









-
Flavonoids are a crucial group of polyphenolic compounds and are among the most abundant secondary metabolites in plants[1]. These compounds are widely distributed across various plant organs, including roots, stems, leaves, flowers, fruits, and seeds. Among them, anthocyanins, a subclass of flavonoids, are water-soluble pigments responsible for imparting vibrant colors such as red, purple, blue, and violet to plant tissues. Anthocyanins primarily include delphinidin, cyanidin, pelargonidin, peonidin, malvidin, and petunidin. Among these, delphinidin is considered the key pigment responsible for the formation of blue flowers and is often absent in many plant species[2]. Studies have shown that these anthocyanin pigments play a crucial role not only in attracting pollinators and facilitating seed dispersal but also in aiding plants to resist various biotic and abiotic stresses[3,4].
Nymphaea, commonly known as water lilies, form a genus within the family Nymphaeaceae, consisting of perennial aquatic plants that are widely distributed across tropical, subtropical, and temperate regions[5,6]. Renowned for their striking diversity in floral coloration and unique morphological characteristics, these plants have emerged as a major focus of investigation within both horticultural and broader scientific research communities globally[7]. As a basal angiosperm[8], Nymphaea holds a crucial position in the evolutionary history of plants, which makes it a focus of considerable scientific interest. In the field of flower color research, alongside traditional hybrid breeding techniques, extensive studies have been conducted to analyze the anthocyanin compositions in cultivars with various flower colors[9]. These studies have revealed that cyanidin and its derivatives are the primary pigments in the blue-violet petals of certain cultivars[10]. In N. colorata, the blue-violet petals, as well as naturally occurring red and white flower variants, offer valuable material for flower color research. In 2020, Zhang et al. conducted transcriptome analysis and identified multiple genes potentially involved in pigmentation, including the ANS gene, suggesting that the ANS gene plays a crucial role in the coloration of blue-violet petals[8].
Anthocyanin synthase (ANS) belongs to 2-ketoglutarate-dependent dioxygenase, also known as leucoanthocyanidin dioxygenase (LDOX). Its primary role is to catalyze the transformation of colorless anthocyanins into their colored anthocyanin[11]. The product of this catalytic reaction is the first chromogenic compound in the anthocyanin biosynthesis pathway, playing a pivotal role in determining the coloration of various plant organs[12]. The ANS gene was first isolated from a corn (Zea mays) mutant through transposon tagging[13], and later cloned into various plant species, including Arabidopsis thaliana[14], litchi (Litchi chinensis)[15], mango (Mangifera indica)[16], apple (Malus pumila)[17], tea tree (Camellia sinensis)[18], and grape (Vitis vinifera)[19]. This research led to the identification of ANS as a key regulator of anthocyanin accumulation and color deposition. Furthermore, the ANS genes in pomegranate (Punica granatum)[20] and grape (Vitis vinifera)[21] also play crucial roles in anthocyanin biosynthesis. Specifically, in V. vinifera[21], the expression level of the ANS gene directly determines the accumulation of pigments in the fruit, influencing its color. Certain members of the ANS gene family contribute differently to the synthesis of various types of anthocyanins under specific environmental conditions. In jujube (Ziziphus jujuba), the expression of the ANS gene is regulated by an atypical MYB circadian rhythm protein, exhibiting fluctuations between day and night[22]. Abscisic acid treatment has been shown to upregulate ANS expression in blueberries, leading to increased anthocyanin content[23]. Additionally, studies have demonstrated that low temperatures enhance the expression of NaANS in red-skinned bananas. UV-A and UV-B radiation can induce anthocyanin accumulation in tea plants (Camellia sinensis) by upregulating the ANS gene expression[24].
In the foundational research on Nymphaea, we previously completed the genome sequencing of N. colorata and obtained a high-quality reference genome sequence. This work also led to the identification of candidate genes for blue petals. However, research on the ANS gene family in N. colorata remains limited. In this study, we identified members of the ANS gene family from the N. colorata genome and conducted a systematic analysis of their physicochemical properties, gene structures, and expression patterns. These findings provide a theoretical foundation for further exploration of the functional roles of the ANS genes in N. colorata and their potential applications in molecular breeding.
-
In this study, N. colorata was used as the research material. Petals of N. colorata and its variants, including blue petals, red petals, and white petals, were collected, with three biological replicates taken from each cultivar. The petals were immediately frozen in liquid nitrogen and stored at −80 °C until needed for future analysis. The samples were subsequently used for transcriptome sequencing.
Identification of ANS genes in N. colorata
-
The protein sequences of the ANS genes of Arabidopsis thaliana were used as queries and downloaded from the TAIR database (
www.arabidopsis.org , accessed on 2 November 2024). Using the assembled N. colorata genome, we performed a homology search to align N. colorata sequences with those of Arabidopsis. The results were then integrated with those identified using the 2OG-FE_OXY protein domain HMM model.Examination of the physicochemical properties of the N. colorata ANS genes
-
The physicochemical properties, like molecular weight (MW), isoelectric point (pI), hydrophilicity, and instability index of the N. colorata ANS genes were calculated and analyzed using the TBtools software[25].
Evolutionary tree, conserved motif, and domain analysis
-
Using MUSCLE software[26], we performed multiple sequence alignment and constructed a phylogenetic tree for ANS proteins from Arabidopsis thaliana, Nymphaea colorata, Amborella trichopoda[27] Ginkgo biloba[28], Cycas panzhihuaensis, and Adiantum capillus. A phylogenetic tree was constructed using PhyloForge software[29] and the tree was subsequently modified on online iTOL software for visualization[30]. Motif prediction was conducted using MEME software (
http://meme-suite.org/tools/meme ), and the results were visualized with TBtools, which integrated the MEME output[31], conserved protein domains were identified using the Pfam database[32]. A multi-species evolutionary tree of the ANS proteins was constructed based on these analyses.Examination of promoter cis-acting elements and tissue expression patterns
-
We retrieved the upstream 2,000 bp promoter sequence of the NcANS gene, starting from the transcription start site, using SeqKit software[33]. The cis-acting elements within the promoter sequence were analyzed using the PlantCARE online tool (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )[34]. The results were then visualized using TBtools for graphical representation. The cis-acting element analysis results were classified based on their functions, and the number of occurrences of each cis-acting element ID was quantified. Additionally, the distribution of these elements in the NcANS gene was assessed, and a heatmap showing the distribution of the cis-acting element IDs was generated using DataColor software[35].Expression of ANS genes in different flower colors of N. colorata
-
Total RNA was extracted using a commercial RNA extraction kit and sent to Benna Biological Company for second-generation transcriptome sequencing. After sequencing[36], the data were processed using Fastp software for quality control to remove low-quality reads[37]. Then the filtered reads were aligned to the N colorata genome, and gene expression levels (TMM) were calculated using FeatureCounts[38]. Transcriptome data from different tissues of N colorata were downloaded and subjected to quality control and expression quantification. The expression levels of the 32 NcANS genes were calculated. To investigate the expression profile of the NcANS genes, a heatmap of gene expression was generated using DataColor[35].
