









-
Anthocyanins are a vital class of flavonoid compound that predominantly exist as glycosides within plant vacuoles under natural conditions. Widely utilized as natural food colorants, anthocyanins not only impart vibrant pigmentation, but also confer multiple health benefits, including cardiovascular protection, enhancement of visual and cognitive function, regulation of adiposity and glycemic control, and anti-atherosclerotic, anticancer, and antiviral activities[1]. Consequently, efficient regulation of anthocyanin biosynthesis is of significant importance for plant quality improvement, including the enhancement of pigmentation and nutritional value[2]. Eggplant (Solanum melongena L.), a major crop in the Solanaceae family is highly valued by consumers for its anthocyanin-rich purple skin. The anthocyanin biosynthetic pathway is a specialized branch of the broader flavonoid pathway, conserved across many plant species. This pathway is primarily regulated by the coordinated action of structural and regulatory genes, with structural genes encoding the core enzymes responsible for anthocyanin synthesis. In eggplant, key structural genes—including SmCHS, SmCHI, SmF3H, SmF3'5'H, SmDFR, SmANS, and Sm3GT—have been implicated in pericarp anthocyanin accumulation. Notably, genes involved in the later stages of the pathway, such as F3'5'H, DFR and GT play a critical role in determining anthocyanin content and fruit coloration[3,4].
Regulatory genes encode transcription factors that modulate the expression of structural genes, either activating or repressing anthocyanin biosynthesis. Among these, MYB transcription factors—one of the most ubiquitous transcription factor families in plants—regulate gene expression by binding to MYB-specific cis-elements in the promoters of target genes[5]. MYBs are central to anthocyanin biosynthesis, functioning both independently and through interactions with bHLH and WD40 proteins to form the MYB-bHLH-WD40 (MBW) regulatory complex, which acts as either an activator or repressor. The transient expression assay indicated that SmMYB35, SmMYB44, and a SmMYB86 isoform might involve in the light-induced anthocyanin biosynthesis pathway[6]. Overexpression of SmMYB75 promotes anthocyanin biosynthesis in eggplant, and the color of the callus changes from green to purple[7]. While numerous MYB factors have been identified as positive regulators of anthocyanin biosynthesis, relatively fewer studies have focused on MYB-mediated transcriptional repression[8−12].
MYB44 has emerged as a key repressor of anthocyanin biosynthesis in several species. For instance, StMYB44 from potato (Solanum tuberosum L.) inhibits anthocyanin accumulation in tobacco leaves by directly repressing SmDFR promoter activity[13]. In sweet potato (Ipomoea batatas), IbMYB44s negatively regulate anthocyanin synthesis by disrupting the formation of the MYB340 (PAP1)-bHLH2-NAC56A/B transcriptional complex[14]. Similarly, MrMYB44-Like in begonia (Begonia L.) downregulates anthocyanin biosynthesis, leading to a color shift from red to green leaves[15]. In peony (Paeonia suffruticosa Andrews), PsMYB44 represses anthocyanin accumulation by directly binding to the PsDFR promoter[16]. Various key enzymes, such as chalcone synthase (CHS) and flavanone 3-hydroxylase (F3H), are involved in the biosynthesis process[17]. Therefore, in the present study, we investigated the molecular regulatory mechanism of the SmMYB44-l gene in eggplant anthocyanin biosynthesis by conducting yeast one-hybrid and dual-luciferase reporter assays on the promoters of SmCHS and SmF3H, with SmMYB44-l. Our objective was to elucidate its functional role as a potential repressor, enhance understanding of the anthocyanin regulatory network, and contribute to the theoretical foundation for the genetic improvement of eggplant quality.
-
The eggplant materials used in this study included 'Bailong', 'W185', 'March eggplant', and 'NC7', and were all provided by the Vegetable and Flower Research Institute of the Chongqing Academy of Agricultural Sciences (Supplementary Fig. S1). Additional experimental materials included Nicotiana benthamiana, competent Escherichia coli cells, and various molecular biology vectors and strains: the pEASY®-Blunt Simple Cloning Kit (TransGen Biotech), Agrobacterium tumefaciens strains (Shanghai Weidi Biotechnology Co., Ltd), yeast strains Y1HGold and Y187, the subcellular localization vector pCAMBIA1300-GFP, transient expression vector pGBO, overexpression vector pCAMBIA-2301G, yeast one-hybrid vectors pGADT7 and pAbAi, luciferase reporter vectors pGreen II 62-SK and pGreen II 0800-LUC, and luciferase complementation vectors pCAMBIA1300-nLUC and pCAMBIA1300-cLUC. All vectors and strains were sourced from our laboratory.
Eggplant genomic DNA extraction
-
The DNA was extracted from young leaves of 'March eggplant'. Tiangen Biotech (Beijing) Co., Ltd's Plant Genomic DNA Kit was used in accordance with the manufacturers instructions.
Total RNA extraction and cDNA synthesis in eggplant
-
Total RNA was extracted from young leaves of 'March eggplant' using the Trizol Plant Total RNA Extraction Kit (Tianmo Biotech), followed by evaluation of RNA concentration, purity, and integrity. First-strand cDNA synthesis was carried out using a reverse transcription kit from Takara according to the manufacturer's instructions. The resulting cDNA products were stored at −20 °C for further use.
Gene cloning and bioinformatics analysis
Gene cloning
-
To identify homologous sequences, the SmMYB44-l gene was annotated based on tomato, potato, and other Solanaceae gene sequences from the NCBI database (
www.ncbi.nlm.nih.gov ), and the eggplant sequences from the high-quality genome database (http://eggplant-hq.cn ). Gene-specific primers were designed using Oligo 7 software (SmMYB44-l-F: ATGGCGGCGATTGCACAG; SmMYB44-l-R: TCATGTCTTGCAGAACGG). PCR amplification of the target gene was performed using eggplant cDNA as a template, and the amplified products were sequenced by Tsingke Biotechnology Co., Ltd.Bioinformatics analysis of genes
-
The physicochemical properties of the SmMYB44-l protein were analyzed using the ProtParam tool (
https://web.expasy.org/protparam ). Secondary structure prediction was conducted with the GOR method (http://pfam.xfam.org/search/sequence ), and three-dimensional structure modeling was performed using ExPASy SWISS-MODEL (http://swissmodel.expasy.org ). Sequence alignment and phylogenetic tree construction were completed using DNAMAN and MEGA 8.0 software.Real-time quantitative fluorescence analysis (qRT-PCR)
-
Expression patterns of SmMYB44-l were analyzed using cDNA samples derived from eggplant pericarps of four colors (white, green, purplish red, and purplish black) and from roots, stems, leaves, flowers, peel, and pulp of white eggplant 'Bailong' and purple-black eggplant 'NC7' (Supplementary Fig. S2). GAPDH was used as the internal reference gene.
Based on the gene sequence of SmMYB44-l and standard primer design principles, specific primers for SmMYB44-l, and the reference gene GAPDH were designed using Primer3Plus (Supplementary Table S1). The qRT-PCR reactions were conducted following the protocol provided with the NovoScript® SYBR qPCR SuperMix Plus (Novoprotein Biologicals).
Determination of anthocyanin content
-
Approximately 500 mg of peel or leaf tissue was ground to a fine powder in liquid nitrogen using a mortar and pestle. Anthocyanins were extracted in 5 mL of a methanol: hydrochloric acid solution (99:1, v/v), incubated at 4 °C in darkness for 24 h, and centrifuged at 13,000 rpm for 20 min at 4 °C. The absorbance of the supernatant was measured at 530 nm and 657 nm using a UV spectrophotometer. Anthocyanin content (Q) was calculated using the formula: Q = (A530 − 0.25 × A657) × m−1, where 'm' is the fresh tissue weight in grams[18].
