









-
In higher plants, photosynthesis primarily occurs in the foliage, where chlorophyll and carotenoids, essential for light absorption, reside. These pigments are not only critical for plant energy production but also influence their visual appeal and structural growth. Changes in leaf pigmentation, such as yellowing, often reflects shifts in environmental, genetic, or nutritional factors, impacting plant health and productivity. This phenomenon offers a valuable framework for delving into chlorophyll function and photosynthetic mechanisms, highlighting the necessity of unraveling the genetic drivers and control mechanisms tied to leaf color variations for enhanced agricultural outcomes and deeper botanical understanding.
Leaf variegation, evident as green and white patches on leaves typically arise from mutations impairing chloroplast genesis or pigment synthesis[1]. The genetic pathways responsible for this variegation in Arabidopsis mutants have been elucidated, pinpointing mutations in genes like FtsH that affect plastid stability[2]. In Helianthus annuus L., a mutation in the chlorophyll apoprotein A1 gene induces variegation[3]. In contrast, in Cymbidium sinense, the CsERF2 gene, belonging to the ethylene response factors family plays a key role[4]. Additionally, disruptions in carotene biosynthesis have been associated with abnormal plastids and variegated leaves in Brassica napus[5]. The study of 'Huabai 1', a natural albino tea plant mutant that exhibits a temperature-dependent shift in shoot color, has revealed significant insights into the molecular basis of leaf color change[6]. The discovery of the CsHO1 gene's impact on theanine levels in tea plants has shed light on the complex genetic interplay influencing leaf pigmentation and tea quality[7]. The analysis of the pepper yellowing mutant yl1 under different light conditions further underscores the relationship between light quality, chlorophyll content, and carotenoid metabolism, illuminating the genetic controls that dictate leaf coloration[8].
Research efforts have expanded our knowledge of the genetic elements that influence leaf coloration. For example, the identification of the CsaCNGC gene in cucumber as essential for the maintenance of youthful, green leaves emphasizes the specificity of genetic factors in pigment regulation[9]. Likewise, mutations in the CsVYL gene in cucumber and the role of SNPs in the 13-LOX genes provide a deeper understanding of the genetic intricacies behind leaf color variations[10]. Non-synonymous SNPs in the tandem 13-LOX genes have been recognized as causative mutations for yellow-green leaves[11], and mutations in CsHD, a gene encoding a protein with histidine and aspartic acid domains, lead to the yellow young leaf (yyl-1) phenotype in cucumber[12]. These advancements not only propel our understanding of plant physiological processes forward but also open avenues for genetic and biotechnological approaches to crop improvement. Another cucumber yellow leaf mutant, yl2.1, coding a plastid isoform of triose phosphate isomerase was identified as having high expression in cotyledons and leaves and affecting chlorophyll metabolism and chloroplast development[13].
UMP kinase genes encode enzymes that catalyze the conversion of uridine monophosphate (UMP) to uridine diphosphate (UDP), a crucial step in nucleotide metabolism. Additionally, these genes may have multifaceted roles in regulating gene expression. Previous research investigates the sole plastid UMP kinase (PUMPKIN) in Arabidopsis thaliana, revealing that it binds efficiently to RNA in vitro and associates with specific introns of plastid transcripts in vivo, thereby affecting their metabolism and influencing plant growth, plastid translation, and photosynthetic performance[14].
In this study based on a cucumber yellow leaf mutant, tnyl2, which showed a bleached phenotype. The tnyl2 mutant exhibits severely retarded growth and a photo-bleached phenotype due to compromised chloroplast structure and function. Genetic linkage analysis pinpointed CsUMPK as the candidate gene associated with these phenotypic deviations. CsUMPK, encoding a uridylate kinase, was crucial for chlorophyll synthesis and maintaining chloroplast integrity, as evidenced by reduced chlorophyll levels and disrupted chloroplast architecture in the mutant. These findings underscore the indispensable role of CsUMPK in sustaining photosynthetic efficiency and chloroplast development, providing valuable insights into the molecular mechanisms underpinning photosynthesis in cucumbers and potentially across other plant species.
-
From the Tnt1 retrotransposon insertion mutants of cucumber inbred line 9930[15], a mutant with photo-bleached cotyledons and leaves was identified and named tnyl2. Inbred line S06, known for its greenhouse suitability and regular green cotyledons and leaves, was utilized for comparison. The mutant seedlings displayed stunted growth and did not survive beyond the development of their fourth leaf. To explore genetic inheritance, both homozygous (+/+) and heterozygous (tnyl2/+) plants were self-pollinated and crossed with the S06 line. This crossing resulted in F1 progeny exhibiting a mix of green and yellow leaves, but only F2 offspring showcasing the yellow leaf phenotype were chosen for in-depth analysis. These selected plant materials were subsequently cultivated in the greenhouse of Shanghai Jiao Tong University (Shanghai, China).
