









-
The flowering transition from vegetative to reproductive growth is an important physiological process in higher plants, and a crucial turning point in the development of flowering plants. Controlling the timing of this transition can be particularly important in ensuring high levels of agricultural productivity. Baicaitai (Brassica rapa ssp. pekinensis) is a new variety of Chinese cabbage, whose stalk is desirable because it is both nutritious and has a pleasant taste. As it is a bolting vegetable, early bolting is an important agronomic trait for Baicaitai, but Chinese cabbages are generally late flowering. Therefore, an important objective in breeding new Baicaitai varieties is to identify new germplasm for early bolting. Understanding the bolting mechanism is thus of great significance in agronomic practices, to improve the breeding efficiency of Baicaitai.
Plant flowering time is mainly regulated by environmental and endogenous signals. Many of the genes that control flowering have been identified in the model plant Arabidopsis[1,2], in multiple pathways, including the photoperiod, vernalization, autonomic, and gibberellin pathways[3−6]. Although these pathways independently regulate plant flowering, eventually, they converge on common downstream genes that play important roles in the integral flowering pathway. Generally, it is believed that CONSTANS (CO) and FLOWERING LOCUS C (FLC) may influence the downstream genes of the flowering pathway. CO is the most downstream gene in the photoperiod pathway, with a zinc finger domain, and the accumulation of CO protein can be regulated by plant photoreceptors and biological rhythms[7,8]. FLC is the confluence of the autonomous and vernalization pathways, and encodes a transcription factor with a MADS domain, which inhibits the conversion of leaves into floral organs[4,9−11]. However, vernalization and autonomous pathways inhibit the relative expression levels of FLC and regulate early bolting and flowering[10,12−14]. In the vernalization pathway VIN3 (VERNALIZATION INSENSITIVE 3), VRN1 (vernalization 1), and VRN2, and in the autonomous pathway FLOWERING LOCUS D (FLD), FLOWER LOCUS VE (FVE), FLOWER LOCUS Y (FY), FLOWER LOCUS CA (FCA), FLOWER LOCUS PA (FPA), REF6 (RELATIVE OF EARLY FLOWERING 6), and FLOWER LOCUS K (FLK), were all found to be involved in the inhibition of FLC expression and the promotion of early flowering in plants[15−18].
FLC genes encode a MADS-box protein that can inhibit flowering in the vernalization pathway, as many genes regulate flowering via FLC. Five FLC genes with similar sequences were isolated from Brassica napus[19]. To date, four copies (BrFLC1, BrFLC2, BrFLC3, and BrFLC5) have been cloned in Brassica rapa[2,20], that are respectively located on the A10, A02, A03, and A03 chromosomes, owing to polyploidy evolution[2]. While BrFLC5 is in the non-syntenic region (between I block and J block) and originated from the α replication of a common ancestor of Arabidopsis and Brassica rapa[21], it was lost from Arabidopsis after it diverged from Brassica. The dosage-dependent expression of multiple copies of FLC genes may regulate flowering time in B. rapa[22]. BrFLC1, BrFLC2, and BrFLC3 were transferred to Arabidopsis, and the transgenic plants flowered late, revealing that they played a similar role to AtFLC in the regulation of flowering time[23]. In B. rapa natural populations, spicing variants of BrFLC1 and BrFLC5 were associated with flowering time[2]. A splicing variation of BrFLC5 was significantly related to flowering time variation[24]. Early-flowering phenotypes were observed in the CRISPR/Cas9 knockouts of BrFLC2 and BrFLC3[25]. During the last two decades, with the rapid development of molecular markers and quantitative trait loci (QTL) analyses, the focus of research has been to analyze the genetic basis of the bolting-related traits of B. rapa, and many QTLs have consequently been identified that can be used to evaluate populations in different environments, locations, and seasons based on their phenotypic scores[26−29]. Xiao et al. (2013)[28] suggested that BrFLC2 was a key factor in the regulation of flowering time in Brassica rapa. A major QTL with BrFLC2 was found on A02 in B. rapa, and the gene was identified as the candidate gene for bolting[26,27]. Wu et al. (2012)[3] discovered that a naturally occurring deletion mutation across exon 4 and intron 4 in the BrFLC2 contributed to variations in the flowering time of oil-type B. rapa. Several QTLs have been identified as being associated with BrFLC1, BrFLC3, BrFLC5, BrFT, CLV1, FLM, and LFY that underlie bolting or flowering in B. rapa[20, 26, 29,30].
Chinese cabbage, B. rapa ssp. pekinensis (2n = 2x = 10, genome AA) is an important vegetable in China, and its bolting ability is an important agronomic trait. We identified an early bolting mutant 6 (ebm6) via EMS mutagenesis with a DH line of heading Chinese cabbage to create germplasm for Baicaitai breeding. MutMap and KASP were applied to identify the causal gene of the mutant ebm6. This investigation has produced a new germplasm resource and provided valuable information for future Baicaitai variety improvements.
-
In this investigation, the doubled haploid (DH) Chinese cabbage line 'FT' was obtained by using microspore cultures of variety 'Fukuda 50' as the wild-type line. The 'FT' seeds were treated with 0.8% (w/v) EMS for 12 h to create mutants that possessed the early bolting trait.