Gene localization and synteny analysis of NcANS gene
-
Gene collinearity analysis was conducted using MCScanX software[39], and the results were visualized using Circos[40] and TBtools to generate intra-species synteny plots. To investigate the synteny of the ANS gene across different species, genome and annotation files for O. sativa[41], A. thaliana, and V. vinifera[42] were downloaded from the Phytozome database (
https://phytozome.jgi.doe.gov/pz/portal.html ). The genome and annotation files for G. biloba[43] were obtained from NCBI. Inter-species collinearity analysis was performed using TBtools to visualize synteny across these species.Real-time quantitative PCR analysis
-
To validate the selected NcANS candidate genes, RT-qPCR was performed. The primers were designed and synthesized (Supplementary Table S1) by Sangon Biotech (Shanghai) Co., Ltd (Shanghai, China). RNA was extracted from the petals of N. colorata, and reverse transcribed into single-stranded cDNA, which served as the template for quantitative analysis. The RT-qPCR system was set to a final volume of 20 μL, containing 2 μL of cDNA, 1 μL each of forward and reverse primers, with thermal cycling conditions of 37 °C for 15 min and 85 °C for 5 s. Actin11[44] was used as the internal control gene for primer validation, and quantitative gene expression analysis of the candidate genes was performed.
-
In the genome of N. colorata, a total of 32 NcANS genes were identified and systematically named according to their chromosomal locations. Among the NcANS gene family members, the shortest protein sequence is NcANS4, with a length of 307 amino acids (aa), while the longest is NcANS17, with a length of approximately 500 aa. The molecular weights of these proteins range from 34,988.93 to 54,884.72 kDa, and the isoelectric points (pI) vary between 5.01 (for NcANS25 and NcANS27) and 8.54 for (NcANS30). Notably, the protein with a pI of 7 is classified as a neutral protein. In this study, four basic proteins (pI > 7) were predicted: NcANS29, NcANS30, NcANS17, and NcANS24, while the remaining 28 proteins were acidic (pI < 7) (Fig. 1).
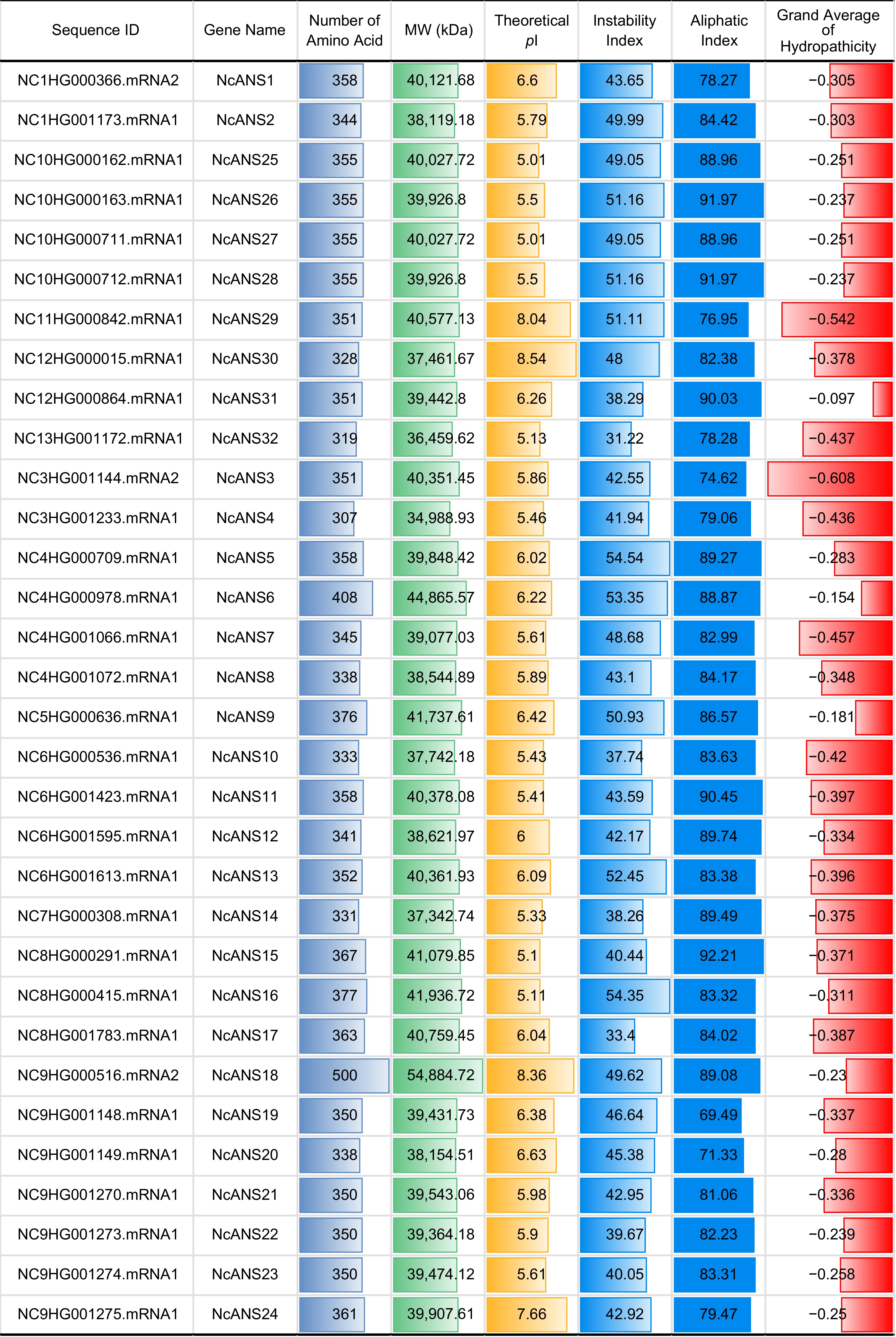
Figure 1.
