








-
The global population boom in the 21st century has led to a significant increase in water consumption. Unfortunately, this rise has been accompanied by escalating water pollution[1,2]. Heavy metals are among the most worrisome pollutants affecting both aquatic and terrestrial environments[3]. These pollutants persist in water and sediment, posing a continuous threat due to their toxicity, even in low concentrations[3]. The non-biodegradability and low solubility of heavy metals, including their ionic forms such as cadmium, make them difficult to eliminate, allowing them to accumulate in aquatic ecosystems. These ionic forms are readily absorbed by living organisms, where they can induce oxidative stress, disrupt steroidogenesis, impair spermatogenesis, and trigger apoptotic toxicity, ultimately impairing biological processes[4−6].
As a result, heavy metals can concentrate within organisms, amplifying as they move up the food chain[7]. This contamination not only reduces biodiversity and ecosystem productivity but also undermines the self-repair abilities of affected water bodies[8]. In addition to the ecological damage, the global economic costs of heavy metal pollution in aquatic systems exceed USD $10 billion annually[9]. Ensuring cleaner water through the removal of these pollutants could preserve biodiversity and yield significant economic benefits[10].
Traditional methods for removing heavy metals from water bodies, such as physicochemical or electrical treatments, are often costly and inefficient[11,12]. Bioremediation offers a promising alternative, utilizing biological agents to transform or degrade pollutants into less harmful substances[3,13]. Fungi, in particular, are effective bioremediation agents due to their high tolerance and absorption capacity for heavy metals, outperforming bacteria and chemical methods in bioleaching studies[3,14].
Researchers have found that fungal isolates sourced from polluted environments, such as industrially impacted rivers, tend to exhibit superior bioremediation capabilities compared to lab-cultured fungi[15,16]. This suggests that environmental pressures may enhance the effectiveness of fungi in mitigating heavy metal contamination. Therefore, exploring diverse natural water sources for potential fungal isolates is crucial[17].
Fungi have shown significant potential as bioremediation agents for environments contaminated with heavy metals due to their remarkable tolerance to toxic substances. They can thrive in extreme conditions, with some species tolerating heavy metal concentrations as high as 400 ppm[18]. Studies have demonstrated that fungi like Aspergillus terreus and Trichoderma viride are capable of effectively removing heavy metals from contaminated media, with A. terreus removing 59.67 mg/g of lead and T. viride removing 16.25 mg/g[18]. Their ability to tolerate and accumulate heavy metals is attributed to their unique structural features, including a rigid cell wall made of chitin, which enhances metal uptake[19]. Fungi also exhibit a high biomass yield and are relatively inexpensive to cultivate, making them an attractive alternative to bacteria for bioremediation. Unlike bacteria, which exhibit a significant decline in activity under high metal concentrations, fungi can maintain or even increase their activity, especially for metals like arsenic and cadmium[20,21]. This increased tolerance is linked to their metabolic differences and high osmotic pressure in their cell walls[19]. Fungi also provide the advantage of a modifiable genetic makeup, which can be optimized for large-scale production and enhanced metal biosorption[22]. Their ability to secrete various metabolites further enhances their potential for treating industrial effluents, making them a promising candidate for the bioremediation of heavy metals[23].
Rivers, particularly those exposed to industrial and urban waste, are prime sampling sites for identifying fungi capable of bioremediation. The Iloilo River in Iloilo City, Philippines, is an example of such a site, as it is frequently subjected to heavy metal pollution due to its location in a densely populated and industrialized area[24]. Water and sediment samples from this river offer valuable opportunities to study the bioremediation potential of local fungal species in combating heavy metal pollution.
This study aims to employ ITS rDNA barcoding for the accurate identification of fungal isolates collected from the Iloilo Ferry Terminal, Iloilo, Philippines, and to conduct a preliminary assessment of their tolerance to cadmium exposure. Through the analysis of these isolates, this research seeks to enhance the understanding of fungal roles in bioremediation within aquatic ecosystems impacted by heavy metal contamination.
-
Water and sediment samples were collected at the river mouth of the Iloilo Ferry Terminal Port, Iloilo City, Philippines (Fig. 1). A total of 500 mL of water samples and 500 g of sediment were collected for analysis. Sediment samples were collected using a sterile corer and stored in sterile zip-lock bags to prevent contamination. Water samples were collected in sterile amber bottles to minimize light exposure, thereby reducing the potential degradation of photosensitive compounds and preserving the integrity of any microorganisms present. Both sediment and water samples were transported to the University of the Philippines Visayas - Microbiology Laboratory, Miagao, Iloilo, Philippines. Water samples were maintained at 4 °C during transport using a cooler to minimize any physicochemical changes that could affect the analysis.
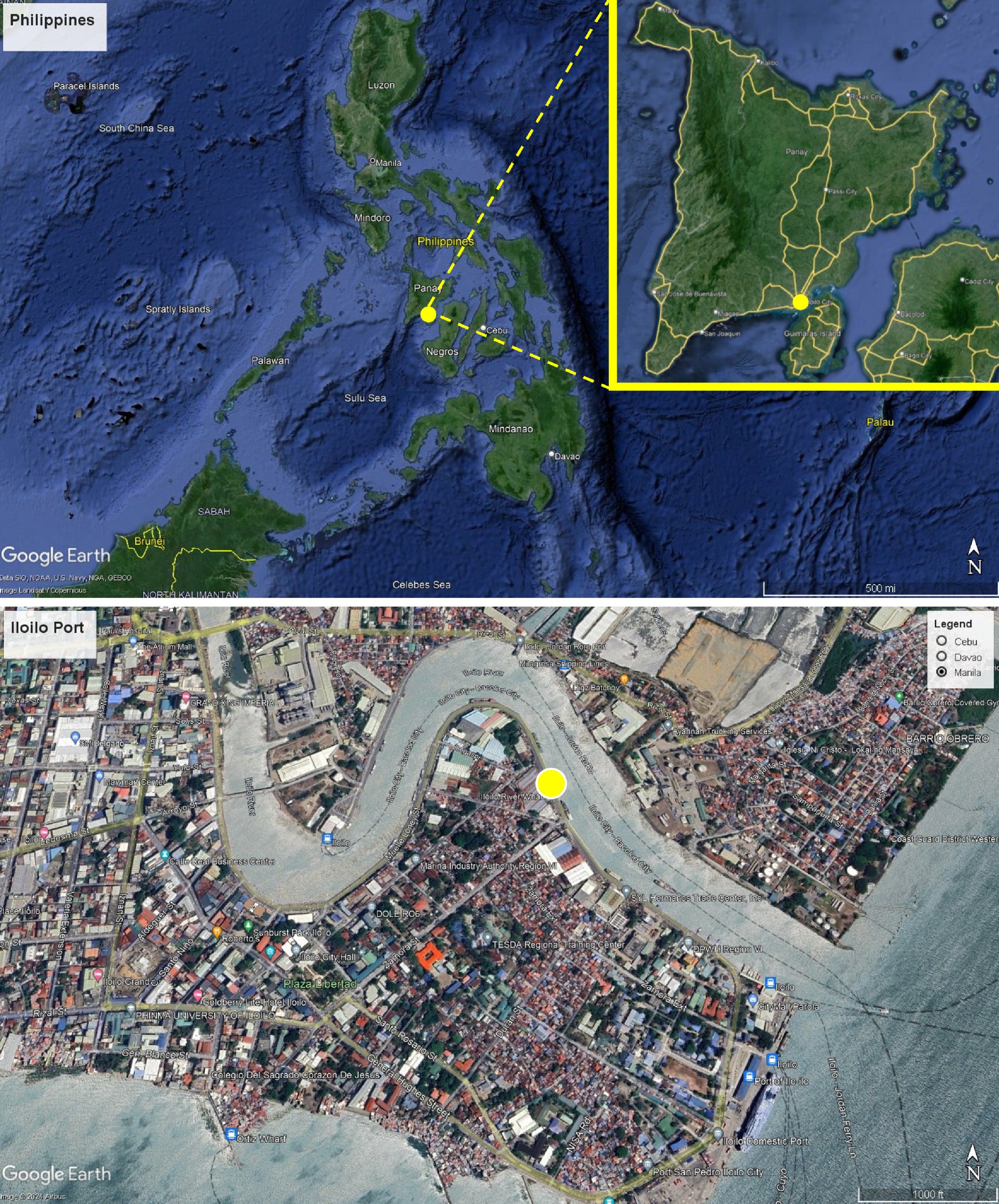
Figure 1.
Map showing the sampling sites (yellow circle) in Iloilo-Guimaras Ferry Terminal Port, Iloilo City, Iloilo, Philippines. The map was created using Google Earth Pro.