Insertion–deletion and single nucleotide polymorphism marker development
-
For the genomic investigation, markers were identified through comprehensive whole-genome resequencing. Genomic DNA from the S06 plants was extracted using the CTAB method[16]. Subsequent resequencing of the S06 genome was conducted using the Illumina HiSeqTM 2000 platform, achieving a sequencing depth of 30×, with all resultant clean reads aligned to the reference cucumber genome of 'Chinese Long'[17]. Primer design for the study was carried out using Primer Premier 5.0, with all primers synthesized by Sangon Biotech (Shanghai, China). Details concerning the primers used are available in Supplementary Table S1.
DNA extraction and molecular marker analysis
-
DNA from the cucumber strains tnyl2, S06, and various F1 and F2 progeny was isolated using young leaf tissue through the CTAB method. PCR reactions were executed using an Applied Biosystems PCR apparatus. The resultant PCR products underwent electrophoresis in an 8% polyacrylamide gel, followed by a 200 V run for 1.5 h. Post-electrophoresis, gels were detached, immersed in a 0.1% solution of AgNO3 for staining, and then placed in a developer solution containing 1.5% sodium hydroxide and 0.4% formaldehyde to visualize the silver-stained DNA bands.
Mapping and fine mapping of tnyl2
-
To elucidate the genetic linkage between the tnyl2 locus and specific markers, the Bulk Segregant Analysis (BSA) strategy was utilized. DNA pools for the wild type and mutant were prepared by mixing equal quantities of DNA from five plants with green leaves and five with yellow leaves from the F2 generation. Indel markers that showed polymorphism between the wt-S06 and tnyl2 were selected for further analysis of these pools. The association of the tnyl2 mutation with these markers was examined using JoinMap 4.0 software, setting the linkage disequilibrium threshold at 4.0, across a cohort of 656 F2 individuals to refine the mapping of the tnyl2 locus.
Candidate gene identification
-
Candidate gene prediction was based on the 'Chinese Long' genome database (
http://cucurbitgenomics.org/ ). The functions of the candidate genes were analyzed using the BLASTP tool from NCBI. DNA fragments corresponding to candidate genes in this region were amplified from wt-S06, and tnyl2 genomic DNA and sequenced. Genome re-sequencing was utilized to identify the Tnt1 insertion sites in the candidate region. The specific Tnt1 insertion detection markers was designed to examine the insertion sites.Phylogenetic analysis and alignment of CsUMPK and its homologous proteins
-
A phylogenetic tree was constructed with the program MEGA X based on the sequences of CsUMPK protein and its homologous proteins in different species downloaded from the NCBI database (
https://blast.ncbi.nlm.nih.gov/ ). Protein sequence alignment was performed with Clustal W and the neighbor-joining tree was constructed, based on 1,000 bootstrap replications in MEGA X.RNA extraction and quantitative real-time PCR
-
Root, stem, and leaf of S06 were collected from seedlings at 10 d after germination. The experimental flower organs were collected from female flowers on the day of blooming. Total RNA was extracted with an Omini Plant RNA Kit (DNase I) (CWBIO, Taizhou, China) according to the manufacturer's instructions. First-strand cDNA was prepared with a HiFiScript cDNA Synthesis Kit (CWBIO, Taizhou, China). The quantitative real-time PCR (qRT-PCR) was conducted with Fast Start Universal SYBR Green Master Mix (ROX) (Roche, Shanghai, China) with a 7500 Fast Real-time PCR. The cucumber CsActin gene was used as the internal control. Quantification of the relative changes in gene expression was performed via the 2−ΔΔCᴛ method[18]. The qRT-PCR experiments were performed with three biological replications. Each biological replication contains at least three different individuals. Three technical repeats were performed for each biological replication.
Measurement of chlorophyll content and chlorophyll fluorescence kinetic parameters
-
Chlorophyll content was assessed by immersing 0.2 g of fresh leaves from both tnyl2 and wt-9930 seedlings in a mixture consisting of acetone, ethanol, and water (4.5:4.5:1 ratio) within 50-mL containers, shielded from light for 12 h. Absorbance readings of the solution were taken using a WFZ UV-3802H spectrophotometer at specific wavelengths pertinent to Chla and Chlb. This procedure was carried out on five yellow and five green leaves, collected during the morning hours under clear sky conditions, and was replicated three times biologically.
Chlorophyll fluorescence was measured using a MINI-PAM (Walz, Effeltrich, Germany). The maximum quantum efficiency of Photosystem II (PSII), Fv/Fm = (Fm–Fo)/Fm (where Fo is the minimal and Fm the maximal fluorescence yield in dark-adapted leaves), was measured at the beginning of the day from leaves that were dark-adapted for 30 min in dark leaf clips DLC-8 (Walz, Effeltrich, Germany). The experiments were performed with five biological replications. Each biological replication contained three different individuals.
