









-
Most Cucurbitaceae crops are annual trailing herbaceous plants characterized by long main vines and numerous lateral branches. For instance, watermelon vines can extend several meters in length, and fruits typically develop on the main stems or lateral branches of the vines. Consequently, substantial manual labor is required in production systems for regular vine training and lateral branch removal to ensure optimal fruit set and marketable quality. Under conditions where cultivation environments remain unchanged, and no additional costs are incurred for mechanized operations, developing short-vine varieties that eliminate the need for manual lateral branch removal plays a significant role in reducing labor costs.
Numerous genes controlling short-vine or dwarfing traits have been identified in Cucurbitaceae crops. Examples include genes involved in hormone biosynthesis and metabolism, such as CmaGA2ox2[1] in winter squash and CsDET2[2], CsDWF5[3], CsDWF7[4], CsSI[5], and CsCLAVATA1[6] in cucumber. Additionally, short-vine genes mdw1[7] and Cmdm1[8] have been reported in melon. Beyond these, CsaVFB1[9] and CsCYP85A1[10], which encodes the F-BOX protein, and cytochrome P450 enzyme, respectively, have also been shown to regulate vine length.
Watermelon ranks among the world's top seven fruit crops. China leads global watermelon production, accounting for over 40% of the cultivated area and more than 60% of the total output worldwide. Research on short-vine traits in watermelon dates back to 1956. The short-vine gene dw-1 confers a phenotype characterized by shortened vines, thickened stems, reduced branching, enlarged leaves, compact growth apices, and incomplete flower development in the material WB-2. In contrast, dw-2 regulates a distinct short-vine phenotype, mainly manifesting as shortened internodes, slender stems, increased branching, and normal floral development. dw-1s is an allelic mutant of dw-1[11]. Besides causing shortened vines, dw-4 leads to delayed-greening traits in cotyledons and shoot apices[12]. The watermelon GA20ox gene, which shares high sequence similarity with its homologs in cucumber and Arabidopsis, has been identified as a regulator of short-vine stature[13,14]. Mutations in the promoter region of ClaGA20ox can modulate its expression, thereby inducing dwarfism in watermelon plants[15]. GID1L2, located on chromosome 9 of watermelon causes both dwarfing and delayed growth and development[16]. However, most of the aforementioned short-vine genes are associated with undesirable agronomic traits beyond reduced vine length, limiting their direct utility in breeding. In contrast, short-vine genes identified later in germplasm resources such as SV-1[17], si302[18], and WCZ[19] hold greater breeding value. The short-vine phenotype in many watermelon accessions is determined by the ClGA3ox gene on chromosome 9, where a 13-bp deletion in its coding region leads to loss of function of the GA3ox protein[20−22]. Apart from gibberellin biosynthesis-related genes, vine-length regulation in watermelon also involves Cldw-1[23], which encodes an ATP-binding transporter protein. This gene participates in auxin metabolism and consequently affects internode elongation. A functional marker, dCAPS3, developed based on this gene, has been successfully applied in breeding for short-vine watermelon varieties[24,25].
For crops with a relatively narrow genetic background and a low natural mutation rate, employing EMS mutagenesis to create germplasm resources is one of the most efficient and convenient methods[26−28]. We constructed a pollen EMS mutant library using G42 as the genetic background and identified a short-vine mutant[29]. The mutant retains the excellent quality traits of G42 and possesses high breeding value. Through the identification of EMS-induced mutants in watermelon, a stably inherited short-vine mutant, G42cs, was obtained. In this study, we conducted a comprehensive investigation of the short-vine trait in this mutant and evaluated its potential breeding utility. The findings provide a cost-effective approach to mitigating labor shortages in watermelon cultivation.
-
The short-vine mutant G42cs used in this study was derived from a pollen EMS mutant library with G42 as the background. The wild-type G42 is one of our breeding backbone lines, exhibiting excellent comprehensive agronomic traits. From 2023 to 2025, at the Jiangsu Huai'an Academy of Agricultural Sciences research base, the short-vine mutant G42cs was crossed with its wild-type G42 to obtain an F2 generation population for this study. All plants were cultivated in greenhouse facilities using a trellising system with standard water and fertilizer management. Phenotypic investigation was conducted approximately 15−30 d after transplanting.
Investigation of agronomic traits
-
Agronomic traits of the short-vine mutant G42cs and wild-type G42 were investigated. Hypocotyl height: the distance from the soil surface to the base of the cotyledons in seedlings. Petiole length: the distance from the point where the petiole of a mature leaf connects to the vine to the point where it connects to the leaf blade. Main vine length: the maximum distance from the cotyledon node to the tip of the main stem. Tendril: the presence or absence, and the length of thread-like structures borne at nodes corresponding to leaves and flowers on mature vines. Fruit stalk length: the maximum length of the stalk connecting the fruit to the vine. Fruit weight: the average weight of mature fruit. Fruit longitudinal and transverse diameter: the maximum distance from the basal edge to the apical edge in the longitudinal section of the fruit, and the maximum distance between two edges perpendicular to the fruit's longitudinal axis. Rind thickness: the distance from the exocarp to the mesocarp (boundary between rind and flesh) at the middle of the sun-exposed side of the rind. Rind and flesh firmness: measured using a texture analyzer at three randomly selected points on the middle of the sun-exposed side of the rind, and three points in the middle of the flesh. Central and edge soluble solids content (SSC): sugar content at the fruit center and the fruit edge, measured with a refractometer. Seed length, width, and thickness: the maximum distance from the beak edge to the tail edge of the seed; the maximum distance between two edges perpendicular to the seed's longitudinal axis on the seed surface; and the maximum distance between the upper and lower surfaces when the seed lies flat, measured with a caliper.
