








-
Tocochromanols, including tocopherols and tocotrienols, are a class of lipid-soluble, plastid-synthesized antioxidants present in all plant tissues. They are essential in the human diet and are known as vitamin E (VTE). In plants, they play a crucial role in physiological homeostasis, primarily as antioxidants and regulators of stress responses[1]. Four forms of tocopherols and tocotrienols are produced by plants (α, β, δ, and γ), and they differ in the positions and numbers of methyl groups added to their chromanol head group (Fig. 1). Studies have shown that the antioxidant capacity of tocopherols to different plant oils mainly depends on the storage temperatures[2]. Besides their role as antioxidants, tocochromanols may interplay with plant hormones influencing plant growth, stress adaptation, and metabolic processes (reviewed by Müller & Munné-Bosch and Sadiq et al.[3,4]). Recent studies have provided insights into this crosstalk, highlighting how tocochromanols modulate and are modulated by phytohormones such as abscisic acid (ABA), salicylic acid (SA), jasmonic acid (JA), auxins, and ethylene. Moreover, a recent review from Kukri et al.[5] highlights about major advances in knowledge on putative α-tocopherol-phytohormones pathways. Here, we critically examine the emerging evidence on the interactions between tocochromanols and plant hormones from a mechanistic point of view, focusing on their roles in stress responses and growth regulation.
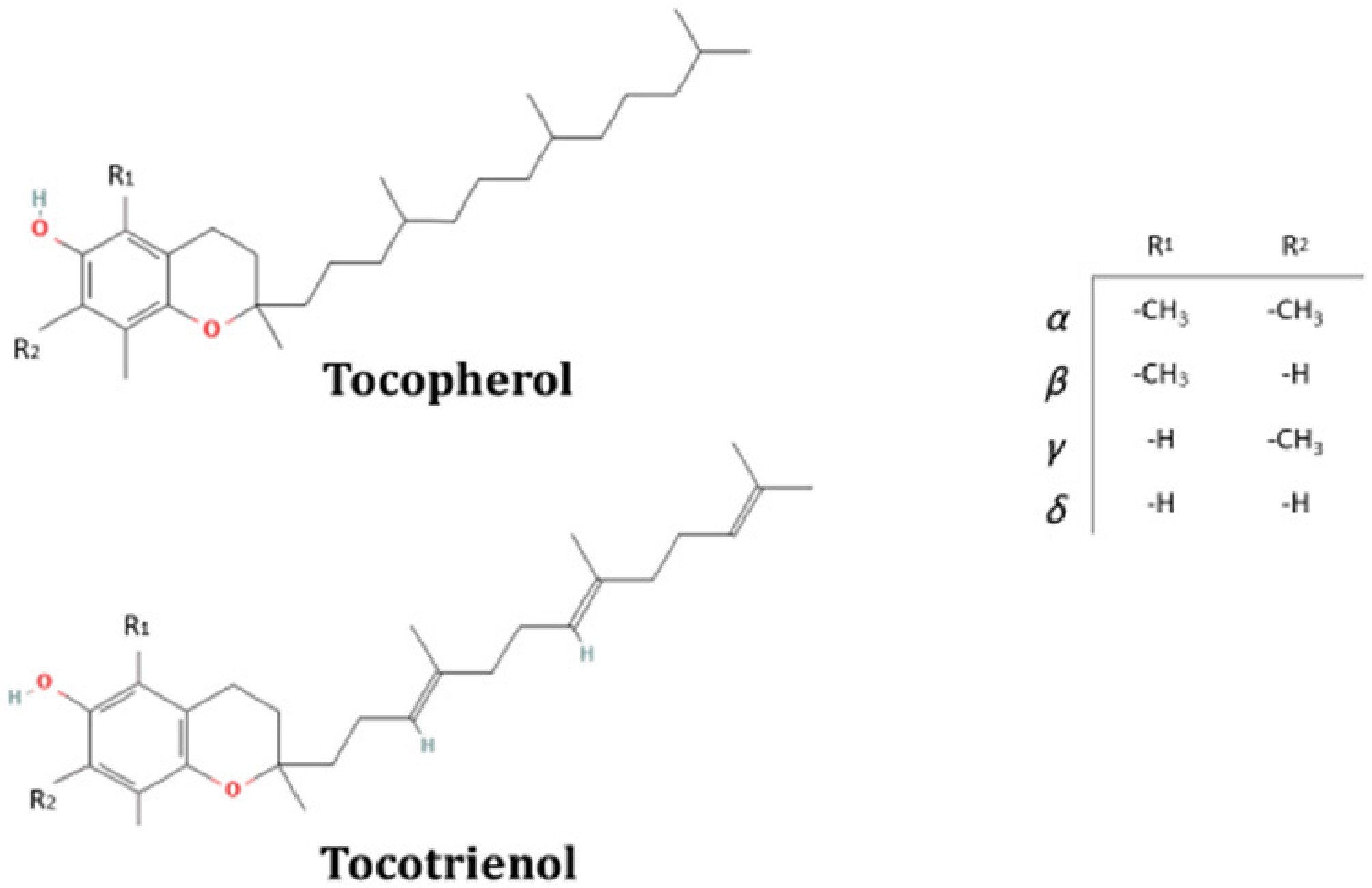
Figure 1.
Tocochromanols molecular structure: A chromanol head (with the differences of α, β, γ, and δ corresponding to the number or position of methyl groups in R1 and R2) and a tail of prenyl (non-saturated and with three unsaturation –3', 7', and 11'- for the cases of tocopherols and tocotrienols, respectively). Figure adapted from PubChem.