Bolting trait evaluation
-
The bolting trait investigation focused on four bolting characteristics: bolting index (BI), days to reach a 5-cm-high elongated stalk (DE5), days to reach a 10-cm-high elongated stalk (DE10), and flowering time, to determine the phenotypic value of the wild-type 'FT' and mutant ebm6. Based on the method described by Yang et al. (2007)[31], when the first flower of the plants was fully open, the bolting scale of the BI was appraised, but the method was modified slightly, due to the different materials used[29]. DE5/10 referred to the number of days from sowing in until the plants were 5/10-cm in height with elongated floral stalks, which was the distance from the cotyledon to the growth vertex. The flowering time was the days from sowing until the complete opening of the first flower. Respectively, 30 wild-type 'FT' and 30 mutant ebm6 were used to identify the bolting characteristics in the natural environment. Each measurement involved three biological replicates, including ten plants per replicate. In the spring of 2019, the bolting trait evaluation experiments were performed at the greenhouse of the Scientific Research Experiment Base of Shenyang Agricultural University. During the investigation, the temperature was 15–23 °C, and the day length was 13–15 h.
Genetic analysis of the mutant trait
-
The early bolting mutant 6 (ebm6) was selected from the obtained mutants. For genetic analysis, the mutant ebm6 (P2) was crossed with the wild-type 'FT' (P1) to construct the F1, F2, and BC1 progenies. The phenotypes of each generation (P1, P2, F1, and F2) were identified, and the segregation ratio of the F2 and BC1 populations was tested using chi-square (
${\textit{χ}}^2 $ ) tests.Construction of the mapping population
-
The F2 population was used for MutMap mapping and SNP genotyping. In autumn of 2019, all plants were grown in the greenhouse of the Scientific Research Experiment Base of Shenyang Agriculture University. Throughout the entire growth stage of the plant, the environmental temperature was 14–18 °C, and the day length was 12–13 h.
MutMap analysis to identify candidate gene for Brebm6
-
DNA was extracted from young and healthy leaf samples of the wild-type 'FT', mutant ebm6, and the 40 early bolting individuals of the F2 population. According to the description of Murray and Thompson (1980)[32], the CTAB method was used to extract the DNA with some modifications. The DNAs of the 40 plants were combined in equal ratios to build a mutant bulk. The DNA of the wild-type 'FT', mutant ebm6, and mutant bulk were then used for genome resequencing with an Illumina NovaSeq 6000. To ensure the quality of the data analysis, high quality clean reads were obtained by filtering out low quality reads. The Burrows-Wheeler Aligner software was applied to align the clean reads to the Brassica reference genome (BRAD: http://brassicadb.cn), and Samtools was used to sort the alignment file[33]. Then SNPs were identified using the software GATK[34] and ANNOVAR software[35]. The distribution of the SNPs in different functional regions of the genome was confirmed, and non-synonymous or stop-gain mutations inferring the protein function were identified. The SNP-index was calculated using the method of Abe et al. (2012)[36].
KASP analysis
-
Kompetitive allele specific PCR (KASP) genotyping technology was used to detect the co-segregation of each SNP, confirming the candidate gene Brebm6. For the KASP, 208 F2 plants were used to validate the genotypes of the early bolting mutation sites. The experiment was carried at the Beijing Vegetable Research Center of Agriculture and Forestry Academy. Allele-specific primers are listed in Supplemental Table 1.
Protein sequence and phylogenetic analysis
-
The full-length DNA and the encoded protein sequences for the candidate gene ebm6 were retrieved from the Brassica reference genome database. The full-length sequences of the wild type 'FT' and mutant ebm6 were cloned using gene-specific primers that are listed in Supplemental Table 2. Multiple sequence alignments were visualized using DNAMAN. Smart online (http://smart.embl-heidelberg.de) software was used to analyze the conserved domains of the protein encoded by the candidate genes.
Quantitative reverse transcription-PCR (qRT-PCR)
-
At the flowering stage, the total flower RNAs of the wild-type 'FT' and mutant ebm6 were extracted according to the instructions of the RNA extraction kit (TIANGEN, Beijing, China). The flowers were collected 70 d after the seeds were sown. First-strand cDNA was formed via reverse transcription of the total RNA using the Fast Quant RT Kit (TIANGEN, Beijing, China). The Ultras SYBR Mixture (CWBIO, Beijing, China) was used to configure the 20 μL system, the qRT-PCR reaction was conducted with QuantStudioTM 6Flex Real-Time PCR System (Applied Biosystems, USA). Each reaction was performed for three biological replications. The 2−ΔΔCt method was applied to calculate the relative expression levels[37]. Gene-specific primers and the actin[38] sequences for the qRT-PCR amplification are shown in Supplemental Table 3.
Data availability
-
The datasets generated for this study can be found in BioProject (https://submit.ncbi.nlm.nih.gov/subs/bioproject/) under accession number PRJNA664483.
-
The microspore DH line 'FT' of Chinese cabbage is characterized by heat resistance and an ovoid leaf head (Fig. 1a). Thirteen stably inherited early bolting mutants were created by mutating the wild-type 'FT' (Fig. 1d). Of these mutants, seven were selected for bolting characteristic analysis, and were named ebm5-1, ebm5-2, ebm6, ebm7, ebm8, ebm9, and ebm10, respectively.

Figure 1. Phenotypic characterization of the Chinese cabbage 'FT' and early bolting mutants. (a) Wild-type 'FT' with ovoid leaf heads grown in an open environment. (b) Mutant ebm6, flowering in an open environment. (c) Wild type 'FT' (left) and mutant ebm6 (right) at the flowering stage. (d) Phenotypes of 13 stably inherited early bolting mutants.
Four bolting characteristics, BI, DE5, DE10, and flowering time, indicated that mutant ebm6 greatly differed from the wild type 'FT' in the spring of 2019 (Table 1). Compared with the wild-type 'FT', the mutants had considerably earlier flowering times under long day conditions (LDs). An exhaustive investigation of the flowering time indicated that the mutants had an advantage due to their bolting and flowering characteristics in LD conditions, when either DE5/10 or the days to flower at bolting were measured (Table 1). The late bolting parent, wild type 'FT', had a higher DE5, DE10, and later flowering time than the early bolting mutants, but the wild type had a lower BI.
