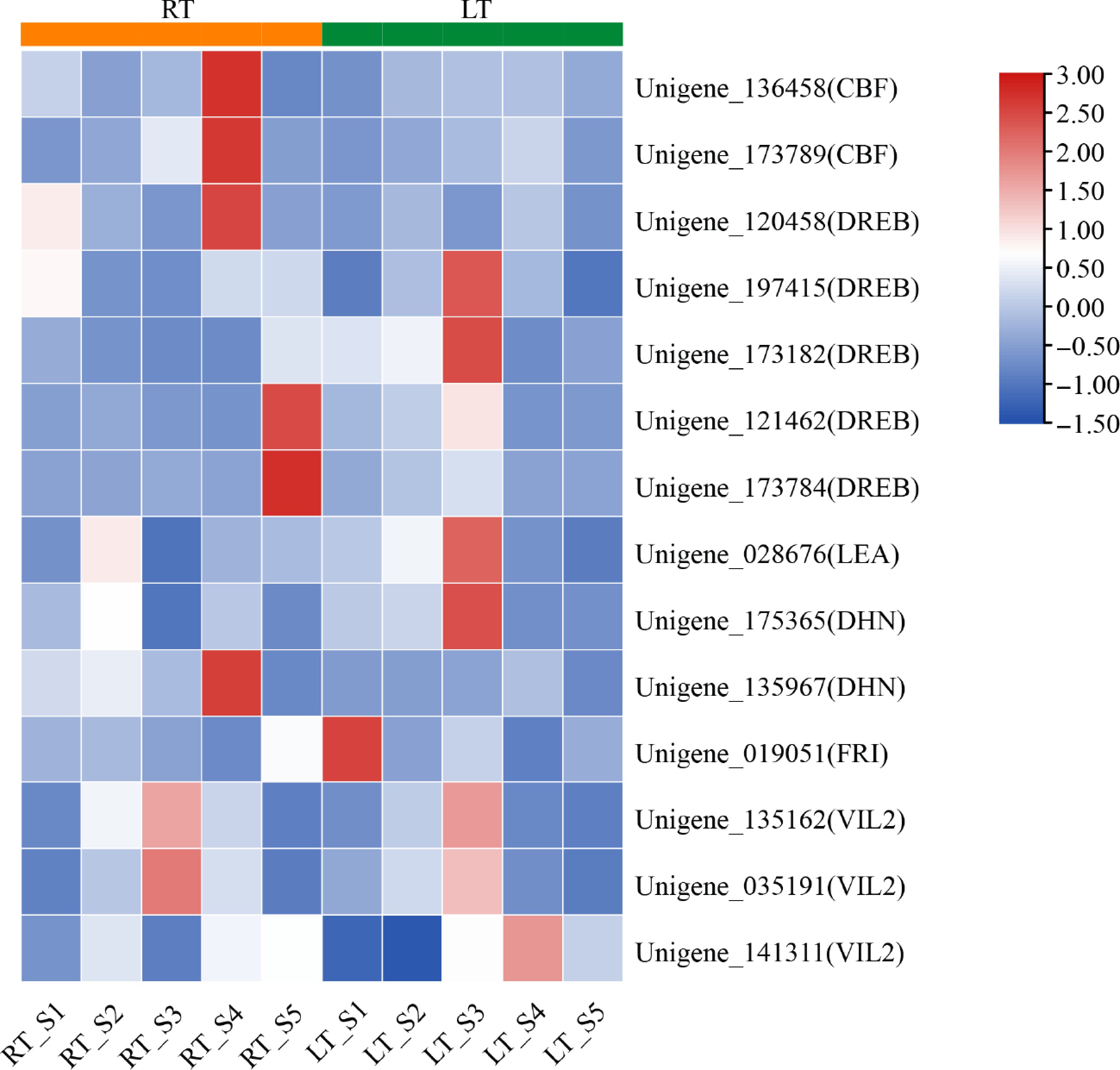
Figure 5.
Heatmap of DEGs associated with cold and vernalization responsive genes in Dendrobium. Expression levels were normalized to log2 counts based on FPKM values and visualized using a color gradient.