DNA extraction and BSA-seq
-
The short-vine mutant G42cs was crossed with wild-type G42 to obtain an F2 population containing 641 individuals. When F2 plants developed two or three true leaves, approximately 0.1 g of young leaf tissue was collected in a 1.5 mL centrifuge tube for DNA extraction using the CTAB method. Based on phenotypic statistics, DNA was extracted from 30 individuals, each with the longest and shortest vine lengths using a kit (0418-50BB) from Beijing Huayueyang Biotechnology Co., Ltd. After concentration measurement, equal amounts of DNA from the 30 individuals were pooled to construct long-vine (L) and short-vine (S) bulks. These bulks, along with parental DNA were sent to Beijing Biomarker Technologies Co., Ltd. for sequencing.
Cell staining and observation
-
Approximately 15 d after transplanting, internodes from the 5th−8th node of G42cs and G42 were collected for paraffin sectioning. Three plants from each G42cs and G42 were selected, and three internodes were taken from each plant for embedding. When G42cs and G42 developed two true leaves, shoot apices were collected and fixed in FAA. Tissues were removed from FAA, dehydrated, embedded, and sectioned longitudinally to 8 μm thickness using a rotary microtome (Leica HistoCore BIOCUT). Sections were stained with toluidine blue following these steps: drying, dewaxing, hydration, staining, dehydration, clearing, and mounting[30]. Three biological replicates were performed, and observations were made using a stereomicroscope (Zeiss SteMI 508).
PCR and molecular marker analysis
-
PCR amplification was performed according to the instructions of the Master Mix for PAGE kit (#P115, Vazyme Biotech, Nanjing, China). Genotyping using the 12982321-dCAPS marker involved digesting the PCR amplicons with the restriction enzyme DdeI. The digestion was carried out at 37 °C for 4–8 h in a reaction mixture prepared following the manufacturer's protocol. The resulting fragments were resolved on a polyacrylamide gel for clear allele discrimination.
Seed production
-
Seed production was conducted in 6-m span plastic greenhouses. G42 and G42cs seedlings were transplanted using a ground-trailing cultivation method with plant spacing of 30 cm and row spacing of 3 m. G42 was pruned to two main vines, while G42cs was not pruned. Standard water/fertilizer management and manual self-pollination were applied, and labor input was recorded. Seeds were harvested 35−55 d after pollination: harvested fruit could be placed in a cool, dry place for 5−7 d of post-harvest ripening before seeds were extracted, promptly washed, dried, and then oven-dried at 40 °C for 2 d. Dried seeds were collected, weighed and recorded for yield comparison between G42 and G42cs.