Table 1. Phenotypic values for wild-type 'FT' and seven Chinese cabbage mutants in the same environment.
Material BI DE5 DE10 Flowering time 'FT' 67.49 ± 0.69 63.50 ± 0.45 67.42 ± 1.16 68.38 ± 0.70 ebm5-1 91.20 ± 0.92** 42.53 ± 0.67** 45.40 ± 1.25** 46.50 ± 1.18** ebm5-2 89.17 ± 0.88** 47.58 ± 0.88** 50.31 ± 1.13** 51.37 ± 0.93** ebm6 88.32 ± 0.45* 37.49 ± 0.77* 40.70 ± 0.51* 55.63 ± 0.88* ebm7 82.38 ± 0.84* 51.34 ± 0.67* 54.47 ± 0.81* 41.36 ± 0.62* ebm8 83.2 ± 0.77* 51.23 ± 0.48* 54.53 ± 0.56* 55.29 ± 0.51* ebm9 83.38 ± 0.74* 50.34 ± 0.55* 53.77 ± 0.31* 44.36 ± 0.55* ebm10 80.2 ± 0.87* 52.33 ± 0.48* 55.76 ± 0.45* 60.31 ± 0.23* Thirty plants from each line were used to identify the bolting characteristics in the natural environment. BI, bolting index; DE5, days to reach a 5-cm-high elongated stalk; DE10, days to reach a 10-cm-high elongated stalk. Mean and standard error (SE)-values were calculated from three independent replicates. Data were analyzed using SPSS16.0 (Chicago, IL, United States). * and ** indicate significant differences at 0.1 and 0.05, respectively. Genetic characterization of the mutant ebm6
-
Among the mutants, ebm6 (Fig.1b, c right) was screened to identify the mutant gene, Brebm6. Except for its flowering time, the other phenotypic characteristics of the mutant ebm6 were the same as those of the wild type 'FT'. For genetic analysis, F1 and F2 populations were prepared with the parents 'FT' (P1) and mutant ebm6 (P2), and the F1 phenotype was the same as that of the wild type 'FT'. The F2 progenies were assessed for trait segregation, and Chi-square (
${\textit{χ}}^2 $ ) tests were conducted to test the 3:1 segregation ratio. The${\textit{χ}}^2 $ test showed that the separation ratio was in accordance with the expected ratio (${\textit{χ}}^2 $ = 0.06 <${\textit{χ}} _{0.05}^2 $ = 3.84). In addition, the plant ratio in the BC1 progenies was approximately 1:1. These results indicated that a single recessive nuclear gene controlled the early bolting characteristic. The detailed segregation data for the mutant trait are displayed in Table 2.Table 2. Genetic analysis of the early bolting Chinese cabbage mutant ebm6.
Generation Total Wild-type
'FT'Mutant
ebm6Segregation
ratio${\textit{χ}}^2 $ textP1 ('FT') 50 50 0 P2 (ebm6) 50 0 50 F1 (P1 × P2) 82 82 0 F1 (P2 × P1) 150 150 0 BC1 (F1 × 'FT') 78 78 0 BC1 (F1 × ebm6) 75 40 35 1.14:1 0.35 F2 186 138 48 2.88:1 0.06 Identification of the candidate gene Brebm6
-
The mutant gene Brebm6 was identified according to the MutMap method introduced by Abe et al (2012)[36]. From the F2 population, 40 individual plants with an early bolting phenotype were selected to extract DNA (bulk-M). The DNA of the wild-type 'FT', mutant ebm6, and the bulk-M were whole genome re-sequenced. We obtained 94,719,968, 128,324,310, and 167,580,256 clean reads for the wild-type 'FT', mutant ebm6, and bulk-M, respectively, with average sequencing depths higher than 30 ×. A total of 98.00%, 99.05%, and 98.83% of the clean reads were aligned to the Chinese cabbage reference genome (http://brassicadb.cn). There were 1,950,825, 1,978,321, and 2,033,988 potential polymorphic SNPs detected in the wild-type 'FT', mutant ebm6, and bulk-M, respectively, using the mutation analysis software GATK. According to the genome annotation file, the ANNOVAR software was used to annotate the filtered SNPs. To ensure the accuracy of the association results, the detected SNPs were strictly filtered, and the SNPs from the bulk-M were deducted from all of the wild-type 'FT', leaving the SNPs associated with the mutant, finally obtaining 718 high-quality SNPs. Then the SNP index of every screened SNP was calculated in all ten chromosomes of Chinese cabbage, and their indices are presented in a scatter plot (Fig. 2). The SNP index was calculated in a one-SNP increment and a two-SNP window. When the SNP index = 0.95 was selected as the threshold, two candidate regions (552,234–2,180,035 and 2,871,165–3,022,398) were localized to a genomic physical distance of 1.78 Mb on chromosome A02. Seven SNP mutations occurred in the exon, but three SNPs (SNP581431, SNP1410912, and SNP2159660) caused non-synonymous amino acid changes, and one SNP (SNP1619528) led to a stop-gain mutation (Table 3).