DEGs associated with flowering-related genes
-
To further dissect the transcriptional regulatory framework governing floral development, we analyzed 26 annotated genes associated with key flowering pathways. One TERMINAL FLOWER1 (TFL1) homolog, and two SHORT VEGETATIVE PHASE (SVP) genes exhibited consistent downregulation across both treatment groups, supporting their conserved roles as repressors of floral transition. In contrast, floral integrators and floral meristem identity regulators, including two FT, two TSF, one LFY, one AP1, and four CAULIFLOWER (CAL) homologs were progressively upregulated under LT conditions, with markedly elevated expression during the S3–S4 interval. Additionally, 12 MADS-box genes associated with floral organ identity were identified, including four B-class, four C-class, and one E-class homolog. Most of these genes exhibited increasing expression throughout development under both RT and LT conditions, although transcript levels were generally higher under RT (Fig. 6).
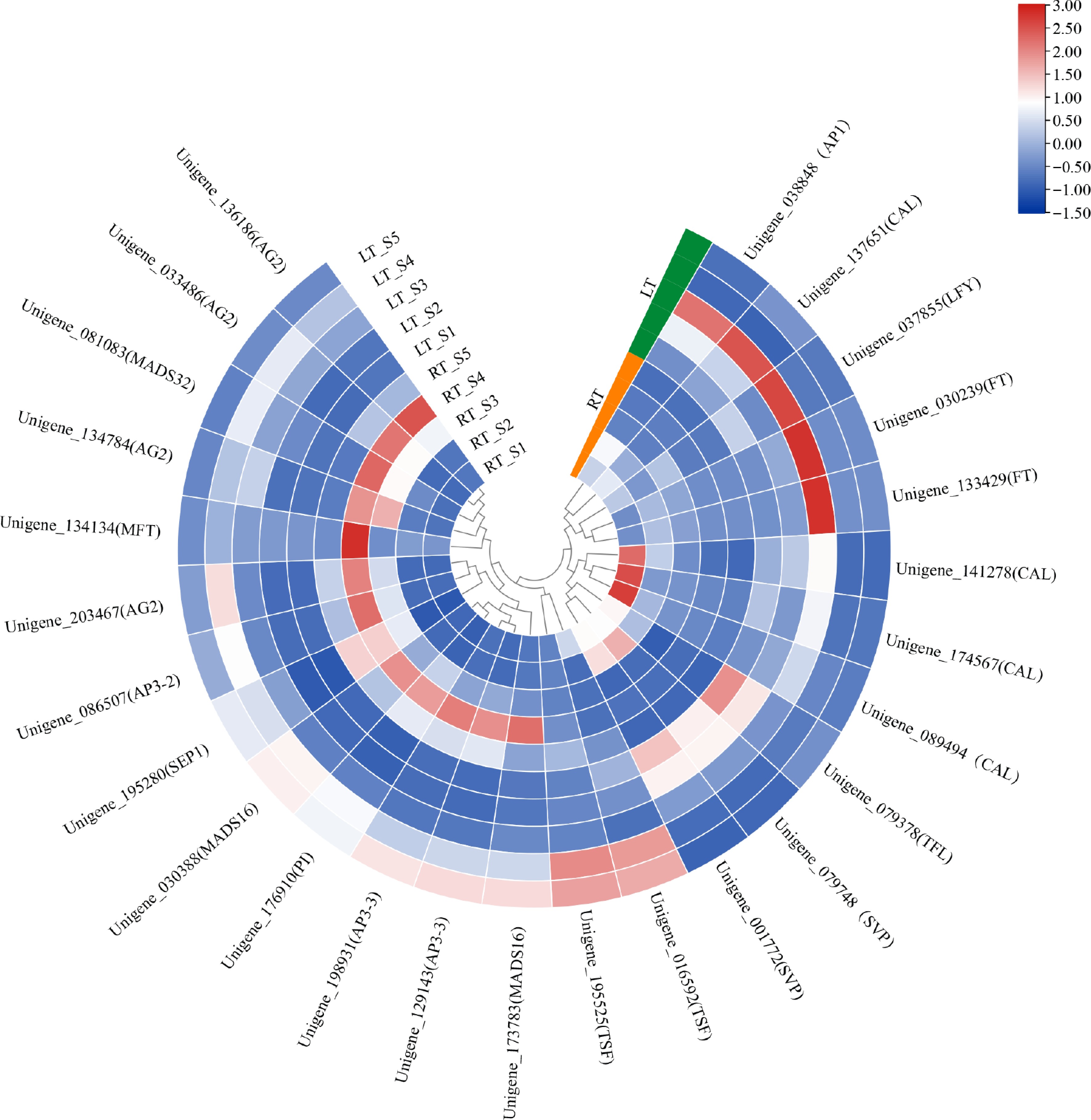
Figure 6.
