








-
Grafting is an ancient plant propagation technique that combines the aerial portion (scion) with another segment (rootstock) to form a new plant[1,2]. Modern fruit cultivation systems are mainly based on scion varieties, grafted with rootstock to control plant-specific characteristics, such as fruit size, early fruiting, and to reduce disease susceptibility[3,4]. Rootstocks provide grafted plants with more favorable tolerance against multiple environmental stresses[5]. The selection of a suitable rootstock is one of the major decisions in establishing a tree garden and achieving excellent tree performance in different ecological regions.
Rootstocks have been used in fruit plants to improve nutrient absorption and transportation by increasing fruit production, taste and quality, and tolerance against several environmental stresses[6,7]. Dwarfing rootstock is an important germplasm resource for high-density cultivations to ensure maximum yield and mechanical harvesting[8]. In recent years, dwarfing rootstocks have become popular due to reduced vegetative growth of scion varieties, increased production, and reduced labour costs[9]. It is speculated that scion and rootstock are the essential components in fruit production that interact with each other to release mineral nutrients, hormones and carbohydrates[10]. The hypothesis behind rootstock-induced size control may be triggered by several factors such as water, nutrients, and particularly hormones, which pass through the graft union to influence scion growth[11].
Several studies have reported the diversity and domestication of rootstocks and their influence on plant vigour, hormonal communication between scion and rootstock, and various stresses[1,12−16]. However, due to the complex interaction of rootstock-induced scion vigour, these studies have not fully elucidated the possible dwarfing mechanism. This review summarizes the existing information of rootstock influence on scion growth, nutrient uptake, hormonal communication, carbohydrate distribution, and the molecular mechanism.
-
Rootstocks have a substantial effect on the physiological features of grafted plants and other aspects of growth and development, among which the reduction of scion growth is one of the most interesting phenomena (Fig. 1). Previous studies have demonstrated that dwarfing rootstock and interstock contribute to restricting plant height, crown size, and tree volume. However, the plants grafted on vigourous rootstocks have more nutritional properties but lower yield[17−20]. Gjamovski and Kiprijanovski[21] documented that 'Granny Smith' apple trees, grafted with taller rootstock (Supporter 4), have higher vegetative growth than size-controlling rootstocks (Panjam 1, M.9 T337 and Mark 9). Zhou et al.[22] stated that 'Red Fuji' apple scion grafted on size-controlling rootstock M.9 had the lowest vegetative growth, canopy width, and trunk cross-sectional area (TCSAs), compared with vigorous rootstocks.
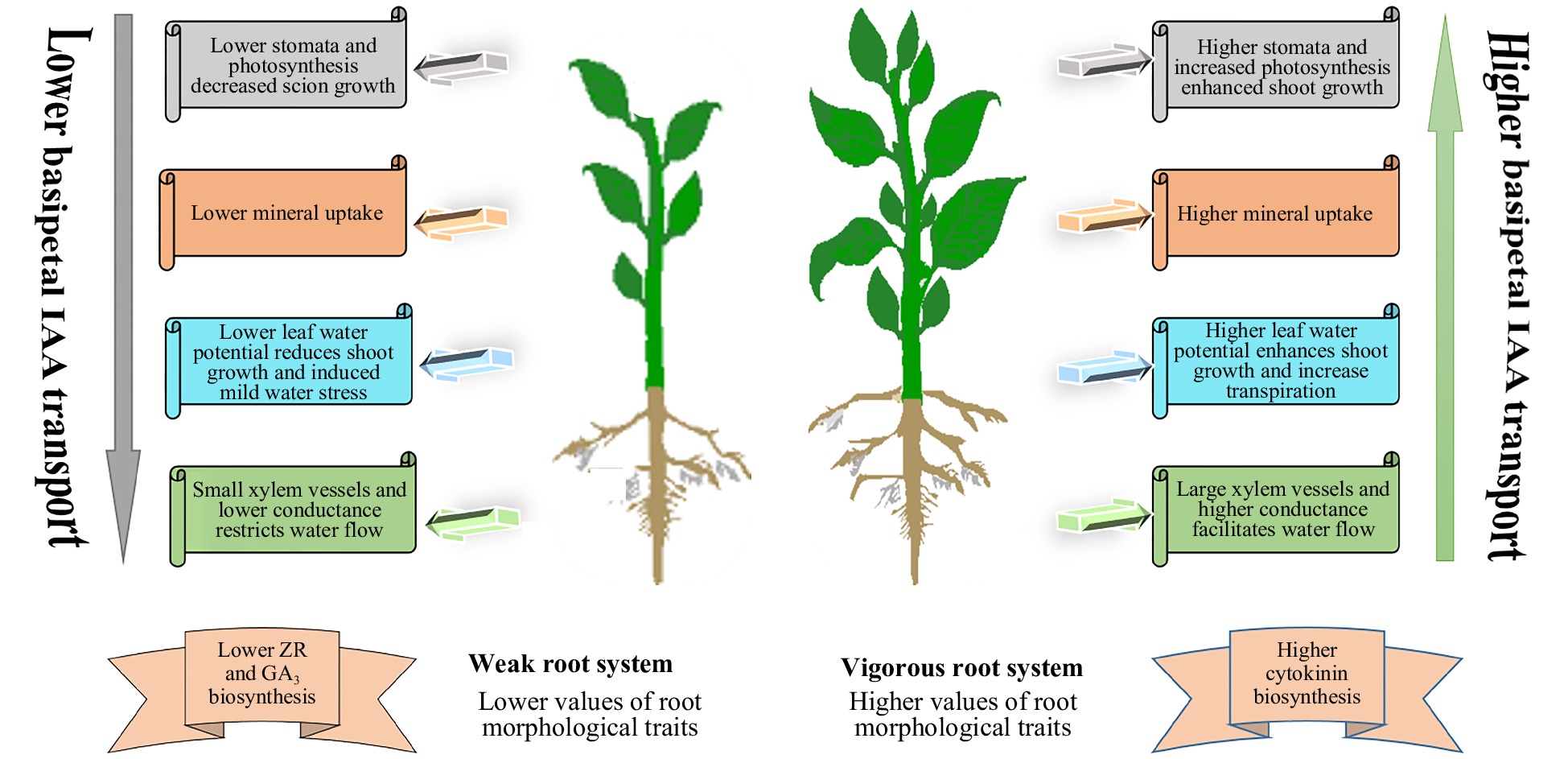
Figure 1. Schematic diagram of rootstock control scion vigour.