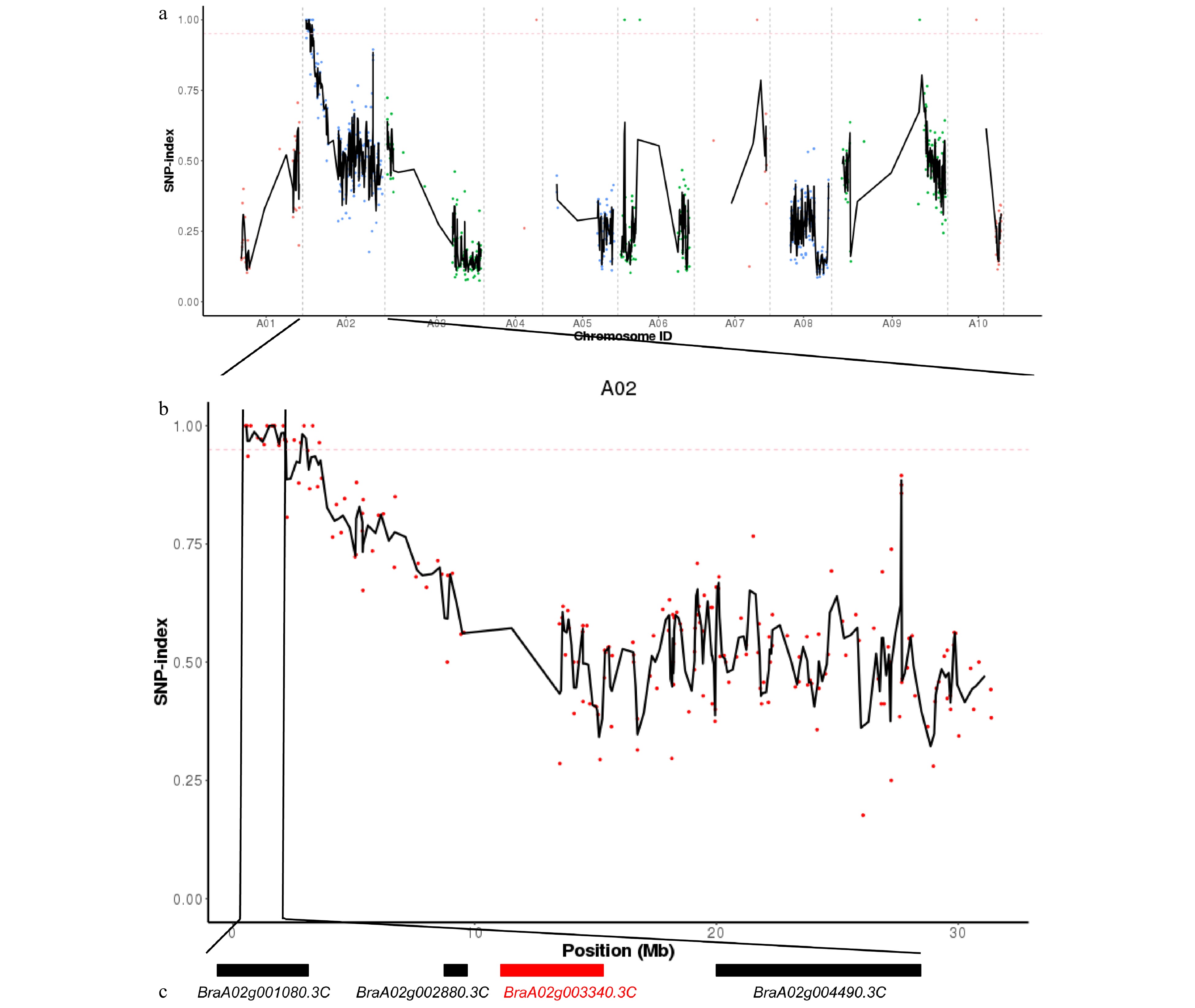
Figure 2. Distribution of the early-bolting mutant site in Chinese cabbage, as determined with Mutmap and analysis of the candidate genes. (a) SNP index of ten chromosomes plotted using MutMap. The x-axis indicates the positions of the ten chromosomes. The y-axis indicates the SNP index. The dotted pink line is the threshold of the SNP index (0.95). Black lines were created by averaging the SNP indices from a moving window spanning two consecutive SNPs and a moving window with one SNP at a time. (b) The SNP index plot from MutMap identifying the candidate regions on chromosome A02. (c) Four candidate genes in the mapping region. Genes indicated by black boxes are listed in detail in Table 3. The red box represents the mutant gene.
Table 3. Candidate gene information from the candidate interval.
Name Gene ID Chr Variation site Variation base Exon ID Mutation type SNP index SNP581431 BraA02g001080.3C A02 581431 G-A exon1 nonsynonymous SNV 1 SNP1410912 BraA02g002880.3C A02 1410912 G-A exon1 nonsynonymous SNV 1 SNP1619528 BraA02g003340.3C A02 1619528 G-A exon1 stopgain 1 SNP2159660 BraA02g004490.3C A02 2159660 G-A exon3 nonsynonymous SNV 0.97 To confirm the candidate SNP, we utilized KASP technology to carry out genotyping analysis with the mutation information of these four SNPs in 208 F2 individuals (112 wild-type phenotypic plants and 96 mutant phenotypic plants). The KASP results revealed that SNP1619528 of BraA02g003340.3C was an A:A genotype in the 96 F2 early bolting plants and A:G or G:G genotype in the 112 F2 wild-type phenotypic plants, which suggested that SNP1619528 co-segregated with the early bolting phenotype (Fig. 3). In contrast, the other three SNPs (SNP581431, SNP1410912, and SNP2159660) were detected for A:A and A:G in 96 early bolting plants, and three mentioned genotypes were all detected in the 112 F2 wild-type phenotypic plants, implying that the three SNPs did not co-segregate with the early bolting phenotype. BraA02g003340.3C was determined to be a FLOWERING LOCUS C − transcription factor (FLC) that encodes a MADS-box protein, which functions as a repressor of floral transition and contributes to temperature compensations for the circadian clock. Therefore, these observations suggested that BraA02g003340.3C is the most likely candidate gene for ebm6, which was annotated as BrFLC2 in the B. rapa reference genome.