Expression heatmap of a selected set of DEGs associated with flowering in Dendrobium. Expression levels were normalized to log2 counts based on FPKM values and visualized using a color gradient.
Validation of DEGs during flowering development
-
To verify the accuracy and reliability of the transcriptome sequencing results, 12 representative genes associated with the plant hormone signaling (LOG, GA3OX, CYP707A, PP2C, COI1), cold and vernalization pathway (LEA, DHN, FRI), and floral induction and meristem identity (FT, LFY, AP1, CAL) were selected for qRT-PCR validation. Under LT conditions, the expression levels of GA3OX, CYP707A, LEA, DHN, FT, LFY, AP1, and CAL were markedly elevated, reaching substantially higher transcript abundance in S3 compared with plants grown under RT conditions, further supporting the notion that chilling accelerated floral induction and enhanced key hormonal adjustments. Conversely, several genes involved in hormone signaling, specifically LOG and COI1, displayed reduced expression during the later developmental stages in the LT group relative to RT, indicating differential hormonal regulatory dynamics under cold exposure. In addition, FRI expression decreased abruptly after S1 under LT treatment, consistent with a chilling-associated release of floral repression. Overall, the qRT-PCR profiles closely mirrored the transcriptomic trends, providing strong validation of the RNA-Seq dataset and reinforcing the robustness of the gene expression analyses (Fig. 7).
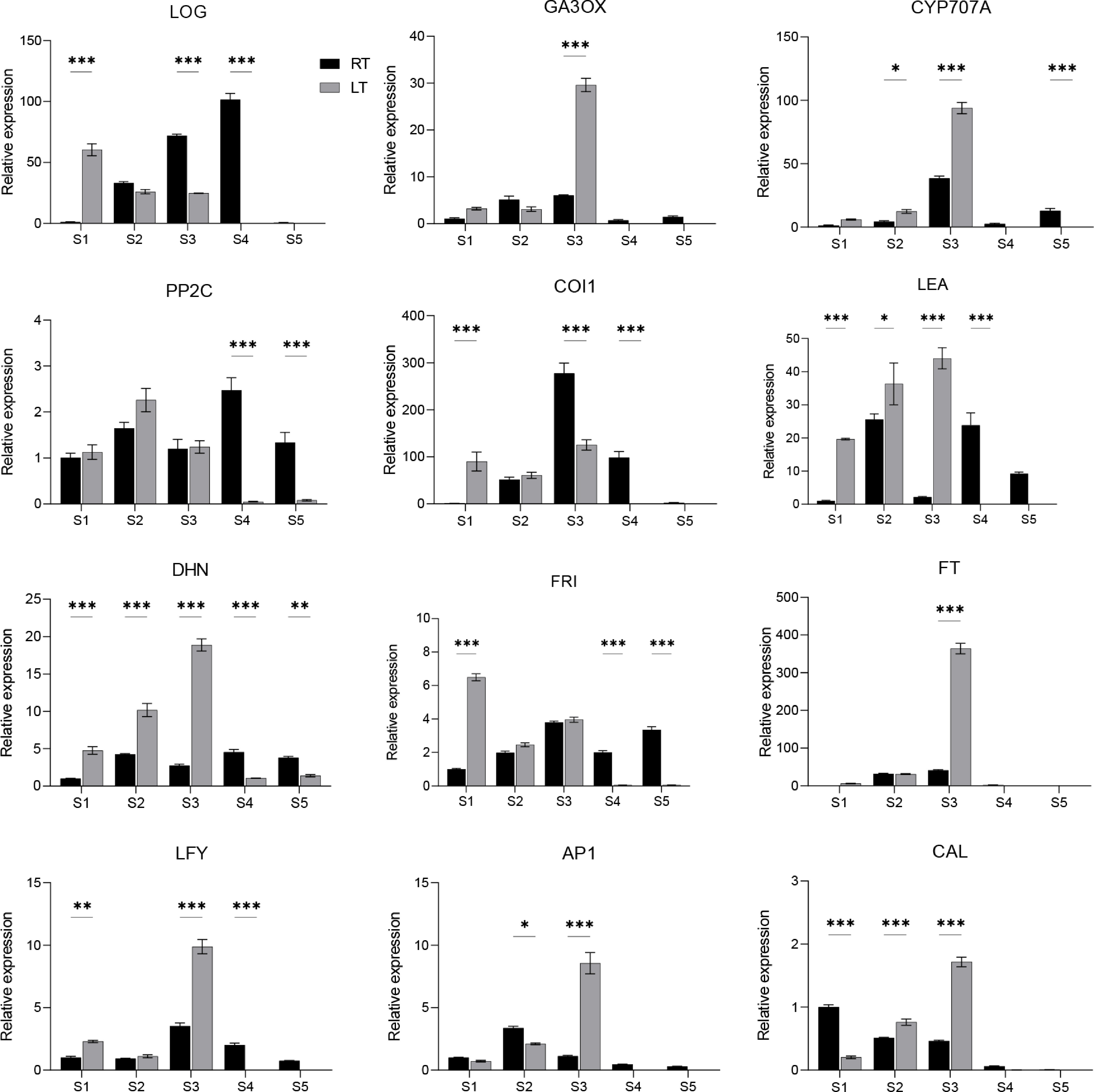
Figure 7.
RT-qPCR validation of selected candidate genes associated with flowering under RT vs LT conditions in Dendrobium. Data is shown as mean ± SD from three biological replicates. Asterisks indicate statistically significant differences relative to the RT group (* p < 0.05; ** p < 0.01; *** p < 0.001).
-
The transition from vegetative growth to reproductive development in plants is initiated by the formation of floral buds, a process that typically requires the accumulation of adequate nutritional reserves before induction occurs[21]. Extensive research has demonstrated that chilling exposure serves as a crucial environmental signal for dormancy release or flowering promotion in numerous plant species including litchi, blueberry, lily, wheat, and orchid taxa[5,22−25]. In the present study, we closely monitored and compared the flowering transition of Dendrobium grown under normal temperature conditions, and artificially imposed low-temperature regimes. Our