Statistics of Nymphaea colorata ANS gene family members.
From the perspective of protein stability, six of the 32 NcANS genes (18.75%) were predicted to with an instability index lower than 40, classifying them as stable proteins, while the majority of the proteins were predicted to be unstable. Additionally, all these proteins were predicted to be hydrophilic. Based on these physicochemical characteristics, it is inferred that different members may play diverse roles in biological functions, especially in the pathways related to anthocyanin biosynthesis in N. colorata, reflecting functional diversification within the gene family.
The seven subfamilies of the ANS genes in flowering plants
-
To investigate the evolutionary relationships of the ANS gene family, we constructed a maximum-likelihood (ML) phylogenetic tree using 388 ANS proteins from six different plant species. These include 21 ANS genes from A. capillus, 52 ANS genes from A. trichopoda, 63 ANS genes from A. thaliana, 119 ANS genes from C. panzhihuaensis, 101 ANS genes from G. biloba, and 32 ANS genes from N. colorata. These proteins were ultimately classified, according to the presence of angiosperm ANS genes, into seven major groups (Fig. 2). Overall, ANS genes within the same subgroup are likely to share similar functional characteristics. Notably, NcANS genes were found in all clusters, clustering with homologous genes from other species, suggesting that the ANS gene family is not exclusive to angiosperms. Further analysis revealed that within each cluster, the phylogenetic relationship between NcANS and AtrANS was relatively close. This variation in homologous gene pairs suggests that the evolutionary relationship between N. colorata and A. trichopoda is more recent, whereas their relationships with outgroup species, such as ferns, are more distantly related.
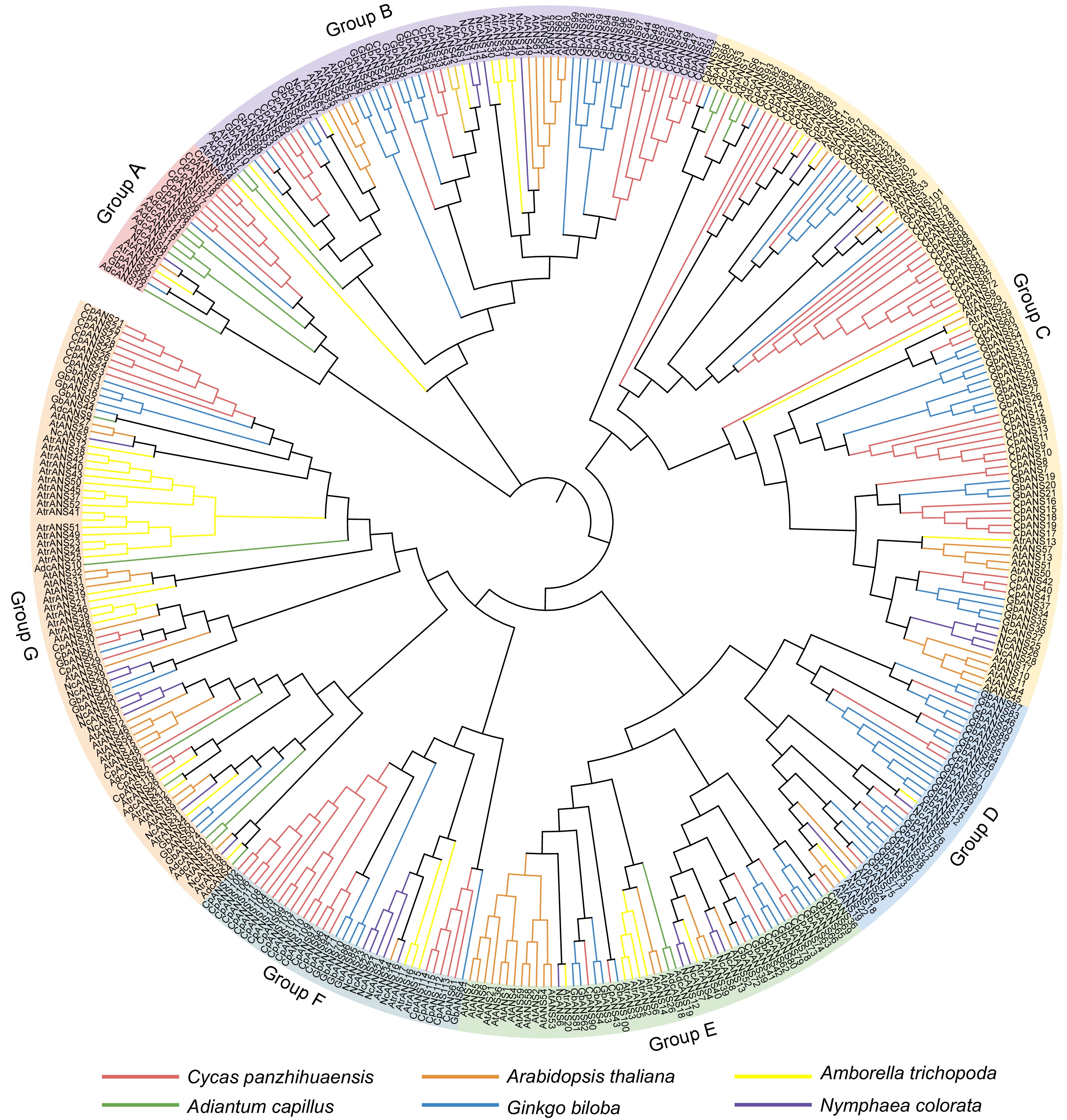
Figure 2.
Maximum-likelihood (ML) phylogenetic tree using 388 ANS proteins from six different plant species. The proteins included from Adiantum capillus, Amborella trichopoda, Arabidopsis thaliana, Cycas panzhihuaensis, Ginkgo biloba, and Nymphaea colorata. The branches of different colors represent different species. Green: A. capillus; Yellow: A. trichopoda; Red: C. panzhihuaensis; Blue: G. biloba; Orange: A. thaliana; Purple: N. colorata.