Figure 3. Genotype detection in 208 F2 individuals. KASP technique was used for genotyping and the genotype of candidate SNP was detected to be A:A in 96 recessive homozygous early bolting plants.
Sequence analysis of BrFLC2
-
Sequence analysis showed that the BrFLC2 gene consisted of seven exons and six introns (Fig. 4a), whose length of genomic and cDNA sequences were 3,259 bp and 591 bp, respectively. And BraA02g003340.3C corresponded to a base transition from C to T at 52 bp of the first exon (Fig. 4b), resulting in a mutation that converted the Q (CAA) in the wild type to a stop-gain codon (TAA) in the mutant ebm6, which leads to a premature termination of translation (Fig. 4a, b). According to the protein structural property analysis, it was indicated that BrFLC2 included a MADS domain (amino acids 1−60) and a Pfam K-box (amino acids 73−164) through the online software SMART program. For the SNP1619528 was located at amino acid 18, the mutant amino acid was located in the MADS conserved domain (Fig. 5c).
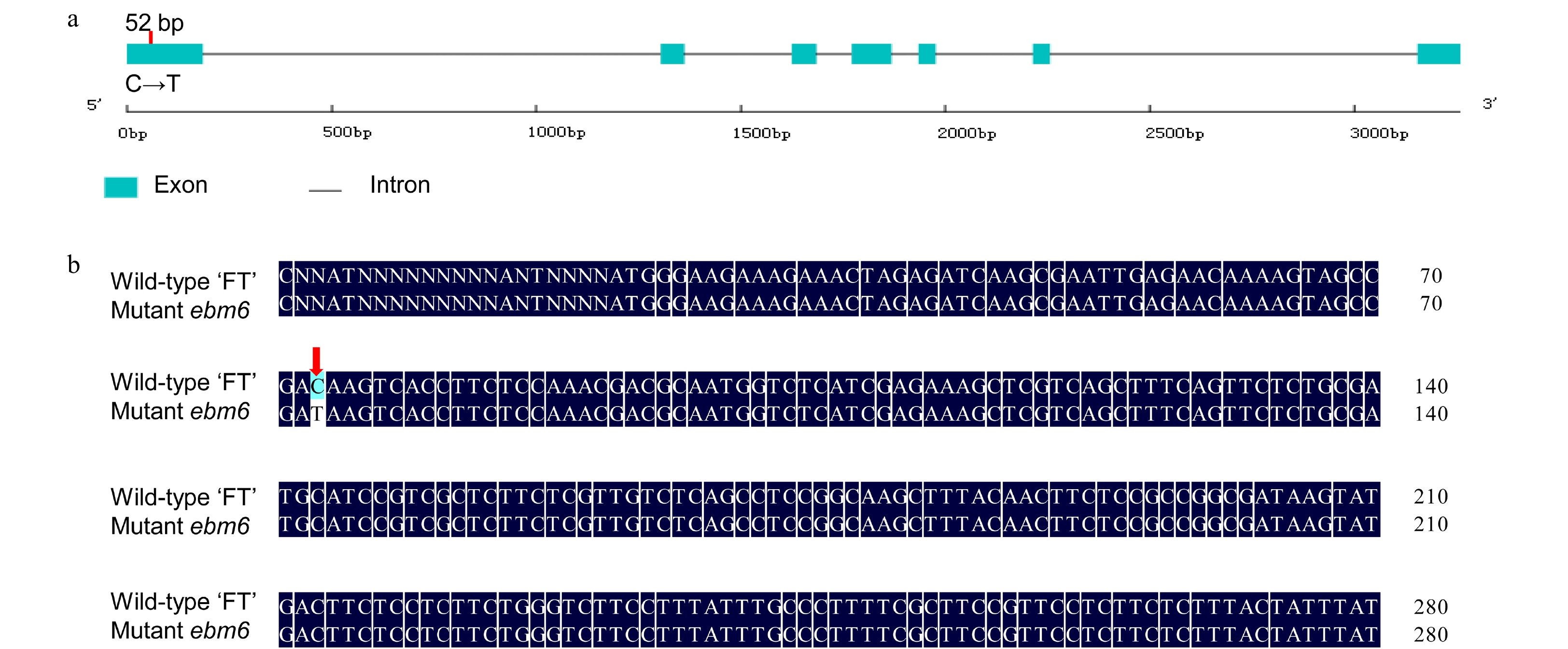
Figure 4. BrFLC2 gene structure and sequence alignment in Chinese cabbage. (a) The gene structure of BrFLC2 and the location of the stop-gain SNP in the mutant ebm6. The red vertical line indicates the SNP site. (b) Alignment of the gene sequences of the BrFLC2 in the wild-type 'FT' and mutant ebm6. The red arrow indicates where the SNP was altered.