observations revealed that artificial chilling markedly shortened the vegetative period and advanced flowering time by more than 1 month. Notably, despite the significant shift in phenology, no evident differences in the floral morphology or structural features were detected between chilled and control plants. This was consistent with previous physiological and molecular studies reporting that low-temperature exposure mainly accelerates the timing of floral induction rather than altering floral form[18,26−29]. Nonetheless, the degree to which chilling influences floral traits in orchids can vary substantially across species, largely depending on both the effective temperature threshold and the duration required for vernalization[30,16,17]. Overall, these findings corroborate the conclusion of this study that an appropriate range of low-temperature treatment primarily affected flowering time while exerting only a minor influence on overall floral quality. Even so, the precise developmental consequences of different chilling regimes in orchids remain incompletely understood and merit additional investigation.
In addition, the flowering process of Dendrobium observed here corresponded well with that reported for other orchid species. The developmental progression could be subdivided into five distinct and orderly stages: S1 (vegetative growth stage); S2 (floral initial differentiation stage); S3 (floral organ development stage); S4 (flower enlargement and development stage); and S5 (blooming stage)[18,26,31,32].
DEGs and KEGG enrichment analysis during the flowering process under RT vs LT conditions in Dendrobium
-
Previous studies have shown that winter cereals maintain a high-energy physiological status during cold periods by enhancing photosynthetic activity, which facilitates adaptation to low temperatures and supports reproductive development following vernalization[33]. Likewise, increased expression of photosynthesis-related genes during vernalization has been documented in orchard grass and has been implicated in the regulation of floral initiation and reproductive capacity[34]. In agreement with these findings, transcriptome comparisons across the five developmental stages in this study revealed pronounced enrichment of DEGs associated with photosynthetic processes, especially in S2. This suggested that artificial chilling might influence the reproductive trajectory of Dendrobium by modulating photosynthesis-related gene expression during the early phases of floral induction.