Transmission electron microscopy
-
To prepare samples for transmission electron microscopy leaves from wt-9930 and tnyl2 seedlings were initially sectioned into small fragments and immersed in a 2.5% solution of glutaraldehyde within a phosphate buffer, maintained at 4 °C for 12 h. Then, they were washed and subjected to a secondary fixation with a 1.0% solution of osmium tetroxide, again at 4 °C, this time for 1.5 h. Following fixation, a sequential dehydration process using ethanol of increasing concentrations was conducted, culminating in embedding samples in Epon 812 resin. Ultrathin sections were prepared using a Leica UC-7 ultra-microtome and stained with uranyl acetate followed by lead citrate for contrast enhancement. These prepared samples were then examined under a Talos L120C G2 Spirit Biotwin transmission electron microscope to visualize the cellular ultrastructure.
-
A bleached mutant plant was identified in the M1 family of the Tnt1 retrotransposon mutant library (Fig. 1a). Compared with that of the wild-type plant (9930), the mutant led to severely retarded growth and a photo-bleached phenotype (Fig. 1b). Given the prior identification of the mutation in CsLPE2 within the Tnt1 mutant library, which leads to a yellow leaf at the seedling stage[19], this mutant was named tnyl2. The phenotype of the tnyl2 mutant is similar to that of cslpe2, with both exhibiting cessation of growth at the seedling stage. The full-length genomic DNA of the CsLPE2 gene from the wild-type material 9930 and the mutant tnyl2 were cloned. Sanger sequencing alignment showed no differences in the CsLPE2 sequences between the two materials, indicating that the yellow leaf phenotype in csyl2 is not caused by a mutation in the CsLPE2 gene (Supplementary Fig. S1). Therefore, we initially excluded the possibility that the tnyl2 phenotype was caused by mutations in the CsLPE2 through PCR analysis.

Figure 1.
Phenotypical characterization of cucumber mutant tnyl2 and wild type (WT) 9930. (a) Cotyledon color of the tnyl2 and WT. (b) Phenotypic identification of the tnyl2 mutant in the segregating population. (c) Chlorophyll contents in the WT and tnyl2 mutant leaves. Error bars represent mean ± SD (n = 5), mg/g. Student's t-test was performed, and statistically significant differences were indicated by **p < 0.01. (d) Phenotypes of the tnyl2 mutant and WT grown in MS medium with and without sucrose, respectively.
Subsequently, the growth potential of the tnyl2 mutant was tested in a sugar-supplied condition. On the sugar-supplemented medium, the tnyl2 mutant cotyledons were light yellow-green and grew for a slightly more extended period, even producing two true leaves. However, the leaf color was lighter than the wt-9930 plants (Fig. 1d). These results indicated that energy-supplied can partially complement the growth defects during the seedling stage of the bleached mutants, consistent with the findings reported for the CsTIC21 mutants[20]. Observing that the tnyl2 mutant, even with sugar supplementation, grows only until the second true leaf is fully expanded, after which its growth slows down (Supplementary Fig. S2). This indicates that the loss-function of CsUMPK cannot enable normal plant growth, even with the supplementation of photosynthetic products.
Mutation in CsUMPK was associated with the tnyl2 mutant
-
Due to the severely retarded growth condition by the mutant, which precludes self-pollination or cross-pollination for seed production, crosses were conducted between nine wild-type M1 plants and the inbred line S06. The progeny of these crosses were self-pollinated to produce F2 seeds, establishing a genetic segregation population. The progeny that contained the mutant phenotype was identified as the F2 segregation population, which was utilized for subsequent map-based cloning. Segregation analysis of leaf color in the F2 population confirms the mutation gene in tnyl2 is a single recessive nuclear gene (Table 1).
Table 1. Segregation analysis of leaf color in the F2 population of cucumber.
Population Total number Green leaf Yellow leaf Expected ratio χ2 F1 (tnyl2 × S06) 10 10 − − − F2 (tnyl2 × S06) 96 69 27 3:1 0.3221 F2 (tnyl2 × S06) 656 501 155 3:1 0.6103 χ2 (0.05,1) = 3.84. From 120 InDel markers distributed on seven cucumber chromosomes, 80 polymorphic markers between the wt-S06 and the tnyl2 were identified. Using the Bulked-Segregant Analysis (BSA) strategy, InDel6-8 and InDel6-9 were polymorphic between the wt-S06 pool and mutant phenotype pools. The two markers were located on chromosome 6 in cucumber. Genetic linkage analysis using 96 individuals of the F2 population showed that tnyl2 was linked with InDel6-7, and InDel6-9 (Fig. 2a). The F2 population, of 656 individuals were further used to fine-map the tnyl2 locus. After characterizing the F2 population, tnyl2 was mapped between the InDel markers InDel6-2-5 and InDel6-2-6, corresponding to a physical distance of 83.6 kb (Fig. 2b).