Preparation of heavy metal stock solution and media preparation
-
Three different concentrations (25, 50, and 100 ppm) were prepared for stock solutions of cadmium [Cd(II)]. Cadmium nitrate tetrahydrate was used as the cadmium source. A 1,000 ppm stock solution of cadmium was prepared by dissolving 2.774 g of cadmium nitrate tetrahydrate in 1 L of distilled water. The prepared 1000 ppm stock solution was used to create the desired working concentrations (25, 50, and 100 ppm) using the dilution formula: C1V1 = C2V2 (where C1 and V1 are initial concentration and volume before dilution, respectively; C2 and V2 are final concentration and volume after dilution, respectively). The diluted solutions were sterilized using 0.22 μm filters with a vacuum filtration setup consisting of a vacuum filter flask and a vacuum pump.
Potato dextrose agar (PDA), supplemented with Pen-Strep, was prepared following the manufacturer's instructions. To achieve a final concentration of 25 ppm cadmium in the medium, a 4 mL cadmium (25 ppm) solution was added per 1 L of cooled, molten PDA. This concentration was used for the initial isolation of fungi tolerant to cadmium.
Fungal isolation and morphological identification
-
Water and sediment samples collected from the port were serially diluted up to 10−6. Aliquots of 0.1 mL from each dilution were then spread onto potato dextrose agar (PDA) supplemented with Cd(II) at concentrations of 25 ppm using a spread plate technique. Each dilution was plated in triplicate to minimize random error. Petri plates were incubated for a week at 28 ± 2 °C and observed daily for fungal growth. Unique fungal colonies were purified and characterized using both colonial characteristics and micromorphological features. Colonial traits include color, texture, elevation, margin, and characteristics of aerial hyphae. For microscopic examination, the slide culture technique was employed to observe traits such as spore morphology, shape, and conidial formation, which were analyzed based on the cultural characteristics of media supplemented with 20 ppm Cd(II)[25].
Primary screening of fungal isolates for cadmium tolerance
-
All fungal isolates were assessed for their tolerance to cadmium [Cd(II)] by culturing them on PDA media supplemented with Cd(II) at higher concentrations (i.e., 50 and 100 ppm). For comparison, fungal isolates grown on PDA medium without Cd(II) served as controls. All cultures were incubated for 5 d at a temperature of 28 ± 2 °C. The growth of each fungal isolate was visually assessed and recorded based on relative growth compared to the control and categorized as follows: ++++ (normal growth), +++ (slightly less than normal), ++(less than normal), + (least growth) and – (absent growth)[15].
DNA extraction, PCR amplification, and sequencing
-
Fungal mycelia from pure cultures grown in PDA for 15 d were scraped using a sterilized scalpel and kept in a sterilized 2 mL microcentrifuge tube. Genomic DNA was extracted using the DNeasy Blood & Tissue Kit (Qiagen) following the manufacturer's protocol. Polymerase chain reaction (PCR) was used to amplify the internal transcribed spacers (ITS) of rDNA using the primers ITS4 and ITS5[26]. Polymerase chain reaction was performed in a volume of 50 μl, which contained 5 μL 10 XPCR buffer, 1 μL of forward primer 10 μM, 1μL reverse primer 10 μM, 1 μL of dNTP, 0.25 μL of Hotstart DNA polymerase, 36.75 μL nuclease-free water, and 5 μL of genomic DNA (5 ng/μL). The PCR thermal cycle program for ITS amplification was as follows: an initial denaturing step of 95 °C for 15 min, followed by 35 cycles of denaturation at 94 °C for 1 min, annealing at 60°C for 30 s, elongation at 72 °C for 1 min, and final extension at 72 °C for 10 min. Agarose gel electrophoresis was carried out to confirm the presence of amplicons at the expected molecular weight. PCR products were purified and sequenced with the primers mentioned above at the Philippine Genome Center – Visayas, University of the Philippines Visayas, Miagao, Iloilo, Philippines.
Phylogenetic analyses
-
Taxonomic classifications were established based on the closest matches obtained from nucleotide BLASTn searches. The ITS sequence data was analyzed in conjunction with additional sequences retrieved from GenBank. Phylogenetic analysis was conducted using maximum likelihood (ML) and Bayesian inference (BI) methods. ML analyses were carried out with IQ-TREE webserver (
www.hiv.lanl.gov/content/sequence/IQTREE/iqtree.html ), while BI analyses were performed with MrBayes v.3.2.6 on XSEDE[27], accessed through the CIPRES web portal (www.phylo.org/portal2 )[28], following the procedures of Dissanayake et al.[29]. The resulting phylogenetic trees were visualized with FigTree v. 1.4.4, and the final layout was created in Microsoft PowerPoint 365. Newly generated sequences have been deposited in GenBank. -
Of the colonies exhibiting tolerance to 25 ppm cadmium supplementation, 15 displayed distinct morphological characteristics. Among these, nine isolates were successfully sequenced and became the primary focus of this study. Table 1 provides detailed information on their taxonomic identities, cadmium tolerance levels, and the original sample sources from which they were isolated. Six isolates (Pseudopestalotiopsis elaeidis, Peroneutypa scoparia, Diaporthe sp., Phlebia sp., Pyrrhoderma noxium, and Trichosporon asahii) from sediment exhibited tolerance to both 50 and 100 ppm cadmium concentrations, indicating their resilience to heavy metal stress. In contrast, water isolates displayed mixed results, with only Trichosporon asahii showing tolerance to both cadmium concentrations. The remaining water isolates (Cerrena sp., Irpex laceratus, and Schizophyllum commune) were not tolerant to 50 and 100 ppm cadmium. This suggests that sediment-derived fungi generally demonstrate a higher tolerance to cadmium contamination compared to those from water sources.
Table 1. Tolerance of isolates to 50 ppm and 100 ppm cadmium supplementation with sample sources.
Taxon Source Growth* 25 ppm 50 ppm 100 ppm Pseudopestalotiopsis elaeidis Sediment ++++ ++++ ++++ Peroneutypa scoparia Sediment ++++ ++++ ++++ Cerrena sp. Water +++ – – Diaporthe sp. Sediment ++++ ++++ ++++ Phlebia sp. Sediment ++++ ++++ ++++ Irpex laceratus Water +++ – – Schizophyllum commune Water +++ – – Pyrrhoderma noxium Sediment ++++ ++++ ++++ Trichosporon asahii Water ++++ ++++ ++++ * ++++ (normal growth); +++ (slightly less than normal); ++ (less than normal); + (least growth); – (absent growth). Phylogenetic analysis
-
The ITS gene dataset comprised 75 taxa from Ascomycota, with Parasympodiella elongata (CBS 522.93) and P. eucalypti (CBS 124767) as the outgroup taxa (Fig. 2). The analyzed dataset, after trimming, comprised a total of 531 characters. Alignment has 75 sequences with 531 columns, 321 distinct patterns, 268 parsimony-informative, 31 singleton sites, and 232 constant sites. The ML analysis for the combined dataset provided the best scoring tree (Fig. 2) with a final ML optimization likelihood value of -5305.142 (ln). Parameters for the GTR model of the combined ITS dataset are as follows: estimated base frequencies; A = 0.250, C = 0.250, G = 0.250, T = 0.250; substitution rates AC = 1.00000, AG = 1.60667, AT = 1.00000, CG = 1.00000, CT = 2.99928, GT = 1.00000; gamma distribution shape parameter α = 0.346. Support values for maximum likelihood (ML) above 75% are given at the nodes.