Plant hormones are recognized as central integrators of endogenous developmental signals and external environmental cues. Previous molecular research has firmly established that phytohormones and their downstream regulatory networks participate in nearly every phase of floral induction and reproductive development, functioning as key coordinators of the complex transition from vegetative growth to flowering[35].
Analysis of plant hormone-related gene expression
-
Phytohormones are widely regarded as central regulators of plant growth and development, functioning through intricate signaling cascades and gene regulatory networks. CK, for instance, interacts either synergistically or antagonistically with auxin to modulate floral bud initiation and subsequent development by coordinating cellular division, expansion, and differentiation[36]. Nonetheless, the precise roles of CK in floral initiation and floral organ differentiation remain contentious, as both their functional significance and temporal expression patterns exhibit marked species-specific variability[37,38]. In the present study, compared with plants grown under RT conditions, the LT-treated group displayed markedly lower expression of LOG and significant upregulation of CKX genes in S3. This suggested that artificial chilling might contribute to CK catabolism during floral development. Interestingly, our findings contrasted with earlier studies reporting activation of CK-related genes in cold-treated Dendrobium, a discrepancy that might be attributable to differences in chilling duration, sampling stages, or physiological states of the plant materials examined[19].
GAs are well-established promoters of floral transition, and their role in regulating flowering time has been documented in a wide range of plant species[27,32]. GA signaling exerts its influence primarily through DELLA proteins, which function as negative regulators and serve as hubs integrating GA availability with flowering-related transcriptional programs[39,40]. In this study, LT-treated plants exhibited enhanced expression of the GA biosynthetic enzyme GA3OX, together with reduced expression of the GA-deactivating enzyme GA2OX and decreased transcript accumulation of DELLA genes such as SLR1 and DWARF8. This transcriptional pattern indicated that low-temperature exposure promoted GA biosynthesis and attenuated GA repression, thereby contributing to the acceleration of flowering.
The function of ABA in flowering remains complex and species-dependent. Earlier work has demonstrated that ABA delays flowering in Arabidopsis, Chenopodium ficifolium, and rice[41−43], whereas it promotes floral transition in DE under long-day photoperiods[44]. In our study, ABA biosynthetic genes (ZEP, NCED) displayed distinct expression patterns under RT and LT treatments, with significantly elevated transcript levels during the later stages under LT. Several CYP707A genes, responsible for ABA catabolism, were strongly up-regulated in S3 under LT conditions. These results implied that chilling might delay ABA biosynthetic gene induction while enhancing ABA degradation during the critical stages of floral initiation and differentiation.
JA signaling is best known for its role in plant defense responses against abiotic and biotic stresses[45]. In JA signaling, JAZ repressors are degraded via the F-box protein COI1, allowing activation of downstream JA-responsive genes. In Arabidopsis, decreased JA levels elevate JAZ transcript abundance, and the resulting accumulation of JAZ proteins promote early flowering by interacting with transcriptional repressors TOE1 (TARGET OF EAT 1), and TOE2, thereby alleviating their repression of FT and accelerating floral induction[46,47]. Similar promotive effects of JAZ overexpression on flowering have been reported in tomato. Consistent with this mechanism, our results showed reduced expression of JAR1 and COI1 under LT treatment, accompanied by elevated expression of five JAZ genes during S4. These observations suggested that chilling suppressed JA biosynthesis and signaling, leading to JAZ accumulation that facilitates early flowering in Dendrobium.
Collectively, hormone homeostasis, maintained through the dynamic balance of biosynthesis and degradation, is essential for coordinating major developmental transitions[48]. Our results demonstrated that the induction of early flowering in Dendrobium under artificial chilling was driven by a concerted reprogramming of multiple hormone pathways, including enhanced expression of CK and ABA catabolic genes, activation of GA biosynthesis, and down-regulation of JA biosynthetic components. These integrated hormonal adjustments formed a crucial regulatory basis for chilling-induced floral acceleration.