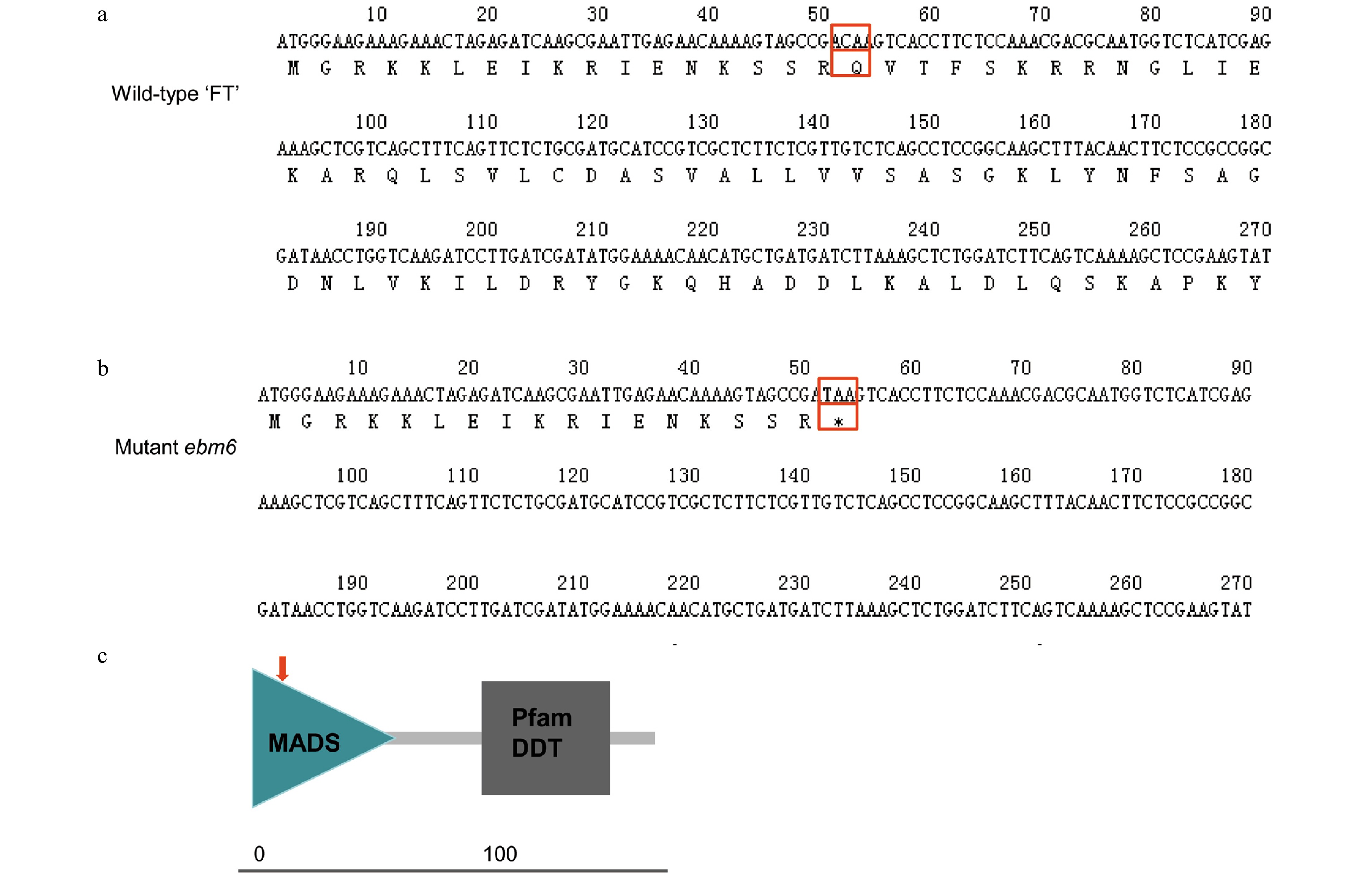
Figure 5. Translation of the protein sequences encoded by the partial nucleotide sequences of the Chinese cabbage 'FT' and mutant ebm6 lines, and analysis of their conserved domains. (a) Partial nucleotide sequence of the wild-type 'FT' translated into its protein sequence. The mutant site is identified with a red empty box. (b) Partial nucleotide sequence of the mutant ebm6 translated into its protein sequence. The red arrow indicates where C mutated to T. (c) The conserved domains of BrFLC2. The MADS domain sequence of BrFLC2 is from amino acids 1−60, and the Pfam K-box is from amino acids 73−164; the red arrow indicates the mutation site.
Expression pattern analysis of ebm6
-
To investigate the relative expression patterns of BrFLC2 in the mutant ebm6, the plant flowers were used for qRT-PCR analysis, and there are significant differences between the mutant ebm6 and the 'FT'. Previous studies have shown that BrFLC2 was a repressor of flowering inhibition and that it could regulate flowering time by inhibiting downstream flower integration genes[3,4,9].
The relative expression levels of BrFLC2 were significantly reduced in the mutant ebm6 flower (Fig. 6a). The expression levels of the other three FLC homologous genes were compared in the wild type and mutant flowers. The expression of BrFLC1 in the mutant ebm6 was slightly lower than in the 'FT' (Fig. 6b). The expression levels of BrFLC3 and BrFLC5 were also significantly lower in the mutant ebm6 than in the 'FT' (Fig. 6c, d). These results suggest that BrFLCs could influence the early bolting trait in mutant ebm6.