Subcellular localization
-
To construct the SmMYB44-l-GFP fusion vector, the stop codon of the SmMYB44-l gene was removed, and subcloning primers were designed (Supplementary Table S2). PCR-amplified fragments were analyzed by electrophoresis, and target bands were recovered from gels. The insert and the pCAMBIA1300-GFP vector were double-digested, gel-purified, and ligated using Solution I ligase. The resulting pCAMBIA1300-SmMYB44-l-GFP plasmid was transformed into competent E. coli, incubated at 37 °C, and positive clones were verified. The plasmid was then introduced into Agrobacterium tumefaciens strain LBA4404. Both the control (pCAMBIA1300-GFP) and recombinant Agrobacterium cultures were infiltrated into the abaxial surfaces of N. benthamiana leaves. After 36–48 h of dark incubation, the subcellular localization of the green fluorescent signal was observed and imaged under a fluorescence microscope.
Overexpression of the SmMYB44-l gene in tobacco
-
Tobacco explants were inoculated with Agrobacterium tumefaciens harboring the pCAMBIA2301G-SmMYB44-l, which came from subcloning primers (Supplementary Table S3) being designed with restriction sites Xba I and Sac I, construct to generate histocultured seedlings containing the resistance gene. Genomic DNA from regenerated tobacco plants was extracted using a rapid DNA release method. PCR amplification was conducted with recombinant plasmid as the positive control and wild-type (WT) DNA and double-distilled water (ddH2O) as negative controls. Amplification products were analyzed by agarose gel electrophoresis.
Total RNA was extracted from the leaves of both wild-type and transgenic tobacco plants overexpressing the SmMYB44-l gene. The RNA was reverse transcribed into cDNA, which served as the template for quantitative real-time PCR (qRT-PCR) to assess the expression of SmMYB44-l and two anthocyanin biosynthesis-related structural genes, NbCHS and NbF3H.
Overexpression of the SmMYB44-l gene in eggplant
-
Hypocotyls and cotyledons of 'March eggplant' were used as explants and infected with Agrobacterium carrying the pCAMBIA2301G-SmMYB44-l construct. Sterile seedlings were dissected to isolate cotyledons and hypocotyls, which were pre-cultured for 2 d. These explants were then infiltrated with Agrobacterium and co-cultivated for an additional 2 d. Following this, healing tissue culture was induced, leading to the formation of adventitious shoots. Once the adventitious shoots developed two leaves, rooting was initiated. Upon successful root formation, seedlings were acclimatized by opening culture bottles for 2–3 d before transplanting them to a soil substrate. Resistant seedlings were obtained via differentiation and rooting culture. Genomic DNA and total RNA were extracted from these transgenic lines, and cDNA was subsequently synthesized. PCR amplification and electrophoresis were performed to confirm transgene integration, while qRT-PCR was used to verify overexpression status. To evaluate the effect of SmMYB44-l overexpression on anthocyanin biosynthesis, the transcript levels of SmCHS and SmF3H were quantified by qRT-PCR in transgenic eggplant lines.
Transient expression of SmMYB44-l and SmMYB75 in tobacco leaves
-
Primers for SmMYB75 gene cloning (SmMYB75-F: ATGAATAATCCTCCTATAATGTG; SmMYB75-R: TTAATCAAGTAGATTCCACAAGTC) were designed using Oligo 7 software. PCR amplification was performed using eggplant cDNA as the template, and target gene identity was confirmed by sequence alignment. For transient expression, the pGBO vector was used, and subcloning primers were designed with restriction sites Sal I and BamH I (Supplementary Table S4). The cloned SmMYB44-l and SmMYB75 plasmids served as templates for subcloning. Standard molecular procedures—electrophoresis, gel purification, restriction digestion, ligation, transformation into competent E. coli, colony screening, sequencing, and bacterial stock preservation—were followed. The empty pGBO vector, as well as the recombinant constructs pGBO-SmMYB44-l and pGBO-SmMYB75, were transformed into Agrobacterium tumefaciens strain LBA4404. Equal volumes of individual and mixed bacterial suspensions (1:1 ratio) were aspirated with a 1 mL sterile syringe (needle removed) and used to infiltrate the abaxial surfaces of healthy Nicotiana benthamiana leaves. Infiltrated plants were kept in a humid, dark environment for 24 h, followed by a 5-d cultivation period under normal light conditions to observe pigmentation changes in the infiltrated leaves.
Protein interaction analysis—luciferase complementary imaging experiment (LCI)
-
For protein–protein interaction analysis, pCAMBIA1300-nLUC and pCAMBIA1300-cLUC vectors were employed[19]. Subcloning primers were designed based on the BamH I and Xho I restriction sites (Supplementary Table S5). PCR amplification products of the target genes were verified by electrophoresis. The amplicons and vectors were double-digested, and correctly sized fragments were purified from gels. The target gene inserts were ligated into the vectors, resulting in SmMYB75-pCAMBIA1300-nLUC and SmMYB44l-pCAMBIA1300-cLUC constructs, which were transformed into competent E. coli. After sequence validation of positive clones, plasmids were extracted and introduced into Agrobacterium tumefaciens strain GV3101. The resulting Agrobacterium liquid cultures carrying SmMYB75-pCAMBIA1300-nLUC and SmMYB44l-pCAMBIA1300-cLUC were co-infiltrated into the abaxial surfaces of tobacco leaves. After incubation in the dark for 24–36 h, infiltrated leaf sections were treated with the luciferase substrate D-luciferin potassium salt. Following a 7-min dark adaptation, luminescent signals indicating proteins interactions were visualized using an in-vivo imaging system.
Yeast one-hybrid assay
-
The linearized plasmids pAbAi-proSmCHS and pAbAi-proSmF3H were transfected into Y1HGold receptor yeast cells, plated on SD/Ura medium supplemented with varying concentrations of aureobasidin A (AbA), and incubated for 3–5 d. Positive clones were screened to determine the minimum AbA concentration required to inhibit the growth of the bait strain. Subsequently, the recombinant plasmid pGADT7-SmMYB44-l was introduced into Y1H (pAbAi-proSmCHS) and Y1H (pAbAi-proSmF3H) yeast strains. These were plated on SD/–Leu solid medium and incubated in an inverted position at 30 °C for 3–5 d to monitor yeast growth.
Luciferase reporter assay
-
The promoters of SmCHS and SmF3H were cloned into the pGreenII 0800-LUC vector, while SmMYB44-l was inserted into the pGreenII62-SK vector. Gene-specific primers containing restriction sites (Supplementary Table S6) were designed for amplification. The constructs were assembled via restriction digestion and ligation, transformed into E. coli, verified by PCR, and plasmids were subsequently extracted and introduced into Agrobacterium tumefaciens GV3101. Agrobacterial cultures harboring pGreenII62-SK, pGreenII62-SK-SmMYB44-l, pGreenII 0800-LUC-proSmCHS, and pGreenII 0800-LUC-proSmF3H were mixed at a 1:9 ratio. Equal volumes of the mixtures were infiltrated into the abaxial side of tobacco leaves using a sterile syringe. Leaves were maintained in the dark under moist conditions for 24–36 h. Subsequently, the leaves were excised, treated with diluted luciferin solution, and imaged after 7-min incubation in the dark to visualize the interaction.