-
Biosynthesis of tocochromanols in plant cells starts in the cytoplasm, and the intermediate and the final steps of their synthesis take place in plastids, where enzymes are localized at the inner envelope or in the plastoglobules[6]. Tocotrienols and tocopherols tails derive from geranylgeranyl diphosphate (GGDP) and from phytyl diphosphate (PDP), respectively. Both are intermediates of the methylerythritol-phosphate (MEP) pathway. Polar chromanol head group derives from homogentisate (HGA), whose synthesis occurs in the shikimate pathway from hydroxyphenylpyruvate by the action of the hydroxyphenylpyruvate dehydrogenase (HPPD)[7]. An additional pathway for PDP production from chlorophyll degradation, also known as the phytol recycling pathway, has been more recently proposed[8]. Condensation of HGA with the corresponding tail is catalyzed by the homogentisate phytyltransferase (VTE2)[9]. Then, the 2-methyl-6-phytyl-1,4-benzoquinol (MPBQ) methyltransferase (VTE3) methylates MPBQ into 2,3-dimethyl-6-phytyl-1,4-benzoquinone (DMPBQ)[10]. Tocopherol cyclase (VTE1) closes the second ring, yielding δ-tocopherol and γ-tocopherol from MPBQ and DMPBQ, respectively[11]. Lastly, the δ-tocopherol and γ-tocopherol methylation is catalyzed by γ-tocopherol methyltransferase (VTE4), producing β- and α-tocopherol, respectively[12] (Fig. 2). In the absence of stress, tocochromanols prevent the excessive accumulation of ROS produced by photosynthesis, which could cause damage to biomolecules. More recent studies have emphasized tocochromanols' involvement in retrograde signalling pathways, which integrate external stress signals to regulate nuclear gene expression[13,14]. These examples extend the role of these lipophilic antioxidants beyond protection against oxidative damage, as they imply a fundamental function in cellular signalling and gene expression modulation.
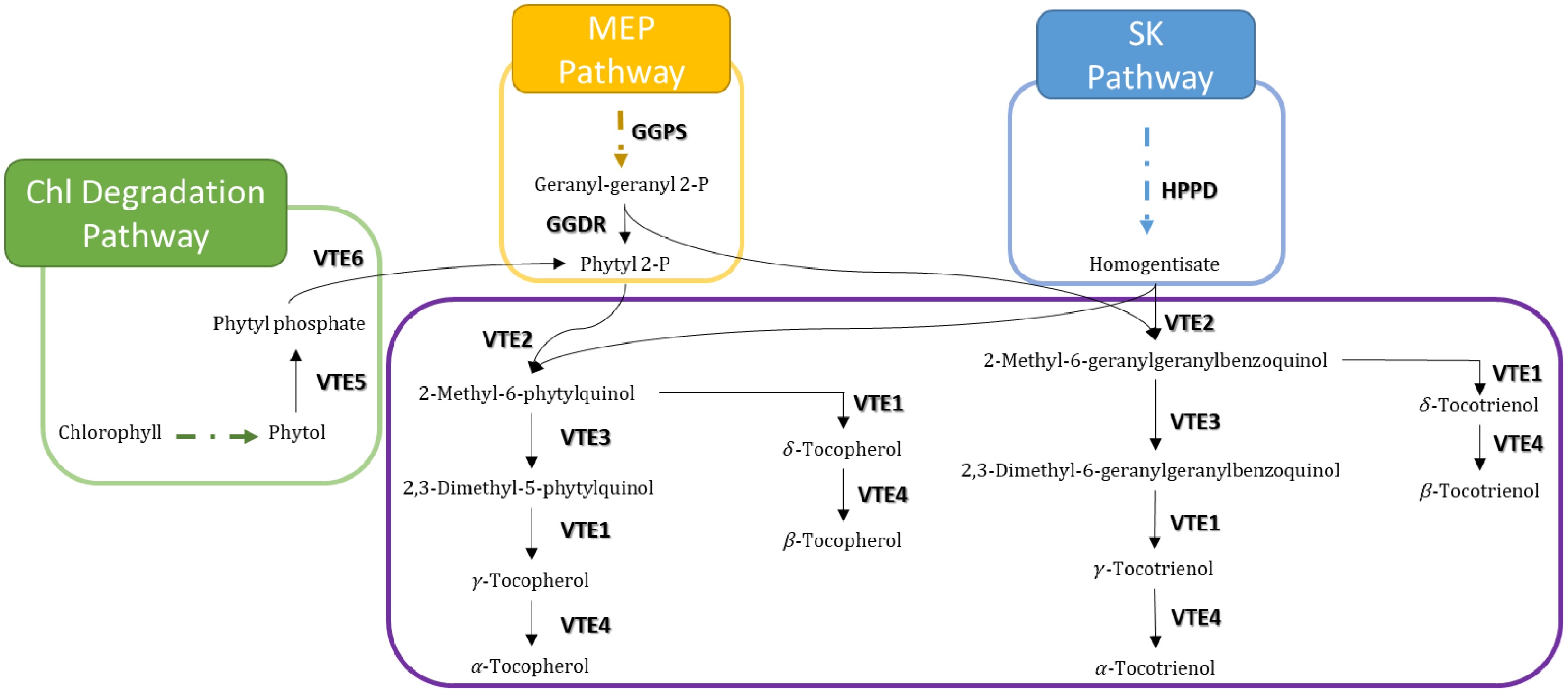
Figure 2.
Tocochromanol biosynthetic pathway. Both tocopherol and tocotrienols are synthesized by the combination of homogentisate and phytyl diphosphate (phytyl 2-P)(tocopherols) or geranyl-geranyl diphosphate (geranyl-geranyl 2-P) (tocotrienols). Violet, blue, yellow, and green boxes depict the tocopherol-core, shikimate (SK), methylerythritol-phosphate (MEP), and chlorophyll degradation pathways, respectively. Filled and dotted arrows mean direct and two or more intermediate enzymatic steps, respectively.
-
ABA is a small sesquiterpene molecule that regulates plant growth and resilience[15]. It was first identified in the 1960s as a growth inhibitor of cotton fruit and leaves of sycamore[16−18]. Since then, it has been shown that this hormone not only regulates plant growth and its responses to environmental stresses but also almost all plant phenological stages; from embryo maturation, seed dormancy, germination, cell division and elongation to floral induction and fruit ripening[19].