Analysis of cold and vernalization-responsive gene expression
-
The genetic architecture underlying cold acclimation and vernalization-induced flowering has been extensively characterized in several model and crop species. These processes are orchestrated by multilayered and often interconnected regulatory systems encompassing diverse gene families and signaling modules[49−52]. At the core of the cold-response framework are CBF/DREB transcription factors, members of the AP2/ERF superfamily, which function as master regulators of freezing tolerance. These transcription factors activate a broad spectrum of cold-inducible genes to enhance cold adaptation in numerous species such as Phalaenopsis aphrodite, Arabidopsis, and alfalfa[53−55]. Consistent with these findings, our study detected pronounced induction of four DREB, one LEA, and one DHN gene under low-temperature conditions, supporting their conserved involvement in chilling responses in Dendrobium.
Genetic studies in Arabidopsis have established FRI and FLC as central repressors of the vernalization-induced flowering pathway[56,57], with VIL2, a component of a VIN3-like (VIL1-containing) chromatin-remodeling complex, contributing to the epigenetic suppression of FLC[8]. In agreement with previous reports in Dendrobium, no FLC homolog was identified in our dataset, suggesting that flowering regulation in Dendrobium operated through an autonomous mechanism distinct from the canonical FRI–FLC module, widely observed in perennial temperate species. Homologs of one FRI and three VIL2 genes were detected. Among these, FRI expression declined substantially under LT conditions, whereas two VIL2 genes exhibited comparable expression dynamics in both RT and LT groups, implying that Dendrobium might utilize a modified or partial vernalization-associated regulatory program.
Studies on cold tolerance mechanisms have increasingly emphasized that cold acclimation and vernalization are functionally intertwined processes. In accordance with this conceptual framework, our results revealed that the expression profiles of cold-responsive genes and those associated with vernalization exhibited notable overlap during chilling exposure, reinforcing the idea that these pathways converge to facilitate low-temperature-induced flowering in plants[24,58].
Analysis of flowering-related gene expression
-
Within the flowering regulatory network, both endogenous developmental cues and environmental stimuli are integrated to modulate the expression of downstream genes essential for floral initiation and meristem specification. Consistent with established genetic models, TFL and SVP function as major repressors of flowering, and their transcript levels progressively decrease during the floral transition. In this study, homologs of TFL and SVP were identified. These genes maintained similar expression patterns under both normal and chilling treatments, suggesting that these genes retained conserved roles, but might not serve as primary determinants of chilling-induced floral acceleration in Dendrobium.
A key observation was the strong accumulation of FT transcripts in S3 and S4 under LT conditions. It is widely accepted that FT acts as a central integrator linking upstream environmental and developmental signals to the activation of floral meristem identity genes[59,60]. Once repression by upstream inhibitors such as FRI is alleviated, elevated FT promotes the expression of LFY, AP1, and CAL, initiating the developmental program that leads to floral organ formation. Consistent with this model, our transcriptomic analyses revealed that LFY, AP1, and CAL were preferentially expressed in S3 under LT, mirroring the expression trajectory of FT. These findings collectively suggested that low-temperature exposure suppressed FRI, enabling the upregulation of floral integrators and meristem identity genes, thereby driving the floral transition in Dendrobium.
Despite this progress, the precise molecular crosstalk linking temperature perception and phytohormone signaling during floral induction in Dendrobium remains insufficiently understood. Further studies dissecting the integration of these pathways will be essential for a more complete mechanistic understanding of chilling-induced flowering in this genus.
-
Low temperature is a pivotal environmental determinant of flowering in Dendrobium and has profound effects on subsequent growth performance and yield. In this study, we integrated comprehensive transcriptomic profiling with detailed microscopic observations to elucidate the molecular framework governing chilling-induced flowering in this species. Our results revealed that artificial chilling markedly accelerated the floral transition by reshaping phytohormone homeostasis and inducing substantial transcriptional reprogramming of cold-responsive, vernalization-associated, and flowering-related genes during floral organ development. Collectively, these findings advance current understanding of the genetic and regulatory basis underlying low-temperature-mediated flowering, and provided valuable guidance for optimizing flowering time in nobile-type Dendrobium under diverse environmental and horticultural conditions (Fig. 8).