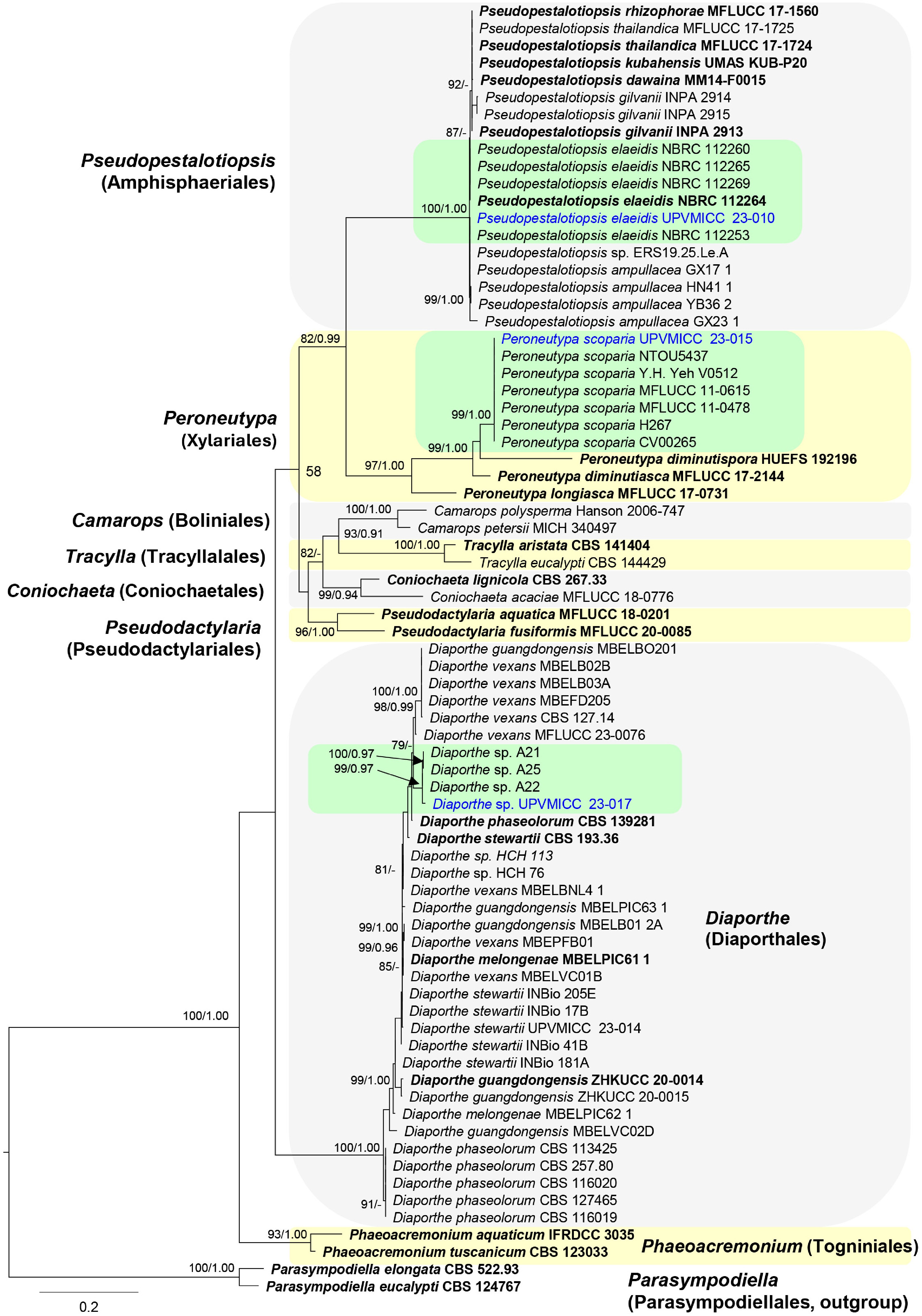
Figure 2.
Phylogram generated from maximum likelihood analysis based on ITS sequence data of Ascomycota taxa. Isolates from this study are highlighted in blue font. Ex-type strains are in bold.
For Basidiomycota, the ITS gene dataset comprised of 242 taxa, with Ustilago abaconensis (CBS 8380) and U. maydis (CBS 380.32) as the outgroup taxa (Fig. 3). The analyzed dataset, after trimming, comprised a total of 1,725 characters. Alignment has 242 sequences with 1,725 columns, 928 distinct patterns, 693 parsimony-informative, 192 singleton sites, and 840 constant sites. The ML analysis for the combined dataset provided the best scoring tree (Fig. 3) with a final ML optimization likelihood value of –22,547.577 (ln). Parameters for the GTR model of the combined ITS dataset are as follows: estimated base frequencies; A = 0.263, C = 0.206, G = 0.227, T = 0.305; substitution rates AC = 1.58477, AG = 2.87131, AT = 1.58477, CG = 1.00000, CT = 4.26806, GT = 1.00000; gamma distribution shape parameter α = 0.611. Support values for maximum likelihood (ML) above 75% are given at the nodes.

Figure 3.
Phylogram generated from maximum likelihood analysis based on ITS sequence data of Basidiomycota taxa. Isolates from this study are highlighted in blue font. Ex-type strains are in bold.
Taxonomy
-
Ascomycota Caval.-Sm., Biol. Rev. 73: 247 (1998)
Sordariomycetes O.E. Erikss. & Winka, Myconet 1(1): 10 (1997)
Amphisphaeriales D. Hawksw. & O.E. Erikss., Syst. Ascom. 5(1): 177 (1986)A
Pestalotiopsidaceae Maharachch. & K.D. Hyde, Fungal Diversity 73: 106 (2015)
Pseudopestalotiopsis Maharachch., K.D. Hyde & Crous, Stud. Mycol. 79: 180 (2014)
Pseudopestalotiopsis elaeidis (C. Booth & J.S. Robertson) F. Liu, L. Cai & Crous, Stud. Mycol. 92: 374 (2018) [2019], Fig. 4a–c
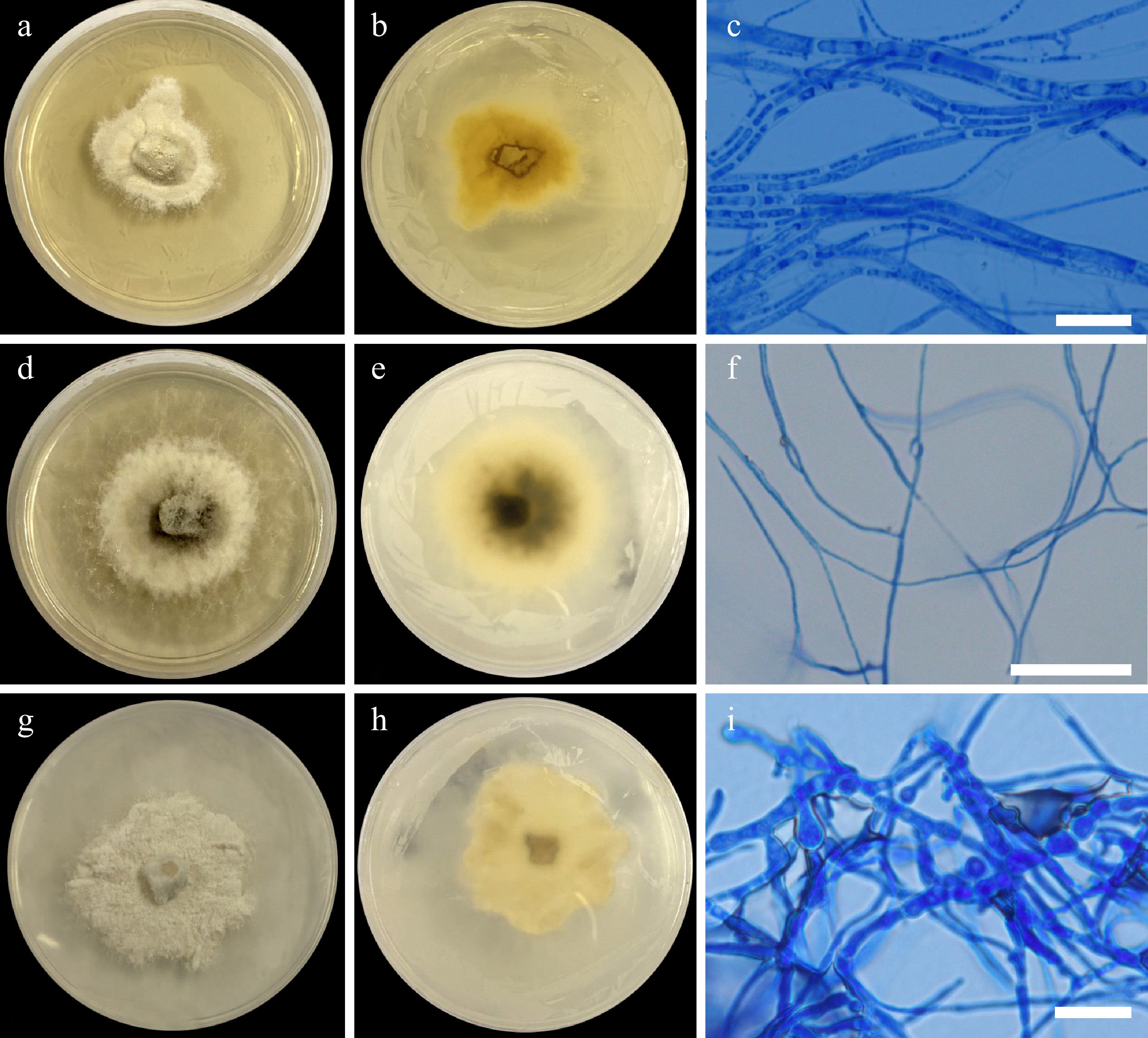
Figure 4.