First steps of ABA biosynthesis take place in the plastid and begin with the ABA and carotenoids common precursor GGDP from the MEP pathway. GGDP is converted to carotenoids, including violaxanthin and neoxanthin, which subsequently undergo cleavage by 9-cis-epoxycarotenoid dioxygenase (NCED), the rate-limiting enzyme, to produce xanthoxin. This plastid-derived xanthoxin is then transported to the cytosol, where it undergoes sequential oxidation via alcohol dehydrogenase and aldehyde oxidase to yield mature ABA. This compartmentalized biosynthetic pathway exemplifies the integration of photosynthetic metabolism with stress hormone triggering, as environmental stresses such as drought upregulate NCED expression, thereby channelling carotenoid precursors toward ABA synthesis rather than photoprotection, ultimately enabling rapid hormonal responses to environmental challenges[20]. In line with this, enzymes of the plastidial MEP pathway are tightly regulated and highly responsive to environmental cues[21]. Abiotic stresses modulate MEP pathway activity, thereby affecting the availability of GGDP and its partitioning among carotenoids, tocopherols, and ABA biosynthesis. This dynamic control of isoprenoid flux enables plants to prioritize ABA production during stress while preserving plastidial redox balance and photoprotection. Despite this apparent crosstalk at the level of the early common precursor GGDP, there is substantial evidence linking ABA signalling to the regulation of tocopherol biosynthesis. Positive correlations between ABA levels and tocochromanol contents have been reported[22]. Buchner et al.[23] showed that α-tocopherol levels rise under heat and drought stress in Senecio incanus and Primula minima, overlapping with increased ABA concentrations, a correlation also observed by Kukri et al.[5] in the succulent plant Sempervivum tectorum L. Kreszies et al.[24] recently confirmed that tocopherol biosynthesis increases in Arabidopsis under drought and osmotic stress, but not under high light or nitrogen deprivation, and that this increase is ABA-dependent, correlating with higher expression of VTE2 and HPPD genes. Such up-regulation in the VTE2 gene expression was also observed in the drought-resistant shrub Cistus creticus, along with a simultaneous increase in ABA and α-tocopherol levels[25]. However, in Arabidopsis exposed to fungal volatile compounds, both ABA and tocopherol levels decrease[26], indicating that environmental context can modulate this interplay.
In Arabidopsis, ABA treatment increases tocopherol accumulation and upregulates several genes (VTE1, VTE4, and HPPD) of the tocopherol biosynthetic pathway[22]. Consistently, ABA-responsive promoter motifs have been identified in the rice genes OsHPPD, OsγTMT (VTE4), and OsMPBQMT1 (VTE3)[27], suggesting direct transcriptional control of tocopherol biosynthesis by ABA. In tomato, Quadrana et al.[28] identified shared promoter domains between several VTE genes (SlVTE4, SlVTE3, SlVTE1, SlHPPD, and SlGGPS) and ABA-related genes, including a binding motif for the transcription factor ABI4, a key regulator of ABA signal transduction[29]. Jiang et al.[30] demonstrated that ABA modulates tocopherol production by regulating HPPD expression, while Tewari et al.[31] identified a cis-regulatory element responsive to ABA in the promoter region of VTE4, highlighting the connection between ABA and VTE metabolism in soybean. These genetic proofs suggest a direct regulation of ABA on VTE biosynthetic genes. However, accumulating experimental evidence suggests that tocochromanols participate in feedback regulation in ABA pathways. Endogenous ABA levels are elevated in Arabidopsis vte1 and vte4 mutants, particularly in roots, and salinity stress further enhances ABA accumulation in vte1[32]. Conversely, overexpression of the VTE4 gene in alfalfa results in reduced ABA levels and repression of ABA biosynthetic (ABA1, ABA3, NCED3) and signalling (ABI3, ABI4) genes, indicating feedback regulation between tocopherol accumulation and ABA pathways[14]. Wang et al.[33] further showed, both in sunflower and Arabidopsis, that VTE1 overexpression results in ABA insensitivity by blocking ABI5 gene expression and by scavenging superoxide contents. On the other hand, Medicago truncatula seeds of the abi5 mutant accumulate higher levels of α-tocopherol, linking ABA signalling competence to tocopherol homeostasis during germination[34]. Together, these data support a bidirectional regulatory relationship: tocochromanols modulate ABA levels and sensitivity, and ABA in turn regulates tocochromanol accumulation.
Salicylic acid (SA) and defense mechanisms
-
SA, first isolated in the 19th century from willow bark[35], is now recognized as a central phytohormone involved in both plant development and stress adaptation. SA regulates key physiological processes—including seed germination, vegetative growth, photosynthesis, flowering, senescence, and thermogenesis—and plays an essential role in mitigating abiotic stresses such as heat, cold, drought, UV radiation, heavy metal exposure, and osmotic shock[26]. Supporting a transcriptional connection with tocochromanol biosynthesis, Tewari et al.[31] found a SA-responsive cis-regulatory element in the promoter region of Glycine max-VTE4 gene. However, Kusajima et al.[36] demonstrated that heat shock-induced tocopherol biosynthesis occurs independently of SA-mediated signalling, suggesting an alternative regulatory mechanism operating under thermal stress. Stahl et al.[37] showed that vte2 Arabidopsis mutants, which are impaired in tocopherol synthesis, exhibit stronger lipid peroxidation and produce less SA, leading to a higher susceptibility to P. syringae. This demonstrates that tocopherol biosynthesis is induced in Arabidopsis upon bacterial infection, and that this induction in turn promotes SA accumulation and enhances basal immunity. Chan[38] reported that seeds of two Arabidopsis SA-deficient mutants, NahG and sid2, accumulate higher tocopherol levels than the wild-type plants, indicating that SA may repress tocopherol synthesis in seeds. However, caution should be taken with these results since auxin levels are also altered in sid2 seed mutants. The authors suggest a negative feedback loop operating in seeds that is not observed during pathogen challenge in leaves. Thus, the relationship between SA and tocopherol metabolism appears to be organ dependent.
Jasmonic acid (JA) and stress signalling
-
JA belongs to the family of oxidized fatty acids and is widely detected across the plant kingdom[39]. JA and its derivatives (collectively known as jasmonates) play central roles in plant adaptation to both biotic and abiotic stresses, and they regulate essential physiological processes including leaf expansion, cell cycle progression, and stomatal dynamics[40,41]. It is well known that JA and Methyl-JA modulate the endogenous level of tocopherol in plants[42,43]. Such regulation operates through the ability of JA to up-regulate genes implicated in the tocopherol-biosynthetic pathway, including HPPD and HPT (homogentisate phytyltransferase) genes[44]. Along the same line, Quadrana et al.[28] identified shared promoter motifs among VTE1, VTE3, VTE4, HPPD, and GGPS genes in tomato, associated with the transcription factor HAHB4, a regulator that integrates JA and ethylene signals during wounding and biotic stress responses[45]. Also, JA-insensitive tomato mutant plants accumulate higher amounts of tocopherols compared to their WT counterparts, and this was accompanied by an elevated expression of the VTE4 gene[46], indicating that active JA signalling normally represses VTE4 expression in fruit. More recently, Zeng et al.[47] identified JA-responsive regulatory elements (and also ABA, SA, auxins, and gibberellins) in the VTE2/HPT gene promoter, in barley, wheat, oat, and Brachypodium, suggesting transcriptional JA-mediated regulation of tocochromanol biosynthesis across multiple species. Allu et al.[48] found that VTE-deficient Arabidopsis mutants accumulate higher JA levels under low phosphate availability, providing genetic evidence linking tocopherol status to the JA-mediated defense pathway. An earlier work by Ellouzi et al.[32] also indicated that α-tocopherol can modulate JA levels, directly affecting plant growth and stress tolerance. Moreover, Casadesús et al.[41] reported that the application of a protein-based biostimulant increased both JA and tocopherol levels in water-stressed tomato plants, suggesting a coordinated hormonal and antioxidant response during stress adaptation.