-
Using eggplant cDNA as a template and primers SmMYB44-l-F/R, PCR amplification yielded the target bands (Supplementary Fig. S3), and sequencing revealed a full-length SmMYB44-l coding sequence (CDS) of 885 bp. Alignment with the open reading frame (ORF) confirmed that the gene lacks introns.
Bioinformatic analysis predicted that the SmMYB44-l protein consists of 294 amino acids, with a high relative abundance of Pro, Ser, Leu, and Ala. The calculated theoretical isoelectric point (pI) was 9.1, the molecular weight was 31.75 kDa, and the instability index was 70.93. The grand average of hydropathicity (GRAVY) score of −0.519 indicates that SmMYB44-l is a hydrophilic protein.
Sequence analysis revealed that SmMYB44-l contained an R2R3 domain, classifying it as a typical R2R3-MYB transcription factor. Comparative alignment with other Solanaceae species identified an EAR (LxLxL) repressor motif at its C-terminus. Secondary structure prediction using GOR software indicated that SmMYB44-l was composed primarily of random coils (78.23%), along with α-helices (12.59%), and β-sheets (9.18%) (Supplementary Fig. S4).
Phylogenetic analysis based on the amino acid sequences of MYB44 homologs from eggplant, potato, tomato, pepper, and Arabidopsis thaliana demonstrated that SmMYB44-l shared high homology with other Solanaceae members, showing the closest relationship to potato StMYB44-l (Supplementary Fig. S5).
Characterization of SmMYB44-l gene expression
-
All the peels and tissues of eggplants were sampled at 10 a.m. and then subjected to quantitative real-time PCR (qRT-PCR). Quantitative real-time PCR revealed significant variation in SmMYB44-l expression across eggplant pericarps of different colors (Fig. 1a). Expression was highest in white pericarp, followed by green and purple-red, and lowest in purple-black pericarp. Further expression profiling of SmMYB44-l across tissues—including root, stem, leaf, flower, pericarp, and pulp—of the cultivars 'Bailong' and 'NC7' (Fig. 1b) indicated elevated transcript levels in the leaves, stems, and pericarps of both cultivars, with expression levels in 'Bailong' exceeding those in 'NC7'. Notably, pericarp expression in 'Bailong' was nearly three-fold higher than in 'NC7', while expression was low in root and pulp tissues.
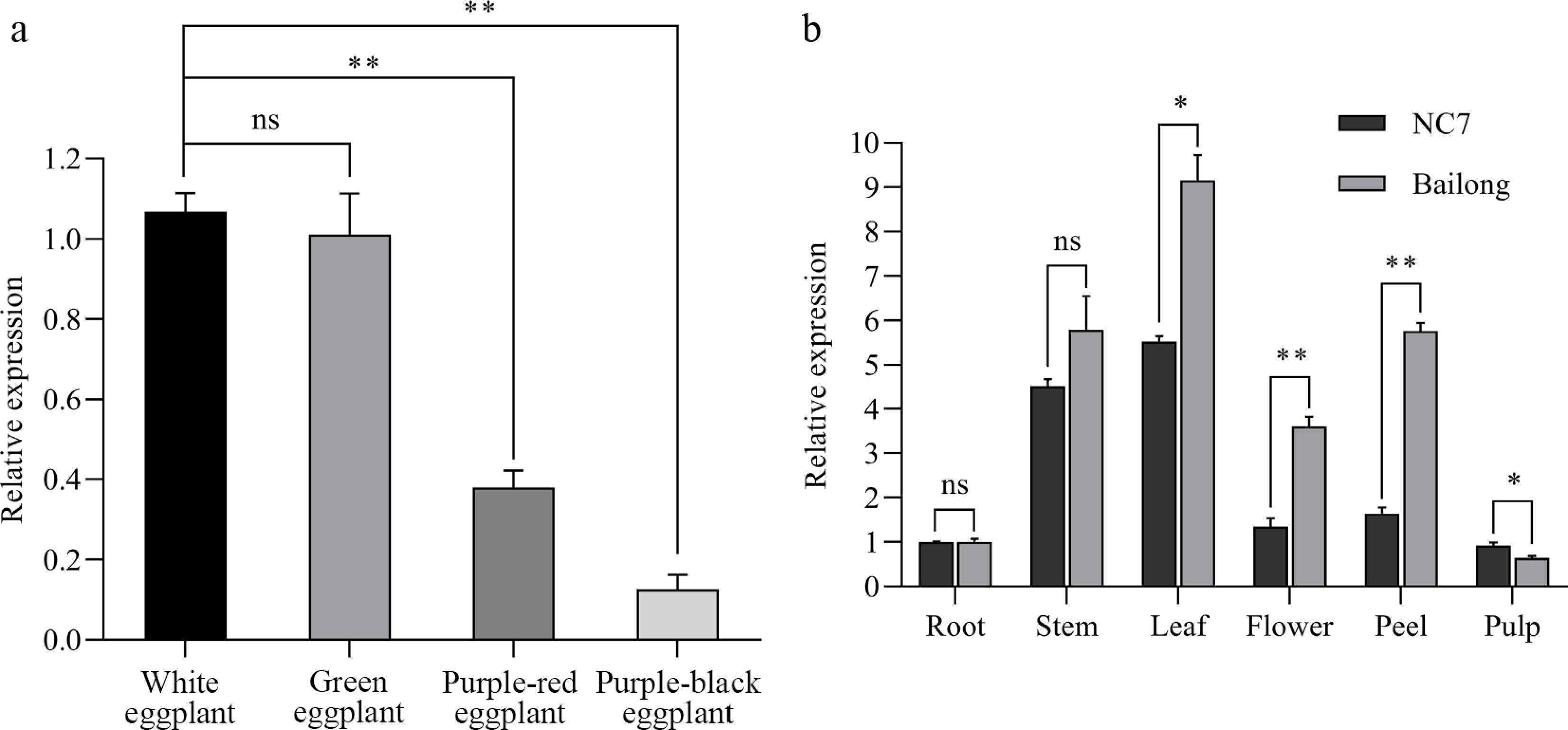
Figure 1.
Relative expression of SmMYB44-l in different colored peels and tissues (* p < 0.05, ** p < 0.01; ns: not significant). (a) Relative expression in different colored peels; (b) relative expression in different tissues.
Subcellular localization of SmMYB44-l protein
-
Transcription factors are a class of proteins that play a crucial role in the regulation of gene expression. They are usually located in the cell nucleus and regulate the transcriptional activity of genes by binding to specific DNA sequences. The AtMYB44 protein was localized in the nucleus[20], and so was the StMYB44 protein[21]. To clarify the subcellular localization of SmMYB44-l, we constructed the pCAMBIA1300-SmMYB44-l-GFP fusion construct in tobacco epidermal cells (Supplementary Fig. S6); the result showed that GFP fluorescence was in the nucleus, indicating that SmMYB44-l functioned within the nucleus.
Effects of genetic transformation of the SmMYB44-l gene on anthocyanin content and structural gene expression
Identification of transgenic tobacco plants
-
To explore the biological function of MYB44-l, we first heterologously expressed SmMYB44-l in tobacco and obtained transgenic plants. PCR amplification using DNA from herbicide-resistant transgenic tobacco leaves confirmed the presence of SmMYB44-l, as target bands were successfully amplified (Supplementary Fig. S7), indicating the successful generation of transgenic lines. To evaluate SmMYB44-l expression in transgenic tobacco, qRT-PCR was performed on four randomly selected lines. As shown in Supplementary Fig. S8, SmMYB44-l transcript levels in SmMYB44-l-OE-8, OE-16, and OE-19 were significantly higher than in the wild type (WT), confirming successful overexpression.