-
The main vine length of G42 continued to increase with the number of days after transplanting, reaching approximately 200 cm at later stages (e.g., 37 d after transplanting), whereas the main vine length of G42cs increased more slowly, with a significantly lower final length of about 100 cm. The vine elongation rate of G42 was approximately 5–6 cm/d, compared to 3–4 cm/d in G42cs, indicating faster vine elongation in G42. Regarding fruit size, G42 exhibited greater longitudinal and transverse diameters as well as longer fruit pedicels than G42cs, with the pedicel length of G42 being nearly three times that of G42cs. During the early growth stage, the hypocotyl of G42 was approximately 2 cm longer than that of G42cs. At maturity, both petiole and tendril lengths of G42 significantly exceeded those of G42cs. Evaluation of fruit and seed traits showed that the fruit weight of G42 was around 2 kg, while the single fruit weight of G42cs was only half that of G42, consistent with the differences in fruit diameters, indicating smaller fruit size in the G42cs mutant compared to the wild-type G42. No significant differences were observed in soluble solid content (center/edge), flesh firmness, pericarp thickness/hardness, or seed morphology, suggesting that the modification in G42cs did not significantly affect internal fruit quality or seed structure (Fig. 1). G42cs exhibited a flowering time approximately 2–3 d later than that of G42, although this difference was not statistically significant. All other developmental stages were largely comparable between the two genotypes (Supplementary Fig. S1).
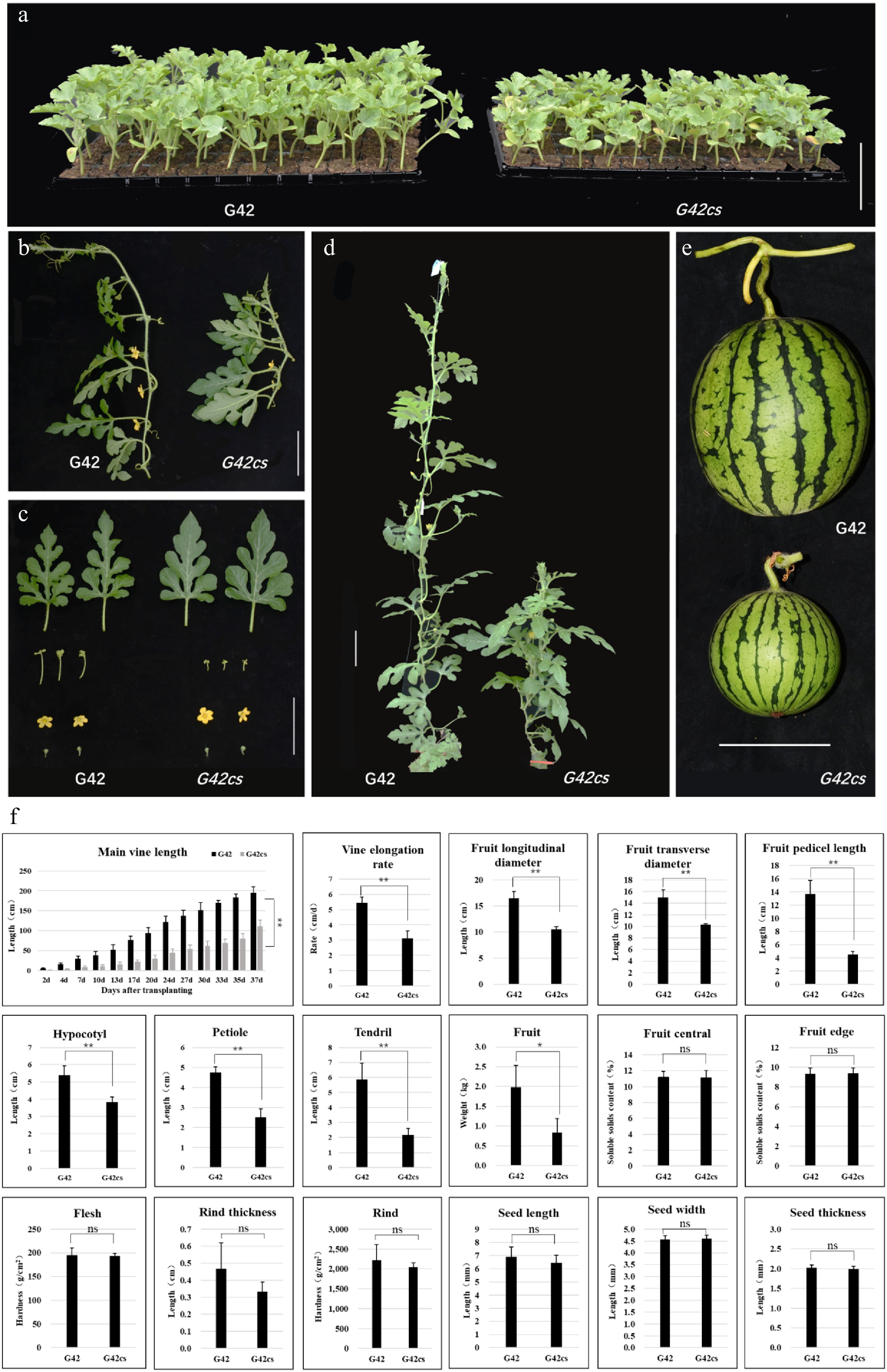
Figure 1.
Phenotypic and morphological characterization of watermelon plants from G42 and G42cs. (a) Seedling morphology of G42 and G42cs at the early growth stage. (b) Vine architecture: G42 exhibits longer, more elongated vines, whereas G42cs shows compacted vine growth. (c) Leaf and floral morphology: the G42cs line exhibits shallower leaf lobation and shorter tendrils compared to its counterpart, while floral structure remains unchanged. (d) Plant architecture: G42 produces a taller, more extended vine, whereas G42cs forms a shorter, more compact vine. (e) Fruit phenotype: G42 yields a larger fruit, while G42cs produces a smaller fruit. (f) Quantitative trait analysis. Main vine length; temporal dynamics of main vine elongation over 37 d after transplanting (DAT), with G42 consistently outperforming G42cs. Vine elongation rate, fruit longitudinal diameter, fruit transverse diameter, fruit pedicel length, hypocotyl length, petiole length, tendril length, and fruit weight: G42 shows significantly greater values