Ethylene signalling and tocochromanol metabolism
-
Ethylene is a gaseous phytohormone widely released by fruits and vegetables and plays a central role in regulating plant growth, fruit ripening, seed germination, and senescence[49,50]. In addition to its developmental functions, ethylene release is triggered by various environmental stresses, including metal toxicity, flooding, drought, and pathogen attack, positioning it as a key mediator of stress adaptation[51]. Its biosynthesis shares a precursor with α-tocopherol: S-adenosyl-L-methionine (SAM). Another metabolic precursor, phytol, enhances nematode resistance in Arabidopsis via the ethylene pathway[52], indicating that tocopherol-related metabolites can activate ethylene-dependent defence mechanisms. On the other hand, Surowka et al.[53] identified VTE1 as a modulator of ethylene sensitivity, influencing how plants perceive and respond to stress cues. This finding places VTE1 as a regulator upstream of ethylene perception, likely through the role of tocochromanols in maintaining membrane integrity and redox balance at sites of ethylene receptor localization. In tomato fruits, ethylene treatments increased α-, β-, and γ-tocopherol contents in a ripening stage–dependent manner, although the magnitude of this effect was lower than that observed upon JA treatment[54]. Consistently, inhibition of ethylene perception by 1-methylcyclopropane (1-MCP) reduced tocopherol levels during tomato fruit ripening, supporting a positive role for ethylene signalling in tocopherol accumulation[55].
In Arabidopsis, genetic studies have revealed that ethylene signalling exerts a regulatory role on tocopherol contents. During the first hours of water stress, mutation in the EIN3 gene (a key transcription factor of the ethylene signalling pathway[56]) abolished the α-tocopherol increments observed in Arabidopsis wild-type leaves. Moreover, Arabidopsis mutants for ethylene perception (etr1-1) and production (eto1-1) showed significant increments in α-tocopherol levels during leaf ageing[1]. However, these responses were not observed when mutant plants were subjected to high light intensity (photodamage)[57]. This implies that VTE gene transcription responds to signalling components downstream of CTR1 (likely EIN3/EIN2-mediated pathways) rather than to ethylene gas concentration per se. Furthermore, Müller & Munné-Bosch[3] proposed that tocopherols may interact with Ethylene Response Factors, particularly ERF1, a key regulator at the crossroads of ethylene and JA signalling, thereby integrating antioxidant status with hormonal stress-response networks. This knowledge allows us to hypothesize on a possible role of tocopherols acting as a molecular modulator of transcription factor activity.
Hormonal regulation of growth and development
-
Here, we discuss research papers on potential cross-talks between tocochromanols and those hormones for which evidence is limited in comparison with those discussed above. Particularly, we focus here on auxins, cytokinins, gibberellins (GAs), and brassinosteroids (BRs).
Auxins regulate a wide range of physiological processes throughout plant development, including the establishment of bilateral symmetry during embryogenesis, root initiation, apical dominance, and environmentally driven responses such as gravitropism and phototropism[58]. Notably, Sereflioglu et al.[59] demonstrated in soybean roots that α-tocopherol-dependent salt tolerance is more closely related to auxin synthesis than to enhancement of antioxidant defence, providing physiological evidence that tocopherol effects on growth are partly auxin-mediated. More recently, links between tocopherols and auxins were suggested by experiments presented by Yu et al.[60] in Arabidopsis and soybean. These authors demonstrated that GGDR, a key enzyme in tocopherol biosynthesis, and the connected carotenoid pathways[61], are transcriptionally induced by IAA, GA3, SA, and MeJA and that its expression levels correlate well with α-tocopherol contents. This observation is in line with the fact that in tomato, IAA modulates carotenoid biosynthetic genes[62]. Moreover, GGDR-overexpressing soybean and Arabidopsis plants accumulated more α- and γ-tocopherol than their WT counterpart. Future research should address whether tocopherols affect auxin pathways through direct transcriptional regulation, through post-translational effects on auxin transporters, or via broader impacts on cellular redox and membrane dynamics.
The scarce knowledge on the auxin-tocochromanol relation is also true for the case of cytokinins, with controversial results. Mesa et al.[1] reported reduced resistance of tocopherol-deficient A. thaliana mutants to fungal infection that correlates with the lack of cytokinin increment in these mutants. This reduction in resistance correlated with a failure to accumulate cytokinins in response to infection, suggesting that tocochromanols are required upstream of cytokinin signalling for the activation of antifungal defenses. This is currently the strongest evidence linking tocochromanols to cytokinin pathways. On the other hand, Alharbi et al.[63] reported that α-tocopherol levels did not show significant changes after cytokinins treatments.
In relation to GAs, it has been shown in Cannabis sativa L. that the effect of GA3 on α-tocopherol content is concentration dependent; tocopherol levels decreased and increased at lower and higher GA3 concentration, respectively[64]. These results, together with the fact that GA3 also modulate the DXS (1-deoxy-d-xylulose-5-phosphate synthase) enzymatic activity, a MEP pathway rate-limiting step, confirm that the GA3 effect is at the very upstream precursor level. Another possibility is that GA3 may influence tocopherol levels through multiple and competing mechanisms, possibly involving GGDP flux partitioning between the tocopherol and GAs pathways at higher concentrations. In wheat, GA3 treatment significantly enhanced the production of tocopherols and elevated the α-/γ-tocopherol ratio in a time- and genotype-dependent manner. Genotypic differences were observed in the effects of GA3 on tocopherol yield and composition in the seeds[65], suggesting that allelic variation in VTE genes or in GA responsive regulatory elements could modulates the responsiveness of tocochromanol biosynthesis to GAs signalling.
BRs also have a dose-dependent effect, inducing the accumulation of tocopherols in leaves of Brassica juncea Czern[66] and also in leaves of salt-stressed plants of Vigna sinensis Walp[67]. Similarly, in pea and lupine plants treated with BRs, increments in seed tocopherols (both α and γ) were observed[68]. More recently, Emamverdian et al.[69] showed that bamboo leaves from plants growing under high lead (Pb) concentration and treated with both JA and BR, resulted in significant increments of total-tocopherol. In all these research papers, the unanswered question remains whether BRs are influencing the production of tocochromanols directly, or rather indirectly by increasing the biosynthesis of other stress hormones, which in turn stimulate tocochromanols accumulation.