Figure 2.
Map-based cloning of genes for the tnyl2 locus of cucumber. (a) Primary mapping with 96 F2 plants placed the tnyl2 locus between InDel6-7 and InDel6-9 on chromosome 6. (b) Fine mapping with 656 F2 plants narrowed down the tnyl2 locus to an 83.6 kb region. (c) Nine genes were predicted in the candidate region. (d) Structural variation of the candidate gene, CsUMPK, with a 5.3 kb Tnt1 retrotransposon inserted. (e) Detection of the insertion mutation using CsUMPK specific molecular markers.
Considering that the occurrence of the tnyl2 mutant is attributed to the insertion of the Tnt1 retrotransposon, a genome Re-Seq of tnyl2 was performed to identify the Tnt1 insertion events within the 83.6 kb region. Within the nine genes in the candidate region (Fig. 2c), only one gene (Csa6G483490), annotated as uridylate kinase, had its exon disrupted by the insertion of a Tnt1 transposon, destroying the gene structure (Fig. 2d). To investigate the association between the mutation and the bleached phenotype, two sets of molecular markers designed to elucidate the linkage between the Tnt1 inserted gene and the bleached phenotype were generated. Within the F2 segregating population (N = 656), the insertion of Tnt1 exhibited a complete linkage (100%) with the bleached phenotype, indicating that this gene is the candidate gene responsible for the albinism trait.
UMP kinase (UMPK), the microbial/chloroplast uridine monophosphate kinase (uridylate kinase) that catalyzes UMP phosphorylation and plays a key role in pyrimidine nucleotide biosynthesis. Interestingly, a homolog of this gene in Arabidopsis encodes a UMP kinase located in the plastid, with Arabidopsis mutants displaying a similar bleached phenotype to that observed in cucumber[14]. Based on this, the gene was named CsUMPK.
Phylogenetic and expression pattern analysis of CsUMPK
-
The amino acid alignment of homologous proteins revealed that the CsUMPK homologs all contain a UMPK domain. This domain is highly conserved across the Cucurbitaceae family, closely related families Quercus, and in diverse species such as rice, maize, Arabidopsis thaliana, and tomato (Supplementary Fig. S3). A phylogenetic tree was constructed based on the amino acid sequences using the neighbor-joining (NJ) algorithm (Fig. 3a). The UMPK protein exhibits close genetic relationships among the Cucurbitaceae family. Within the closely related evolutionary lineage of the Quercus genus, homologous proteins of UMPK were also identified and clustered accordingly. Furthermore, the genetic distances of homologous proteins from tomato, Arabidopsis thaliana, maize, and rice were displayed, indicating comparatively greater evolutionary divergence.
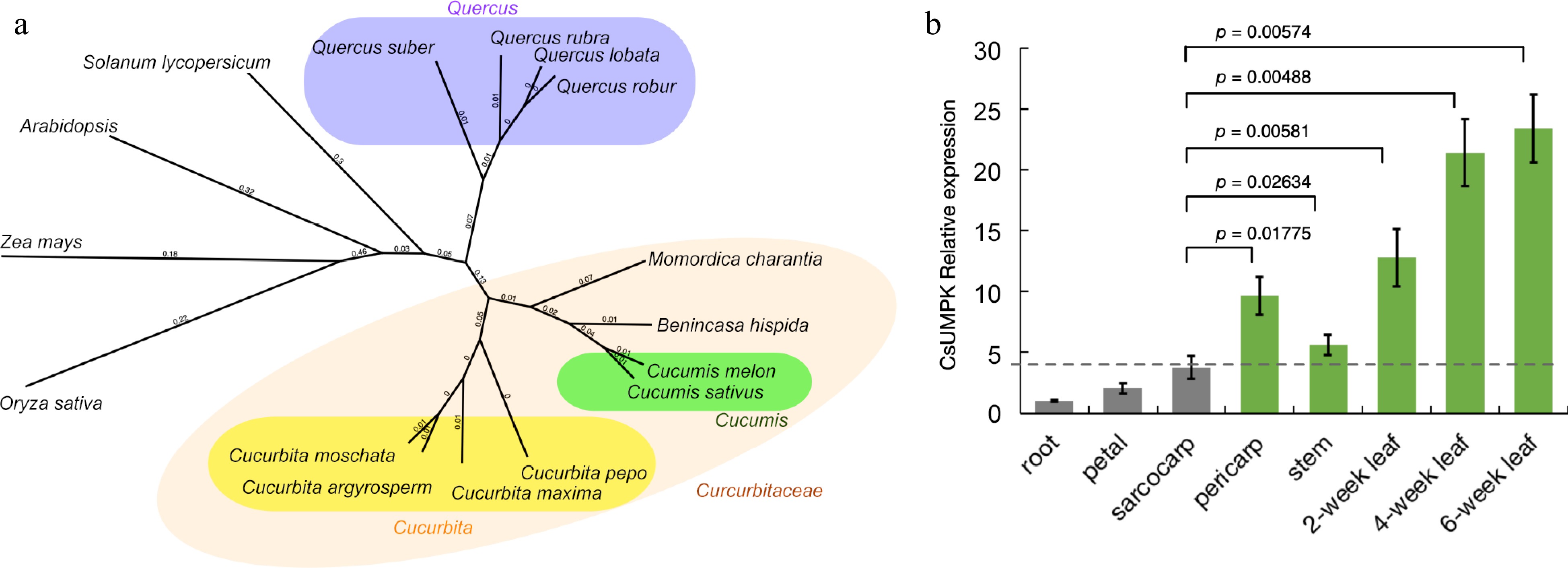
Figure 3.