Structures of the ANS genes
-
Motif analysis of the NcANS genes revealed a total of 10 motifs (Fig. 3a). Most of the NcANS genes contain motif 10 and motif 6 at the N-terminus, while motif 9 is found at the C-terminus. Additionally, genes within the same evolutionary branch tend to share similar motif positions and numbers, indicating a potential functional synergy in their evolution. Notably, with the exception of NcANS18, which contains only one motif, all other genes contain more than four motifs.
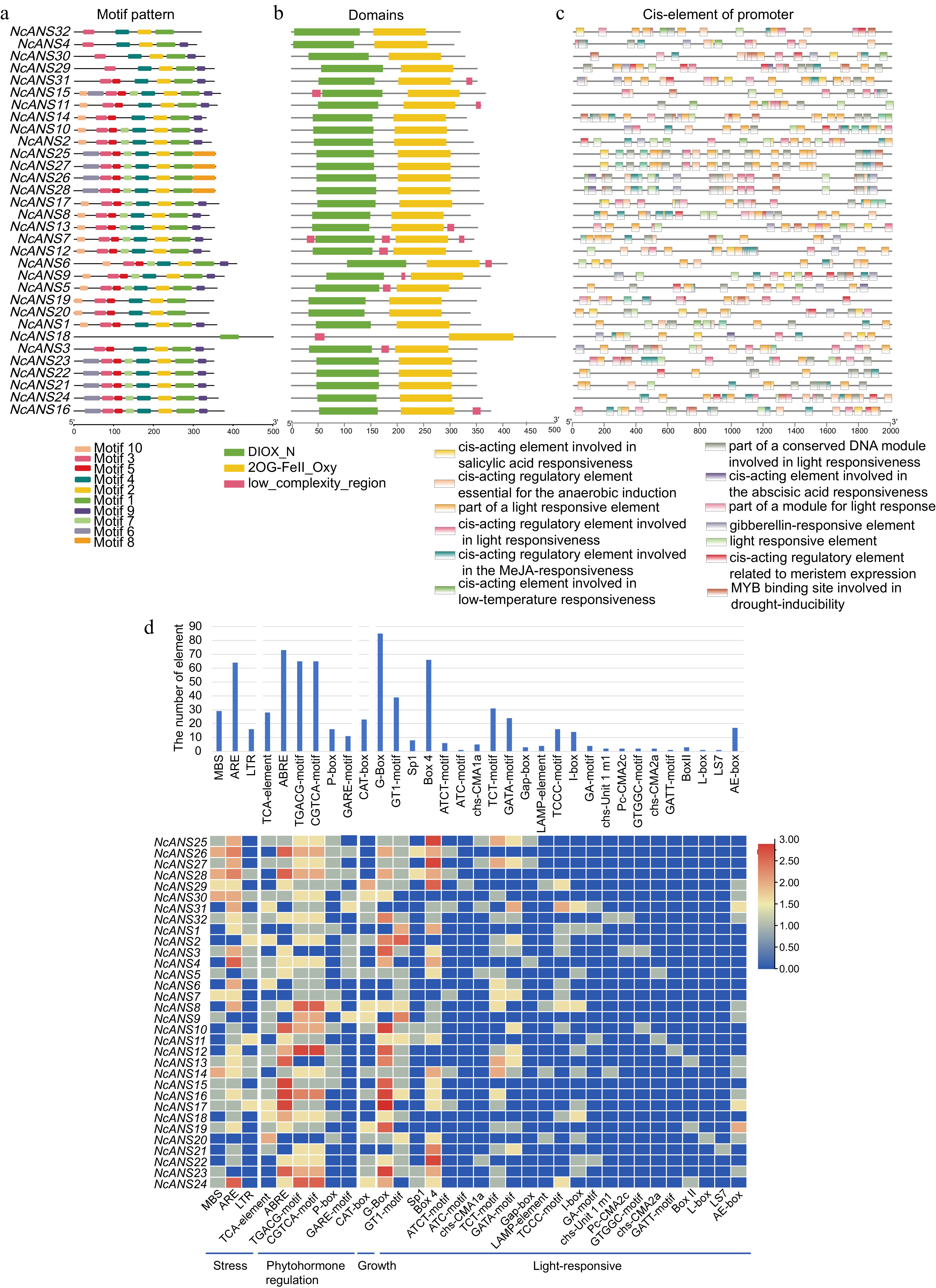
Figure 3.
Conserved motif, domain examination, and cis-regulatory element analysis of NcANS genes. (a) Motif analysis of the NcANS genes revealed that most the number and arrangement of motifs are similar within the same evolutionary branch. (b) Conserved domain analysis of the NcANS genes showed that, except for NcANS16, which contains only one 2OG-FE(II)-dependent oxygenase (2og-fe_oxy) domain, all other genes contain both 2og-fe_oxy and Diox conserved domains. (c) Cis-regulatory element analysis of the NcANS gene promoters revealed that all NcANS genes contain light-responsive elements, suggesting that they may play an important role in light-regulated anthocyanin metabolism. (d) The cis-elements were statistically classified into four categories: stress-responsive elements, hormone-responsive elements, growth-related elements, and light-responsive elements.
Conserved domain analysis using PFAM revealed that 31 genes in the N. colorata ANS genes contain both the 2OG-FE(II)-dependent oxygenase (2og-fe_oxy) and Diox domains, except for NcANS16, which contains only the 2og-fe_oxy domain. The 2og-fe_oxy domain is typically associated with the binding of iron ions and 2-oxoglutarate, serving as the core region for catalysis, while the Diox domain may be involved in the correct folding or activity regulation of the enzyme. These domains play a crucial role in anthocyanin biosynthesis (Fig. 3b).
To study the function of the ANS genes in N. colorata, we predicted and analyzed cis-regulatory elements in the 2,000 bp upstream sequences of their transcription start sites. The result shows that the promoters primarily contain light-responsive elements, hormone-responsive elements, MYB binding sites, salicylic acid-responsive elements, core promoter elements, anaerobic induction elements, and meristematic tissue expression-related elements (Fig. 3c). The cis-elements can be categorized into four groups: stress, phytohormone regulation, growth, and light-responsive elements. Among these, the phytohormone regulation and light-responsive elements are particularly abundant. Among them, the contents of methyl jasmonate (MeJA)-responsive elements and gibberellin (GA)-responsive elements in the hormone-responsive elements are both relatively high. Notably, all NcANS genes contain light-responsive elements in their promoters, suggesting that they may play a crucial role in anthocyanin metabolism under light regulation (Fig. 3d).