Effect of overexpression of tobacco on anthocyanin content and expression of structural genes
-
Phenotypic analysis under identical growth conditions revealed that anthocyanin levels were significantly reduced in SmMYB44-l-overexpressing plants compared to WT (Fig. 2a). Correspondingly, qRT-PCR showed markedly decreased expression of structural genes NbCHS, and NbF3H (Fig. 2b), suggesting that SmMYB44-l functions as a negative regulator of anthocyanin biosynthesis.
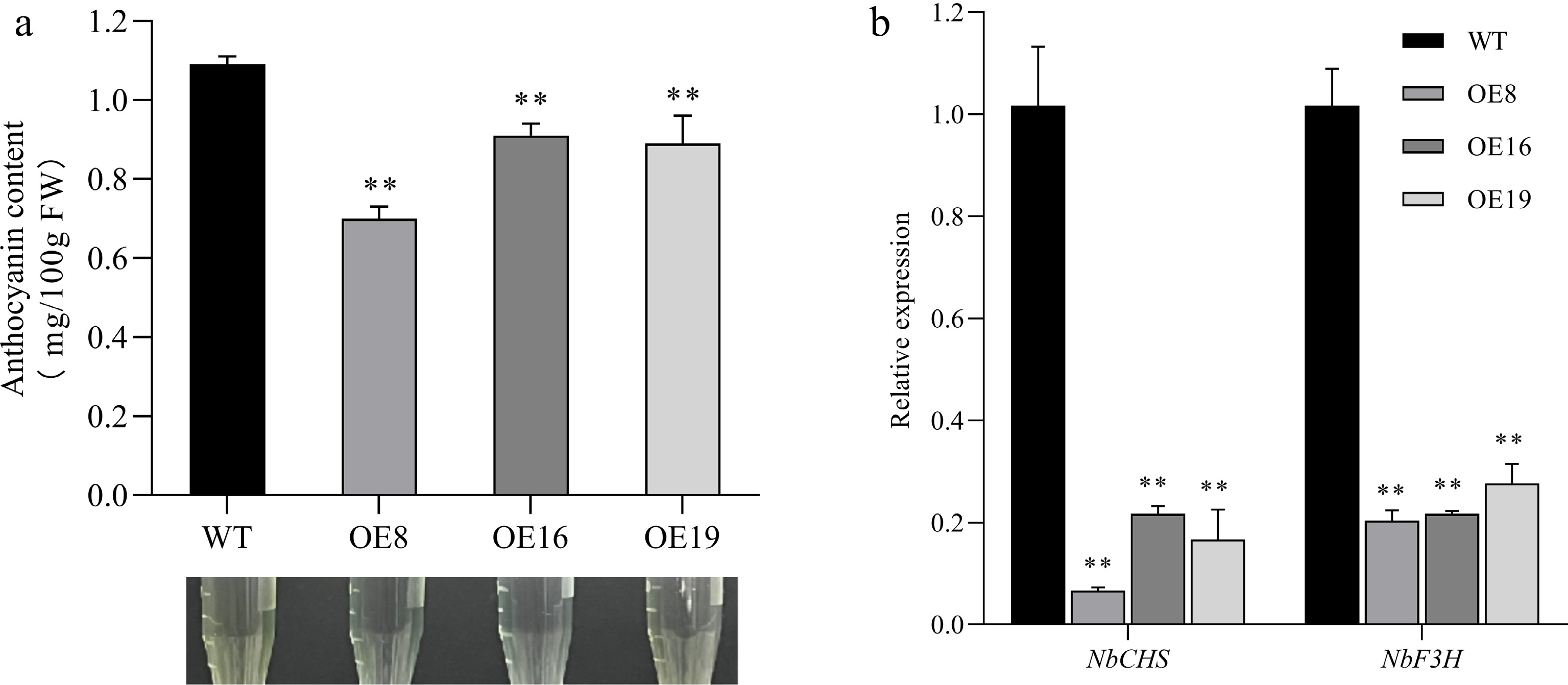
Figure 2.
Effect of SmMYB44-l overexpression on anthocyanin synthesis in tobacco leaves (** p < 0.01). (a) Total anthocyanin content in tobacco leaves with overexpressed SmMYB44-l; (b) expression levels of relevant structural genes in overexpressed SmMYB44-l tobacco leaves.
Identification of transgenic eggplant plants
-
To further clarify the functional role of SmMYB44-l in eggplant, we constructed and obtained the SmMYB44-l overexpression plants. Genetic transformation of cotyledons and hypocotyls in eggplant was performed using Agrobacterium-mediated transformation (Supplementary Fig. S9). It was observed that hypocotyl explants exhibited a higher efficiency of adventitious shoot differentiation compared to cotyledon explants, and that the survival rate of shoots derived from cotyledons was lower. Ultimately, three T0 transgenic eggplant lines resistant to kanamycin and harboring the SmMYB44-l gene were obtained. PCR amplification of genomic DNA from resistant plants confirmed the presence of the transgene, as target bands were successfully amplified (Supplementary Fig. S10), indicating preliminary validation of successful transformation. The phenotypic comparison of flowers, leaves, and fruits revealed that compared to the WT, the colors of petals, leaves, and leaf veins of T1 generation overexpressed eggplant (OE-8) became paler, and the color of the fruit skin lost most of its purple hue (Supplementary Fig. S11).
To further confirm the transgenic nature of these plants, the relative expression levels of SmMYB44-l were analyzed by qRT-PCR. The expression of SmMYB44-l in transgenic lines SmMYB44-l-OE-2 and SmMYB44-l-OE-3 was significantly higher than in the WT plants (Supplementary Fig. S12).
Effect of overexpression of eggplant on anthocyanin content and expression of structural genes
-
To evaluate the impact of SmMYB44-l overexpression on anthocyanin biosynthesis, anthocyanin content was quantified in transgenic lines SmMYB44-l-OE-2 and SmMYB44-l-OE-3, along with WT controls grown under identical conditions. Anthocyanin levels were significantly reduced in both transgenic lines compared to the WT (Fig. 3a). Furthermore, fluorescence-based quantification revealed a marked downregulation of the structural genes SmCHS and SmF3H in transgenic plants relative to the WT (Fig. 3b).