than G42cs (p < 0.01 or p < 0.05). Fruit central/edge soluble solids content, flesh hardness, rind thickness, rind hardness, and seed length/width/thickness: no significant differences (ns) between G42 and G42cs. Statistical significance: *, p < 0.05, **, p < 0.01; ns = not significant (p ≥ 0.05). All data represent mean ± standard error (SE) from at least three biological replicates. Scale bars = 10 cm.
Differences in stem cells determine the vine length
-
Cellular staining observation of the apical tissues in G42cs and G42 seedlings revealed that the mutant exhibited a greater number of bud or leaf primordia (red arrows) and shorter bud axes (yellow arrows) compared to the wild type (Fig. 2a, b). Wild-type stem cells (Fig. 2c) were arranged neatly, while mutant stem cell arrangement (Fig. 2d) was more compact and crowded. Furthermore, stem cells of the mutant were smaller, and the number of cells per unit area (650 μm × 800 μm) was higher (Fig. 2e, f).
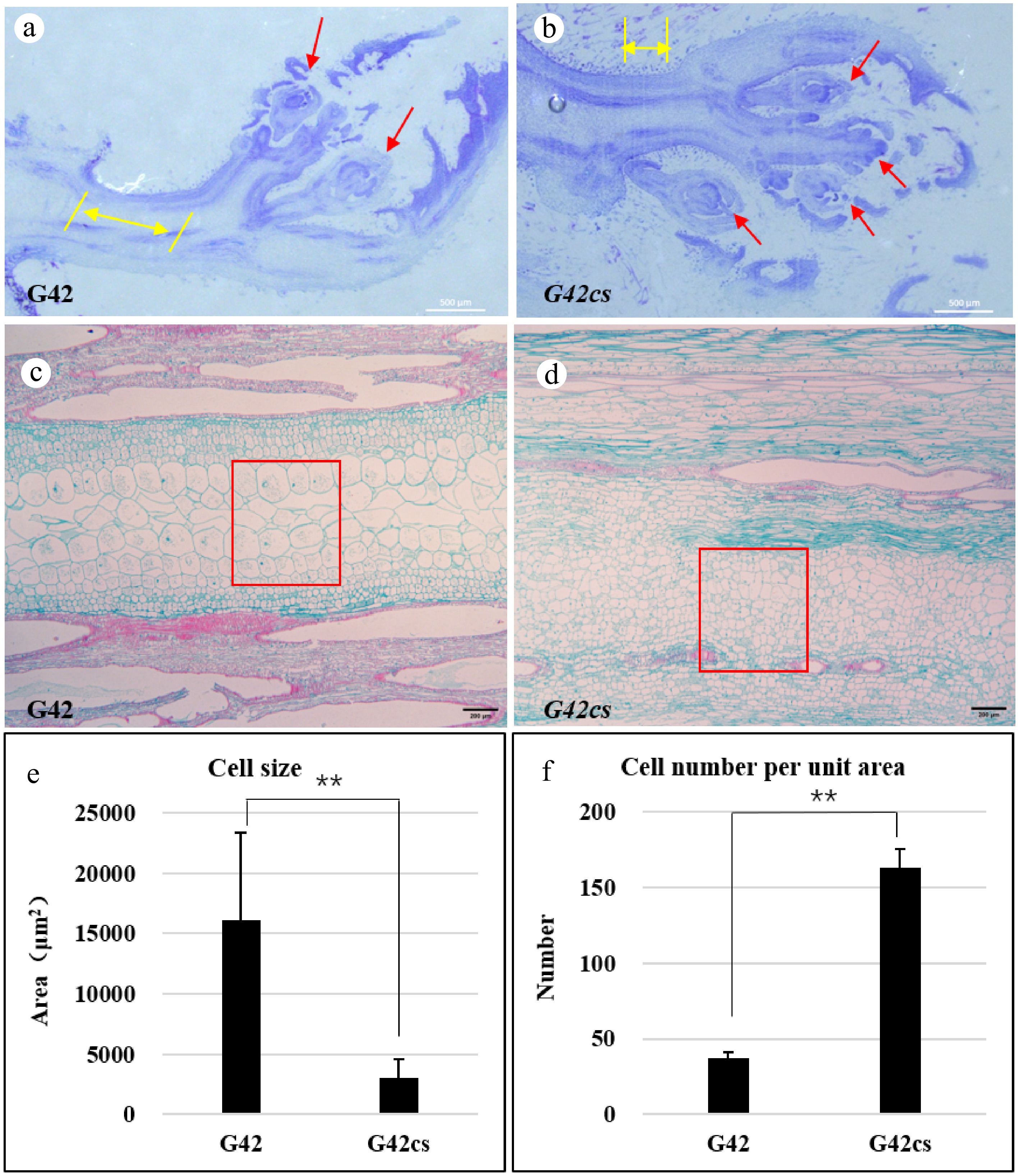
Figure 2.
(a), (b) Shoot apical meristem of wild-type G42 and short-vine mutant G42cs, scale bar = 500 μm (yellow arrows indicate bud axis, red arrows indicate leaf primordia). (c), (d) Observation of longitudinal stem section cells in wild-type and short-vine mutant, scale bar = 200 μm. (e), (f) Comparison of stem cell size and cell number per unit area between wild-type and short-vine mutant. **, p < 0.01.
Genetic analysis and mapping of the short-vine gene
-
An F2 population comprising 634 individuals was generated from a cross between the wild-type G42 and the mutant G42cs. Phenotypic evaluation of plant height revealed that all 30 F1 individuals exhibited a long-vine phenotype, with heights exceeding 100 cm at approximately 30 d after transplanting (prior to the reproductive growth stage). In the F2 population, substantial phenotypic variation was observed as early as 4 d after transplanting. Plant height was assessed around 30 d after transplanting, at which point 177 individuals were classified as short-vine (height < 80 cm) and 457 as long-vine (height ≥ 80 cm). Statistical analysis of plant height in the F2 generation (Table 1) showed a segregation ratio of long-vine to short-vine plants that fitted a 3:1 ratio. The calculated χ2 value was 2.879, which is less than the critical value of 3.841, indicating that the short-vine trait is controlled by a single recessive gene. For bulk segregant analysis (BSA), extreme pools were constructed by selecting 30 plants with the longest vines and 30 plants with the shortest vines from the F2 population. The plant heights of the long-vine pool ranged from 100 to 160 cm, while those of the short-vine pool ranged from 35 to 70 cm (Supplementary Fig. S2). Through BSA-Seq analysis and molecular marker development and validation, the candidate gene regulating the short-vine trait was preliminarily mapped to an interval between positions 9,078,429 and 13,817,428 on chromosome 1 (Fig. 3 and Supplementary Tables S1 and S2).