Chromosomal localization and collinearity analysis
-
To investigate the evolutionary relationships of the NcANS genes, an intraspecific synteny analysis of the N. colorata genome was conducted. The analysis identified five syntenic gene pairs within the NcANS gene family, located on chromosomes chr1, chr4, chr6, chr9, chr10, chr11, chr12, and chr13. Specifically, NcANS1 showed synteny with NcANS19; NcANS25 was syntenic with NcANS27; NcANS29 was syntenic with NcANS30; NcANS8 was syntenic with NcANS13 and NcANS7 was syntenic with NcANS12. These findings suggest that some members of the NcANS gene family may have arisen through gene duplication events and may possess functional similarities (Fig. 4a). To further investigate the evolutionary dynamics of the NcANS gene in N. colorata, we conducted synteny analysis using four representative model plant species with high-quality genome assemblies Arabidopsis thaliana, Oryza sativa, Ginkgo biloba, and Vitis vinifera along with the water lily genome. Figure 4b exhibited that N. colorata shares six syntenic ANS gene pairs with A. thaliana and O. sativa, five pairs with G. biloba, and up to 17 pairs with V. vinifera (Fig. 4b). These results suggest that the homologous genes of N. colorata were more abundant in dicotyledons than in monocotyledons.
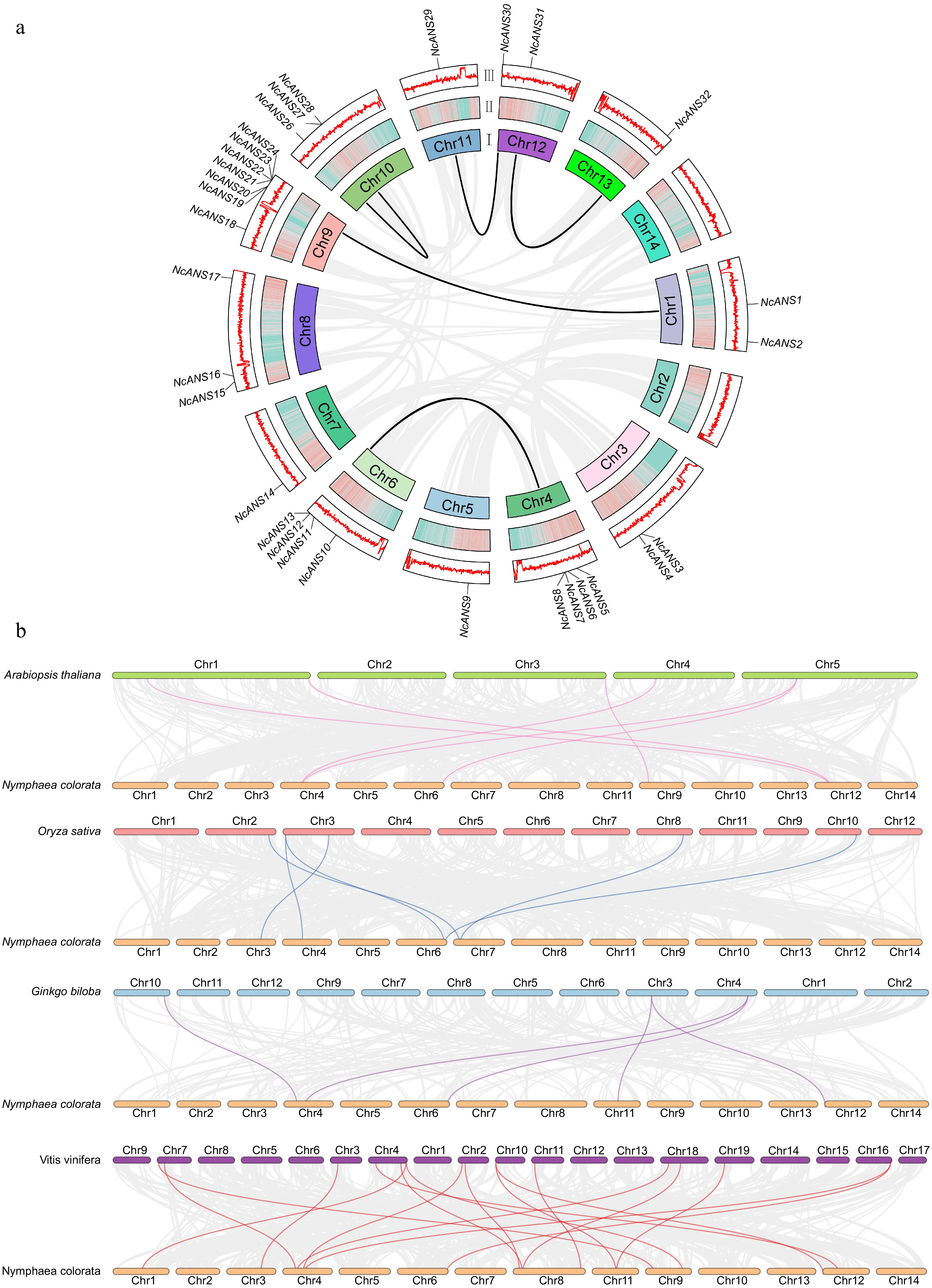
Figure 4.
Synteny analysis of the NcANS gene family. (a) Synteny analysis of NcANS genes. The gray lines represent all syntenic blocks across the N. colorata genome, while the black lines denote gene pairs between NcANS genes. The circular plot is divided into three layers: (I) Chromosome names; (II) Chromosome density; (III) Distribution of GC content across the chromosomes. (b) Synteny analysis of NcANS genes between N. colorata and four other representative plants. The gray lines indicate synteny between the N. colorata genome and the genomes of A. thaliana, O. sativa, G. biloba, and V. vinifera. The pink, blue, purple, and red lines represent syntenic gene pairs between NcANS genes and ANS genes from Arabidopsis, rice, Ginkgo, and grape, respectively.
Selection pressure analysis
-
The Ka /Ks ratio is a crucial indicator for assessing the evolutionary selection pressure on genes and is important for understanding the evolutionary mechanisms of the NcANS gene family (Table 1). Based on the five pairs of collinear genes in the NcANS family, we calculated the Ka/Ks ratios for four of the collinear gene pairs, all of which were less than 1. This result indicated that the NcANS gene family in N. colorata may have undergone purifying selection, which helps maintain the stability and conservancy of the gene family.
