










-
Diterpenes or diterpenoids, are the most complex and abundant plant metabolites, approximatly 13,000 compounds have been recorded to date[1]. Among them, over 7,000 are classified as labdane-type diterpenes[2], which usually perform significant functions in primary metabolism as basic plant growth hormones like gibberellin (GAs) and in secondary metabolism such as phytoalexins[3]. As the ancestral members of the labdane-type diterpenes[4], GAs are widely found in higher plants, and required for many aspects of plant growth and development, including seed germination, stem elongation, leaf expansion, and flower development[5−7]. GA biosynthesis in gymnosperms and angiosperms starts with the cyclization of the common diterpenoid precursor (E, E, E)-geranylgeranyl diphosphate (GGPP) by a pair of sequential diterpene synthases (diTPS), which gives rise to the formation of a diterpene skeleton[8]. The diterpene skeletons are then further oxidized by two cytochrome P450 monooxygenases and 2-oxoglutarate-dependent dioxygenases to form diverse GAs[4,9].
Ent-kaurene synthase (ent-KS) catalyses ent-copalyl diphosphate (ent-CPP) to generate ent-kaurene, which is the key step for GA biosynthesis. Based on previous phylogenetic analysis of TPSs, they were divided into seven subfamilies: TPS-a, TPS-b, TPS-c, TPS-d, TPS-e/f, TPS-g, and TPS-h, the ent-KSs from gymnosperms and angiosperms all belong to TPS-e clade[10]. Plant TPSs are classified into two groups: class I and class II, according to the reaction mechanism and products[10]. Ent-KS is a class I diterpene synthase with two conservative motifs 'DDxxD' and 'NSE/DTE', which locate at the entrance of the ligand-binding pocket and bind to the diphosphate moiety of the substrate by magnesium ions[11]. The dissociation of the pyrophosphate group and further cyclization or rearrangement of intermediate carbocations lead to the formation of tetracyclic diterpene skeleton ent-kaurene[12].
In addition to serving as the important precursor of GA biosynthesis, ent-kaurene is also associated with plant secondary metabolism[4]. Over 800 known natural diterpenoid products are derived from ent-kaurene, collectively designated as ent-kaurenoids[4]. Ent-kaurenoids extensively present in the plants of Poaceae and Lamiaceae[1,4,12−18], and most of these natural products have strong antibacterial activity such as rice diterpene phytoalexins and maize kauralexins[4,13]. Others serve as pharmaceuticals, like the vasodilator drug forskolin, which is a diterpene of Plectranthus barbatus[19].
Ilex latifolia Thunb, a species belonging to genus Ilex of family Aquifoliacea, is widely distributed in China[20]. In China and some Southeast Asian countries, leaves and buds of I. latifolia are used to make a traditional tea beverage named as 'Kudingcha' due to its bitter taste[21] (Fig. 1). Kudingcha is one of the most popular teas in China as well as teas produced using tea plant (Camellia sinensis) leaves[22]. Like tea, 'Kudingcha' also contains abundant beneficial compounds, such as terpenoids, flavonoids, polyphenols, polysaccharides and alkaloids, which have potential benefits from their anti-microbial, anti-oxidant and anti-inflammatory properties[23−25]. In addition to being consumed as tea, I. latifolia has also been used in traditional Chinese medicine for nearly 2,000 years[26]. It has been used to cure the common cold, rhinitis, itchy eyes, conjunctival congestion and headache[20]. Modern pharmacological research indicates it also has anti-obesity, anti-atherosclerotic, neuroprotective, anti-tumor and anti-diabetic effects[27−32]. Though many terpenoids have been identified as the functional components, how diterpenes are biosynthesized is unclear in I. latifolia. In this paper, we first identified a diterpene synthase gene responsible for ent-kaurene biosynthesis in I. latifolia by transcriptome sequencing, and then confirmed its biochemical function using a co-expression system in E. coli, finally detecting gene expression patterns in tissues. This study would help us to discover diterpenoids metabolism in I. latifolia, to better understand its growth and development and to better cultivate this plant for application in 'Kudingcha' tea manufacture.