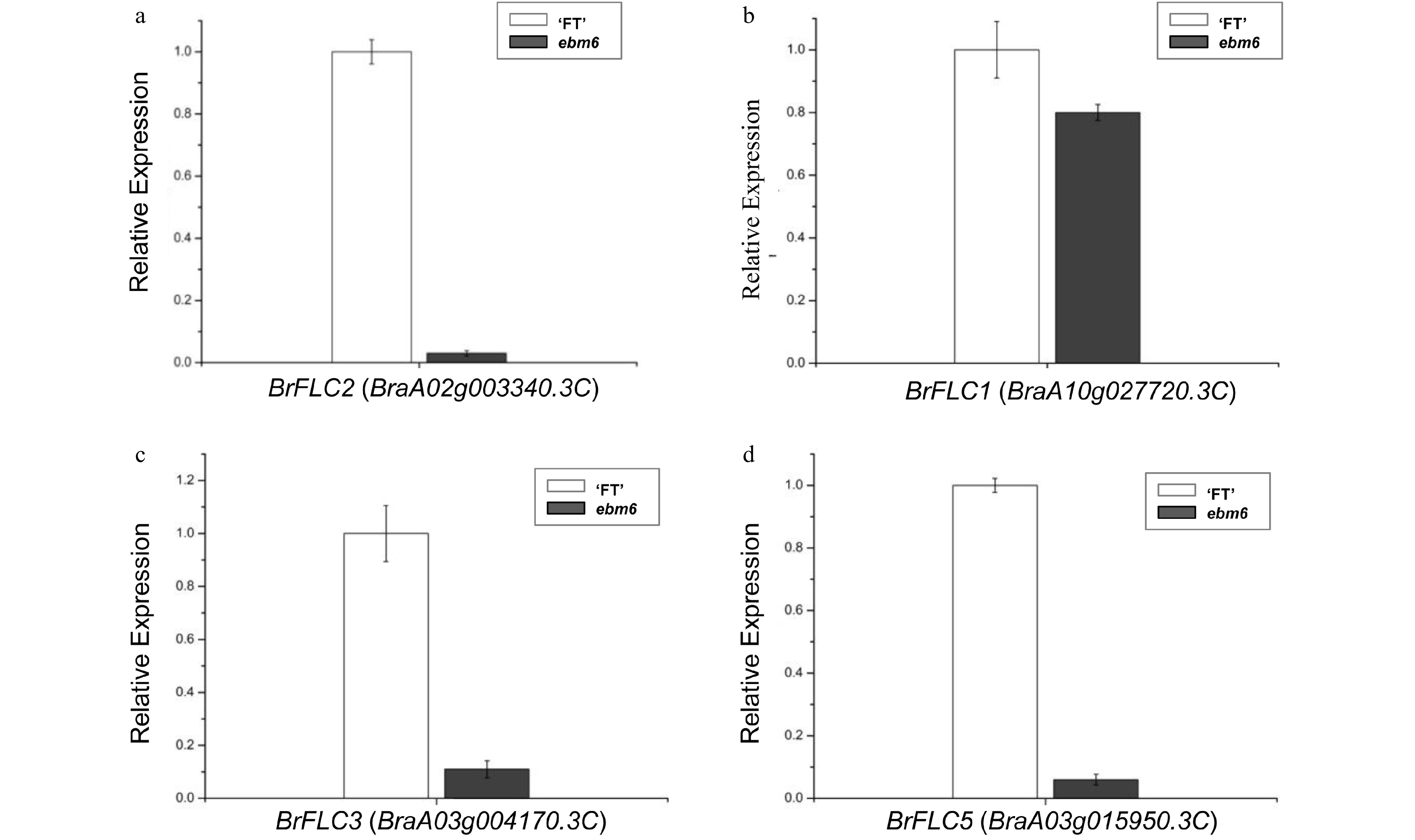
Figure 6. Gene expression levels, determined via qRT-PCR, for Chinese cabbage flowers from the 'FT' and mutant ebm6 lines. (a) Analysis of the BrFLC2 expression levels in the flowers of the 'FT' and mutant ebm6. (b−d) BrFLC1, BrFLC3, and BrFLC5 expression levels in the 'FT' and mutant ebm6, respectively. Standard errors from three replicates are represented with the error bars.
-
Baicaitai is a Chinese vegetable whose stalk emergence is a bolting phenomenon. Like other flowering plants, it originates from the process of flower bud differentiation and development. Bolting is an important agronomic trait, and it is one of the main signals of the switch from vegetative to reproductive growth, and flowering time is determined by external environmental factors and endogenous signals. In the model plant Arabidopsis, the pathways involved in the regulation of flowering are interlinked, and a complex regulatory network is formed to influence it. Bolting mutants are valuable resources used to study the mechanisms of early flowering in Chinese cabbage. It is necessary and essential to understand the genetic regulation mechanisms of the bolting trait. Hence, in this study, 13 stably inherited bolting mutants were obtained via EMS treatments of the 'FT' seeds, and were named ebm5-1, ebm5-2, ebm6, ebm7, ebm8, ebm9, and ebm10, respectively. Mutant ebm6 was then used to identify the early bolting gene, BrFLC2 (BraA02g003340.3C) that encodes a MADS-box, responding as a transcription factor to inhibit flowering conversion. The flowering time of the mutant ebm6 was discovered to be earlier than that of 'FT'. The mutant ebm6 was determined in the genetic analysis to be regulated by a single recessive gene, BrFLC2. The causal mutation of the early bolting trait may be a non-synonymous base change in BrFLC2. The early bolting mutant gene was located between two regions on chromosome A02, with a total length of 1.78 Mb, and it contained three non-synonymous SNPs (SNP581431, SNP1410912, and SNP2159660) and one stop-gain SNP (SNP1619528). The SNP1619528 of BraA02g003340.3C was further identified as a candidate gene using the KASP method, as it co-separated with the early bolting phenotype. In combination, cloning with the functional annotation of BraA02g003340.3C resulted in the speculation that it may be a potential mutant gene functioning as the early bolting mutant ebm6. BraA02g003340.3C, an AtFLC homologous gene of Arabidopsis, encodes a transcription factor with a MADS-box protein, the mutation of which can promote the early-flowering phenotype in Arabidopsis[9], so the gene was named BrFLC2.
Bolting is generally considered to be a quantitative trait, whose associated QTLs have been mapped[30, 39−41]. Like the early bolting traits of ebm1 and ebm5-1/2 that were previously studied by our team[42,43], in this investigation, the early bolting character of mutant ebm6 was also determined to be a quality trait. A stable early bolting mutant ebm6 was created from the DH line 'FT' that was treated with EMS. BrFLC2 was identified to be a candidate gene for the mutant ebm6 in Chinese cabbage. Compared with the wild-type 'FT', a base substitution of C to T at the first exon of BrFLC2, co-segregated with the early-bolting character, which mutated the codon CAA (glutamine) into a terminator TAA, leading to the early termination of translation. Accordingly, BrFLC2 was predicted to be the causal gene for the early bolting mutant ebm6.