Table 1. Selection pressure analysis of NcANS homologous gene pairs.
Seq_1 Seq_2 Ka Ks Ka_Ks NcANS1 NcANS19 0.21 0.88 0.24 NcANS25 NcANS27 0 0 / NcANS29 NcANS30 0.131 0.94 0.14 NcANS7 NcANS12 0.18 1.06 0.16 NcANS8 NcANS13 0.17 1.32 0.13 Expression analysis of the NcANS genes in different flower colors of N. colorata
-
This study analyzed the flowering characteristics of three varieties of N. colorata and the expression patterns of the NcANS genes in different organs. The results indicated that, overall, NcANS genes were expressed across all organs. However, the expression levels were higher in blue and red flowers compared to white flowers. Figure 5a showed that the expression levels of most NcANS genes in both mature leaves and young leaves are not significantly pronounced. Notably, the expression levels of NcANS17, NcANS6, and NcANS3 were significantly elevated in floral organs, particularly in the petals and sepals, suggesting that these genes may be involved in biological processes related to floral organ development, such as pigment biosynthesis. NcANS18 consistently exhibited high expression in petals and sepals, while its expressions in other tissues were very low, suggesting that it may be a flower organ-specific expressed gene. In contrast, NcANS19 showed low expression across all tissues, with significant upregulation observed only in the anthers.
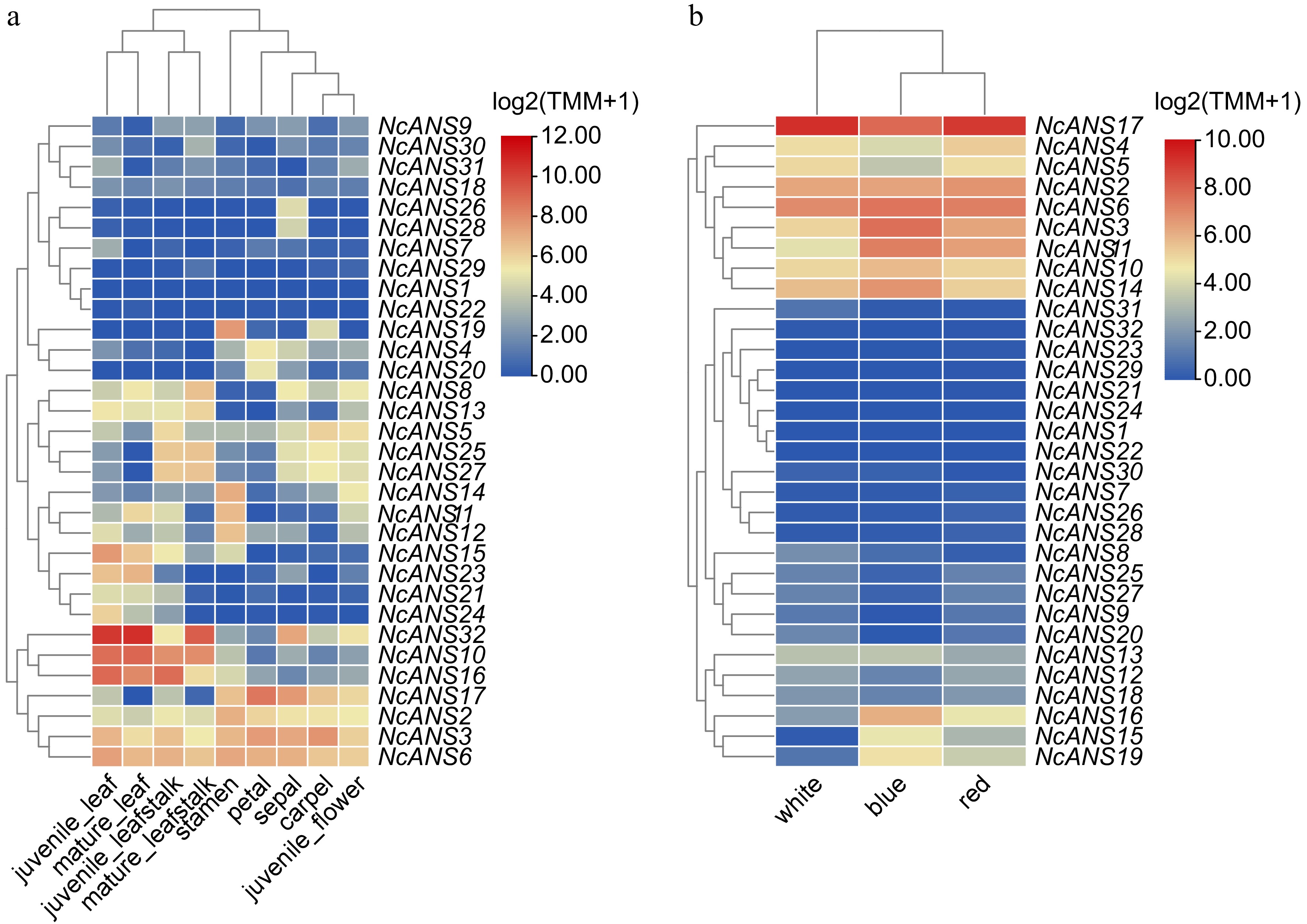
Figure 5.
Expression of NcANS genes in different tissues and flower colors of N. colorata.
To investigate the expression characteristics of NcANS genes in different flower colors of N. colorata, we analyzed their expression levels in white, blue, and red flowers (Fig. 5b). The results revealed that NcANS17 exhibited significantly high expression in all three flower colors, particularly in red and blue flowers, with expression in white flowers slightly higher than in red flowers. Additionally, NcANS3, NcANS6 and NcANS15 also showed relatively high expression levels, with the highest expression observed in blue flowers, while NcANS4 and NcANS11 exhibited slightly higher expression in red flowers compared to the other colors. In contrast, NcANS19 and NcANS16 exhibited clear flower color-specific expression patterns, with both genes being significantly upregulated in blue and red flowers and showing markedly reduced expression in white flowers.