Colonial morphology on 25 ppm Cd (II). Pseudopestalotiopsis elaeidis UPVMICC 23-010. Colony on PDA from (a) surface, and (b) reverse. (c) Septate hyphae. Peroneutypa scoparia UPVMICC 23-015. Colony on PDA from (d) surface, and (e) reverse. (f) Septate hyphae. Diaporthe sp. UPVMICC 23-017. Colony on PDA from (g) surface, and (h) reverse. (i) Septate hyphae. Scale bars: (c), (f), (i) = 25 μm.
Description: Colonies on PDA reaching 20–25 mm diameter after 7 d at 25 °C, colonies circular, margin entire, flat, velvety to sparse cottony appearance, colony from surface: initially white, became greyish white; reverse: verrucose, yellow. No sporulation even on prolonged incubation (6 months). Hyphae 1.69–3.63 μm (
$\bar {\rm x} $ Material examined: PHILIPPINES, Iloilo, Iloilo City, Iloilo Ferry Terminal Port, on sediment samples, 31 October 2023, F.M. Dela Cruz, SHD-633, culture UPVMICC 23-010.
GenBank number: PQ304372 (ITS)
Notes: Pseudopestalotiopsis elaeidis was introduced by Liu et al.[30] based on their phylogenetic analysis and comparison of nucleotide differences of the ex-type strains of Leptosphaeria elaeidis (CBS 413.62) and P. myanmarina (NBRC 112264). Based on nomenclatural priority, P. myanmarina was synonymized as P. elaeidis. The taxon was reported in China, Indonesia, Myanmar, and Nigeria on various hosts[30−32]. In the phylogenetic analysis, UPVMICC 23-010 clade with other strains of P. elaeidis (NBRC 112260; NBRC 112264; NBRC 112265; NBRC 112269) with high bootstrap support (100% ML, 1.00 BYPP) (Fig. 2). This is the first country report of P. elaeidis in the Philippine aquatic habitats. The isolate was able to grow in all 25, 50, and 100 ppm cadmium and thus can potentially be applied for cadmium bioremediation.
No other studies were found that report heavy metal tolerance or bioremediation applications for this species, making this the first study to document its tolerance to heavy metals.
Xylariales Nannf., Nova Acta R. Soc. Scient. upsal., Ser. 4 8(no. 2): 66 (1932)
Diatrypaceae Nitschke [as 'Diatrypeae'], Verh. naturh. Ver. preuss. Rheinl. 26: 73 (1869)
Peroneutypa Berl., Icon. fung. (Abellini) 3(3-4): 80 (1902)
Peroneutypa scoparia (Schwein.) Carmarán & A.I. Romero, Fungal Diversity 23: 84 (2006), Fig. 4d–f
Description: Colonies on PDA reaching 27–29 mm diameter after 7 d at 25 °C, colonies circular, margin entire, flat, cottony, effuse, fluffy but are flat and dense, colony from surface: denser mycelia at the center, initially white, with grey pigmentations appearing at the center when mature; reverse: white with grey at center. No sporulation even on prolonged incubation (6 months). Hyphae 1.34–4.27 μm (
$\bar {\rm x} $ Material examined: PHILIPPINES, Iloilo, Iloilo City, Iloilo Ferry Terminal Port, on sediment samples, 31 October 2023, F.M. Dela Cruz, SHD-432, culture UPVMICC 23-015.
Notes: Carmarán et al.[33] resurrected the genus Peroneutypa based on morphological and phylogenetic analyses of Diatrypaceae, transferring Sphaeria scoparia to Peroneutypa. Peroneutypa scoparia is a known pathogen causing diseases on various plant hosts[34−40]. Additionally, the taxon has been reported as saprobic[33,41], and endophytic[42] in nature. The isolated P. scoparia UPVMICC 23-015 represents the first record of this species in the Philippine aquatic habitats. Phylogenetic analysis revealed that it clustered with other documented P. scoparia species with strong bootstrap support (99% ML, 1.00 BYPP) (Fig. 2). Growth in 25, 50, and 100 ppm cadmium was observed, which suggests potential application for cadmium bioremediation.
There is limited documentation on the use of P. scoparia as a bioremediation agent, with most studies focusing on its enzyme-based remediation of dyes, particularly using laccase. While no applications for heavy metal remediation have been documented, there is considerable potential given that the enzymes employed for dye remediation have demonstrated effectiveness against other toxic substances, including heavy metals[43,44].
Diaporthales Nannf., Nova Acta R. Soc. Scient. upsal., Ser. 4 8 (no. 2): 53 (1932)
Diaporthaceae Höhn. ex Wehm., Am. J. Bot. 13: 638 (1926)
Diaporthe Nitschke, Pyrenomyc. Germ. 2: 240 (1870)
Description: Colony on PDA has a diameter ranging from 25–27 mm after 7 d at 25 °C incubation, dense and velvety, white on the surface view, similar to its reverse side. Chlamydospores 4.97–6.58 μm (
$\bar {\rm x} $ $\bar {\rm x} $ Material examined: PHILIPPINES, Iloilo, Iloilo City, Iloilo Ferry Terminal Port, on sediment samples, 31 October 2023, F.M. Dela Cruz, SHD-431, culture UPVMICC 23-017.
Distribution: Diaporthe is a genus with a wide geographical distribution, encompassing pathogens, endophytes, and saprobes that infect diverse hosts[45,46]. It has been associated with economically important crops like citrus and coffee, as well as ornamental plants such as Clematis[47−49]. Phylogenetic analyses placed this taxon within the Diaporthe sojae species complex[47] and clustered with other unidentified Diaporthe spp. (99% ML, 0.97 BYPP) (Fig. 2). Growth in 25, 50, and 100 ppm cadmium was also observed, which suggests potential application for cadmium bioremediation.
Basidiomycota R.T. Moore, Bot. Mar. 23(6): 371 (1980)
Agaricomycetes Doweld, Prosyllabus Tracheophytorum, Tentamen Systematis Plantarum Vascularium (Tracheophyta) (Moscow): LXXVIII (2001)
Polyporales Gäum., Vergl. Morph. Pilze (Jena): 503 (1926)
Cerrenaceae Miettinen, Justo & Hibbett, Fungal Biology 121(9): 817 (2017)
Cerrena Gray, Nat. Arr. Brit. Pl. (London) 1: 649 (1821)
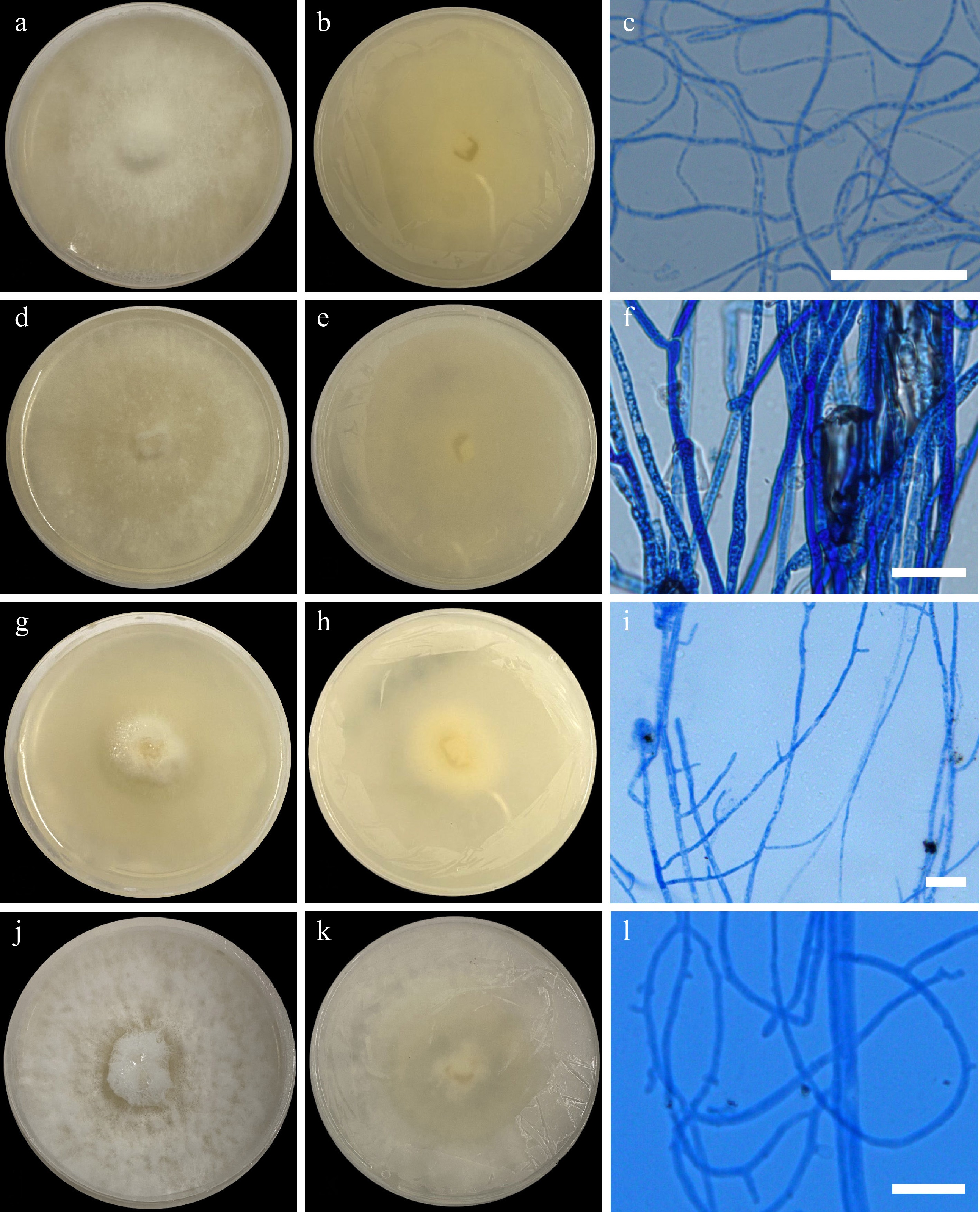
Figure 5.