In Brassica rapa, BrFLC1, BrFLC2, BrFLC3, and BrFLC5 are copies of FLC[20], the first three of which are homologous with AtFLC genes, as they have high sequence similarities[44]. In Arabidopsis, AtFLC is a key gene controlling flowering time variation[45]. While in Brassica rapa, BrFLC5 was discovered to be a weak regulatory factor of bolting and flowering[24]. A previous investigation by Kim et al (2006)[23] indicated that BrFLC1, BrFLC2, and BrFLC3 played similar roles to AtFLC in the regulation of flowering. Furthermore, BrFLC2 was determined to be a major regulatory factor of flowering in B. rapa, by means of transcriptional co-expression networks[24,28]. Here, gene expression analysis indicated that the expression of BrFLC2 in mutant ebm6 flowers was remarkably lower than in 'FT'. However, the expression of BrFLC1 in mutant ebm6 was slightly lower than in the 'FT', probably owing to the differences in function of BrFLC1 and BrFLC2[3]. Additionally, the expression levels of the other two BrFLCs were also lower in the mutant ebm6 than in the 'FT'. In early-flowering Chinese cabbage, studies have shown that the relative expression levels of the BrFLC1, BrFLC3, and BrFLC5 were lower, implying the BrFLC transcript levels were related to flowering[44]. Since the base substitution occurred at the first exon in BrFLC2, which resulted in the premature termination of the amino acid encoded in mutant ebm6, we considered this mutation may have affected the expression level of BrFLC2, and a loss-of-function protein was created, which influenced the function of the BrFLC2 protein. Furthermore, there were very high levels of sequence similarity for the FLC homologous genes in Chinese cabbage, leading to speculation that this may be the reason for the low expression levels of the other three genes (BrFLC1, BrFLC3, and BrFLC5) in the mutant ebm6. Therefore, we inferred that BrFLC2 was the candidate gene for mutant ebm6, with the early bolting phenotype.
Baicaitai originated in the Yangtze River basin of China in the 1980s and has since become a popular variety of vegetable in Southern China. The term Baicaitai refers to the easy bolting Chinese cabbage that was obtained by selection and breeding processes, with the young flower stalks being the main edible part. Among the seven early bolting mutants identified, ebm6 seems to have the best potential in Baicaitai breeding as it has the strongest stem and best growth. Due to its popular potential and demand has developed in the markets around the country, it has consequently become the focus of research. In future Baicaitai breeding, the BrFLC2 variation could be used to obtain easy bolting Chinese cabbage varieties. By using the BrFLC2 mutant in crosses with B. rapa, the Baicaitai breeding materials could be created by sexual hybridization and gene recombination, to address the current lack of germplasm resources for Baicaitai.
In summary, we identified gene BrFLC2 in Chinese cabbage line ebm6, a mutant created via EMS- mutagenesis and characterized by an early flowering trait. We conjectured that BrFLC2 might act as a negative regulator in flowering, and this gene mutation affected the early flowering phenotype in Chinese cabbage. Our results will help further facilitate understanding of the biological and molecular mechanisms of early flowering.
- The research was supported by the National Natural Science Foundation of China (Grant No. 31730082). We would like to thank Editage for English language editing.
- The authors declare that they have no conflict of interest.
- Supplemental Table 1 List of primer sequence of KASPs.
- Supplemental Table 2 Primer sequence for BraA02g003340.3C.
- Supplemental Table 3 The primer sequences used for qRT-PCR.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fu W, Huang S, Liu Z, Gao Y, Zhang M, et al. 2021. Fine mapping of Brebm6, a gene conferring the early-bolting phenotype in Chinese cabbage (Brassica rapa ssp. pekinensis). Vegetable Research 1: 7 doi: 10.48130/VR-2021-0007

Fine mapping of Brebm6, a gene conferring the early-bolting phenotype in Chinese cabbage (Brassica rapa ssp. pekinensis)
- Received: 09 February 2021
- Accepted: 24 July 2021
- Published online: 06 August 2021
Abstract: In flowering plants, bolting signals the transition from vegetative to reproductive growth, and it is consequently an important agronomic trait for the stalk-use Chinese cabbage Baicaitai. There is an urgent need to create germplasm resources for Chinese cabbage that have easy bolting characteristics. In this investigation, early bolting mutant 6 (ebm6) was created by treating Chinese cabbage double haploid (DH) 'FT' seeds with an ethyl methanesulfonate (EMS) solution. Phenotypic analysis showed that the mutant ebm6 flowered remarkably earlier than the wild-type 'FT'. Genetic analysis indicated that this early bolting trait was controlled by a recessive nuclear gene. In mutant ebm6, the causal gene BraA02g003340.3C (BrFLC2) was predicted using MutMap and Kompetitive Allele Specific PCR genotyping (KASP), as the gene encodes a MADS-box transcription factor. The 3259-bp BrFLC2 gene possessed seven exons and six introns, and in the first exon, a single nucleotide C-to-T substitution in the highly conserved MADS-box domain resulted in the premature termination of translation of the BrFLC2 protein sequence. This research supported a critical role of BrFLC2 in controlling flowering time in B. rapa. The early flowering ebm6 mutant could also be a useful material in Baicaitai breeding.
-
Key words:
- Early-bolting /
- Chinese cabbage /
- Mutmap /
- BrFLC2