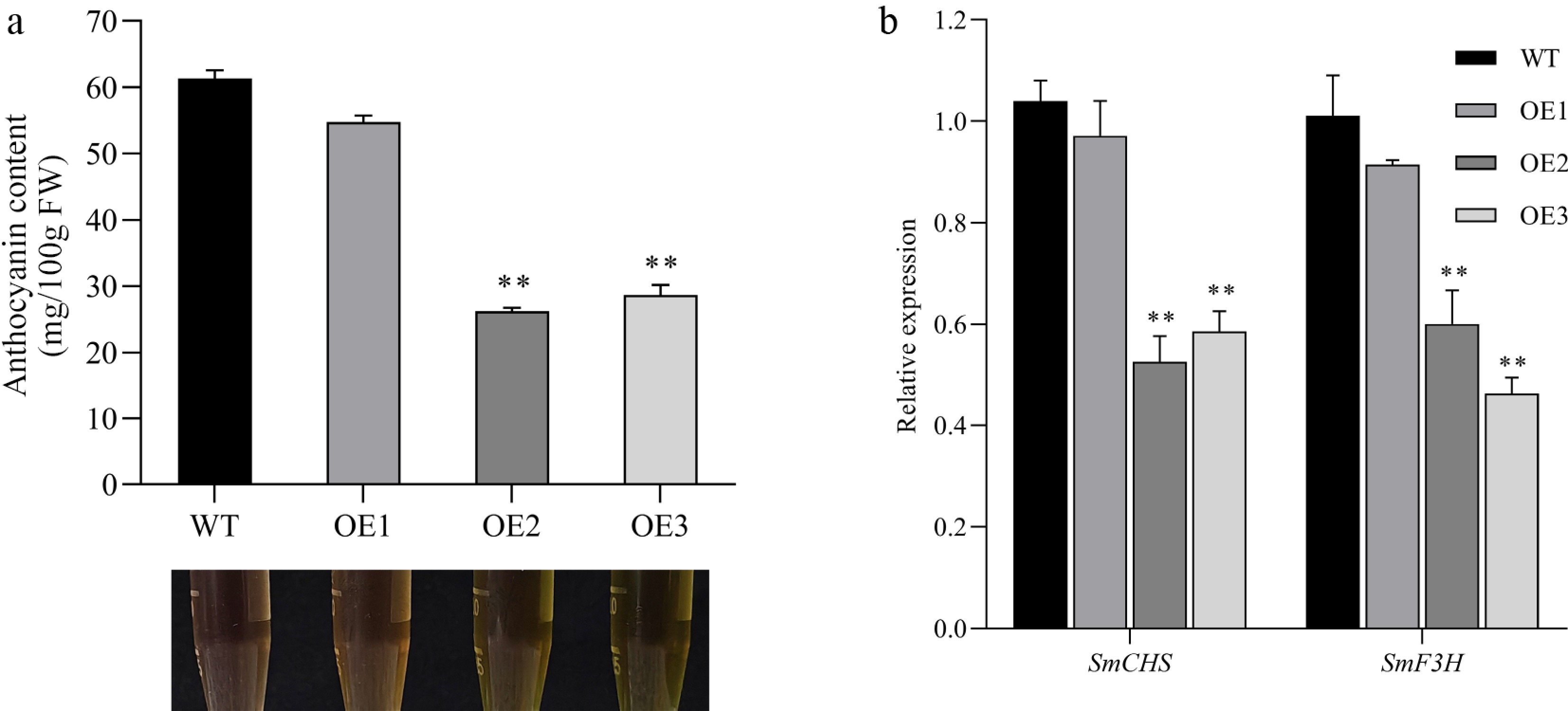
Figure 3.
Effect of SmMYB44-l overexpression on anthocyanin synthesis in transgenic eggplant leaves. (a) Total anthocyanin content in eggplant leaves with overexpressed SmMYB44-l; (b) expression levels of relevant structural genes in overexpressed SmMYB44-l eggplant leaves (** p < 0.01).
Analysis of transient expression of SmMYB44-l and SmMYB75
-
To verify the relationship of SmMYB44-l and SmMYB75 with anthocyanin biosynthesis, we carried out transient expression assays in tobacco leaves. The results demonstrated that expression of SmMYB44-l alone did not cause visible pigmentation compared to the empty vector control. In contrast, SmMYB75 alone induced pronounced pigmentation, and co-expression with SmMYB44-l resulted in visibly lighter pigmentation (Fig. 4a).
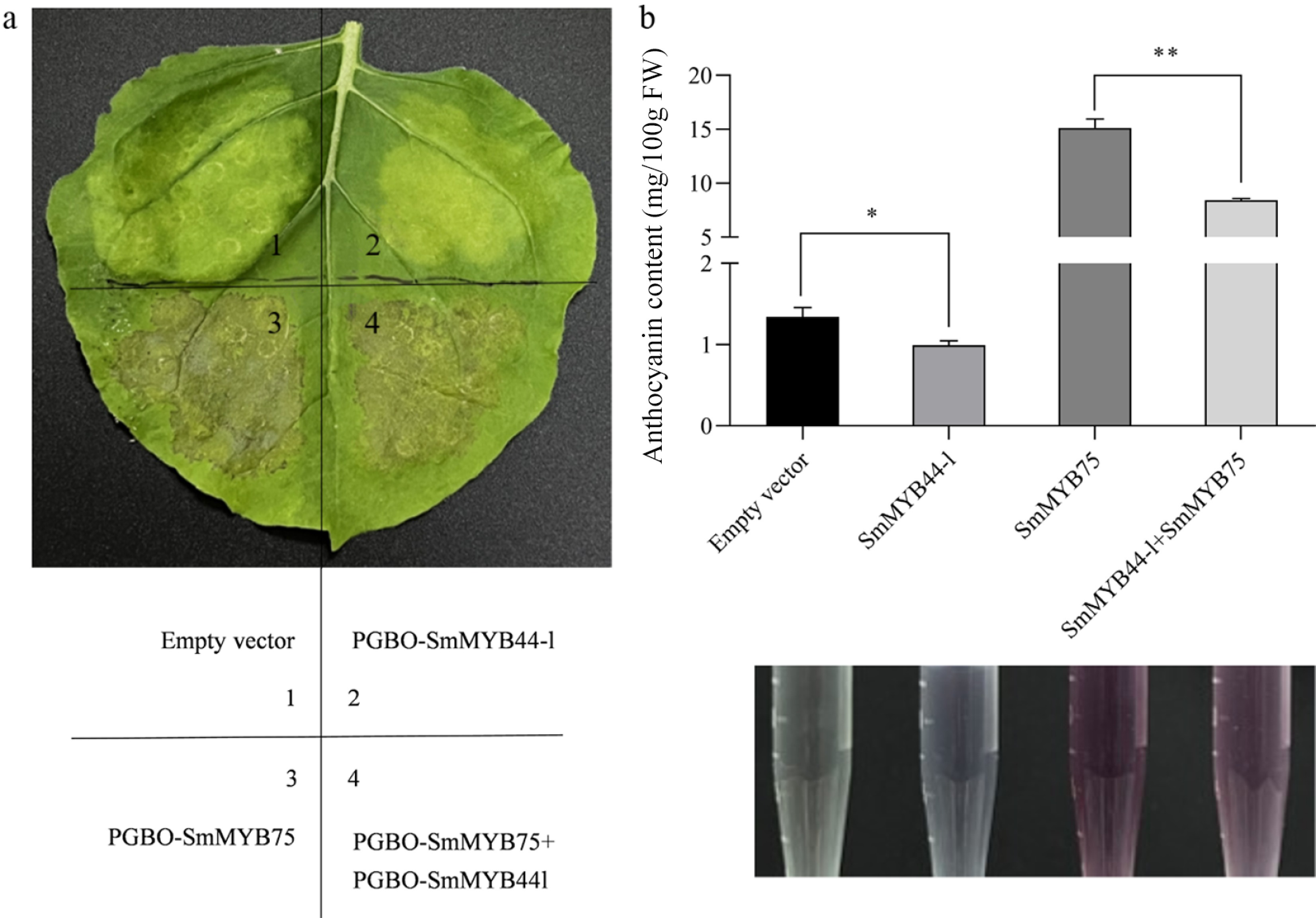
Figure 4.
Expression analysis of SmMYB44-l and SmMYB75 in Nicotiana benthamiana leaves (* p < 0.05, ** p < 0.01). (a) Transient overexpression of SmMYB44-l and SmMYB75 in Nicotiana benthamiana leaves. Numbers in the figure indicate infected genes; (b) total anthocyanin contents in Nicotiana benthamiana leaves agroinfiltrated with different gene constructs.