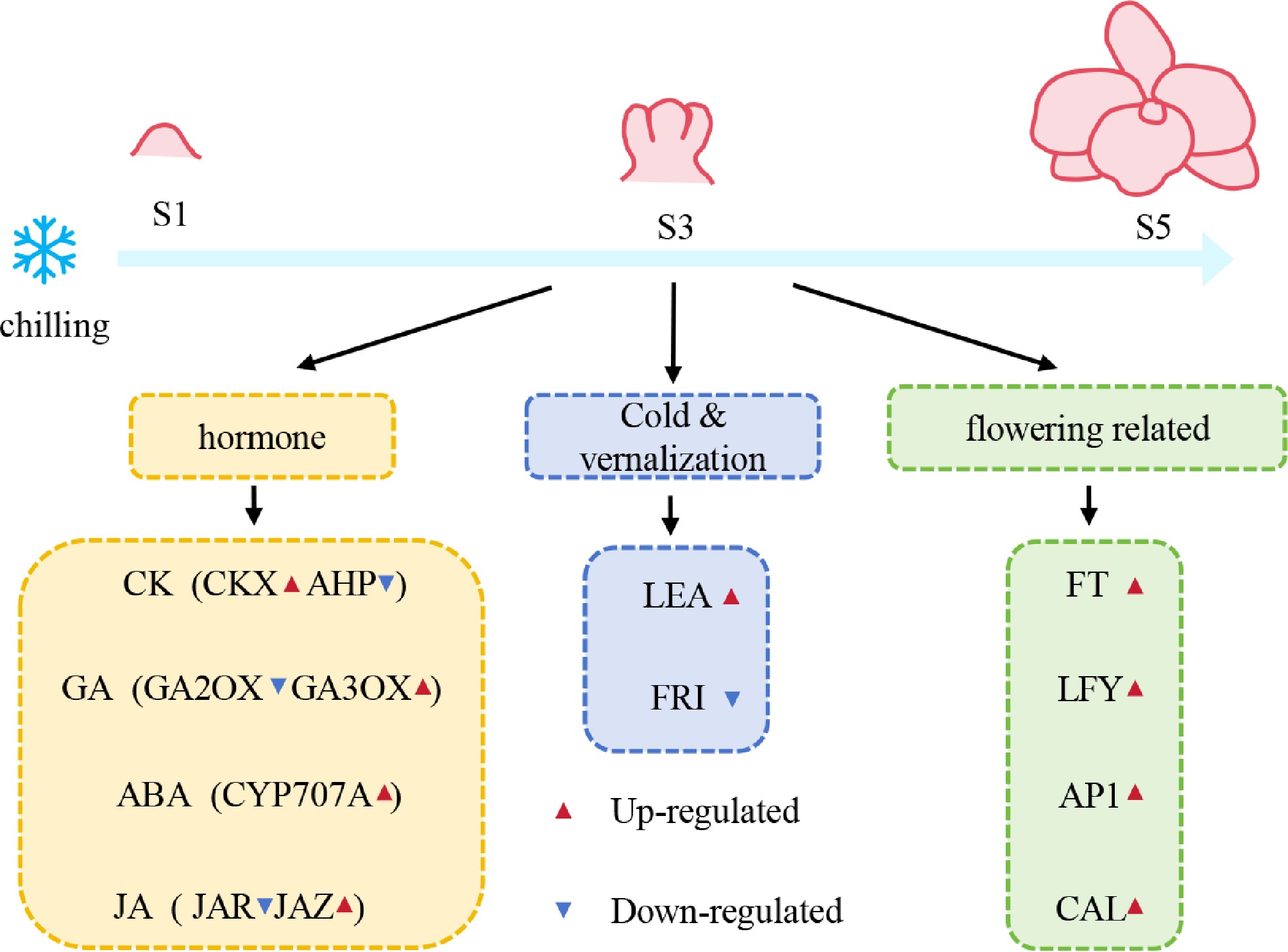
Figure 8.
Schematic model for the gene regulatory network of chilling-mediated flowering in Dendrobium.