Analysis of the evolutionary and expression characteristics of CsUMPK in cucumber. (a) Phylogenetic tree of CsUMPK protein in cucumber and its orthologs in other species. The phylogenetic tree was constructed by the Neighbor-Joining method with 1,000 bootstrap sampling. The numbers represent genetic distances. The protein sequences used in phylogenetic tree construction were from following species: Cucumis sativus (accession No. XP_004148645), Cucumis melo (XP_008441028), Benincasa hispida (XP_038880866), Cucurbita pepo (XP_023517300), Cucurbita moschata (XP_022962708), Momordica charantia (XP_022132364), Cucurbita argyrosperma (KAG7027137), Cucurbita maxima (XP_023003069), Quercus suber (XP_023896406), Quercus robur (XP_050292523), Quercus rubra (KAK4585494), Quercus lobata (XP_030975424), Solanum lycopersicum (XP_004235078), Oryza sativa (XP_015638950), Zea mays (XP_008654512), Arabidopsis thaliana (AT3G18680). (b) Relative expression of the CsUMPK gene. Relative expression of the CsUMPK gene in wt-9930 was measured by qRT-PCR in the root, petal, sarcocarp, pericarp, stems, and different stages of the leaf. For each sample, three independent biological replications were performed. Data are means ± SD (n = 3). Transcript levels of CsUMPK were normalized with CsActin and expression was shown relative to the root of wt-9930, the values of which were set as 1. The p-values were calculated using the Student's t-test.
The expression level of the CsUMPK gene in different organs (root, petal, sarcocarp, pericarp, stem, 2-week, 4-week, 6-week leaf) was examined by RT-qPCR. The results showed that CsUMPK was significantly highly expressed in the pericarp, stem, 2-week, 4-week, and 6-week leaves, compared with the sarcocarp (Fig. 3b). The high expression of CsUMPK in the green tissues suggest that its primary functional role is within these photosynthetically active regions. These results illustrate the evolutionary conservation of the CsUMPK gene, which appears to be integrally associated with photosynthetic activity in plants, as evidenced by its expression pattern.
CsUMPK is required for chlorophyll synthesis and chloroplast development
-
Transmission electron microscopy (TEM) analysis of the first true leaves from the wt-9930 and the tnyl2 mutant provided insightful revelations into the subcellular disruptions caused by the mutation. The ultrastructural examination delineated that chloroplasts in the tnyl2 mutant were marked by pronounced defects in the architecture of grana stacks and exhibited perturbations in the thylakoid membrane system (Fig. 4a−c). These morphological aberrations underscore a compromised photosynthetic capacity within the mutant chloroplasts. In stark contrast, the chloroplasts from the control specimens presented a well-organized structural integrity characterized by regular granal and stromal arrangements conducive to optimal photosynthetic function (Fig. 4d, e). Furthermore, a conspicuous sparsity in chloroplast distribution was observed in the tnyl2 mutant, with the grana being loosely organized and the starch grains markedly diminished.