Sitarek and Bartosiewicz[23] studied that trees grafted on M46 and P. divaricata had substantially higher values of trunk cross-sectional area. In comparison, those grafted on Wangenheim Prune showed lower values and dwarfing characteristics. Several studies have shown that apple scions grafted onto dwarfing rootstocks have shorter shoot lengths, lower trunk cross-sectional area (TCSA), and weak growth characteristics[21,24,25]. Tworkoski and Fazio[23] also studied the effect of various apple scion cultivars grafted onto rootstocks of Geneva and Malling series. They observed that plants grafted with G.41 and G.935 rootstocks had the maximum shoot growth and stem diameter compared with other rootstocks.
Moreover, the application of interstocks with greater dwarfing effects had lower net photosynthesis rate and root-shoot ratio but improved yield efficiency and fruit quality of Red Fuji apple trees[26]. In another study, Hayat et al.[27] reported that apple scion grown onto dwarfing M.9 rootstock had the smallest plant height and reduced scion diameter, and the lowest hydraulic conductance (Kleaf). In contrast, plants grown onto more vigorous rootstocks M.26, Chistock-1, and Baleng rootstocks had longer shoot length, greater scion diameter, and hydraulic conductance.
-
Root hydraulic conductivity can influence shoot growth by modifying water supply to the aerial part; however, its role in rootstock-induced vigour is still controversial[28]. The physiological mechanisms of the size reduction of rootstock or interstock are triggered by restricting water flow to the scion[29]. In another study, Basile et al.[30] reported that water status might significantly affect the rootstock-induced dwarfing effects in peach trees. They studied the influence of different size-controlling rootstocks on the early maturing cultivar 'Flavorcrest' and found significant variations in stem water potential (ΨSTEM) and stem extension growth. Solari et al.[31] and Tombesi et al.[32] explained the relationship between dwarfing rootstocks and hydraulic conductance. They found that dwarfing rootstocks limit the water supply to the aerial portion.
Lowered hydraulic conductivity of size-controlling rootstocks is probably due to the greater hydraulic resistance and limited water absorption capacity of size-controlling rootstocks. In pear, Chen et al.[33] studied the anatomical structure of leaves and stem of vigorous and dwarf type pears to clarify the mechanism of a short tree structure. They found that the dwarf type had a smaller vessel diameter and a greater stem cross-sectional area than the vigorous type. In citrus, Saeed et al.[34] explored the relationships among growth vigour and anatomical characteristics of leaves, stems and roots. They found that vigorous rootstocks, rough lemon (C. jambhiri), have larger xylem vessel elements in the stems and roots with lower phloem percentage (%), when compared with dwarfing rootstocks- flying dragon (P. trifoliata). Zoric et al.[35] noticed that dwarfing rootstocks had lower theoretical hydraulic conductance (kh) and a smaller vessel lumen size, especially when compared with vigorous rootstocks. Martínez-Alcántara et al.[36] also demonstrated lower leaf water potential and decreased hydraulic conductance during times of high evaporative demand, leading to reduced stomatal conductance in plants grafted onto size-controlling rootstocks. In apple, Zhou et al.[37] studied the effect of different vigour controlling rootstock combinations (vigorous, semi-dwarfing and dwarfing characteristics) grafted with 'Red Fuji' scion cultivar. They concluded that reduction in scion growth of dwarfing rootstocks (M.9 and B.9) was related to lower palisade/spongy parenchyma ratios and hydraulic conductance stomatal density compared with more vigorous rootstocks. Therefore, it is concluded from the above literature (Table 1) that the lower hydraulic conductance/ limited supply of water to the aerial part resulted in the closure of stomata and decreased CO2 assimilation rate, possibly limited scion growth in most dwarfing rootstocks.
Table 1. Important physiological and anatomical evidences underlying rootstock induced vigour control.