Colonial morphology on 25 ppm Cd (II). Cerrena sp. UPVMICC 23-008. Colony on PDA from (a) surface, and (b) reverse. (c) Hyphae. Irpex laceratus UPVMICC 23-007. Colony on PDA from (d) surface, and (e) reverse. (f) Hyphae. Phlebia sp. UPVMICC 23-009. Colony on PDA from (g) surface, and (h) reverse. (i) Hyphae. Schizophyllum commune UPVMICC 23-012. Colony on PDA from (g) surface, and (h) reverse. (i) Hyphae. Scale bars: (c), (f), (i), (l) = 25 μm.
Description: Colony on PDA reaching a diameter of 35 to 36 mm after 7 d at 25 °C, white (surface and obverse), cottony, and round; Mycelia fluffy, short, dense; septate hyphae 3.15–4.76 μm (
$\bar {\rm x} $ Material examined: PHILIPPINES, Iloilo, Iloilo City, Iloilo Ferry Terminal Port, on water samples, 31 October 2023, F.M. Dela Cruz, WHD-431, culture UPVMICC 23-008.
Notes: Cerrena species are widely distributed across Europe, Africa, and South America[50−52]. Known for its potent wood-decaying ability, this fungus commonly parasitizes living trees before adopting a saprobic lifestyle on dead wood[53]. The isolate clustered with other unidentified Cerrena strains [e.g., NTOU5117; NTOU 4204; 7-SU-3-B-77(M)-B; 6-L-3-C-32(M)][54−56] with strong bootstrap support (100% ML, 1.00 BYPP) (Fig. 3). However, the presence of Cordyceps militans (CH9) within this cluster raises the possibility of misidentification and must be correctly identified as Cerrena sp. In the phylogenetic analysis, given its distinct lineage compared to established Cerrena taxa (i.e., C. albocinnamomea, C. cystidiata, C. gilbertsonii, C. multipileata, C. unicolor, and C. zonata), this strain might represent a novel species and warrants further investigation through sequencing of its protein-coding regions. In aquatic environments, the genus Cerrena was isolated from decaying mangrove wood[57]. This is the first report of Cerrena isolated from brackish water. Growth was only observed on 25 ppm cadmium and not on higher concentrations (50 and 100 ppm).
Cerrena is a white rot fungus that possesses the same enzyme as Peroneutypa scoparia, namely the laccase enzyme, and has been reported to play a role in environmental contaminant biodegradation[58]. This enzyme has been associated with the tolerance of fungal species to heavy metals, as it exhibits broad-spectrum activity against various contaminants, including heavy metals[59]. Furthermore, this enzyme is known for its ability to decolorize and detoxify industrial effluents, aiding in wastewater treatment[60]. It has been observed to degrade a multitude of contaminants, including heavy metals[61]. Interestingly, the promoter regions of the laccase genes harbor distinct recognition sites tailored for xenobiotics and heavy metals, making heavy metals significant activators of this enzyme[62]. When present in the medium, xenobiotics and heavy metals can attach to the gene's recognition sites, thereby triggering the production of laccase[60]. For example, when Cerrena sp. is exposed to Cu2+ and Zn2+, increased laccase production is observed[63]. Cadmium has also been observed to specifically increase laccase activity and is considered an inducer of the enzyme[64−66].
Irpicaceae Spirin & Zmitr., in Spirin, Mycena 3: 48 (2003)
Irpex Fr., Syst. orb. veg. (Lundae): 81 (1825)
Irpex laceratus (N. Maek., Suhara & R. Kondo) C.C. Chen & Sheng H. Wu, Fungal Diversity 111: 411 (2021), Fig. 5d–f
Description: Colony on PDA has a diameter that ranges 45–48 mm after 7 d of incubation at 25 °C, white (surface and obverse), velvety, evenly flat, and translucent with some areas having moderate fluffiness while others seem to be waxy. Hyphae 2.94–4.23 μm (
$\bar {\rm x} $ Material examined: PHILIPPINES, Iloilo, Iloilo City, Iloilo Ferry Terminal Port, on water samples, 31 October 2023, F.M. Dela Cruz, WHD-622, culture UPVMICC 23-007.
Distribution: Irpex laceratus is typically found on angiosperms and exhibits a vast distribution across Southeast and East Asia[67−69]. Additionally, this fungus contributes to white rot in wood and has a global presence, with its primary concentration in the boreal region of the Northern Hemisphere[70]. The new isolate clustered with other I. laceratus strains (BRPET15, ATCC42010, Y.H. Yeh l1215, J5-2, hlb5_4_1) (Fig. 3) with strong statistical support (98% ML, 1.00 BYPP). Interestingly, it also grouped with I. latemarginatus with moderate support (93% ML, 1.00 BYPP). While traditionally known as a terrestrial fungus, I. laceratus has been isolated from aquatic environments in Acanthaster planci[71] and Ostrea denselamellosa[72], demonstrating bioactivity. It is considered a marine fungus by Jones et al.[73] and Calabon et al.[74]. This is the first report of I. laceratus isolated from brackish water in the Philippines. Growth was observed on 25 ppm cadmium, while no growth was observed on both 50 and 100 ppm.
The only mention of I. laceratus in relation to bioremediation pertains to its wood-degrading capabilities, with no references to its effectiveness in heavy metal remediation[75].
Meruliaceae Rea, Brit. basidiomyc. (Cambridge): 620 (1922)
Phlebia Fr., Syst. mycol. (Lundae) 1: 426 (1821)
Description: The colony on PDA reaches a diameter of 48–49 mm after 7 d at 25 °C, white, dense, and cottony center on the obverse side, as the colony grows outwards, the cottony texture becomes more flat and yellow coloration of the agar occurs. Hyphae 2.94– 4.23 μm (
$\bar {\rm x} $ Material examined: PHILIPPINES, Iloilo, Iloilo City, Iloilo Ferry Terminal Port, on sediment samples, 31 October 2023, F.M. Dela Cruz, SHD-632, culture UPVMICC 23-009.
Notes: Species of the genus Phlebia are widely distributed across North America and Europe, with additional records from New Zealand[76]. The Phlebia sp. isolated in this study clustered with uncultured fungal clones (L042881-122-061-B10, L042881-122-061-B11, L042881-122-061-B12, and L042886-122-066-F07), two unidentified Phlebia species (MG60, MGEF50), and a fungal endophyte, GFM3, with robust statistical support (100% ML, 0.97 BYPP) (Fig. 3). Notably, this clade also contained Aspergillus versicolor, a clear case of misidentification. This taxon was included in the phylogenetic tree based on BLASTn results. Phylogenetic analysis revealed that Phlebia sp. UPVMICC 23-009 forms a distinct lineage separate from other known Phlebia species (P. floridensis and P. radiata), suggesting it may represent a novel taxon. Further investigation through sequencing of its protein-coding regions is warranted. While Phlebia species have been reported in aquatic environments, such as mangrove stands in Japan, with biodegradation capabilities[77], this study represents the first record of the genus in brackish waters within the Philippines. There was observed growth in all three concentrations of cadmium 25, 50, and 100 ppm and has the potential to be used for the bioremediation of cadmium.