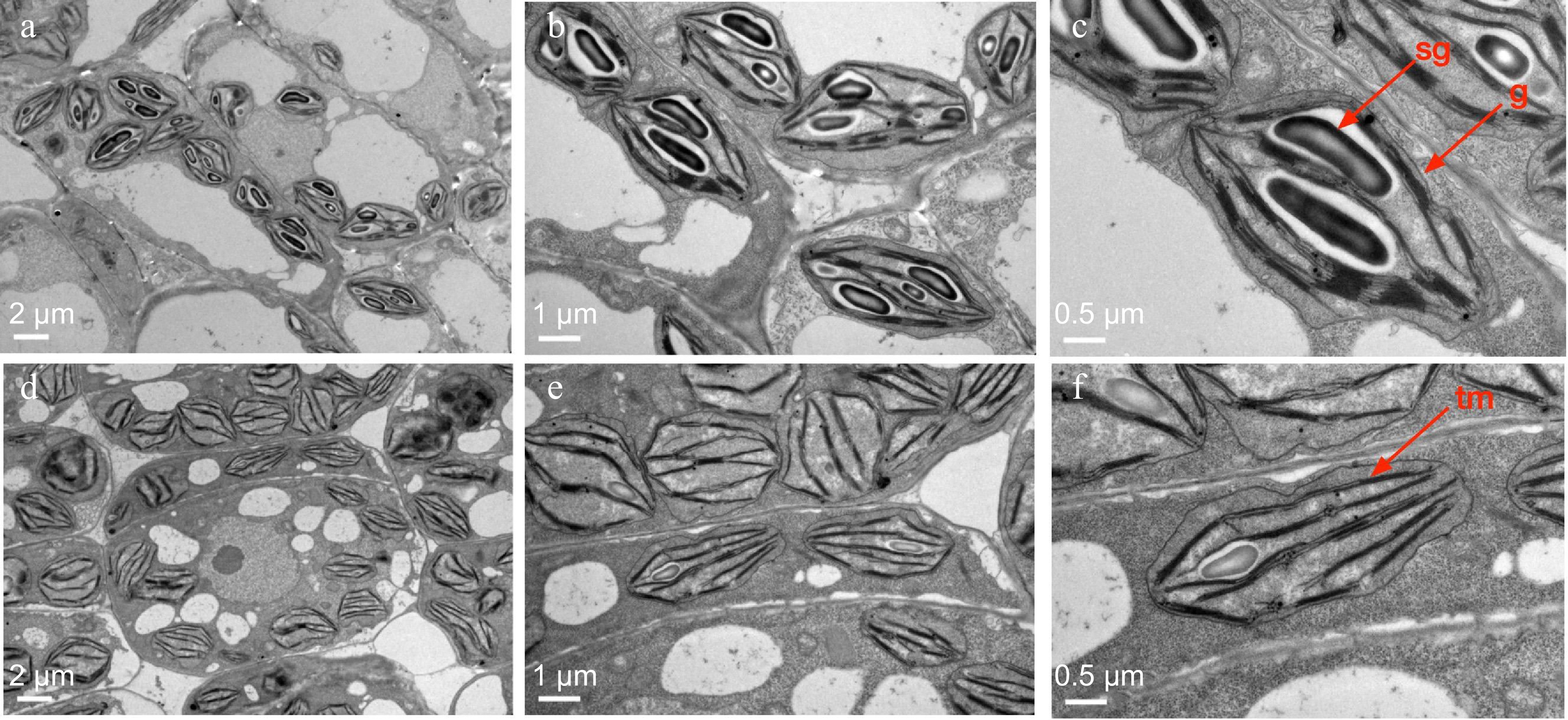
Figure 4.
Chloroplast ultrastructure of cucumber wt-9930 and tnyl2. (a)−(c) Chloroplast structure of WT, and (d)−(f) tnyl2. sg = starch granules; tm = thylakoid membranes; g = grana. Scale bars are shown in the figure.
Complementing the morphological observations, the quantification of leaf pigment content—encompassing total chlorophyll, Chla, and Chlb—was conducted for both the wt-9930 and the tnyl2 mutant. The measurements revealed a stark decrease in pigment levels in tnyl2 leaves, with total chlorophyll, Chla, and Chlb reaching only 59.7%, 34.3%, and 50.9% of the content in the tnyl2 leaves underpins the manifestation of the yellow leaf phenotype, delineating a clear correlation between pigment composition and leaf coloration phenotypes (Fig. 1c). To confirm if tnyl2 affected the photosynthetic capacity or not, several chlorophyll fluorescence parameters were measured, including minimum initial fluorescence (F0), maximum coefficient of fluorescence (Fm), photochemical quenching (qP), the coefficient of non-photochemical quenching (NPQ), and the actual photosynthetic efficiency of photosystem II (YII). The results demonstrated that almost all the chlorophyll fluorescence parameters of tnyl2 were lower than that of the 9930 (Table 2). These findings collectively attest to the mutation's pivotal impact on chloroplast functionality, implicating it as the culprit behind the observed phenotypic deviations and the ensuing photosynthetic insufficiency.
Table 2. Chlorophyll fluorescence kinetic parameters of young leaves of cucumber 9930 and mutant tnyl2.