Name of crop Rootstock/treatments Traits Key findings References Peach K146-43, Hiawatha, Nemaguard Vegetative growth, stem elongation, and water status measurements Water relations were linked with rootstock-induced dwarfing mechanism. [25] Tree canopies were partially covered (0, ~30, and ~60%), and three rootstocks were used, including (K146-43, Hiawatha, and Nemaguard) Water usage and transport Tree water status were possibly involved in the dwarfing of scion growth by rootstocks. [30] ‘K146-43’, ‘Nemaguard’ and ‘P30-135’ Theoretical axial xylem conductance Larger vessel dimensions were found in the vigourous rootstock than dwarfing rootstocks (‘K146-43’). The vessel density per xylem area in ‘Nemaguard’ rootstock was also lower than those of the ‘K146-43’ and ‘P30-135’ rootstocks. [31] Apple JM5, JM1, JM7, JM2, and Marubakai Anatomical traits, sap flow, and hydraulic conductance The degree of dwarfing were closely associated with the phloem to xylem ratios of roots. Therefore, it could be a useful index for screening the dwarfing potential of rootstocks. [32] Baleng, Chistock-1, SH.6/Baleng, SH.6/Chistock-1, M.26, M.9, SH.6, B.9 Hydraulic conductance, non-structural carbohydrates, leaf anatomy, gas exchange, and chlorophyll fluorescence, The reduction of photosynthetic rate has been attributed to the decreased hydraulic conductivity, which causes a decline in instantaneous photosynthetic assimilation of the whole canopy. [29] Pear Dwarfing and vigourous rootstocks Anatomical investigations of young stems and mature leaves Dwarfing rootstock had lower vessel diameter and vessel density compared with the vigourous type. The lower growth vigour of dwarf pear may be associated with anatomical characteristics. [26] Cherry Gisela 5, Mahaleb, Mazzard, Colt PHL-A Stem and root anatomical characteristics and theoretical hydraulic conductance (kh) Lower hydraulic conductance in dwarfing rootstock were possibly due to smaller vessel lumens and percentage. [28] Citrus Troyer citrange, Rough lemon, Sweet lime, Carrizo citrange, sour orange and Flying dragon Anatomical features of leaf, stem and root Vigourous rootstocks possessed larger vessel elements in the stem and roots as well as lower proportions of bark (phloem) compared with dwarfing rootstocks. [27] Flying Dragon and Rubidoux trifoliate rootstocks Leaf water potential, hydraulic resistance, gas exchange measurements, xylem anatomy, and 13C photoassimilates transport Lowered hydraulic conductance might be the possible reason for rootstock-induced dwarfing in citrus scion when grafted with dwarfing rootstock. [33] Olive Cairo7, Manzanill, Aggezi, Frantoio, Coratina, Koroneiki, Picua, Arbequina, Teffahi, Chemlali, Dolce Histological studies of stem Lower phloem and higher xylem (%) in vigourous rootstock are correlated with growth vigour. [34] Mango Alphonso, Fajri Kalan, Sukkary and Zebda, Tommy Atkins, Sediek, Keitt, 13/1, Naomi, Kent, Mabrouka, Ewais, Hindi Besinnara, Maya Anatomical studies of stem Xylem and phloem percentage could be used as a dwarf indicator for the screening of rootstocks. [35] -
Phytohormones play a significant role in vegetative and reproductive growth and are considered essential for root-shoot communication[38−40]. Hormones are transported as signal molecules, which induce tissue differentiation and perform above and below the graft junction[41] Hormonal signals can also control rootstock-mediated vigour by modifying gene expression in the scion part[11]. In earlier literature (Table 2), reduction in plant growth-promoting hormones (IAA, ZR and GA3) and increase in growth-inhibiting (ABA) in dwarfing rootstocks have been studied. However, the effective hormonal communication of scion vigour is still unclear[28].
Table 2. Important hormonal and carbohydrates evidences underlying rootstock induced vigour control.