Anthocyanin content analysis at the infiltration sites corroborated these observations: SmMYB44-l alone reduced anthocyanin levels from 1.3 mg/100 g (control) to 1.0 mg/100 g. Co-infiltration of SmMYB44-l with SmMYB75 led to a highly significant reduction in anthocyanin content compared to SmMYB75 alone (Fig. 4b).
qRT-PCR analysis further revealed that SmMYB44-l expression was elevated both when expressed alone, and when co-expressed with SmMYB75, relative to the control (Fig. 5). Interestingly, SmMYB75 expression was significantly repressed by SmMYB44-l alone, but was upregulated when expressed alone or co-expressed with SmMYB44-l, though co-expression resulted in lower expression than SmMYB75 alone. These results suggest that SmMYB44-l negatively regulates SmMYB75. Analysis of structural gene expression showed that SmMYB44-l overexpression reduced the expression of NbCHS and NbF3H compared to the control. While SmMYB75 alone significantly upregulated both genes, co-expression with SmMYB44-l suppressed this induction. This indicates that SmMYB44-l may repress the expression of key structural genes in the anthocyanin biosynthetic pathway.
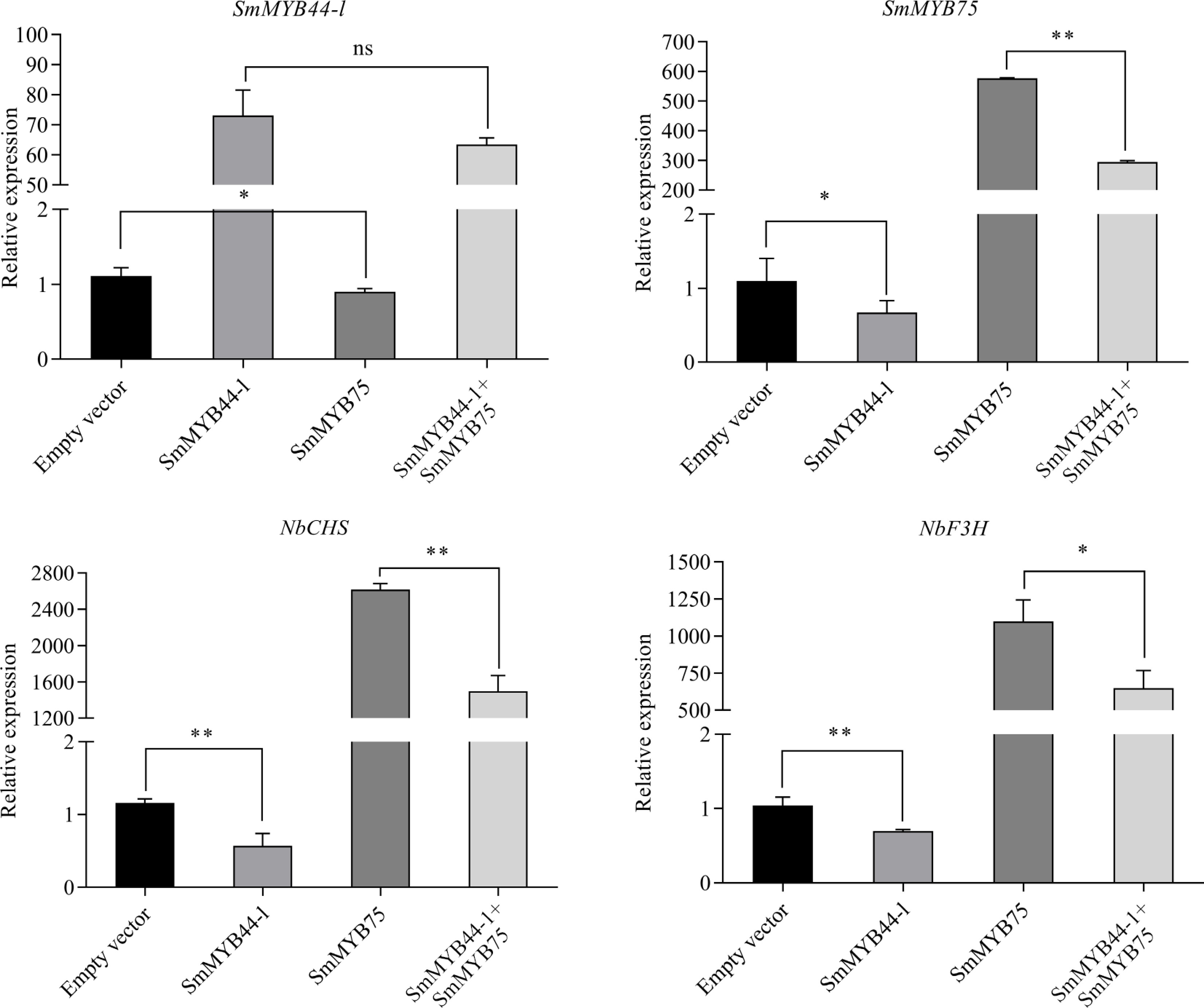
Figure 5.
Expression levels of the MYB gene (SmMYB44-l and SmMYB75) and structural genes (NbCHS and NbF3H) in Nicotiana benthamiana leaves (* p < 0.05, ** p < 0.01).
Analysis of protein interactions between SmMYB44-l and SmMYB75
-
The above results indicate that the expression of MYB44 is influenced by MYB75. Then, is there a protein-protein interaction regulatory relationship between MYB4 and MYB75? To investigate potential protein–protein interactions, bacterial suspensions were co-infiltrated into tobacco leaves with nluc+cluc (negative control), SmMYB75-nluc+cluc, nluc+SmMYB44-l-cluc, and SmMYB75-nluc+SmMYB44-l-cluc (experimental group). No fluorescence was observed in either the experimental or control groups, indicating that SmMYB44-l does not directly interact with SmMYB75 in tobacco (Supplementary Fig. S13).
Analysis of promoter interactions between eggplant SmMYB44-l and the anthocyanin structural genes SmCHS and SmF3H
-
The previous results indicated that CHS and F3H were expressed in MYB44-overexpressing plants (Figs 2, 3). To verify whether MYB44S directly targets the promoters of CHS and F3H for expression regulation, we conducted yeast single-hybrid and dual-luciferase reporter experiments.
Yeast one-hybrid
-
Yeast one-hybrid (Y1H) assays were conducted to assess the interaction between SmMYB44-l and the promoters of SmCHS and SmF3H (Fig. 6). The positive control (PC) was Y1H (p53-AbAi), and the negative control (NC) included Y1HGold strains with pGADT7+proSmCHS and pGADT7+proSmF3H. On selective medium containing 500 ng/mL AbA, yeast strains harboring pGADT7-SmMYB44-l and either proSmCHS or proSmF3H grew robustly, forming white colonies. This suggests that SmMYB44-l can bind to the promoters of SmCHS and SmF3H.