Determination of anthocyanin content and expression analysis of NcANS genes in water lily at different colors
-
The expression patterns of NcANS19, NcANS16, and NcANS15 genes across different flower colors in N. colorata were analyzed using quantitative PCR (Fig. 6). The results revealed significant variations in expression among the three flower colors. NcANS19, the expression in blue flowers was higher than in red flowers, while in white flowers, it was nearly undetectable. Similarly, NcANS16 exhibited the highest expression in blue flowers, showing a marked difference compared to red and white flowers. NcANS15 followed a comparable pattern, with its expression in blue flowers significantly exceeding that in red flowers, and almost no expression observed in white flowers.
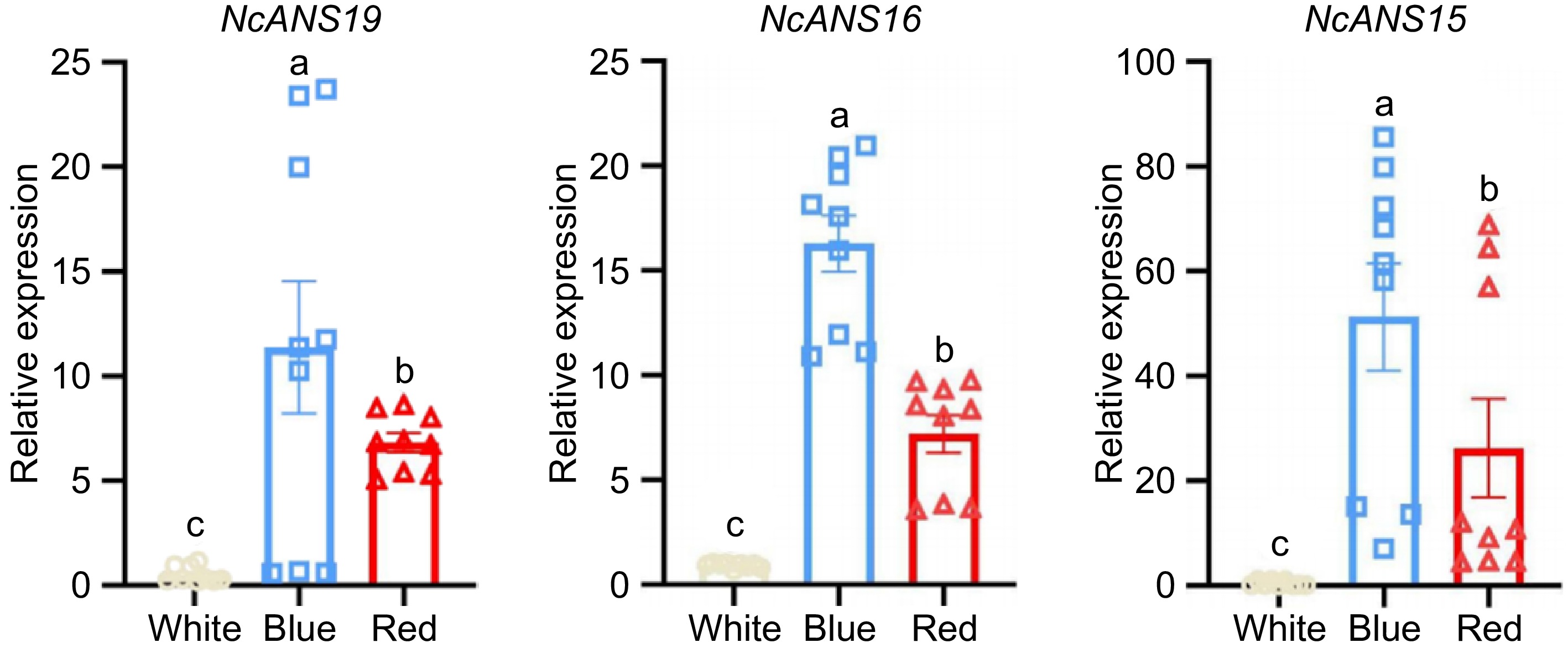
Figure 6.
Expression levels of the NcANS gene in different flower colors of N. colorata. Different lowercase letters indicate significant differences at the 0.05 level.
These findings show that the elevated expression of NcANS19, NcANS16, and NcANS15 in blue and red flowers likely promotes the upregulation of anthocyanin biosynthesis, resulting in darker pigmentation of the petals. Conversely, their minimal expression in white flowers may lead to insufficient anthocyanin production, accounting for the white or colorless petal appearance. This indicates that the ANS gene family plays a pivotal role in flower coloration in N. colorata and exhibits clear specificity among different floral varieties.
-
The foundational Nymphaea colorata genome has deepened the understanding of the traits of water lilies. The diverse flower colors of N. colorata are significant horticultural traits. This study focuses on the identification of key genes involved in the formation of rare blue anthocyanins. We systematically identified and analyzed 32 NcANS genes in N. colorata, all of which contain the 2OG-FE_OXY domain critical for anthocyanin biosynthesis. The presence of this domain not only provides the necessary enzymatic activity for anthocyanin synthesis but also reflects the high evolutionary conservation of the ANS genes. This characteristic is widely observed across different plant species, indicating that ANS genes are subject to strong selective pressures for conservation during evolution. Studies have shown that the RrANS of rose (Rosa rugosa)[45] and the IbANS of sweet potato (Ipomoea batatas)[46] both exhibit high sequence similarity and conservation with ANS genes from other plants, particularly in the conserved domain of the 2OG-FE(II) dioxygenase family. This further supports the evolutionary conservation of ANS genes.
The expression of ANS genes is closely associated with anthocyanin accumulation. Comparative transcriptome analysis of different flower color varieties of N. colorata revealed significant differential expression of NcANS genes at the transcriptional level, demonstrating distinct variety-specific regulatory patterns. The results indicated that the expression levels of NcANS genes in blue and red floral organs are significantly higher than those in white-flowered varieties. This pattern has been validated across multiple species: Feng et al.[22] further confirmed that differential expression of VvANS genes in grapes is closely correlated with color variation; Xiao et al.[47] found that the expression level of the ANS_1 gene is very high in the ornamental species of Musa ornata (Purple Banana) and Musa velutina (Velvet Banana). Yang[46] demonstrated that the expression of IbANS genes in purple-fleshed sweet potato (Xu Zi Shu 3) is markedly higher than in white-fleshed varieties (Xu Shu 18). Similarly, the transcriptional expression of StANS genes in colored potato tubers significantly exceeds that in yellow-fleshed cultivars[48]. Notably, red callus tissues of snow lotus (Saussurea involucrata) accumulate anthocyanins through specific upregulation of ANS gene expression, with expression levels far surpassing those in white and green tissues[49], further supporting the central role of ANS genes in pigment biosynthesis.