With the aim to provide readers with a compendium of all research papers discussed above, Table 1 summarizes pieces of evidence on each hormone-tocochromanol possible cross-talk, categorized by plant species and organs. Of note is the fact that only research papers showing evidence about transcriptional control on VTE genes are listed in this table.
Table 1. Summary table on each hormone-tocochromanol cross-talk evidence.
Tocochromanol/hormone changes cis-regualtory motifs in VTE genes Key VTE genes (*) Species Tissue Genetic context (evidence type) Ref. ABA ↑ α-Tocopherol, ↑ γ-Tocopherol VTE1, VTE4, HPPD ↑ Arabidopsis thaliana Leaves ABA treatment (transcriptomic, metabolomic) [22] ↑ ABA in vte1, vte4 mutants VTE1, VTE4 (loss) Arabidopsis thaliana Rosettes/roots VTE-deficient mutants + salt stress (genetic/mutant, biochemical) [32] ↑ α-Tocopherol VTE3 and VTE4 ↑ Medicago truncatula Seeds ABA-insensitive abi5 mutant (genetic/mutant) [34] ABI4 motifs in VTE gene promoters SlVTE4, SlVTE3, SlVTE1, SlHPPD, SlGGPS Solanum lycopersicum Fruits ABI4-binding motifs in VTE gene promoters (transcriptomic, promoter analysis) [28] ↑ β-Tocotrienol, ↑ VTE, ↓ ABA after MsHPPD overexpression MsHPPD Arabidopsis thaliana Seeds MsHPPD OE (genetic, transcriptomic) [30] ↑ α-Tocopherol ABA dependent/independent VTE2, HPPD ↑ Arabidopsis thaliana Leaves Drought/osmotic stress (metabolomic, transcriptomic) [24] ABRE and IIb motifs in VTE
gene promotersOsγTMT (VTE4), OsHPPD, OsMPBQMT1 (VTE3) Oryza sativa Multiple tissues Promoter analysis (promoter/
cis-element analysis)[27] ↑ α-Tocopherol ↑ ABA HPT/VTE2 Cistus creticus Leaves Drought stress (biochemical) [25] Hordeum vulgare; Triticum sp.; ABA-responsive elements in VTE2 promoter VTE2 (HPT) Thinopyrum intermedium; Avena sp.; Brachypodium distachyon Leaves/stems/
inflorescencesPromoter analysis (promoter/cis-element analysis) [47] SA ↓ Tocopherols in vte2
mutant → ↓ SAVTE2 (loss) Arabidopsis thaliana Leaves vte2 mutant + P. syringae (genetic/mutant, biochemical) [37] ↑ SA in VTE-deficient mutants (low phosphate) VTE1, VTE4 (loss) Arabidopsis thaliana Seedlings vte1, vte4 mutants, low-P (genetic/mutant, metabolomic) [48] SA-responsive cis-element in VTE4 promoter GmVTE4 Glycine max Leaves Promoter analysis (promoter/cis-element analysis) [31] ↑ Tocopherols in heat-shock HPPD, VTE1, VTE2, VTE3, VTE4 Arabidopsis thaliana Seedlings Heat shock treatment (biochemical/metabolomic/genetic) [36] ↑ tocopherol (α, γ, δ, total) GGDR Arabidopsis thaliana; Glycine max Hordeum vulgare; Triticum sp.; Seeds and leaves GGDR OE (biochemical, physiological, expression analysis) [61] SA-responsive elements in VTE2 promoter VTE2 (HPT) Thinopyrum intermedium; Avena sp.; Brachypodium distachyon Leaves/stems/
inflorescencesPromoter analysis (promoter/cis-element analysis) [47] JA HAHB4 motifs in VTE gene promoters SlVTE4, SlVTE3, SlVTE1, SlHPPD, SlGGPS Solanum lycopersicum Fruits HAHB4 binding motifs in VTE gene promoters (transcriptomic, promoter analysis) [28] ↑ JA in vte1, vte4 mutants VTE1, VTE4 (loss) Arabidopsis thaliana Rosettes/roots VTE-deficient mutants + salt stress (genetic/mutant, biochemical) [32] ↑ JA in VTE-deficient mutants (low phosphate) VTE1, VTE4 (loss) Arabidopsis thaliana Seedlings vte1, vte2, vte4 mutants, low-P (genetic/mutant, metabolomic) [48] ↓ α-Tocopherol ↑ JA levels VTE1 (loss) Arabidopsis thaliana Leaves vte1 mutant analysis (biochemical, genetic) [5] ↑ Tocopherols in JA-insensitive mutants; ↑ VTE4 VTE4 ↑ Solanum lycopersicum Fruits JA-insensitive tomato mutants (genetic/mutant, transcriptomic) [46] ↑ tocopherol (α, γ, δ, total) GGDR Arabidopsis thaliana;