Phlebia species utilized as bioremediation agents have shown promising results, particularly in the remediation of heavy metals[78]. For instance, it has been observed that Phlebia species remove up to 97.1% of cadmium, 97.5% of lead, and 72.7% of nickel[79]. However, the same study also illustrated changes in the morphology of the fungus when exposed to heavy metals[79]. Similar to Cerrena sp., Phlebia sp. is another example of white rot fungi[80], which are generally renowned for their biodegradability and have recently gained attention for their remarkable results in heavy metal-specific bioremediation[81,82]. Phlebia species is also capable of producing the previously mentioned enzyme laccase[79]. Similarly, laccase production in Phlebia sp. is intensified by the presence of metals, acting as inducers. It is believed that the main mechanism for Phlebia sp. tolerance and potential bioremediation application involves intracellular compartmentalization of metals inside vacuoles and biosorption[79]. Known Phlebia species with capabilities for heavy metal bioremediation and tolerance include P. brevispora, P. floridensis, and P. radiata[79,81].
Hymenochaetales Oberw., Beitr. Biol. Pfl.: 89 (1977)
Hymenochaetaceae Donk, Bull. bot. Gdns Buitenz. 17(4): 474 (1948)
Pyrrhoderma Imazeki, Trans. Mycol. Soc. Japan 7: 4 (1966)
Pyrrhoderma noxium (Corner) L.W. Zhou & Y.C. Dai, Mycologia 110(5): 882 (2018), Fig. 6
Description: The colony on PDA has a diameter that ranges 38–40 mm after 7 d of incubation at 25 °C, light greyish brown at the surface view, round, irregular, with a dense and cottony center; as the colony grows outwards, the cottony texture disappears, and the colony becomes flat and white which later develops brown pigmentation as time passes. Hyphal system 1.88–2.96 μ (
$\bar {\rm x} $ $\bar {\rm x} $ Material examined: PHILIPPINES, Iloilo, Iloilo City, Iloilo Ferry Terminal Port, on sediment samples, 31 October 2023, F.M. Dela Cruz, WHD-621, culture UPVMICC 23-011; ibid, on water samples, 31 October 2023, F.M. Dela Cruz, SHD-411, culture UPVMICC 23-016.
Notes: Pyrrhoderma noxium was formerly classified as Phellinidium noxium but was transferred to the genus Pyrrhoderma based on morphological and phylogenetic analyses by Zhou et al.[83]. As a significant pathogen, P. noxium infects a broad host range of over 200 plant species, with a particularly devastating impact on more than 100 tropical tree species, including economically and culturally important plants like breadfruit and mango[84,85]. This pathogen poses a substantial threat to Pacific Island ecosystems due to its widespread distribution across pan-tropical regions, encompassing Asia, Australia, Africa, and Oceania[85−87]. Phylogenetic analysis based on the ITS gene region placed the isolated UPVMICC 23-016 within a clade of P. noxium strains (Fig. 3). In aquatic environments, P. noxium has been isolated from marine sediment in the South China Sea[88]. The isolated Pyrrhoderma noxium UPVMICC 23-016 was the first record of its species in the brackish waters of the Philippines. Growth was observed on 25, 50, and 100 ppm cadmium.
No previous literature has reported on the bioremediation potential or applications of P. noxium. This study is the first to identify and demonstrate its tolerance to cadmium, highlighting its potential as a novel species for bioremediation.
Agaricales Underw., Moulds, mildews, and mushrooms. A guide to the systematic study of the Fungi and Mycetozoa and their literature (New York): 97 (1899)
Schizophyllaceae Quél., Fl. mycol. France (Paris): 365 (1888)
Schizophyllum Fr. [as 'Schizophyllus'], Observ. mycol. (Havniae) 1: 103 (1815)
Schizophyllum commune Fr. [as 'Schizophyllus communis'], Observ. mycol. (Havniae) 1: 103 (1815), Fig, 5j–l
Description: Colony on PDA reaches 48–49 mm in diameter after 7 d at 25 °C, white and woolly cottony colony in surface view; obverse, white. Mycelia density is compact. Hyphae 2.94–4.23 μm (
$\bar {\rm x} $ Material examined: PHILIPPINES, Iloilo, Iloilo City, Iloilo Ferry Terminal Port, on sediment samples, 31 October 2023, F.M. Dela Cruz, SHD-622, culture UPVMICC 23-012.
Notes: Schizophyllum commune is widely distributed across temperate and tropical regions[89]. It typically exists as a weak parasite or saprophyte on various woody plants, although it can occasionally be found on herbaceous species[89]. This fungus has been isolated from diverse terrestrial habitats and exhibits an unexpected versatility in aquatic environments. In marine ecosystems, S. commune has been recovered from subseafloor and deep-sea sediments[90−93], as well as associated with marine macroalgae, Flabellia petiolata[94] and Padina pavonica[95], and seagrass Posidonia oceanica[96]. In the phylogenetic analysis, the isolate UPVMICC 23-012 clustered with other strains of S. commune (CBS 124811; CBS 227.57; CBS 342.58; IHEM:25479; IHEM:27140; KoRLI047393; T28)[97]. Notably, the isolation of Schizophyllum commune UPVMICC 23-012 represents the first record of this species in the brackish waters of the Philippines. The isolate was only able to grow on 25 ppm cadmium with no growth in 50 and 100 ppm.
This fungal species has been previously proven to withstand high heavy metal concentrations and effectively bioremediate them[81,98]. The species' tolerance can be attributed to the up-regulation of glutathione S-transferase[99,100]. Another mechanism for tolerance observed in this species is their ability to bind heavy metals in their cell walls. The protein β-glucans found in the walls of S. commune can bind heavy metals such as Cu2+[101]. It has also been revealed that binding, rather than uptake, of heavy metals, is perhaps the main mechanism for tolerance. Binding to the cell wall can serve as a storage mechanism, limiting the availability of these molecules to competitors, or preventing toxic influx into the cytoplasm[102]. Additionally, S. commune is another fungal species capable of producing the enzyme laccase, with overexpression of this enzyme observed alongside the accumulation of heavy metals such as Zn, Pb, and Cd inside its organelles. Overexpression of this enzyme has also been associated with higher heavy metal tolerance[103].
The effectiveness of S. commune as biosorbents is attributed to their rapid kinetics, impressive biosorption capacity, and ability to selectively remove metal ions from electroplating industrial effluents[104]. It has also been demonstrated that S. commune can transport heavy metals along their hyphae[105].
Tremellomycetes Doweld, Prosyllabus Tracheophytorum, Tentamen Systematis Plantarum Vascularium (Tracheophyta) (Moscow): LXXVIII (2001)
Trichosporonales Boekhout & Fell, Int. J. Syst. Evol. Microbiol. 50: 1363 (2000)
Trichosporonaceae Nann. [as 'Trichosporaceae'], Repert. mic. uomo: 285 (1934)
Trichosporon Behrend, Berliner Klin. Wochenschr. 21: 464 (1890)
Trichosporon asahii Akagi ex Sugita, A. Nishikawa & Shinoda, J. gen. appl. Microbiol., Tokyo 40(5): 405 (1994), Fig. 7
Description: Colony on PDA has a diameter that ranges from 29–31 mm after 7 d at 25 °C, white, creamy to dry, powdery on the surface and obverse view, the center is farinose and agar fissures on its reverse side. Blastoconidia 2.58–5.04 μm (
$\bar {\rm x} $ Material examined: PHILIPPINES, Iloilo, Iloilo City, Iloilo Ferry Terminal Port, on sediment samples, 31 October 2023, F.M. Dela Cruz, SHD-022, culture UPVMICC 23-013.
Notes: Trichosporon, a fungus resembling yeast, is widely distributed in nature and found in soil, water, plants, animals, and even within the natural flora of the human body[106]. Although typically harmless, it can cause illness in humans under certain circumstances. Research spanning several decades has documented instances of Trichosporon asahii infection, totaling 140 cases between 1996 and 2019, with reports from various regions worldwide. Asia accounted for most of these cases, comprising 77.1% of the total[107]. The isolate's ITS gene region gave 93% ML support as its clade with several T. asahii strains. Together with the other type strains, they grouped with strains of Trichosporon aquatile, T. japonicum and T. asteroides with 100 % ML and 1.00 BYPP support (Fig. 3). In aquatic environments, T. asahi was isolated from sand[108], and sediments[109−113]. The isolation of T. asahii UPVMICC 23-013 represents the first record of this species in the brackish waters of the Philippines. The isolate has potential for use in cadmium bioremediation, as it grew in all tested concentrations of cadmium (25, 50, and 100 ppm).