Table 1. Genetic analysis of short vine trait in watermelon.
Material/population Total plants Long vine Short vine Expected segregation ratio χ2 G42 30 30 G42cs 30 30 F1 30 30 F2 634 457 177 3:1 2.879 χ2 0.05 (1) = 3.841. 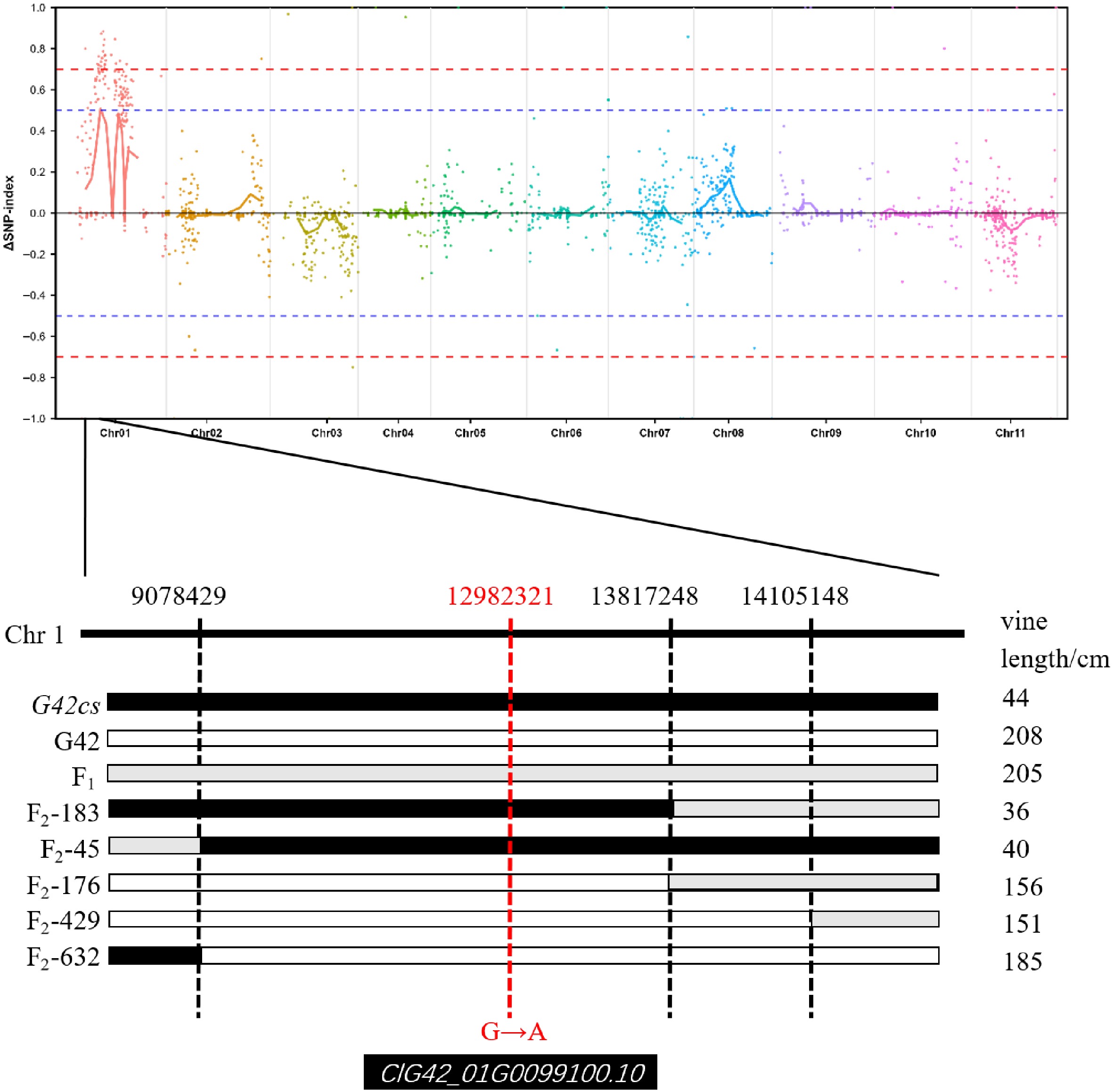
Figure 3.
Preliminary mapping of the short-vine gene by BSA-seq. Manhattan plot of SNP-index values across chromosomes (Chr01–Chr11). The red dashed lines indicate significance thresholds (± 0.7), and the blue dashed lines represent the baseline (0.0). A prominent peak is observed on Chr01, suggesting a strong association with vine length. Genotype-phenotype correlation for vine length (cm) in G42cs (44 cm), G42 (208 cm), F1 hybrid (205 cm), and F2 progeny (F2-183, F2-45, F2-176, F2-429, F2-632). Black bars indicate homozygous G42cs genotype, gray bars indicate heterozygous genotype, and white bars indicate homozygous G42 genotype. The red dashed line indicates that the differential SNP located within the candidate interval is in linkage with the vine length phenotype, and this SNP resides within the coding sequence (CDS) of the gene ClG42_01G0099100.10.
Candidate gene functional analysis and marker development
-
Within the candidate region, SNPs were found in only three genes: two in exons and one in an intron; the remaining SNPs in the interval were in intergenic regions. Thus, two candidate genes were identified (Supplementary Table S3): ClG42_01g0099100.10 and ClG42_01g0101900.10. ClG42_01g0099100.10 is annotated as a phosphate transporter. In G42cs, a G→A mutation occurs at position 1,580 bp in the CDS, resulting in an amino acid change from glycine to aspartic acid. ClG42_01g0101900.10 is annotated as belonging to the TRAFAC class dynamin-like GTPase superfamily. In G42cs, a G→A mutation occurs at position 1,576 bp in the CDS, changing the encoded amino acid from aspartic acid to asparagine. These G→A transitions are the primary mutation type induced by EMS treatment.
Based on sequence differences in these two genes between G42 and G42cs, dCAPS markers were designed to distinguish long/short-vine phenotypes. Validation showed that marker 12982321-dCAPS (Supplementary Table S1), developed based on the ClG42_01G0099100.10 sequence difference was essentially linked to the phenotype and can assist in screening for the short-vine trait (Fig. 4 and Supplementary Table S2).
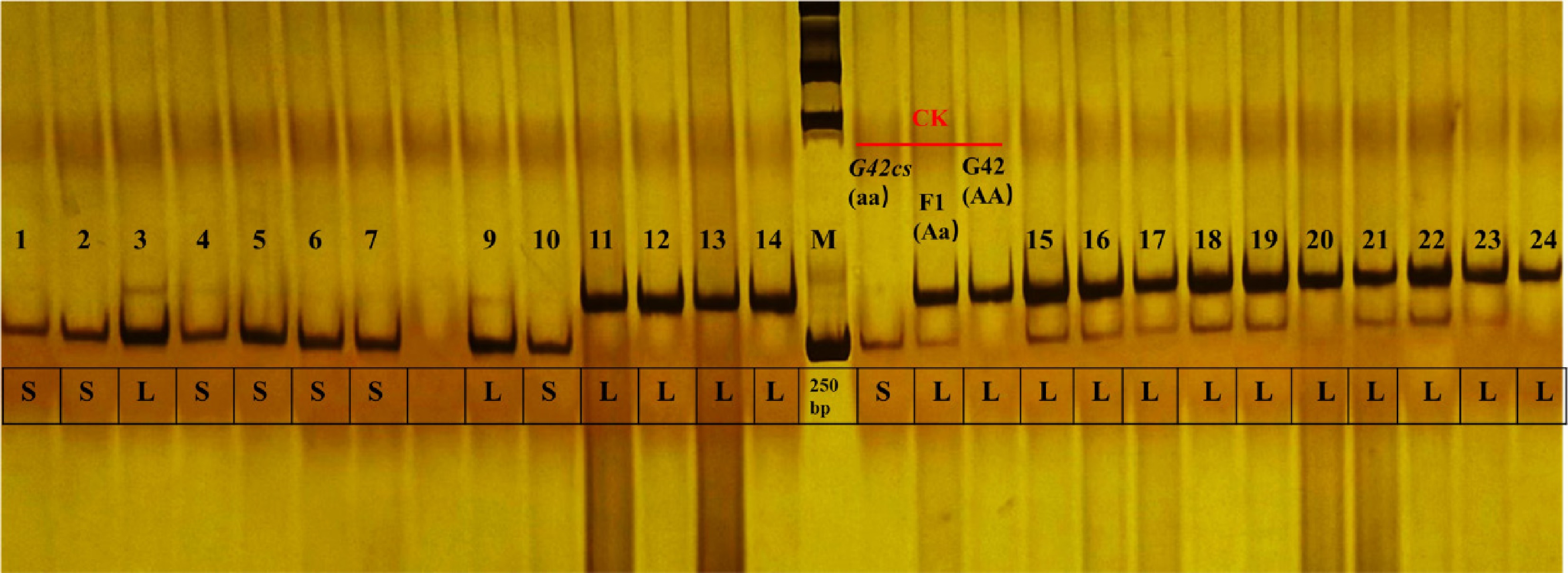
Figure 4.