Glycine maxSeeds and leaves GGDR OE (biochemical, physiological, expression analysis) [61] JA-responsive elements in VTE2 promoter VTE2 (HPT) Hordeum vulgare; Triticum sp.;
Thinopyrum intermedium; Avena sp.;
Brachypodium distachyonLeaves/stems/
inflorescencesPromoter analysis (promoter/cis-element analysis) [47] Ethylene HAHB4 motifs in VTE gene promoters SlVTE4, SlVTE3, SlVTE1, SlHPPD, SlGGPS Solanum lycopersicum Fruits HAHB4 binding motifs in VTE gene promoters (transcriptomic, promoter analysis) [28] ↑ ethylene in vte1, vte4 mutants VTE1, VTE4 (loss) Arabidopsis thaliana Rosettes/roots VTE-deficient mutants + salt stress (genetic/mutant, biochemical) [32] ↑ α-Tocopherol in ein2-1 mutant (ethylene insensitive) VTE5 Arabidopsis thaliana Roots vte5 /ein2-1 mutants + phytol/nematode treatment (biochemical, genetic) [52] Auxin ↑ tocopherol (α, γ, δ, total) GGDR Arabidopsis thaliana;
Glycine maxSeeds and leaves GGDR OE (biochemical, physiological, expression analysis) [61] Auxin-responsive elements in VTE2 promoter VTE2 (HPT) Hordeum vulgare; Triticum sp.;
Thinopyrum intermedium; Avena sp.;
Brachypodium distachyonLeaves/Stems/Inflorescences Promoter analysis (Promoter/cis-element analysis) [47] Cytokinin ↓ Tocopherols in vte1/vte4 mutants → ↓
cytokinin increment during fungal infectionVTE1/VTE4 (loss) Arabidopsis thaliana Leaves vte Arabidopsis mutants + Botrytis (Genetic/mutant, biochemical) [1] Gibberellin ↑ Tocopherols, altered α-/γ-ratio (genotype- dependent) VTE1/VTE4 Brassica napus L; Arabidopsis thaliana Seeds GA3 exogenous treatment (Metabolomic) [65] ↑ tocopherol (α, γ, δ, total) GGDR Arabidopsis thaliana;
Glycine maxSeeds and leaves GGDR OE (biochemical, physiological, expression analysis) [61] GAs-responsive elements in VTE2 promoter VTE2 (HPT) Hordeum vulgare; Triticum sp.;
Thinopyrum intermedium; Avena sp.;
Brachypodium distachyonLeaves/stems/inflorescences Promoter analysis (Promoter/cis-element analysis) [47] * Only research papers showing evidence about transcriptional control on VTE genes are listed in this table. A critical question is whether the hormonal regulation of tocochromanol biosynthesis documented in model systems represents conserved regulatory networks or species-specific adaptations. Systematic comparison of the three main lines of evidence—tocochromanol content changes, VTE gene expression modulation, and hormone-responsive cis-elements—across different plant species suggests partial conservation, strongest for the ABA axis. Particularly, ABRE elements are present in VTE gene promoters in Arabidopsis, rice, tomato, and soybean, and ABA-induced VTE gene expression has been confirmed in Arabidopsis, Medicago, rice, and alfalfa, suggesting a broadly conserved regulatory connection. For JA, JA-responsive elements have been identified in VTE2 in barley, wheat, and oat[50], and the functional interaction between JA signalling and tocopherol accumulation has been shown in both Arabidopsis and tomato. The mechanism may be conserved but is more context-dependent than the ABA axis. For SA, evidence is primarily from Arabidopsis and soybean; conservation in other species is plausible but not yet demonstrated. For growth hormones (auxins, GAs, BRs), the available evidence is largely from single-species studies, and the degree of regulatory conservation remains an open question that merits systematic comparative genomic and functional analysis.
-
Tocochromanols influence phytohormone signalling through several interconnected mechanisms, which are depicted in Fig. 3 and collectively shape plant responses to environmental stress:
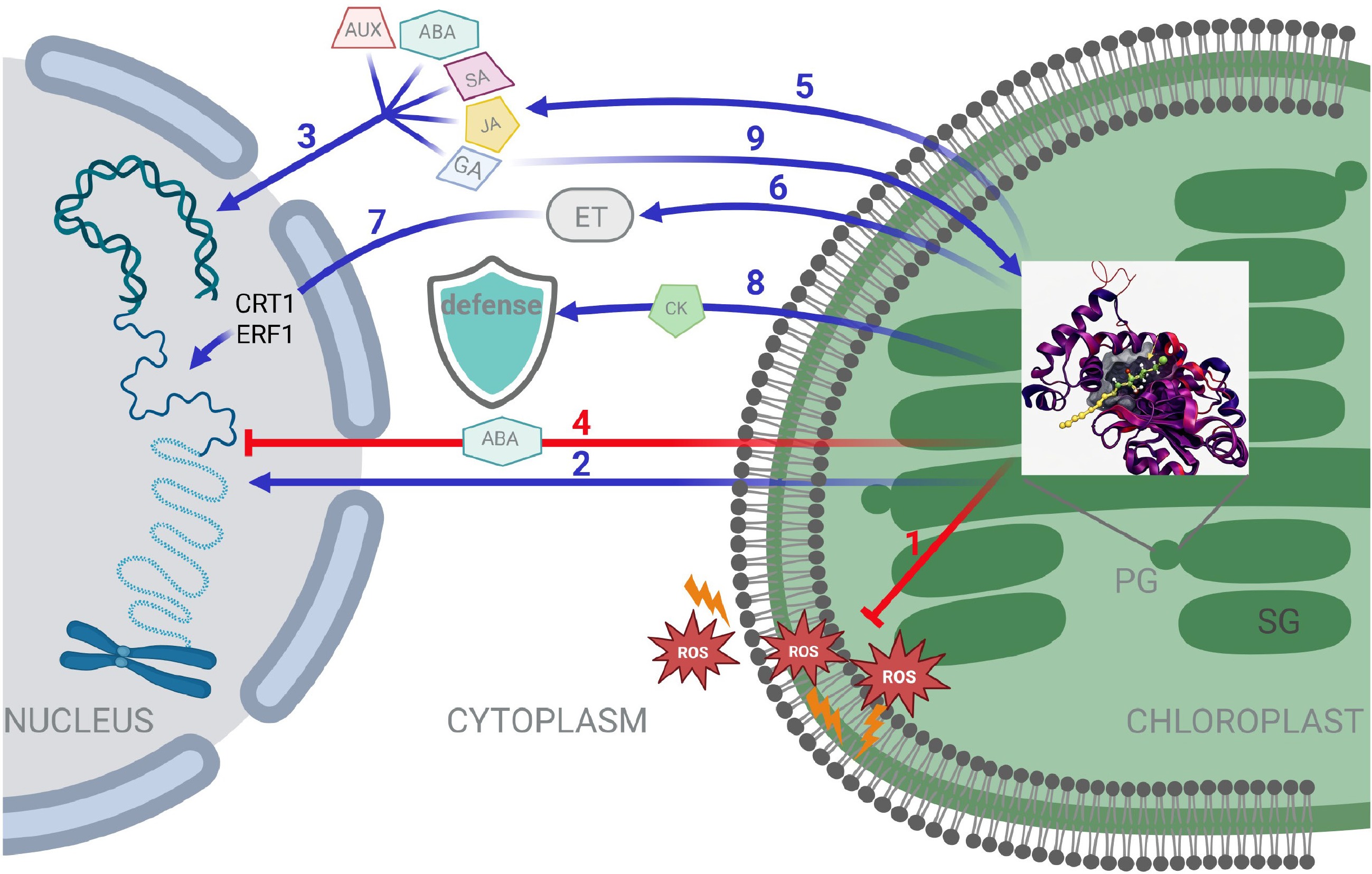
Figure 3.