This species has been observed to tolerate not only cadmium but also other heavy metals, including Pb, Cu, and As. The resistant strains exhibited multiple heavy metal binding sites in their cell walls[114,115]. However, it has also been observed that the involvement of the cell wall in the heavy metal stress response caused phenotypic changes, particularly in the cell wall structure[116]. Additionally, the glutathione (GSH)-glutathione disulfide (GSSG) system, the most prevalent redox system in fungi and yeasts, was downregulated in T. asahi when exposed to heavy metals, except for cadmium, where it was increased[111,114]. GSH is thought to be a tolerance mechanism of the yeast against high levels of oxidative stress caused by metal exposure. Yeasts are also observed to have efficient heavy metal accumulation methods[114]. This suggests that cadmium has a specific and positive impact in the tolerance and bioremediation capability of T. asahii.
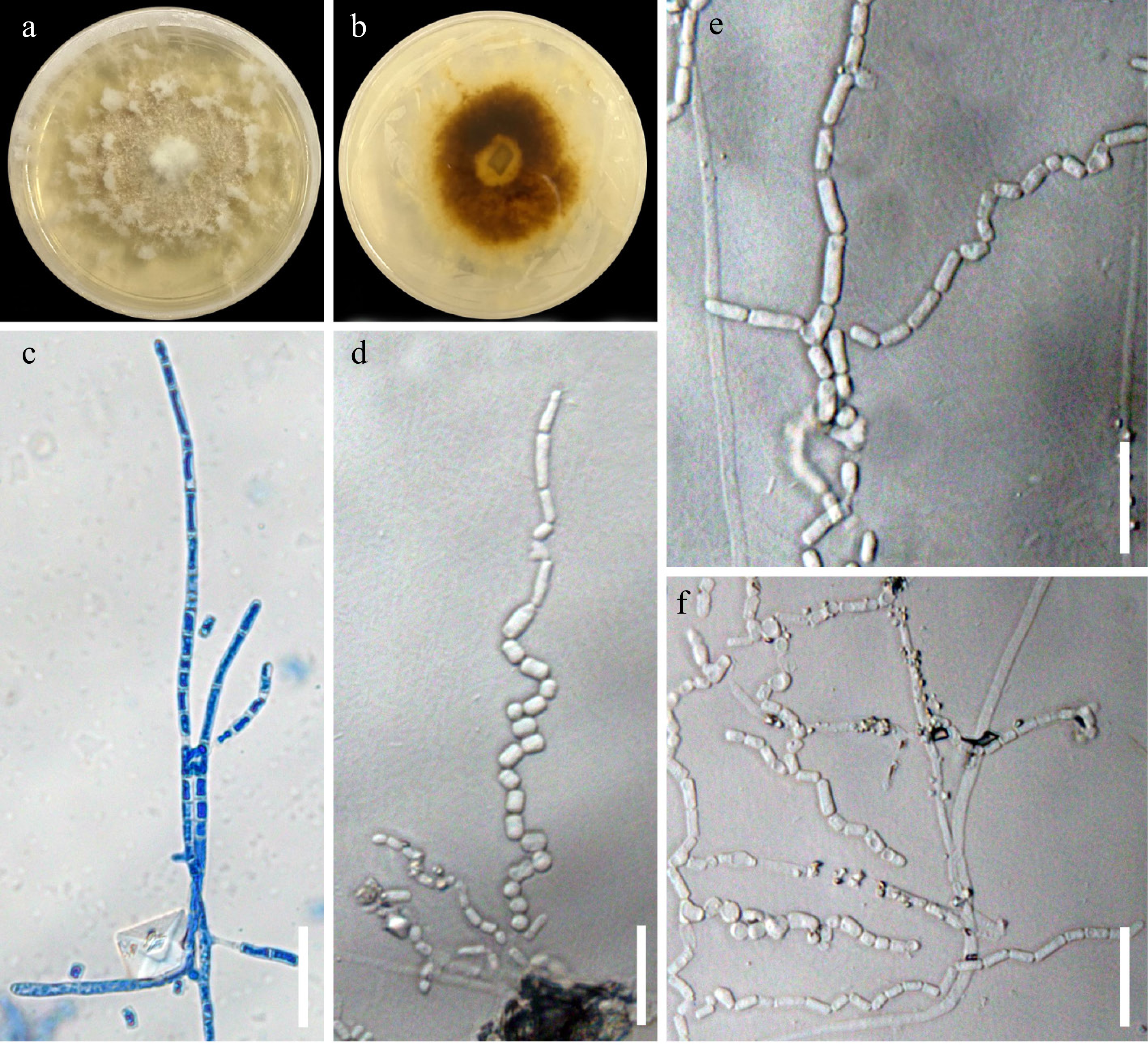
Figure 6.
Pyrrohoderma noxium UPVMICC 23-016. Colony on PDA with 25 ppm Cd(II) from (a) surface, and (b) reverse. (c) Septate hyphae beginning to fragment. (d)–(f) Arthrospores. Scale bars: (c)–(f) 25 μm.
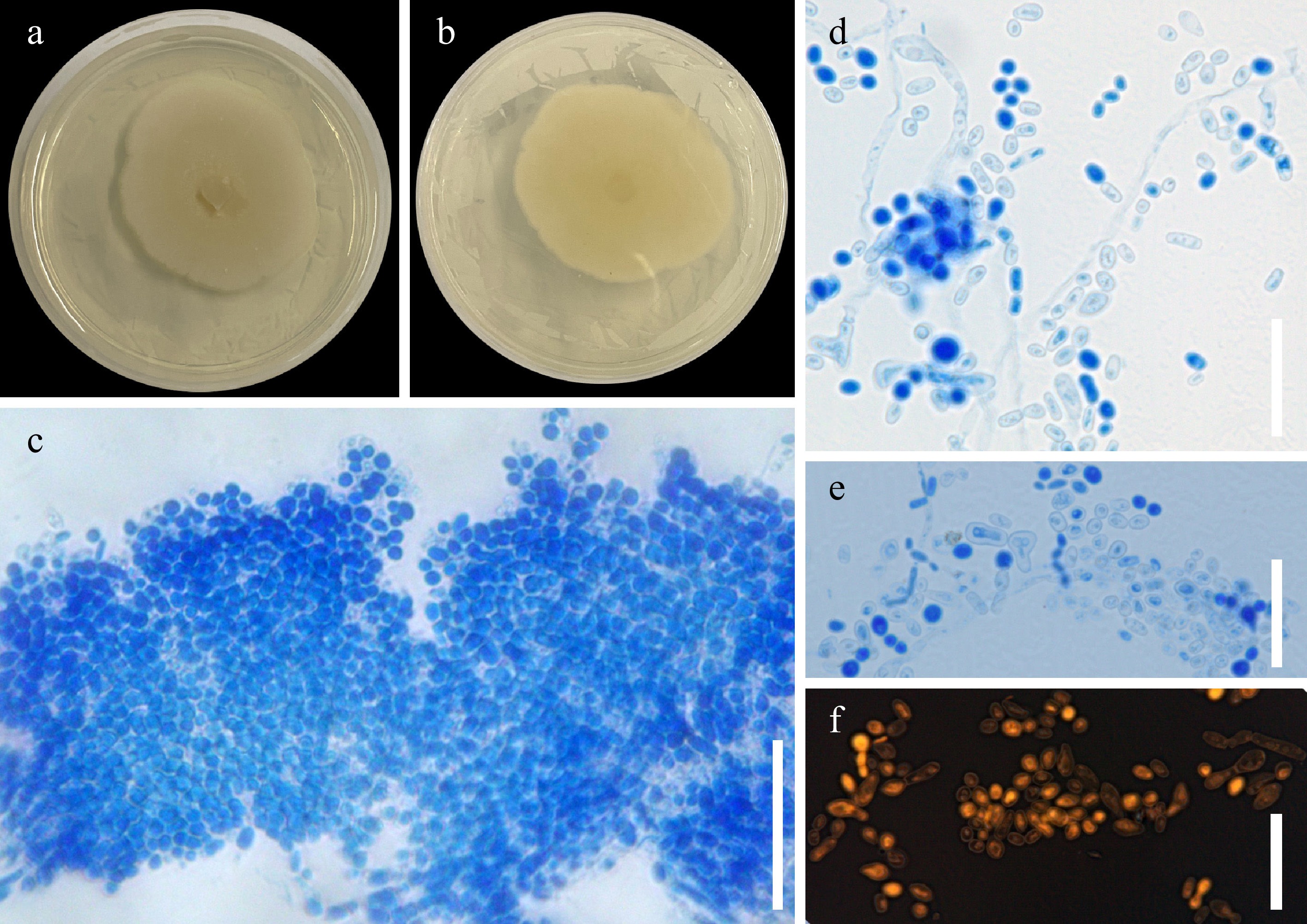
Figure 7.