F2 individuals were genotyped using the 12982321-dCAPS marker, and the resulting restriction fragments were separated by polyacrylamide gel electrophoresis. Genotypes were scored as follows: lower band, homozygous for the short-vine allele; upper band, homozygous for the long-vine allele; two bands, heterozygous. The size marker is 250 bp. L indicates that the plant exhibits the long-vine phenotype (plant height > 80 cm), and S indicates that the plant exhibits the short-vine phenotype (plant height < 80 cm). CK represents the parental lines of the F2 population, and F1.
Application of the short-vine mutant G42cs in breeding
-
G42 is one of our breeding backbone lines, and also the material for the T2T genome assembly of watermelon. The mutant library constructed from this inbred line has yielded many valuable materials, such as fruit shape variants and male-sterile mutants[19]. The short-vine mutant in this study also originated from this library. Due to its excellent agronomic traits, we used the 12982321-dCAPS marker for hybrid selection, developing it into a short-vine backbone line, G42cs-1. Hybrid combinations were made, resulting in a high-quality small-fruited watermelon F1 hybrid (Fig. 5). For hybrid seed production, large-scale propagation of female parent seeds is typically required annually. The short-vine trait, which eliminates the need for manual lateral branch removal and vine training, allows for higher planting density, and is controlled by a single recessive gene, and can be utilized for propagating seed producing female parents.
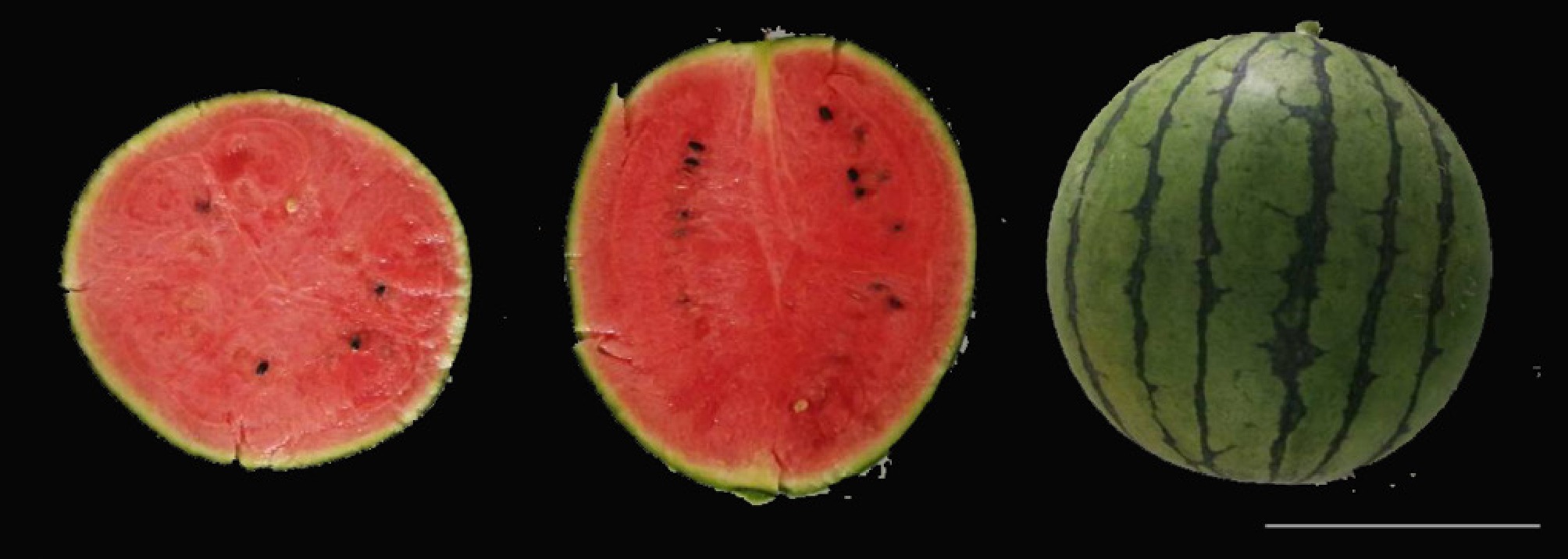
Figure 5.
A novel small-fruit watermelon hybrid was developed utilizing a dwarf trait: the short-vine gene was introgressed into the maternal parent G42cs-1, which was then crossed with a paternal parent to produce the new cultivar (Scale bar = 10 cm).