Name of crop Rootstock Scion Traits Key findings References Peach Mr.S.2/5, GF677, Armking 'Armking' Xylem exudates Tree growth was positively associated with the xylem transport rate of cytokinin. [37] Apple MM.111, M.27, M.7, and M.9 MM.111, M.9, M.7, M.27, Gala, Fuji Scion above the graft union, rootstock below the graft union, leaf, roots, exudates, current year stem Reduced GA levels and higher ABA levels were linked with a degree of dwarfing. [48] 'Royal Gala', 'M.793', 'MM.106', and 'M.9' 'Royal Gala' Application of growth regulators, Measurements of tree growth, shoot-root-shoot hormonal signalling Dwarfing rootstocks reduce the basipetal IAA transport to the root, thus decrease the amount of root synthesized gibberellin and cytokinin, transported to the aerial part. [34] 'Royal Gala', 'M.793', 'MM.106' and 'M.9' 'Royal Gala' Measurements of scion growth, root growth, total growth per tree, total dry weight quantification of indole-3-acetic acid, cytokinins, and gibberellins. Size-controlling apple rootstocks may restrict root-produced GA19 supplied to aerial part. [18] 'M.9', 'MM', and interstock combinations, 'M.9'/'MM' 'Red Fuji' Hormone determination and the expression of hormone related genes. Lower expression levels of IPT3 gene in dwarfing rootstocks resulted in lowered cytokinin synthesis in roots, which resulted in inadequate supply of cytokinin to the aerial parts, resulting in a reduction of auxin content and scion growth. [49] M.9 and Baleng Crab Scion/rootstock combinations (Fuji/M9), Fuji/
M9/Baleng Crab, Fuji/Baleng CrabDetermination of phytohormone contents and relative expressions of MdPINs genes Dwarfing effect was initiated by inherently lower expression of MdPIN8 in M9 interstem. [50] 'M.9' and 'MM.106' 'Fuji' Investigation of tree growth and measurement of IAA to explain the function of auxin in controlling the dwarfing rootstocks The lower amount of auxin transported from the shoots along with the root auxin synthesis deficiencies reduced the root growth and then decreased the supply of root-produced substances to the shoots in dwarfing rootstocks. [33] 'M27', 'M9' and 'M793' 'Royal Gala' ('RG') Non-structural carbohydrate analysis, RNA sequencing and quantitative reverse transcriptase PCR. Over-accumulation flavonoids and reduced MdAUX1 and MdLAX2 expression contribute to the reduced auxin transport observed in dwarfing rootstocks. [51] Six kinds of apple rootstock ('Mailing 9', 'Mailing 26', 'Budagovsky 9', 'Malus xiaojinensis', 'Malus robusta', 'Malus baccata') and two grafting complexes ('Fuji/M9' and 'Fuji/Mr') 'Fuji' Trans-zeatin determination, and expression of cytokinin metabolic pathway genes Low IPT5b expression with high-level methylations in the promoter region, leading to poor root trans-zeatin biosynthesis in the M9 rootstock, which may induce dwarfing. [52] Pear Dwarf and standard rootstocks − Comparative transcriptomic analysis between standard and dwarf pears of the young apical stem Major differences between dwarf and standard pears were associated with ribosome biogenesis and photosynthesis. [53] Zhongai 1 and Jinxiang − RNA-Seq analysis and the determination of phytohormones (GA3, IAA, and ABA) The dwarf mechanism of different genotypes may be diverse, and even in the same genotype, the dwarf character may be controlled by many factors [54] Sweet cherry Ten different size controlling (Prunus Mahaleb L.) genotypes − Vegetative growth parameters and hormonal ratio The proportion of ABA/IAA hormones was lesser in sturdy rootstocks than that in the dwarf rootstocks. Moreover, the available ABA concentration demonstrates a relationship between growth vigour and ABA in shoot bark of rootstock. [47] Litchi vigourous cultivar (Feizixiao) and dwarf cultivar (Ziniangxi) − Anatomical observation, transcriptome profiling, and mechanisms of litchi dwarfism Upregulation of GA2ox in dwarf cultivar (ZNX) samples only, showing GA may assume a significant role in managing an immense difference between cultivars of dwarf and vigourous litchi cultivars. [43] Sweet Persimmon Rootstock (Diospyros lotus L.) and interstock/rootstock (Nantong-xiaofangshi/Diospyros lotus L.) 'Kanshu' Hormone levels and water conductance The expression of GA2ox, DELLA, and SPINDLY genes were upregulated and associated with a decrease level of GA in scions grafted on the interstock. [55] Citrus Fragrant orange, Rough lemon, Shatangju mandarinCanton lemon, and Red tangerine 'Shatangju' Dwarfing mechanisms and the influence of rootstock on scion growth IAA and GA were significantly and positively related to growth vigour. the qRT-PCR analysis showed that expression levels of ARF1, ARF8, GH3, and IAA4 were negatively correlated with the growth vigour and IAA content. [56] Auxin
-
Indole-3-acetic acid (IAA) is a plant growth hormone directly involved in a variety of biological mechanisms such as cell elongation, differential growth, tissue patterning and embryogenesis[38]. Lockard and Schneider[42] speculated that the reduction in the downward movement of auxin transport in size-controlling rootstocks would limit root growth. Soumelidou et al.[43] checked the auxin transport capacity of different size-controlling rootstocks and found that auxin (IAA) transport capacity was lower in dwarfing rootstock (M.9) than that of vigorous rootstocks (MM.11) in both woody and nonwoody segments. This observation was confirmed by Kamboj et al.[44], who measured the uptake of radio labelled [3H]-IAA of stem segments from five apple rootstocks which differed in their dwarfing effect, and found a reduced uptake and transport in dwarfing rootstock stems compared to stems from non-dwarfing rootstocks.