Trichosporon asahii UPVMICC 23-013. Colony on PDA with 25 ppm Cd(II) from (a) surface, and (b) reverse. (c)–(f) Blastoconidia. Scale bars: (c)–(f) = 25 μm.
-
The Iloilo River, particularly the Iloilo-Guimaras Ferry Port, serves as a significant sample site, revealing the presence of heavy metal-tolerant fungi. This observation suggests potential heavy metal contamination in the area, as such fungi are typically found in polluted environments[14,117−122]. The level of heavy metal tolerance displayed by these fungi reflects the conditions of their isolation site. Fungi isolated from metal-contaminated areas tend to exhibit higher resistance compared to those from uncontaminated sites, indicating that heavy metals exert selective pressure favoring resistant microorganisms[123]. Studies on the Iloilo River have consistently shown that heavy metal concentrations exceed the guidelines set by the Philippine Department of Environment and Natural Resources (DENR)[124−127]. The contamination is likely driven by activities at the ferry port, including leachates from reclamation areas, port operations, untreated industrial effluents, and domestic sewage[128−130].
Sediment samples yielded more fungal isolates than water samples, highlighting the critical role of sediments as a source for identifying diverse fungal species. This can be attributed to sediment's vital function in river ecosystems, where it acts both as a sink and source of heavy metals[131,132]. As a dynamic component of the river basin, sediment shapes diverse habitats and environments[133]. Heavy metals tend to accumulate in sediments at significantly higher concentrations than in the water column, particularly in river mouths, which are often more polluted than other zones[134−137]. The proximity of the ferry port further supports the sampling choice, as port infrastructure and operations are linked to elevated heavy metal concentrations[138]. Sediments not only collect heavy metals from river systems but can also emit them back into the environment. Elevated metal levels in sediments often reflect the geochemical composition of the parent material[139]. Heavy metals in sediments and water commonly originate from effluents released by metal processing, recreational, municipal, and agricultural industries. These pollutants undergo chemical and microbial transformations, impacting environmental quality and public health[140,141]. Furthermore, sediments provide a more stable and nutrient-rich environment for fungal growth compared to water. The stability of the substrate supports the development of vegetative hyphae in filamentous fungi while offering refuge from environmental fluctuations, contributing to the higher fungal diversity observed in sediments. In contrast, aquatic environments are more dynamic and less conducive to fungal stability, leading to lower isolation rates[142−146].
Phylogenetic analysis of the fungal isolates identified two phyla: Ascomycota and Basidiomycota. Basidiomycota accounted for 66.7% (six isolates), and Ascomycota for 33.3% (three isolates). All isolates demonstrated growth on PDA plates supplemented with 25 ppm of cadmium during the initial screening. Within the Basidiomycota phylum, all isolates belonged to the subphylum Agaricomycotina, while all Ascomycota isolates were classified under Pezizomycotina. These isolates were further categorized into three classes: Agaricomycetes (five isolates; 55.6%), Sordariomycetes (three isolates; 33.3%), and Tremellomycetes (one isolate; 11.1%). The subclasses represented were Xylariomycetidae (two isolates; 22.2%), Incertae sedis (four isolates; 44.4%), Agaricomycetidae (two isolates; 22.2%), and Diaporthomycetidae (one isolate; 11.1%). The orders identified included Polyporales (three isolates; 33.3%), and Trichosporonales, Agaricales, Hymenochaetales, Xylariales, Amphisphaeriales, and Diaporthales, each with one isolate (11.1%). At the family level, Diaporthaceae, Trichosporonaceae, Schizophyllaceae, Hymenochaetaceae, Meruliaceae, Irpicaceae, Cerrenaceae, Diatrypaceae, and Pestalotiopsidaceae were each represented by one isolate (11.1%).
These findings align with a metagenomic study[147,148], investigating fungal communities in heavy metal-contaminated sites, which also identified Ascomycota and Basidiomycota as dominant phyla. Similar to this study, Sordariomycetes was the most prominent class, supporting the assertion that it includes heavy metal-tolerant fungi. Within Basidiomycota, Agaricomycetes was the most abundant class, followed by Exobasidiomycetes and Tremellomycetes, consistent with the results of this study, where five of six Basidiomycota isolates belonged to Agaricomycetes, and one was classified as Tremellomycetes. Metagenomic analysis further revealed that isolates from these phyla possessed a higher number of genes associated with metabolic pathways, gene information processing, biosorption processes, and DNA repair, which were absent in isolates from non-contaminated sites. These genetic adaptations likely underpin the heavy metal tolerance of these fungal communities. Greater fungal diversity was also observed in less-contaminated sites, as reported in related studies[148].
Ascomycota and Basidiomycota exhibit remarkable potential for heavy metal bioremediation and are often the dominant fungal groups in contaminated sites[80,148−150]. The heavy metal tolerance of Basidiomycota members is attributed to their ability to produce a variety of degradative enzymes, such as peroxidases, H2O2-generating oxidases, and laccases, commonly found in white rot fungi species[81,82,151−155]. These enzymes enhance survival and metabolic activities while supporting other fungal species in the consortium[80,145]. Similarly, members of the Ascomycota phylum demonstrate heavy metal tolerance through biosorption and metabolic capabilities, alongside their robust survival across diverse habitats, allowing them to adapt rapidly to environmental changes[80−82,147−157].
-
This study elucidates the considerable fungal diversity within the Iloilo River, revealing nine taxa with tolerance to cadmium exposure. The identification of taxa from both Ascomycota and Basidiomycota emphasizes the ecological resilience of fungi in response to anthropogenic pollution. These findings not only augment our understanding of fungal biodiversity within this aquatic environment but also provide a foundational basis for subsequent investigations into the bioremediation potential of these isolates. By harnessing the inherent metabolic capabilities of these fungi, we can develop innovative and sustainable strategies for mitigating heavy metal contamination. This approach holds promise for the restoration and protection of aquatic ecosystems, ultimately enhancing public health outcomes by reducing environmental toxicity.
The authors would like to thank the Department of Agriculture (DA) - Bureau of Fisheries and Aquatic Resources (BFAR) for the issuance of the gratuitous permit. MS Calabon is grateful to the UP System Balik PhD Program (OVPAA-BPhD-2022-02) entitled, 'Unraveling the hidden diversity of aquatic fungi from Panay Island, Philippines'.
-
The authors confirm contribution to the paper as follows: study conception and design: Dela Cruz FM, Calabon MS, Bermeo-Capunong MRA; data collection: Dela Cruz FM, Bagacay JFE; analysis and interpretation of results: Dela Cruz FM, Bagacay JFE, Calabon MS, Canto CM; draft manuscript preparation: Dela Cruz FM, Calabon MS. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated during the study is available within the article.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Dela Cruz FM, Bermeo-Capunong MRA, Bagacay JFE, Canto CM, Calabon MS. 2025. Taxonomy, phylogeny, and preliminary screening of fungal isolates for cadmium tolerance from Iloilo Ferry Terminal, Iloilo, Philippines. Studies in Fungi 10: e001 doi: 10.48130/sif-0025-0001 

Taxonomy, phylogeny, and preliminary screening of fungal isolates for cadmium tolerance from Iloilo Ferry Terminal, Iloilo, Philippines
- Received: 21 October 2024
- Revised: 24 December 2024
- Accepted: 25 December 2024
- Published online: 23 January 2025
Abstract: Water pollution by heavy metals poses a growing threat to ecosystems and human health. Bioremediation using fungi offers a promising solution due to their tolerance and absorption capabilities. This study aimed to isolate and identify cadmium-tolerant fungi from the Iloilo River, a potentially polluted river in Iloilo City, Philippines using morpho-phylogenetic analysis. Water and sediment samples were collected, and fungi were isolated by serial dilution on PDA plates supplemented with cadmium ions [Cd(II)] (25 ppm). Isolates were further tested for tolerance to varying cadmium concentrations (50 and 100 ppm). Nine fungal species were identified, including three ascomycetous taxa (Pseudopestalotiopsis elaeidis, Peroneutypa scoparia, Diaporthe sp.) and six basidiomycetous taxa (Cerrena sp., Irpex laceratus, Phlebia sp., Pyrrhoderma noxium, Schizophyllum commune, Trichosporon asahii). The majority of the isolates originated from sediment samples. Morphological illustrations, descriptions, and taxonomic notes are provided for each taxon. These findings provide insight into the fungal diversity of the Iloilo River and their tolerance to cadmium.
-
Key words:
- Aquatic fungi /
- Bioremediation /
- Fungal biodiversity /
- Fungal systematics /
- Heavy metals