Material F0 Fm Fv/Fm qP NPQ Y(II) tnyl2 0.07 ± 0.02** 0.20 ± 0.03** 0.43 ± 0.05** 0.25 ± 0.08** 0.34 ± 0.02** 0.19 ± 0.05** 9930 0.14 ± 0.03 0.45 ± 0.02 0.69 ± 0.01 0.69 ± 0.02 0.53 ± 0.01 0.53 ± 0.01 Five independent biological replications were performed. Each independent biological replication contains three different individuals. Three technical repeats were performed for each biological replication. Data are means ± SD (n = 5). Asterisks indicate statistically significant differences compared with the wild-type at ** p < 0.01 by Student's t-test. Loss-function-of CsUMPK impacts expression of CABs and chloroplast genes
-
Chlorophyll A/B binding protein (CAB) proteins, essential components of the Light-Harvesting Complex (LHC) complexes, play a significant role in plant photosynthesis. A comparison of CAB protein expression levels between the tnyl2 mutant and the wild type has revealed a marked reduction in the expression of multiple CAB proteins in the mutant (Fig. 5a). This decrease in CAB protein levels suggests potential disruptions in the light-harvesting capability of the tnyl2 mutant, which could lead to a diminished efficiency in photosynthetic energy conversion and overall reduced photosynthetic activity.
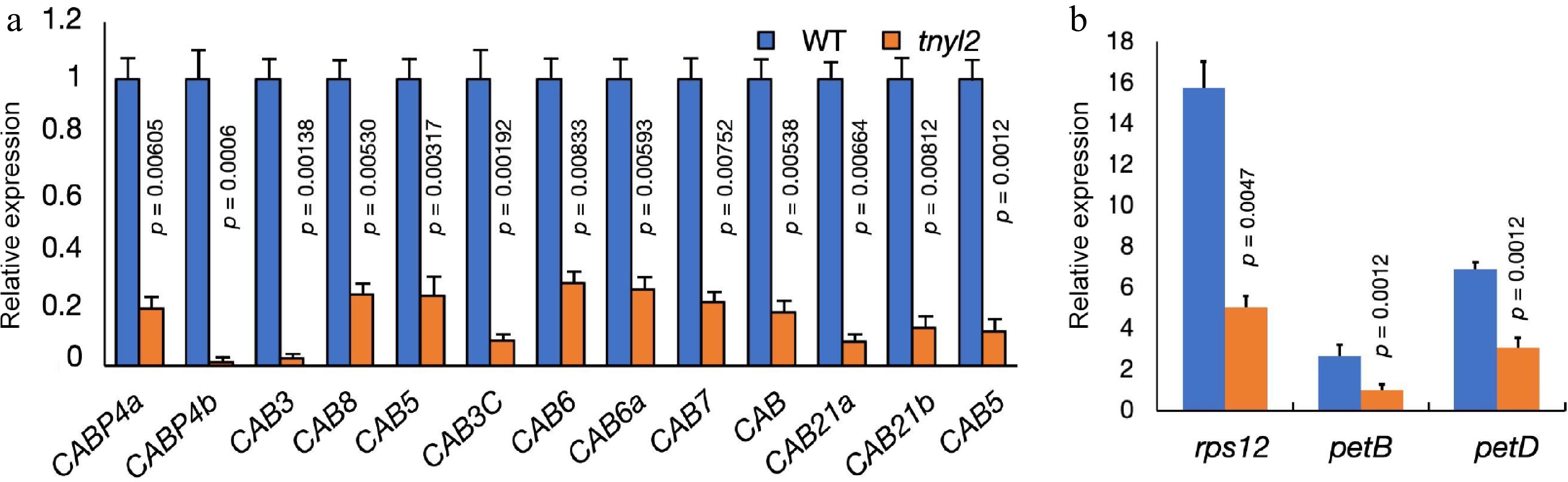
Figure 5.
Expression levels of chlorophyll-related genes in cucumber leaves. (a) Expression levels of the Chlorophyll a/b binding protein (CAB) related genes in wt-9930 and tnyl2. (b) Expression levels of chloroplast gene introns in the wt-9930 and tnyl2. For each sample, three independent biological replications were performed. Data are means ± SD (n = 3). The p-values were calculated using the Student's t-test.
In the model plant Arabidopsis thaliana, mutations in the PUMPKIN gene, which is homologous to CsUMPK in cucumber, result in the incorrect splicing of chloroplast gene introns. To investigate the effect of CsUMPK gene loss-of-function on the splicing of chloroplast gene introns in cucumber, RT-qPCR methods were utilized to assess the expression levels of intron regions within the chloroplast genome. The results indicated that the expression levels of introns rps12, petB, petD within the mutant were significantly higher than those in the wild type (Fig. 5b). This suggests that the loss-function-of CsUMPK function similarly affects the splicing of chloroplast gene introns in cucumber.
-
The investigation into the tnyl2 mutant presents significant insights into the complex interplay between genetic mutations, chloroplast development, and photosynthetic capacity in plants. The present findings delineate a multifaceted impact of the tnyl2 mutation, elucidating its consequences on chloroplast architecture, pigment synthesis, and the regulation of photosynthesis-associated genes.