According to Li et al.[45], when apple scion was grafted onto dwarfing rootstock, the expression of auxin transporter gene PIN1 was substantially decreased, resulting in an inadequate supply of IAA to the roots, causing dwarf phenotype. Song et al.[46] reported that IAA levels of apple tree grafted on vigorous ('Fuji'/MM111) rootstock was higher than that of grafted on dwarfing ('Fuji'/M9) rootstock. Besides, the expression levels of auxin synthesis gene MdYUCCA10a in leaves and roots of size-controlling rootstock were markedly lower than taller/stronger rootstocks. Therefore, a lower amount of IAA was transported from the aerial part to the roots, which decreased the supply of root-produced substances to the shoots. Van Hooijdonk et al.[47] stated that dwarfing apple rootstocks could change the morphology of scion cultivar. 'Royal Gala' grafted with dwarfing M.9 rootstock substantially decreased the node numbers and shoot length. In addition, IAA transport inhibitor, N-1-naphthylphthalamic acid (NPA), injected into the stem of taller/vigorous rootstocks, resulted in reduced shoot length and architectural modifications closely resembled with M.9 rootstock. Therefore, it is suggested that the roots of dwarfing rootstocks obtain less IAA from the scion, thus, reduce the amount of cytokinin and gibberellins produced by roots for transportation to the scion. Consequently, the decrease of cytokinin supply may alter the scion structure by reducing branching.
Cytokinin
-
In contrast to auxins, cytokinin (CKs) are synthesized in the root and transported to the aerial part, where they regulate some of the main developmental processes, including shoot growth and production[15]. The control of vigour may also be due to decreased xylem solutes (e.g., cytokinin) caused by the graft junction between rootstock and scion[43,48]. They also noticed that the cytokinin concentration of dwarfing apple rootstocks in root pressure exudates and leaf xylem sap was lower than that of vigorous rootstocks. Orange trees grafted on less vigorous 'Troyer' (Poncirus trifoliate × Citrus sinensis) rootstock also have lower cytokinin-like activity in branch sap than grafted on vigorous 'Volkamer' lemon rootstock[49]. The content of cytokinin in the stem fluid from different rootstocks was different. The cytokinin concentrations were significantly higher in vigorous rootstock (MM.106) as compared with size-controlling (M.27 and M.9) rootstocks.
Similarly, Sorce et al.[50] also noticed a positive correlation between tree vigour and cytokinin transport rate in the xylem of both un-grafted and grafted peach trees. Trees grafted with dwarfing rootstocks contained lower cytokinin levels than vigorous rootstocks[45]. Most recent studies showed that cytokinin biosynthesis is necessary for bud break initiation and transfer of auxin from buds in an apple tree through primary shoot apex or decapitated apple tree. The relative expression of MdPIN1 indicates that the export of IAA from axillary buds is limited due to the absence of CK synthesis[51]. In apple, the MdIPT5b gene was suggested to have a pivotal role in stimulating the dwarfing characteristics of M.9 rootstock[52]. Taken together, these evidences suggested that the poor root cytokinin biosynthesis and transportability from roots to shoots are key factors responsible for dwarf tree morphology.
Gibberellin
-
Gibberellins play a fundamental role in plant growth and are a controlling factor in plant architecture [53−55]. There is substantial evidence suggesting that disruption in GA metabolism plays a vital role in the dwarfing of the scion[22,28]. Bulley et al.[56] described the role of gibberellins in apple shoot growth. They explained that down-regulation of GA3ox and GA20ox genes reduced the GA level, which resulted in dwarf phenotypes.
In apple, Van Hooijdonk et al.[19] observed that in the first growing season after grafting, the average concentration of GA19 in the xylem sap of the 'Royal Gala' grafted on the M.9 rootstock was twice than that of vigorous rootstocks. Interestingly, the node number of primary and secondary shoots of dwarfing rootstocks restores after scions treated with GA application. In contrast, over-expression of GA2ox genes, which encodes enzymes that convert active forms of GAs into inactive forms, also produces dwarf plants[57]. Scion cultivars grafted with dwarfing rootstocks contained lower levels of GA19 in the xylem exudates compared with other vigorous rootstocks[58].
Hu et al.[59] noted that the GA2ox gene was up-regulated in dwarf litchi cultivar, demonstrating that GA may play a vital role in controlling the substantial differences between dwarf and vigorous litchi cultivars. Similarly, the transcriptome of apple tree graft with dwarf rootstock revealed that GA biosynthesis genes are decreased[60]. The higher expression levels of a GA catabolic gene and reduced GA level in persimmon scion stem grafted with size-controlling interstocks[61]. Furthermore, a dwarf plum hybrid with elevated transcript levels of a major GA catabolic gene, GA2ox, exhibits shorter internodes and reduced stem elongation; when used as rootstocks, it also decreases the amount of bioactive GAs in scions and reduces scion growth[62]. In summary, this literature shows that dwarfing rootstocks decreased GA concentration and altered gene expression associated with GA synthesis, resulting in a dwarf phenotype.