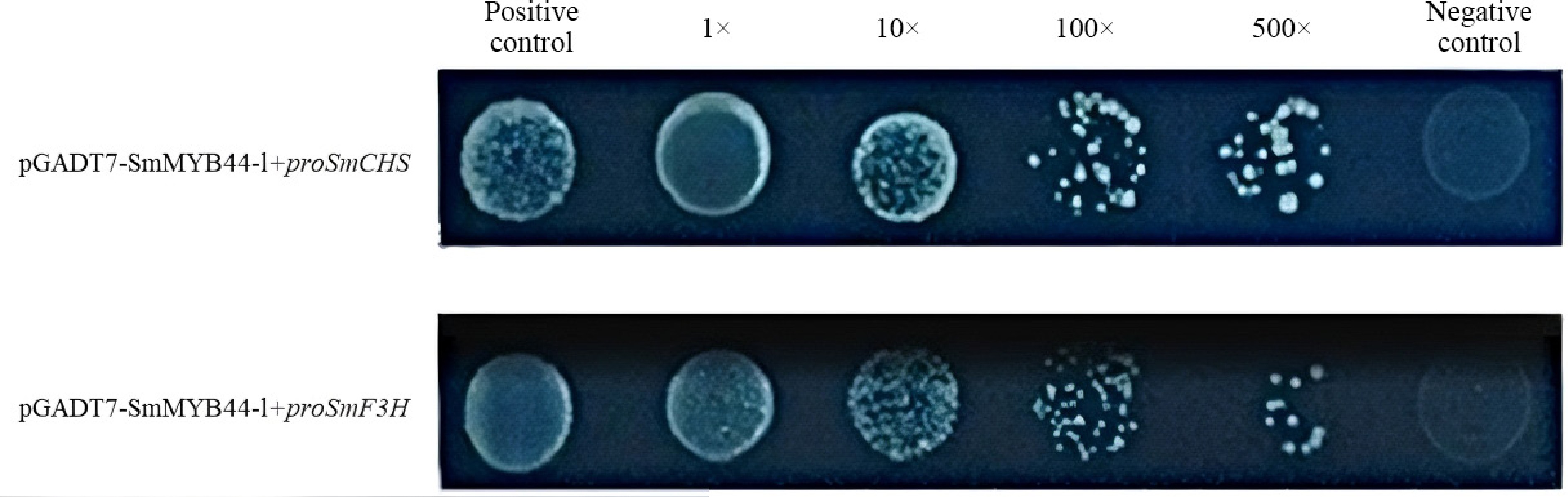
Figure 6.
Interaction results of SmMYB44-l with the proSmCHS and proSmF3H.
Luciferase reporter assay
-
Luciferase reporter assays further supported this finding. Co-expression of SmMYB44-l with promoter–reporter constructs resulted in a shift from red to blue fluorescence compared with the empty vector control, indicating reduced promoter activity. These results suggest that SmMYB44-l interacts with the SmCHS and SmF3H promoters and suppresses their transcriptional activity (Fig. 7).
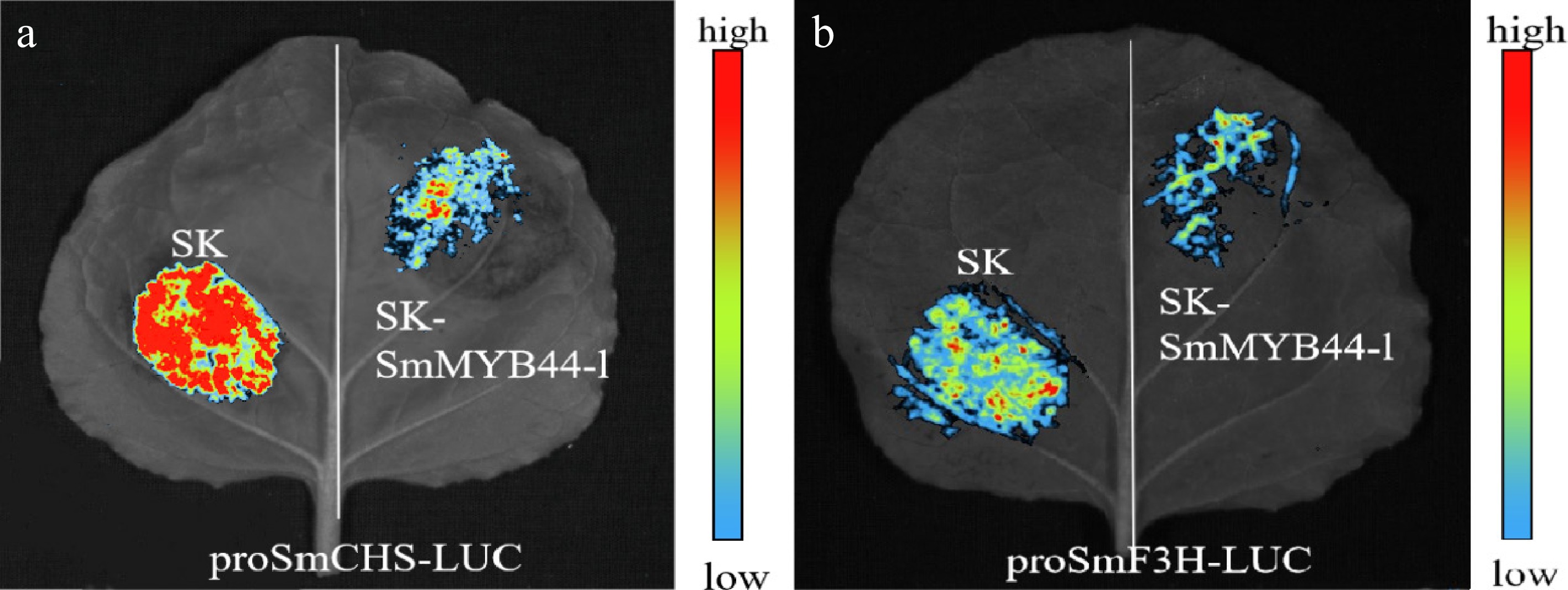
Figure 7.
DNA–protein interactions were verified by LUC. (a) proSmCHS activity with SmMYB44-l; (b) proSmF3H activity with SmMYB44-l.
-
The pericarp of eggplant exhibits a diverse range of colors, including purple-black, purple-red, green, white, and pink. This coloration is primarily determined by the relative concentrations of two key pigments: anthocyanins and chlorophyll. Our study revealed that the anthocyanin concentration in the peel of purple eggplant was significantly higher than in both white and green eggplant varieties.
Regulatory genes encode transcription factors that control the expression of structural genes, either activating or repressing their function. Anthocyanin biosynthesis is tightly regulated by a network of transcription factors, among which MYB proteins play a central role. With advances in plant molecular biology, transient gene expression systems have become widely adopted for functional gene analysis[22]. In tobacco leaves, transient expression experiments demonstrated that the potato MYB activators StAN1-R0, StAN1-R1, and StAN1-R3 significantly promoted anthocyanin biosynthesis when infiltrated individually. However, co-expression with StMYB44s resulted in suppressed anthocyanin accumulation, suggesting that StMYB44s act as negative regulators of anthocyanin synthesis[13]. Similar repressive effects have been observed in other species, including Narcissus[23], sweet potato[24], and Chinese cabbage[25], where the expression of key structural genes for anthocyanin biosynthesis was significantly downregulated. These findings suggest that BcMYB44 and its homologs may function as transcriptional repressors by inhibiting the expression of structural genes in the anthocyanin biosynthetic pathway. In our study, we used the MYB transcriptional activator SmMYB75 as a positive control and found that infiltration of PGBO-SmMYB44-l alone into tobacco leaves caused no visible pigmentation change. However, both anthocyanin content and expression levels of key structural genes (NbCHS and NbF3H) were markedly reduced. In contrast, infiltration with PGBO-SmMYB75 alone enhanced anthocyanin accumulation. When PGBO-SmMYB44-l was co-infiltrated with PGBO-SmMYB75, anthocyanin biosynthesis was suppressed, and the expression of structural genes was significantly downregulated. These results indicate that SmMYB44-l can inhibit anthocyanin accumulation. Furthermore, transgenic tobacco and eggplant plants overexpressing SmMYB44-l also exhibited reduced expression of structural genes, further confirming its role as a negative regulator of anthocyanin biosynthesis.