Tocochromanols influence phytohormone signalling through several interconnected mechanisms. Tocopherol binding protein(s) (TBP) might be one of the carriers for the interactions hypothesized in this scheme. (1) Prevention of ROS accumulation and damage (membranes and cytoplasm). (2) Retrograde signalling pathway. (3) ABA, SA, JA, Aux, and GAs up-regulate the expression of several VTE genes. Aux, GA3, SA, and JA induce GGDR gene expression. (4) Feedback regulation of tocochromanols-ABA. (5) α-tocopherol modulates JA levels. (6) Regulation of ethylene perception by tocopherols. (7) VTE gene transcripts respond to ethylene signalling via CTR1/EIN1. (8) Tococrhomanols act upstream of CKs for activation of antifungal defense. (9) GA3 may influence tocopherol levels through multiple mechanisms. PG: plastoglobuli. SG: stacked grana.
● ROS modulation: ROS function as secondary messengers in numerous hormone pathways, including ABA (H2O2 in stomatal closure), SA (oxidative burst in immunity), and JA. By mitigating oxidative damage and maintaining cellular redox balance, tocopherols indirectly modulate hormone activity through ROS limitation. This balance is essential during stress responses: tocopherols prevent harmful ROS accumulation while preserving the controlled oxidative bursts required to activate protective hormonal signalling.
● Gene expression regulation: Tocopherols participate in plastid-derived retrograde signalling, influencing the expression of nuclear genes responsive to hormones. Through modulation of key transcription factors and signalling cascades, tocopherols help fine-tune hormonal responses to environmental stimuli, ensuring coordinated adjustments to stress.
● Membrane stabilization: As key lipid antioxidants, tocopherols protect cellular membranes from peroxidation, thereby preserving membrane fluidity, integrity, and function. This stabilization is critical for proper hormone transport, receptor activity, and signal transduction, particularly under stress conditions where membrane lipids are highly vulnerable to oxidative damage.
● Hormonal crosstalk in biotic and abiotic stress responses: Tocopherols interact with multiple hormone networks, enabling plants to integrate signals from diverse stressors. For example, tocopherol-mediated modulation of ABA and JA contributes to enhanced drought tolerance and pathogen resistance, while interactions with ethylene and SA help fine-tune immune and defense responses. These synergistic interactions highlight tocopherols as key hubs in stress-signalling networks.
● Regulation of growth-development balance: Under stress, plants must reallocate resources to prioritize survival or overgrowth. Tocopherol-hormone interactions help regulate this balance by influencing hormone homeostasis and signalling outputs. Through their effects on ROS, membrane stability, and transcriptional regulation, tocopherols contribute to optimizing trade-offs between growth and defense.
Among the hormone interactions reviewed here, the ABA–tocochromanol relationship is supported by the most consistent experimental evidence. Direct genetic and biochemical data support a bidirectional feedback loop between ABA levels and VTE gene expression across Arabidopsis, rice, tomato, soybean, and alfalfa. In addition, competition for GGDP represents a plausible metabolic node of crosstalk, although direct flux-partitioning measurements are still lacking. Despite this relatively strong evidence base, several mechanistic aspects remain unresolved. Key open questions include: (i) Are the ABI4-binding sites in SlVTE promoters functionally active? This could be addressed using promoter-deletion GUS reporter assays in tomato. (ii) How does ABA signalling post-translationally regulate VTE enzyme activity? Comparative phosphoproteomic analyses of VTE enzymes in ABA-treated plants vs ABA-signalling mutants would help clarify this regulatory layer.
The SA–tocochromanol relationship is bidirectional but context-dependent. In leaves under pathogen attack, tocochromanols affect SA production by limiting lipid peroxidation and maintaining membrane integrity (biochemical, indirect evidence). At the same time, promoter analyses revealing SA-responsive elements in GmVTE4 suggest the possibility of direct transcriptional regulation. In contrast, in seeds, SA seems to suppress VTE gene expression through an as-yet uncharacterised repressor mechanism (genetic evidence). This apparent tissue-specific duality raises important mechanistic questions. Do SA-responsive elements in VTE4 promoters function as activators in leaves but as repressors in seeds? Stage- and tissue-specific promoter–GUS reporter lines would help resolve this regulatory complexity. Furthermore, identifying the SA-dependent repressor active in seed tissue represents a key step toward understanding how developmental context shapes this interaction.
The JA–tocochromanol relationship also appears to be bidirectional and partly dependent on the tissue. In vegetative tissues under biotic stress, JA may promote VTE gene expression through HAHB4 transcription factor (transcriptomic and promoter evidence)[28]. In contrast, during tomato fruit ripening, JA signalling represses the VTE4 gene, as suggested by genetic evidence from JA-insensitive mutants[47]. This apparent tissue-dependent regulatory switch raises several mechanistic questions. Does the JA-mediated induction of VTE genes in vegetative tissues and their repression in fruit reflect the involvement of distinct cis-regulatory elements, or differences in the co-regulators interacting with transcription factors such as HAHB4? Chromatin immunoprecipitation (ChIP) experiments targeting the HAHB4-binding site at VTE promoters in fruit vs leaf tissues would help address this possibility. In addition, the identity and functionality of the JA-responsive elements reported in oat, barley, wheat, and Brachypodium VTE2/HPT promoters remain to be validated through transactivation assays.
Despite the SAM-mediated shared step in their biosynthesis, ethylene–tocochromanol interaction appears to cross-talk by downstream signalling components (CTR1/EIN2/EIN3) rather than by changes in ethylene levels themselves, according to the genetic evidence from mutant analyses. Tocopherols may influence ERF1-dependent transcription, potentially through membrane-mediated effects on receptor or co-regulator activity, although current support for this model is indirect. In addition, the tocopherol precursor phytol has been shown to activate ethylene-dependent defense responses. These observations raise the possibility that tocopherols may directly interact with components of the ethylene transcriptional machinery. In particular, the hypothesis that tocopherols physically interact with ERF1[3] points to a key target for experimental validation. Molecular docking of α-tocopherol against the ERF1 AP2/ERF DNA-binding domain could provide an initial assessment of structural plausibility. This could be followed by co-immunoprecipitation or thermal shift assays in VTE mutant backgrounds expressing ERF1–GFP fusions. In addition, yeast two-hybrid screening of tocopherol-treated vs untreated libraries could help identify additional ERF-family interactors.
Clarifying the potential regulatory link between auxins and tocochromanols will require targeted functional analyses. Future research should address: (i) whether auxin treatment induces VTE gene expression in a dose-dependent manner, which could be tested by RNA-seq following IAA application in Arabidopsis seedlings; and (ii) whether AuxRE motifs in VTE promoters are functionally active, which could be examined through promoter-deletion GUS reporter assays.