Abscisic acid
-
Abscisic acid (ABA) is known to regulate various aspects of plant development and growth attributes such as leaf abscission, stomatal closure and root growth inhibition[63−65]. ABA controls shoot and root growth in plants that regulate tolerance responses against different stress factors, and it is also capable of inducing dwarfism in higher plants[25,66]. Similarly, Jindal et al.[67] compared ABA-like activity in the shoots of dwarf and normal type mutants of 'Golden Delicious' and 'Cortland' apples and found that maximum levels of ABA were reported in dwarf mutants; similar findings were observed during terminal bud formation, rapid shoot elongation, and termination of cambial growth phases. Kamboj et al.[48] showed that the size-controlling rootstocks had maximum levels of ABA and ABA-IAA ratio in shoot bark compared with vigorous rootstocks. Noda et al.[68] reported that citrus trees grown on dwarfing rootstock had a higher concentration of ABA in new shoots than vigorous rootstocks, and the higher ABA levels were believed to be responsible for the reduction of plant growth. Tworkoski et al.[69] reported that ABA concentrations found in the shoot of apple plants grown with dwarfing 'M.9' rootstocks were higher than those found on vigorous 'Antonovka' rootstocks. A high level of ABA in the xylem sap of trees grafted onto dwarfing rootstock seems important because of its role in vigour control and drought tolerance. Another possibility is that the abnormal xylem configuration at the graft union reduces the value of hydraulic conductance (HC) and increases ABA level in the xylem stream[29,69]. Moghadam and Shabani[70] worked on sweet cherry (Prunus Mahaleb L.) rootstocks and reported that the ratio of ABA/IAA in vigourous rootstocks was minimum compared with more dwarfing rootstocks. Furthermore, they observed a close relationship between ABA concentrations in shoot bark and growth vigour. Briefly, ABA concentrations were found to be higher in the tissues of dwarfing rootstocks (act as a growth inhibitor) and impaired growth characteristics by suppressing the accumulation of other phytohormones.
-
Nutrient and water capture through more efficient root system architecture and/or uptake and transport mechanisms are major rootstock traits that regulate plant growth and crop yield[71]. The influence of rootstocks on leaf mineral content is related to the mineral uptake capacity of rootstocks due to their specific root morphology[72]. Al-Hinai and Roper[73] also reported that rootstock-scion interactions resulted in the difference of transportation and absorption of mineral nutrients to the scion. Previous studies have shown (Table 3) that the nutritional status of trees is related to the vigour controlling capacity of rootstocks[74−78].
Table 3. Influence of rootstock on the vegetative growth, yield, fruit quality, and mineral uptake of grafted plants.
Name of crop Family Name of rootstocks Name of scion cultivar Traits References Apple (Malus domestica Borkh.) Rosaceae M.9, M.26, Chistock-1 and Baleng 'Red Fuji' Dwarfing traits, scion morphology, and mineral concentration [8] M.9, MM.106, MM.111 and local seedling (Malus domestic cv. 'Local' 'Golden Delicious' and 'Royal Gala' Growth vigour and mineral concentration [22] M. hupehensis Rehd., M. prunifolia Borkh., M. robusta Rehd., M. sieversii Roem, and M. rockii − Plant growth and potassium (K) use efficiency [58] Peach (Prunus persica L.) Rosaceae Seven hexaploid plum rootstocks (Adesoto, Monpol, Montizo, P. Soto 67 AD, PM 105 AD, GF 655/2 and Benasque) 'Catherina' Leaf mineral concentration and fruit quality traits [59] almond × peach hybrids rootstocks (Adafuel, Adarcias, GF 677, Cadaman, Garnem and Felinem) Queen Giant and Tebana Vigour, yield and mineral concentration [52] Pear (Pyrus communis L.) Rosaceae Seedlings of Local and colonal pyrus, MA, MC and BA 29 Quince (Cydonia oblonga M.) Santa Maria Fruit quality, yield and leaf uptake of mineral elements [63] Plums (Prunus domestica L.) Rosaceae Marianna GF8-1 and Marianna GF10-2 (Prunus cerasifera × munsoniana) Rainha Claudia Verde Tree vigour, fruit quality and fruit mineral concentration [64] Apricot (Prunus armeniaca L.) Rosaceae Evrica, Krymsk 86, Torinel, PAC 00-08 and PADAC 01-47 Two apricot cultivars ('E-101' and 'E-404') Fruit yield and quality [65] Pistachio (Pistacia vera L.) Anacardiaceae Badami-sefid, Sarakhsi, Kalle-ghouchi, Daneshmandi, Barg-Seyah and Akbari Akbari and Barg-seyah Mineral uptake [66] Grape (Vitis
vinifera L.)Vitaceae 'Couderc 161-49', 'Sori', 'Kober 125AA', 'Börner', 'Kober 5BB' 'Regent' Mineral nutrition and heavy metals uptake [67] Two different rootstocks including (low and high vigour) Cabernet Sauvignon Control of scion vigour, mineral nutrition and whole plant biomass [68] 'IAC 766', 'IAC 572', 'IAC 313' and 'IAC 571-6' 'Venus' Yield and nutrient uptake [69] Citrus Rutaceae Nine different citrus rootstocks Kinnow mandarin (Citrus reticulata Blanco L.) Leaf nutrient status, yield and fruit quality [70] Rough lemon, Karna Khatta, Carrizo citrange, Rangpur lime, Troyer citrange, Jatti Khatti and sour orange Kinnow mandarin Root morphology, mineral uptake and nutrient absorption capacities [71] Hirst and Ferree[79] reported that trees grafted on a local seedling were less prone to potassium (K) and calcium (Ca) deficiency than those grafted on MM.106 rootstock. Aguirre et al.[80] noted that the low mineral absorption rate of dwarfing rootstocks is one of the possible reasons for mineral deficiencies in 'Golden Delicious'. Previous studies have shown that the concentration of potassium (K) and magnesium (Mg) in leaves of trees grafted on vigorous rootstocks were higher than those grafted on dwarfing rootstocks. The nitrogen (N) concentration of trees grafted onto dwarfing rootstocks was higher than that of vigorous rootstocks[80]. Zarrouk et al.[76] reported that different peach rootstocks significantly affect the concentrations of essential nutrients, namely [(Nitrogen (N), Phosphorus (P), Potassium (K), Calcium (Ca), Magnesium (Mg), Manganese (Mn), Iron (Fe), Sodium (Na), Zinc (Zn), and Copper (Cu)]. Furthermore, the effect of five rootstocks grafted with 'Imperial Double Red Delicious' on leaf mineral contents[81]. The results showed that the concentration of N and K in the leaves of M7 and MM106 rootstocks was significantly lower than seedling rootstock. Absorption efficiency and accumulation of potassium were observed differently in different rootstocks; especially under the conditions of K- deficiency, these differences turn out to be more and more evident[82].