Both the short-vine backbone line G42cs-1 and the normal-vine line G42 were cultivated in 40-m-long plastic greenhouses with a span of 6 m, with approximately 400 plants per greenhouse. By recording and calculating the labor input costs for short-vine and long-vine plants, it was found that the key difference in input costs mainly lay in the pruning of lateral branches. Compared with the normal female parent, the short-vine female parent G42cs-1 saved 130 CNY in labor cost per greenhouse, which was entirely attributed to its trait of requiring no pruning, thereby significantly reducing labor input for field management (Table 2). In the most labor-intensive stage of emasculation and pollination, the operational costs for cultivating short-vine and long-vine plants were equivalent. This result confirms that the short-vine trait does not impede the efficiency of the essential hybridization procedure. Planting the short-vine female parent G42cs-1 holds a clear advantage in seed-production cost control. Although its seed yield per unit area is slightly lower, this can be compensated by adopting an appropriate higher planting density. Moreover, the reduced management costs may grant it better overall economic efficiency in large-scale production. In summary, the short-vine trait demonstrates applicable value in the propagation of female parents for watermelon hybrid seed production, and offers a new strategy for watermelon seed breeding.
Table 2. Labor cost input for F1 maternal seed propagation.
Per 400 plants (6 m × 40 m greenhouse) Propagating short-vine female parent G42cs-1 seeds Propagating normal female parent G42 seeds Seedling raising and transplanting 16.25 CNY/h × 2 h = 32.5 CNY 16.25 CNY/h × 2 h = 32.5 CNY Pruning lateral branches 0 16.25 CNY/h × 8 h = 130 CNY Emasculation and pollination 16.25 CNY/h × 32 h = 520 CNY 16.25 CNY/h × 32 h = 520 CNY Harvest 16.25 CNY/h × 2 h = 32.5 CNY 16.25 CNY/h × 2 h = 32.5 CNY Total (CNY) 585 715 Seed yield (kg) 7.4 8 -
Vine length is a crucial agronomic trait directly related to watermelon production management. Most current cultivars have long main vines and internodes, leading to low land use efficiency in ground cultivation and increased costs (e.g., trellising ropes) in vertical cultivation. Long vines also increase difficulty and labor intensity for pruning, spraying, and harvesting. In contrast, short-vine watermelons have a more compact structure, allowing higher planting density without pruning, optimizing land use, and reducing production and management costs. Recently, with urbanization and improved living standards, home gardening has diversified. Short-vine watermelons, suitable for potted vertical planting, easy to manage, and with small fruit ideal for single-family consumption, are gaining popularity. Consequently, demand for short-vine watermelons is increasing. However, due to limited germplasm resources, short-vine watermelon breeding progress has been relatively slow. US researchers improved undesirable traits like late maturity and disease susceptibility in short-vine materials through hybridization and selection, developing varieties suitable for pot or garden cultivation, such as 'Bush Charleston Gray', 'Bush Desert King'[11], and 'Bush Sugar Baby'[31]. Japan's 'Dwarf Red Gem Watermelon' is a representative short-vine variety. 'Wucha Zao' was the first short-vine, branchless watermelon cultivar bred in China[32], followed by 'Short Vine No.1', 'Short Vine No.2', 'Short Vine No.3', 'Hei Jin Bao', 'SS17', etc. However, due to various reasons like small fruit size, thick rind, poor disease resistance, low quality, and low farmer acceptance, these have not been widely adopted. Thus, high-quality short-vine resources are fundamental for breeding. The short-vine resource used here, G42cs, is an EMS-induced mutant from the small-fruited inbred line G42, retaining G42's excellent traits like high sugar, crack resistance, and high fruit set. Agronomic investigation showed significant differences from G42 only in plant height, tendrils, petioles, fruit stalk, and fruit size, with other traits largely similar (Fig. 1), making it valuable for breeding. Cellular observations further indicate that cell size and density directly affect internode length (Fig. 2), consistent with findings in cucumber and melon, where smaller cells and increased cell number per unit area lead to dwarfing[3,4,33,34]. To identify the gene regulating the short-vine trait, we constructed an F2 population from a cross between G42 and G42cs (Table 1). Through BSA-Seq and subsequent analysis, the candidate gene controlling the short-vine trait was delimited to a 4.5 Mb interval on chromosome 1 (Fig. 3). The marker 12982321-dCAPS, developed based on ClG42_01G0099100.10, successfully distinguished G42 and G42cs and co-segregated with the short vine phenotype in the F2 population (Fig. 4 and Supplementary Table S2). The developed short-vine inbred line G42cs-1 was used as the female parent in hybrid combinations, successfully yielding a high-quality F1 hybrid (Fig. 5). To ensure sufficient supply of F1 hybrid seeds, large-scale propagation of the female parent G42cs-1 is required. A comparison of the production costs between G42cs-1 and G42 during seed propagation revealed that the short-vine trait of G42cs-1 reduces labor inputs, resulting in a cost saving of approximately 18.18% (Table 2).