MYB transcription factors can function either as activators, by upregulating downstream structural genes, or as repressors, by directly inhibiting gene transcription. R2R3-MYB repressors are known to bind the promoters of structural genes and suppress their activity[26]. For instance, SmMYB86 has been shown to bind and repress the promoters of SmCHS, SmF3H, and SmANS[17], while NtMYB3 in Narcissus inhibits the promoter activity of NtFL[23], and MdMYB6 represses MdANS and MdGSTF12 in apple[27]. In the current study, yeast one-hybrid and dual-luciferase reporter assays reveal that SmMYB44-l directly binds to the promoters of SmCHS and SmF3H, and inhibits their transcriptional activity. These findings strongly support that SmMYB44-l functions as a transcriptional repressor by directly targeting structural gene promoters to inhibit anthocyanin biosynthesis and accumulation. However, as promoter regions often contain multiple cis-regulatory elements, the specific DNA motifs recognized by SmMYB44-l and the nature of its interaction with them remain to be elucidated through further investigation.
Additionally, some R2R3-MYB repressors inhibit anthocyanin biosynthesis indirectly by competing with activator MYBs for binding to bHLH proteins within the MYB–bHLH–WD40 (MBW) transcriptional complex. This competition weakens the transcriptional activation of target genes. Such repressive mechanisms have been documented for VvMYB4b in grape[28], MdMYB15 in apple[29], and StMYB3 in potato[30]. However, based on amino acid sequence alignment, some R2R3-MYB repressors lack the conserved bHLH-interaction motif (D/E)Lx2(R/K)x3Lx6Lx3R and thus are unable to physically interact with bHLH proteins. In our analysis, SmMYB44-l was found to lack this motif, suggesting that it likely does not interact with bHLH partners. Nevertheless, this hypothesis requires experimental validation to clarify the precise mechanism of repression employed by SmMYB44-l.
MYB proteins contain a conserved MYB DNA-binding domain that recognizes specific cis-regulatory elements and facilitates protein–protein interactions, playing a pivotal role in regulating plant growth and development[31]. Bimolecular fluorescence complementation (BiFC) analysis has shown that MYB5 and MYB14 interact synergistically to promote proanthocyanidin (PA) accumulation in Medicago truncatula[32]. In plants, heterodimerization among R2R3-MYB transcription factors is well-documented[33]; for example, the heterodimer formed by MdMYB306-like and MdMYB17 in apple suppresses anthocyanin biosynthesis[34]. In the present study, no interaction was detected between SmMYB44-l (a transcriptional repressor) and SmMYB75 (an activator) using luciferase complementation imaging (LCI) assays. While the SmMYB44-like protein can bind to the specific cis-elements within the SmMYB75 promoter, the electrophoretic mobility shift assay (EMSA) can be utilized to find the mechanism by which it regulates the transcription of SmMYB75. Compared to the extensive research on MYB protein complex formation, studies on MYB–MYB interactions remain limited. However, continued exploration of these interactions will provide deeper insights into the regulatory networks involving MYB transcription factors in plant pigmentation.
-
The authors confirm their contributions to the paper as follows: Wang Z, Yang H, Tao T and Yang Y designed the research; Yang H, Li H, and Tao T performed the molecular biology experiments; Yang H, Wei D, and Tang Q carried out the bioinformatics analysis; Li H, Yang H, and Wang Z analyzed the data and wrote the manuscript. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
This work was supported by the earmarked fund for the China Agriculture Research System (CARS-23-A08), the Chongqing Natural Science Foundation Project (CSTB2024NSCQ-MSX1020), and the Major Core Technology Research of Chongqing Academy of Agricultural Sciences Supported by Municipal Finance Project (cqaas2023sjczhx002).
-
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
-
accompanies this paper online at: https://doi.org/10.48130/vegres-0026-0014.
-
# Authors contributed equally: Hangchun Li, Huiqin Yang, Tao Tao
- Supplementary Table S1 Primers sequences of qRT-PCR.
- Supplementary Table S2 Primers for pCAMBIA1300-SmMYB44-l-GFP.
- Supplementary Table S3 Subcloning primers of pCAMBIA-2301G-SmMYB44-l.
- Supplementary Table S4 Primers used for the subcloning of SmMYB44-l and SmMYB75.
- Supplementary Table S5 Subcloning primers of pCAMBIAI1300-nLUC and pCAMBIAI1300-cLUC vector.
- Supplementary Table S6 Subcloning primers for construcing pGreenII 0800-LUC and pGreenII62-SK recombinant plasmids.
- Supplementary Fig. S1 The four eggplant materials with different colors.
- Supplementary Fig. S2 The eggplant materials of Bailong and NC7.
- Supplementary Fig. S3 Electropherograms of PCR amplification of the SmMYB44-l gene.
- Supplementary Fig. S4 Prediction of the secondary structure.
- Supplementary Fig. S5 Phylogenetic tree analysis.
- Supplementary Fig. S6 Subcellular localization of the SmMYB44-l protein.
- Supplementary Fig. S7 Obtaining genetically modified tobacco and PCR identification of some T0 generation transgenic tobacco.
- Supplementary Fig. S8 SmMYB44-l gene expression in tobacco (**p < 0.01).
- Supplementary Fig. S9 Obtaining genetically modified eggplant.
- Supplementary Fig. S10 PCR identification of T0 generation of transgenic eggplant.
- Supplementary Fig. S11 Phenotypic comparisons of flowers, leaves and fruits of T1 generation of transgenic eggplant.
- Supplementary Fig. S12 SmMYB44-l gene expression in transgenic eggplants.
- Supplementary Fig. S13 Identification of protein-protein interactions by luciferase complementary imaging technology.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li H, Yang H, Tao T, Wei D, Tang Q, et al. 2026. Regulatory role of the SmMYB44-l gene in anthocyanin synthesis in Solanum melongena L.. Vegetable Research 6: e020 doi: 10.48130/vegres-0026-0014 

Regulatory role of the SmMYB44-l gene in anthocyanin synthesis in Solanum melongena L.
- Received: 22 July 2025
- Revised: 24 March 2026
- Accepted: 01 April 2026
- Published online: 08 June 2026
Abstract: Eggplant (Solanum melongena L.), a major crop of the Solanaceae family, is valued for its rich nutritional profile, particularly the high anthocyanin content in the purple peel. Anthocyanin biosynthesis is primarily regulated by a network of transcription factors. Current research on MYB transcription factors in eggplant has predominantly focused on their roles as positive regulators, with limited insight into negative regulatory mechanisms. In this study, we cloned the SmMYB44-l gene. Bioinformatic and phylogenetic analyses revealed the presence of an EAR (LxLxL) repressor motif at the C-terminus of its encoded protein, and high sequence homology to StMYB44-l from potato. Expression analysis indicated a negative correlation between SmMYB44-l transcript levels and pericarp anthocyanin content. Subcellular localization experiment showed that SmMYB44-l localized to the nucleus. Transient expression assays demonstrated that SmMYB44-l repressed anthocyanin biosynthesis in tobacco (Nicotiana benthamiana) by downregulating both the MYB activator SmMYB75, and structural genes (NbCHS, NbF3H). Luciferase complementation imaging confirmed the absence of physical interaction between SmMYB44-l and SmMYB75. Overexpression of SmMYB44-l in both tobacco and eggplant suppressed anthocyanin accumulation. Yeast one-hybrid and luciferase reporter assays validated its direct repressive role in the regulation of anthocyanin biosynthesis.
-
Key words:
- Eggplant /
- SmMYB44-l /
- Inhibition /
- Anthocyanin