The leaves of nectarine showed a significant difference in mineral nutrient concentration when grafted onto Prunus rootstocks[83]. Several studies have shown that there are substantial differences in nutrient absorption and transport among different rootstocks. Moreover, the effect of most rootstock studies was inconsistent from one site to another, the type of scions and change over time. No common rootstocks can accomplish all the requirements of climate and soil conditions. Many of the rootstocks are utilized for adaptability to different environmental conditions[84]. Based on the anatomical study of apple rootstocks, it is concluded that the anatomical structure of the xylem affects the mineral absorption rate[85]. The ability of hydraulic conductance to supply mineral nutrients to leaves through roots and stems are linked to the anatomical structure of rootstock[84]. The lower mineral absorption rate will decrease mineral concentrations, which may lead to a mineral deficiency. In previous studies, reduced hydraulic conductivity also reduced the rate of mineral absorption, growth and yield performance suggested as a possible rootstock dwarfing mechanism[86].
-
Photosynthetic efficiency is generally considered the main physiological parameter for evaluating plant growth activity, biomass and yield potential[87,88]. Net photosynthetic rate is one of the critical factors affecting fruit yield[89,90], various studies have shown that rootstocks directly impact gas exchange characteristics[91,92]. There is a positive correlation between the photosynthetic performance and the production ability of plants[93]. In fruit trees, leaf photosynthetic characteristics are significantly affected by dwarfing rootstocks throughout the growing season. Still, there is a lack of consensus regarding the correlation between dwarfing rootstocks and increased leaf photosynthesis.
However, Baugher et al.[94] have highlighted a negative effect of size-controlling rootstocks on photosynthesis in scion leaves. Other studies have however reported that apple trees grafted onto size-controlling rootstocks with more significant dwarfing effects, had a lower photosynthetic rate[81,95]. In contrast, some studies reported that the Pn rate of field planted apple trees grown on dwarfing rootstock (M-9) was higher than that of M.7[91]. The effect of rootstock on the Pn rate may represent a mechanism through which rootstock employs its impact on scion growth[96]. These conflicting findings may be due to trees of different ages, differences in sampling methods, or differences between the years of testing. In another study, Zhou et al.[37] provide evidence that the limited photosynthetic capacity and leaf area are significant causes of the reduced canopy photosynthetic assimilation observed in dwarfing grafted apple trees.
Moreover, Hayat et al.[27] reported that a significant decline in Pn rate was observed in 'Red Fuji' grafted onto dwarfing (M.9) rootstock compared with more vigourous rootstocks (M.26, Chistock-1 and Baleng). The application of interstock with greater dwarfing abilities causes a decrease in photosynthesis and root-shoot ratio, but yields better fruit qualities in 8-year-old Red Fuji trees[26]. These studies have indicated the notable role of photosynthesis in regulating plant growth through which rootstock employs its impact on scion growth.
-
To better study the dwarfing mechanism and find reliable dwarfing markers is a fast method. Numerous studies have emphasized that DNA markers can identify rootstocks more accurately than morphological markers because developmental stages and environmental factors inhibit physiological and biochemical markers. DNA markers remain the most reliable genetic markers[97]. Apple dwarfing rootstock (M9) is used globally as a genomic asset to breed new rootstocks[98]. Pilcher et al.[99] reported for the first time that the dwarfing ability of the apple rootstock was named Dwarfing 1 (Dw1), which was detected at the position of 2.5-cM between the RAPD DNA marker NZraAM18_700 and simple sequence repeat (SSR) marker CH03a09 at the linkage group 5 of apple rootstock 'Malling 9'. Dw1 is a major element because most of the dwarf and semi-dwarf apple rootstocks carry this allele. Gene identification of Dw1 is a major achievement in studying the dwarfing mechanism of apple rootstocks.
Furthermore, Foster et al.[98] identified Dw1 and Dw2 dwarfing loci in a cross between apple rootstock 'M9' and non-dwarf 'Robusta5'. Dw2 was identified on LG11, and four small QTLs with little effect on LG6, LG9, LG10, and LG12. Dw1 and Dw2 had the most significant impact on rootstock dwarfing, and Dw1 had greater influence than Dw2. The screening of several dwarfing and semi-dwarfing apple rootstocks found marker alleles related to Dw1 and Dw2, which indicates that most of the apple dwarfing rootstock had similar genetic sources.