This research was supported by the Provincial Rural Revitalization Strategy of Dongguan in 2021 (20211800400062), the Innovation Team of the Modern Agriculture Industry Technology System in Guangdong Province (2024CXTD12) and Modern Seed Industry Innovation Capability Enhancement Project of Guangdong Academy of Agricultural Sciences (2025ZYTS). The authors gratefully acknowledge these funding sources for enabling the completion of this work.
-
The authors confirm that their contributions to this work are as follows: study conception and experimental design: Lu C, Zhu G, Yang F, Zheng G; sample preparation and transcriptome sequencing: Lu C, Wu J, Li J, Jin J; data analysis and interpretation: Lu C, Wu J, Lin Z, Li S, Zhang K, Wei Y, Xie Q; manuscript drafting and initial preparation: Lu C, Gao J, Lin Z. All authors reviewed the data, contributed to manuscript revisions, and approved the final version for publication.
-
The datasets generated and analyzed during the current study are not publicly available at this time, as the research group intends to publish additional findings derived from the dataset. However, the data is available from the corresponding author upon reasonable request.
-
The authors declare that they have no competing financial or non-financial interests related to the work presented in this manuscript.
-
# Authors contributed equally: Chuqiao Lu, Jieqiu Wu
- Supplementary Table S1 Primers used for qRT-PCR.
- Supplementary Table S2 Effect of temperature treatment on flowering of Dendrobium.
- Supplementary Table S3 Sample sequencing data evaluation statistics table.
- Supplementary Table S4 Total number of transcripts and unigenes identified in the transcriptome.
- Supplementary Fig. S1 Morphology change of complete process of flower development in D. nobile.
- Supplementary Fig. S2 Overview of de novo assembled unigenes in Dendrobium based on transcriptomic data.
- Supplementary Fig. S3 Correlation analysis and principal component analysis (PCA) of all transcripts and expressed genes in D.nobile.
- Supplementary Fig. S4 Functional analysis of DEGs from samples at five different flower development stages of D. nobile.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lu C, Wu J, Gao J, Lin Z, Li S, et al. 2026. Comparative transcriptomic analysis identifies key regulatory factors in low temperature-induced flowering in Dendrobium. Ornamental Plant Research 6: e017 doi: 10.48130/opr-0026-0006 

Comparative transcriptomic analysis identifies key regulatory factors in low temperature-induced flowering in Dendrobium
- Received: 07 September 2025
- Revised: 13 December 2025
- Accepted: 06 January 2026
- Published online: 14 May 2026
Abstract: Low temperature is a key environmental cue governing floral transition and inflorescence development in nobile-type Dendrobium, and chilling treatments are routinely applied in horticultural practice to manipulate flowering time. Nevertheless, a robust genetic strategy for modulating flowering independent of temperature signals has yet to be established in this genus. To elucidate the molecular framework through which plants perceive and respond to chilling, we integrated cytological analyses with RNA sequencing (RNA-seq) to characterize transcriptional dynamics and chilling-responsive pathways under different low-temperature regimes. Continuous exposure to chilling markedly hastened floral induction, advancing flowering by nearly 1 month relative to plants maintained under standard growth conditions. Comparative transcriptomic analysis identified 13,708 differentially expressed genes (DEGs) between chilled and control plants. KEGG enrichment revealed strong overrepresentation of pathways related to photosynthesis and plant hormone signal transduction. Further analyses indicated that chilling triggered extensive reprogramming of hormone-related, cold-responsive, and floral-developmental genes. Specifically, low temperature reshaped hormonal homeostasis by elevating transcripts associated with CK and ABA catabolism, stimulating GA biosynthetic enzymes, and repressing JA biosynthesis. Concurrently, chilling suppressed FRI expression, while activating key cold-responsive genes, floral integrators, and meristem identity regulators, including LEA, FT, LFY, AP1, and CAL, collectively accelerated floral transition and organ initiation. These results not only offered actionable insights for horticultural control of flowering time, but also provided a valuable genetic resource for understanding chilling-induced flowering in nobile-type Dendrobium.
-
Key words:
- Dendrobium /
- Chilling /
- Transcriptome analysis /
- Flowering /
- Phytohormone