With the advancement of gene-editing technologies, targeted modification of gene sequences has become an efficient strategy to generate variation in plant architecture[35,36]. Knockout of ClNAC100 via CRISPR-Cas9 results in significantly reduced plant height and fruit size in watermelon, accompanied by decreased gibberellin (GA) levels[37]. Similarly, knockout of ClaCSLH1 leads to excessive hemicellulose synthesis, which inhibits internode cell elongation and ultimately produces a dwarf phenotype[38]. These two proteins influence plant dwarfing through pathways related to hormone biosynthesis and cell-wall synthesis, respectively. DELLA proteins in plants are negative regulators of the GA signaling pathway[39,40], directly inhibiting expression of cell elongation-related genes (e.g., expansin genes EXPA[41]), causing dwarfing. Phosphorylation of DELLA proteins is also crucial for height regulation[42]. Reducing GA biosynthesis in the sd1 mutant to promote DELLA protein accumulation can increase rice yield[43]. Recent research shows the rice SD1 gene not only regulates plant height, but also phosphorus uptake. In dwarf rice, the DELLA protein OsSLR1 interacts with the transcription factor OsMYB48-1, leading to reduced expression of inorganic phosphate (Pi) transporter genes (OsPT2 and OsPT10) and thereby restricting Pi uptake[44]. The Arabidopsis PHT4;6 phosphate transporter loss-of-function mutant displays dwarfism and is characterized by concurrent defects in protein glycosylation and cell wall hemicellulose biosynthesis[45]. This study identified the candidate gene ClG42_01g0099100.10 regulating the short-vine trait, which is annotated as a phosphate transporter (Supplementary Table S3). Its primary function involves phosphorus uptake, as well as the translocation and intercellular distribution of phosphorus within the plant. Through quantitative measurement, we found that the expression of this gene was significantly higher in the leaves and stems of G42cs than in those of G42 (Supplementary Fig. S3). This result is consistent with the observed phenotypic similarity between the two lines, i.e., no difference in floral morphology was detected between G42 and G42cs. We measured the hormone content in the stems of G42 and G42cs (Supplementary Fig. S4) and found that GA9 accumulated in the stems of the mutant G42cs. Previous studies have shown that the accumulation of GA9 can reduce GA3ox activity, thereby inhibiting the biosynthesis of active gibberellins and leading to a dwarf phenotype[46]. Meanwhile, ethylene levels in the stems of G42cs were higher than in G42, and elevated ethylene concentrations are known to inhibit cell elongation[47]. No difference in indole-3-acetic acid content was observed between the two lines. Based on these findings, we hypothesize that the phosphate transporter encoded by ClG42_01g0099100.10 may act as an upstream regulator in the network by influencing phosphate uptake and translocation in watermelon, which subsequently modulates both gibberellin biosynthesis and ethylene metabolism. Future work will focus on functional characterization of this phosphate transporter in the short-vine mutant G42cs to further elucidate the molecular regulatory network underlying the short-vine trait in watermelon.
-
This study conducted an agronomic trait investigation and stem histological analysis on a watermelon short-vine mutant, finding it has excellent comprehensive traits for breeding. Its short vine results from compact cell arrangement and smaller cells in stems. BSA-seq analysis mapped the controlling gene to chromosome 1. SNP and functional annotation analysis preliminarily identified ClG42_01g0099100.10, encoding a phosphate transporter, as the candidate. Molecular marker 12982321-dCAPS was developed for breeding. The mutant was used to develop new cultivars, and its application was shown to save approximately 18.18% of production costs.
This work was supported by the Open Research Project of Key Laboratory of Vegetable Biology of Hainan Province (HAASSC2025TJPT04); the Modern Agro-industry Technology Research System (CARS-25); and the Doctoral Research Startup Fund of Huai'an Academy of Agricultural Sciences (0012023011B).
-
The authors confirm contributions to the paper as follows: study conception and design: Liu X, Wang M, Sun Y; data collection: Deng Y, Shi X, Zhao S, Tang Y; analysis and interpretation of results: Liu X, Cheng R, Zhu Y, Xu B, Bai T, Xu W, Luo D; draft manuscript preparation: Liu X. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/vegres-0026-0013.
-
# Authors contributed equally: Xin Liu, Min Wang
- Supplementary Table S1 Marker information.
- Supplementary Table S2 Linkage analysis between the marker and the vine length.
- Supplementary Table S3 Candidate gene functional annotation.
- Supplementary Fig. S1 Phenotypic data of the extreme pools.
- Supplementary Fig. S2 Growth cycle of G42cs.
- Supplementary Fig. S3 Relative gene expression of CIG42_01G0099100.10 in different tissues (leaf, vine, and flower) between wild-type G42 and mutant G42cs.
- Supplementary Fig. S4 Endogenous levels of gibberellin A9 (GA9), ethylene, and indole-3-acetic acid (IAA) in the vine of G42 and G42cs.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu X, Wang M, Shi X, Zhao S, Tang Y, et al. 2026. Identification and gene mining of a short-vine watermelon mutant for breeding applications. Vegetable Research 6: e019 doi: 10.48130/vegres-0026-0013 

Identification and gene mining of a short-vine watermelon mutant for breeding applications
- Received: 29 January 2026
- Revised: 26 March 2026
- Accepted: 10 April 2026
- Published online: 30 May 2026
Abstract: In this study, a stably inherited short-vine mutant, G42cs, was identified and obtained from an EMS-mutagenized watermelon mutant library. Phenotypic characterization, genetic analysis, preliminary mapping of the short-vine gene, and research on its application in breeding were subsequently conducted on this mutant. Investigation of field agronomic traits revealed that the mutant exhibited significant differences from the wild type in traits such as plant height, tendrils, petioles, fruit pedicel length, and fruit size, while other traits showed no notable differences. Cellular staining observation of mutant tissues revealed that, compared to the wild type, the apical tissues of the mutant possessed more bud primordia or leaf primordia. The stem cells of the mutant were smaller than those of the wild-type watermelon, exhibiting a more compact and crowded arrangement, along with a higher number of cells per unit area. Measurements of plant height of the F2 population revealed that the short-vine trait is controlled by a single pair of recessive genes. Subsequently, BSA-Seq was performed using the two parents and the F2 population, which preliminarily mapped the short-vine gene to a 4.5 Mb region on chromosome 1. A dCAPS marker was developed to assist in screening for short-vine materials. Breeding backbone lines were developed using this short-vine mutant and applied in seed production. Assessments of seed yield per unit area and labor inputs showed that utilizing this short-vine trait in seed production can effectively reduce overall costs, demonstrating certain practical value. This model offers a new approach for watermelon seed production.
-
Key words:
- Watermelon /
- Short-vine /
- Breeding