The available evidence suggests that tocochromanols may operate upstream of cytokinin accumulation during defense responses (genetic evidence), although cytokinin regulation of VTE gene expression has not yet been established. The relationship may therefore be indirect, potentially mediated by redox changes that affect cytokinin stability or signalling competence. Discriminating between these possibilities will require targeted experimental approaches. For example, do tocopherol-deficient mutants fail to accumulate cytokinins during infection because of altered ROS homeostasis or changes in membrane dynamics? Measuring cytokinin levels in vte1 mutants treated with antioxidants (e.g., ascorbate) during infection would help distinguish between these scenarios. In addition, direct testing of cytokinin-inducible VTE gene expression by qRT-PCR following cytokinin treatment in Arabidopsis would clarify whether cytokinin signalling feeds back onto the tocochromanol biosynthetic pathway.
Future research should determine whether GA-deficient mutants (e.g., ga1) show altered tocochromanol profiles and whether GA treatment affects HPPD promoter activity in germinating seeds. For the case of BRs, even more basic experiments would be designed to contribute to elucidating the relationship between this hormone action and tocopherol metabolism. A good collection of Arabidopsis BR mutants is publicly available for such a task[70].
-
The evidence discussed above converges on three major conclusions, and highlights several unresolved mechanistic questions regarding the basis of tocochromanol–hormone interactions.
First, the ABA–tocochromanol regulatory axis represents the best-supported bidirectional interaction described in this review. Converging evidence from genetic studies (e.g., Arabidopsis vte1/vte4 and Medicago abi5 mutants), transcriptomic analyses, promoter architecture (including ABRE motifs in VTE promoters across Arabidopsis, rice, tomato, and soybean), and physiological experiments across multiple species collectively support a reciprocal regulatory relationship between ABA signalling and the VTE pathway[14,22,24,27,28,32,34]. Notably, this interaction appears conserved at the level of promoter organization and functional response.
Second, JA and SA show credible but context-dependent interactions with the VTE pathway. In both cases, complementary transcriptional, genetic, and biochemical evidence supports regulatory connections in model species such as Arabidopsis and tomato. However, the conservation of these interactions across plant lineages, as well as their directionality across tissues and developmental stages, remains less clear and requires further investigation[5,31,36−38,41,45−49].
Third, ethylene appears to regulate tocopherol accumulation primarily through downstream signalling components rather than through hormone levels per se, as illustrated by the contrasting phenotypes of the eto1-1 and ctr1-3 mutants[1,56,57]. This observation suggests that transcriptional targets downstream of CTR1 may play a central role in regulating VTE gene expression.
Despite the relative progress in the field that this review may contribute to, several important ambiguities remain. And perhaps most critically, the intracellular transport of tocochromanols remains unresolved. Tocochromanol biosynthesis occurs in plastids, yet many of the proposed signalling interactions involve cytosolic, membrane-associated, or nuclear processes. Without a clear understanding of how tocochromanols—or their derivatives—move from plastids to these sites of action, the mechanistic basis for their role in hormone signalling remains largely inferential. In this context, the discovery of plant proteins with the ability to bind tocochromanols and putatively transport to the different cell compartments and/or from cell to cell adds another dimension to this problem[71]. Up to now, two plant proteins with this feature have been characterized in plants; the tomato tocopherol-binding protein (SlTBP) capable of mediating tocopherol function during inter- and intra-organellar communication[1], and more recently, Hornbergs et al.[72] characterized a second plant protein binding α-tocopherol, PATL2, another member of the same SEC14-like protein family[73]. SlTBP knockdown tomato plants have a massive change in their transcriptomes; classification of the genes exhibiting altered patterns of expression includes a high proportion of transcripts belonging to the hormone metabolism category. Moreover, PATL6, the closest Arabidopsis orthologue, was shown to be part of a co-expression cluster with an enrichment of auxin-related genes[73]. These seminal contributions, together with the identification of homologous tocopherol-binding proteins in other plant species, offer a promising starting point for understanding how tocochromanols may be transported or targeted to hormone-signalling compartments. Further characterization of TBP family members across plant species will likely provide important insights into how tocopherols interact with retrograde signalling pathways to coordinate plant responses to developmental cues and environmental stress. The conceptual model proposed by Muñoz & Munné-Bosch[71], which places tocochromanol metabolism at the interface of stress signalling and crop nutritional improvement, gains additional support from the interactions summarized in this review.
-
During the preparation of this work, the author(s) used ChatGPT (4.0) for Table 1 structure and formatting. The author(s) reviewed and edited all content of the table produced with the assistance of this tool, verified its accuracy, and take full responsibility for the integrity and originality of the final manuscript. This work represents the author(s)' own intellectual contribution, and no AI tool is credited as an author.
-
The authors confirm their contributions to the paper as follows: conceived the topic and structure of the review: Carrari F, Asís R; conducted the literature survey and drafted the manuscript: Santoro LJ, Echeverría M, Perez-Flores LJ, Asís R, Carrari F; prepared the figures: Santoro LJ, Carrari F. All authors discussed the content and approved the final version of the manuscript.
-
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
-
This work was supported by the ANPCYT (Grant No. BID PICT 2017-1907, Argentina), the CONICET (Grant No. 11220200102248, Argentina), the Universidad de Buenos Aires (Grant No. PIDAE 2025 RESCS-2024-1909 1-371, Argentina), and the Universidad Autónoma Metropolitana – Iztapalapa (México). The authors acknowledge Miss Ainhoa De Amico for her help in producing Fig. 3.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Santoro LJ, Echeverría M, Perez-Flores LJ, Asís R, Carrari F. 2026. Tocochromanols - plant hormones crosstalk. Plant Hormones 2: e015 doi: 10.48130/ph-0026-0013 

Tocochromanols - plant hormones crosstalk
- Received: 05 January 2026
- Revised: 31 March 2026
- Accepted: 22 April 2026
- Published online: 22 June 2026
Abstract: Tocochromanols, the group of compounds with vitamin E activity, are lipid-soluble antioxidants synthesized by photosynthetic organisms. Their biosynthesis is restricted to plastids, while their action sites are in different cellular compartments (i.e., cytosol, plasma membrane, and nucleus). Although a key open question concerns how these compounds are transported to the different cell compartments, a considerable piece of evidence suggests their crosstalk with different plant hormones acting in many different physiological mechanisms. In this review, we discuss the current knowledge about these crosstalks.
-
Key words:
- Tocochromanols /
- Vitamin E /
- Plant hormones /
- Crosstalk /
- Intracellular transport /
- Plastids