The precocity and intensity of trees greatly stimulate their effectiveness in commercial production. Dwarfing rootstocks permit a higher number of plantations with efficient flowering allowing early fruit-bearing apple. A previous study reported the first QTL linked to vigour and flowering characters in pear rootstocks. A high-density genetic map (SNP-based) was constructed, consisting of 597 and 113 polymorphic molecular markers in pear and apple respectively, which allowed finding QTLs for precocity and scion vigour in the linkage groups LG5 and LG6. The position of LG5 QTL was consistent with that of apple 'M9'. An early maturing and dwarfing allele linked to apple Dw1 was isolated from pear germplasm. This LG5 QTL from pear and apple increases the possibility of Dw1 dwarfing locus in other Rosaceae species[100]. In apple, two QTLs identified the homology of dwarfing QTLs Dw1 and Dw2, and a third QTL identified on chromosome 13. The closely linked QTL flanking sequence-tagged site markers improve the resolution of the loci, thereby leading to the discovery of dominant and epistatic interactions among loci and further reported the significant negative correlation between stem diameter and root bark. The relationship between rootstock-induced scion dwarfing and root bark percentage was confirmed, and a three-locus model was proposed. Therefore, the newly identified QTL (Rb3) on chromosome 13 might be related to the third dwarfing QTL Dw3, not previously identified[101]. These results will help to expand the scope of marker-assisted selection and finding the basis for dwarfing apple rootstock breeding.
The achievement of whole-genome sequencing delivers a chance to apprehend the phylogenetic relationship of the ABC transporters family genes in Rosaceae species. ATP-binding cassette transporter genes are a varied, omnipresent and wide superfamily that performs several processes. In nine Rosaceae genomes, 1323 ABC transporter genes were detected: 179, 141, 174, 138, 122, 118, 191, and 98 respectively from Rosa chinensis, Prunus dulcis, Pyrus communis, Prunus persica, Fragaria vesca, Prunus avium, Malus domestica, and Rubus occidentalis. Furthermore, transmembrane structures, subcellular localization, and protein motifs were studied. Based on structural features and phylogenetic analysis, these ABC transporters were assembled into eight subfamilies. In apple rootstocks, the W-box deletion in the promoter of MdABCG28 was linked to gene expression and dwarfing phenotype. The overexpression of MdABCG28 enhanced shoot growth in mutants of Arabidopsis[52].
-
Grafting is a centuries-old technique used in several horticultural species to improve biotic and abiotic stress resistance, yield and quality. The alterations caused by grafting, as well as the physiological and fewer genes involved in molecular mechanisms related to dwarfing, remain unclear. The use of dwarfing rootstocks or interstock is a primary approach to produce dwarfing fruit trees, as these rootstocks and interstock result in reduced tree volume, height, canopy diameter and circumference. The current evidences showed that size control of rootstock is regulated by a complex interaction between scion and rootstock. The dwarfing rootstocks have a smaller root system (i.e., root volume, root diameter, number of root tips and branches), so the soil absorbs less water and nutrients. A lower volume of water is transported to the scion due to lower hydraulic conductance and reduced size of xylem vessels that influence net photosynthetic rate (Pn) and plant growth. The IAA absorbed from the scion of dwarfing rootstock was less, which affects the production of cytokinin (CK) and gibberellins (GA) produced by the root system, and ultimately less are transported to the aerial part and reduce plant growth.
- The authors, thanks to all the researchers who contribute in this study. This study was supported by the National Key Research and Development Program of China (2020YFD1000203) Project for Crop Germplasm conservation and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD) for funding this research in materials collection, data analysis, and experiment.
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Hayat F, Iqbal S, Coulibaly D, Razzaq MK, Nawaz MA, et al. 2021. An insight into dwarfing mechanism: contribution of scion-rootstock interactions toward fruit crop improvement. Fruit Research 1: 3 doi: 10.48130/FruRes-2021-0003

An insight into dwarfing mechanism: contribution of scion-rootstock interactions toward fruit crop improvement
- Received: 20 March 2021
- Accepted: 25 May 2021
- Published online: 23 June 2021
Abstract: Grafting has been commonly practiced for many centuries in the cultivation of horticultural crops. The use of dwarfing rootstocks has enabled a high-density plantation to produce maximum yield. Rootstock regulates scion phenotype, including precocity, fruit size, yield, quality characteristics, and tolerance to various environmental stresses. This review summarizes the existing information on the influence of rootstocks on scion growth and dwarfing mechanisms induced by multiple factors, including hormone signaling, photosynthesis, mineral transport, water relations, anatomical characteristics, and genetic markers. It has been shown that the complex interactions between scion and rootstock can regulate plant development and its structure. This information will provide interesting insights for future research related to rootstock-mediated dwarfing mechanisms and accelerate the breeding progress of dwarfing rootstocks.
-
Key words:
- Fruits /
- dwarfing rootstock /
- hormonal signaling /
- dwarfing mechanism