Phenotypic manifestations of the tnyl2 mutation
-
The observable phenotypes of the tnyl2 mutant—characterized by retarded growth, photo-bleached leaves, and compromised structural integrity of chloroplasts—underscore the pivotal role of intact chloroplast development in maintaining plant vigor and photosynthetic efficiency. The morphological aberrations within the chloroplasts, particularly the disorganized grana stacks and the dilated thylakoid membranes hint at the underlying disruptions in the photosynthetic machinery. Such structural disarray within the chloroplasts unequivocally translates to the observed photobleaching, linking the mutation directly to the physiological processes central to plant survival.
Comparative phenotypic analysis of CsUMPK loss-of-function mutants
-
The phenotypic manifestations of CsUMPK loss-of-function mutants in cucumber are significantly more severe than those observed in Arabidopsis PUMPKIN mutants, with the former displaying pronounced difficulties in true leaf development. Conversely, Arabidopsis PUMPKIN mutants can complete their life cycle under greenhouse conditions. This disparity could stem from species-specific compensatory mechanisms or adaptive capacities to the loss of this gene's function. In cucumber, CsUMPK may play a more central role in photosynthetic processes and energy conversion, directly impacting plant development, especially true leaf formation. In contrast, Arabidopsis may possess greater biological redundancy or alternative pathways to mitigate the absence of PUMPKIN function, allowing survival and reproduction under certain conditions.
Impact of CsUMPK loss-of-function on photosynthetic gene expression and efficiency
-
The diminished expression of CAB proteins in the tnyl2 mutant provides compelling evidence of the downstream effects of CsUMPK loss-of-function on the light-harvesting complex. Given the crucial role of CAB proteins in capturing and transferring light energy, their reduced expression likely contributes to the impaired photosynthetic capacity observed in the mutant. Additionally, the altered splicing of chloroplast gene introns, notably in rps12, petB, and petD, further complicates the photosynthetic apparatus's efficiency, pointing to a systemic disruption in chloroplast gene regulation and expression. This dual impact on the photosynthesis pathway's structural and functional components encapsulate the critical nature of CsUMPK in plant photosynthesis.
In conclusion, the tnyl2 mutation and the associated loss of CsUMPK function unveils a critical dependency of plant photosynthesis on the proper development and maintenance of chloroplasts. The collective findings from this study not only shed light on the genetic underpinnings of photosynthetic deficiencies but also underscore the broader implications of chloroplast functionality on plant health and productivity.
This work was supported by the Department of Science & Technology of Liaoning (2022JH1/10200004).
-
The authors confirm contribution to the paper as follows: study conception and design: Pan JS, Pan J; experiments performing: Pan M; experiments assisting: Xia C, Gu S, He H; data analysis and draft manuscript writing: Pan M; assisting in data analysis and draft manuscript writing: Wang G, Weng Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primer information used in this study.
- Supplementary Fig. S1 Alignment of full-length genomic DNA sequence of the CsLPE2 gene in wild-type material 9930 and the mutant tnyl2. DNA sequence of CsLPE2 with 100% identity between the wild-type material 9930 and mutant tnyl2 are dark background.
- Supplementary Fig. S2 Phenotypes of the tmyl2 mutant and WT grown in MS medium with and without sucrose, respectively.
- Supplementary Fig. S3 Alignment of CsUMPK homologs from diverse species. Amino acid residues with 100% identity between the homologs are dark background.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Pan M, Xia C, Gu S, He H, Wang G, et al. 2024. Loss-function-of a UMP kinase leads to impaired chloroplast development and photosynthesis efficiency in cucumber. Vegetable Research 4: e034 doi: 10.48130/vegres-0024-0035 

Loss-function-of a UMP kinase leads to impaired chloroplast development and photosynthesis efficiency in cucumber
- Received: 23 May 2024
- Revised: 23 July 2024
- Accepted: 01 August 2024
- Published online: 06 November 2024
Abstract: Photosynthesis, a critical process occurring the chloroplast, supports essential metabolic pathways crucial for plant survival and adaptive responses. This study explores the cucumber tnyl2 mutant's impact on photosynthesis and chloroplast development, highlighting critical metabolic pathways essential for plant adaptability. The tnyl2 mutant, characterized by stunted growth and a photo-bleached phenotype, reveals a compromised chloroplast structure and function. Through genetic analysis, CsUMPK (Csa6G483490) emerged as the pivotal gene associated with these abnormalities, playing a vital role in chlorophyll synthesis and chloroplast integrity. Reduced chlorophyll levels and altered chloroplast architecture in the mutant underscore CsUMPK's significance. Moreover, the decline in CAB genes expression and potential intron splicing abnormalities due to CsUMPK dysfunction emphasize its essentiality in the photosynthetic apparatus. The present findings highlight the crucial contribution of CsUMPK to photosynthetic efficiency and chloroplast formation, offering insights into the molecular basis of photosynthesis across plant species.
-
Key words:
- Cucumber /
- Chloroplast development /
- Photosynthesis /
- CsUMPK