The expression of the ANS genes exhibits light-induced characteristics and hormone responsiveness. Analysis of the NcANS genes promoter region revealed several functional cis-regulatory elements, including light-responsive elements, hormone-responsive elements, stress-responsive elements, and growth-responsive elements. The presence of these elements suggests that the NcANS gene may be involved in anthocyanin biosynthesis through light regulation and hormone response mechanisms, which is consistent with previous studies on light-induced anthocyanin synthesis in tea plants[50]. For example, Jin et al.[19] found that the expression of the CsANS gene in tea plants is significantly regulated by light intensity, with a notable decrease in expression under low light conditions. The abundance of methyl jasmonate (MeJA)-responsive elements identified in this study aligns with the established role of jasmonates in promoting anthocyanin accumulation. For instance, MeJA treatment enhances the biosynthesis and accumulation of flavonoids, including anthocyanins, by regulating the expression of MYC transcription factors, which subsequently influence ANS gene expression[51]. However, a paradoxical phenomenon was observed regarding gibberellin (GA)-responsive elements. While GA typically suppresses anthocyanin synthesis in Arabidopsis through DELLA protein-mediated inhibition, the NcANS promoters in N. colorata exhibit an unusually high density of GA-responsive elements[52]. This contradiction may reflect aquatic adaptation in water lilies, where GA signaling potentially coordinates submergence responses and pigment production. This hypothesis is supported by the co-occurrence of 64 anaerobic response elements in the NcANS promoters identified in our experiments.
Expression patterns of ANS genes in different organs. Analysis of NcANS genes expression in different organs of N. colorata revealed that NcANS6 and NcANS3 are highly expressed not only in vegetative organs but also maintain relatively high expression levels in reproductive organs. This finding contrasts with studies on species such as peach[53] and orchid[54], where ANS genes are predominantly concentrated in floral organs. In this study, we hypothesize that this phenomenon may be related to the unique pigment distribution in the leaves of N. colorata—its leaves are not entirely green, with blue-purple pigments accumulating in the leaf axils. This may also be associated with anthocyanin biosynthesis.
Future research directions. Although this study revealed the expression patterns of NcANS genes in N. colorata, several questions remain to be addressed. First, while we predicted the expression patterns and relevant features of the ANS gene family using bioinformatics methods and identified key genes, experimental validation is lacking. Future research could integrate experimental approaches such as yeast two-hybrid assays and transgenic expression systems to validate the functions of NcANS genes. Second, this study was limited to the analysis of different organs and flower colors. Future studies could incorporate multiple time points to explore the dynamic changes of NcANS genes at various developmental stages.
In conclusion, this study, through comprehensive bioinformatics analysis, reveals that the expression patterns of NcANS genes may be regulated by both environmental signals (such as light and hormones) and endogenous developmental signals, exhibiting certain varietal specificity in N. colorata. The findings provide valuable insights into the role of NcANS genes in anthocyanin biosynthesis and color formation in N. colorata, laying a theoretical foundation for further research into the molecular mechanisms underlying flower color variation in this species.
This work was supported by the National Natural Science Foundation of China (32172614), and Hainan Provincial Natural Science Foundation of China (324RC452). We thank the editor and reviewers for their insightful comments and suggestions.
-
The authors confirm contribution to the paper as follows: study design and supervision: Chen F, Bai Y; data collection, and experiments: Yang S; data analysis: Yang S, Khan WU, Zhang J, Zhang J, Liang Y; draft manuscript preparation: Yang S, Chen F. All authors reviewed the results and approved the final version of the manuscript.
-
The raw sequencing data supporting the results of this study are stored in the Genome Warehouse in the China National Center for Bioinformation, under Accession No. PRJCA023065 (https://ngdc.cncb.ac.cn/gwh).
-
The authors declare that they have no conflict of interest. Fei Chen is the Editorial Board member of Tropical Plants who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0006)
-
Received 15 January 2025; Accepted 25 February 2025; Published online 24 March 2025
-
We successfully identified 32 NcANS genes in Nymphaea colorata and analyzed their cis-elements, revealing that ANS genes are regulated by light-induced expression and hormonal control.
The expression of NcANS genes varies significantly among different flower color varieties, showing distinct variety-specific expression patterns. Higher expression levels were observed in red and blue flowers.
Notably, the expression of NcANS was also detected in the leaves of N. colorata. This is attributed to the unique pigment distribution in the leaves of N. colorata (the leaves are not entirely green, with anthocyanin accumulating in the leaf axils), further indicating that this gene is associated with anthocyanin biosynthesis.
- Supplementary Table S1 Reference gene and candidate gene primer sequences.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yang S, Khan WU, Zhang J, Zhang J, Liang Y, et al. 2025. The sequence and expression analysis of anthocyanin synthase (ANS) genes in Nymphaea colorata with different flower colors. Tropical Plants 4: e011 doi: 10.48130/tp-0025-0006 

The sequence and expression analysis of anthocyanin synthase (ANS) genes in Nymphaea colorata with different flower colors
- Received: 15 January 2025
- Revised: 21 February 2025
- Accepted: 25 February 2025
- Published online: 24 March 2025
Abstract: Anthocyanins, a subclass of flavonoids, are key pigments responsible for diverse flower colors in plants. While the role of anthocyanin synthase (ANS) genes in regulating anthocyanin accumulation is well established in model plants, their evolutionary diversification and expression patterns in Nymphaea colorata remain largely unexplored. This study systematically identified 32 ANS genes (NcANS) from the N. colorata genome and analyzed their physicochemical properties, gene structures, evolutionary relationships, promoter cis-acting elements, and expression patterns across blue, red, and white petals. Synteny analysis demonstrated conserved and species-specific expansion of ANS genes across plant species. Phylogenetic analysis grouped these genes into seven subfamilies, suggesting functional diversification. Expression profiling and RT-qPCR validation revealed specific NcANS genes with higher expression in blue petals, highlighting their potential roles in pigment accumulation. These findings provide critical insights into anthocyanin biosynthesis in N. colorata and lay a foundation for future molecular breeding aimed at flower color modification.
-
Key words:
- Water lily /
- ANS gene family /
- Expression analysis