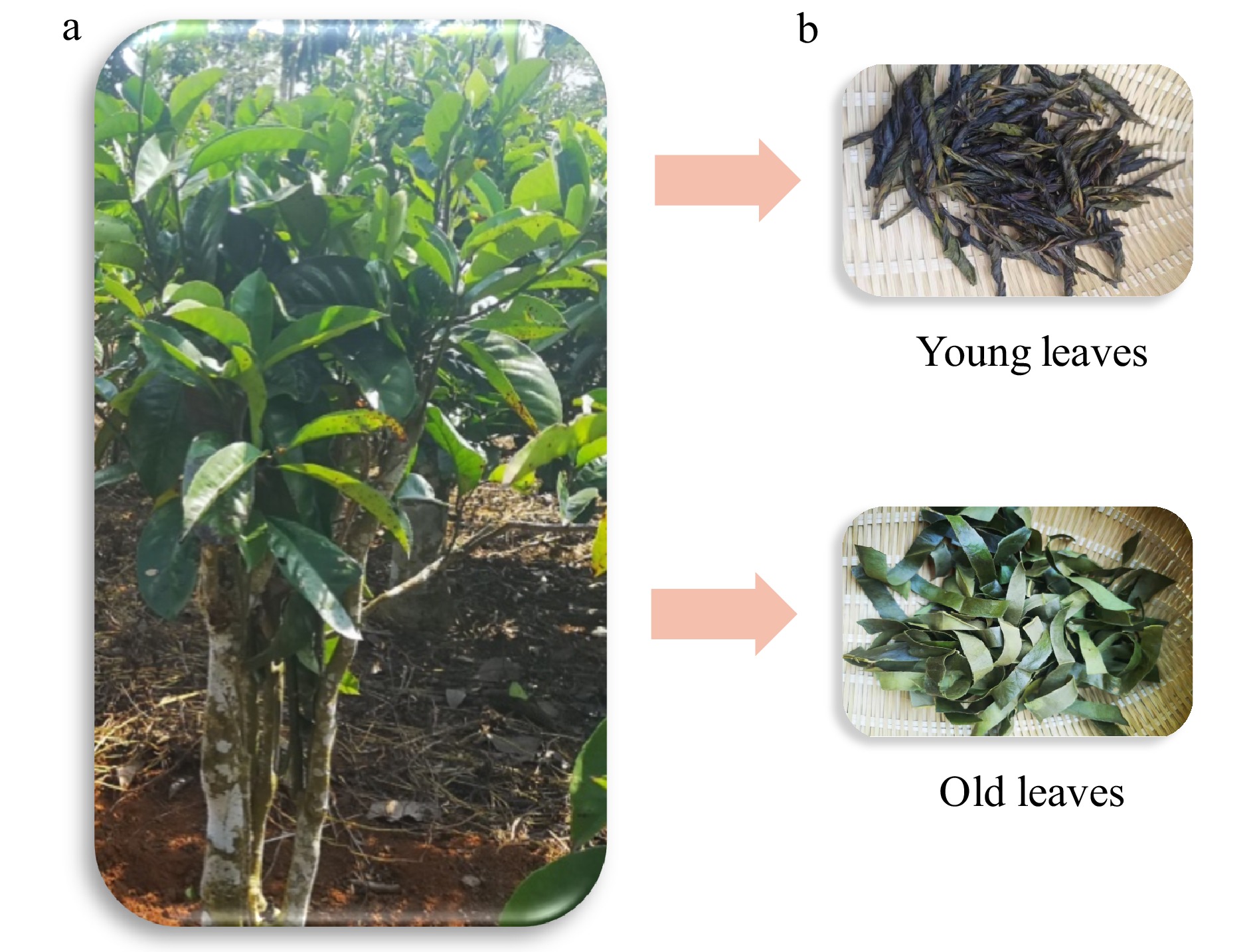
Figure 1. I. latifolia and 'Kudingcha'. (a) I. latifolia tree, (b) two types of 'Kudingcha' processed using young and old leaves.
-
After searching using two PFAM motifs: the N-terminal domain (PF01397) and the metal-binding domain (PF03936) by HMMER method, a single unigene as putative ent-kaurene synthase gene was obtained. The candidate gene of ent-KS was further confirmed by RT-PCR cloning from cDNA of leaves. We first used two primers P1 and AUP2 for full-length sequence amplification, however no product was obtained. We then ran PCR reactions with P1 and P3 for 5'-end fragment, and with P2 and AUP2 for 3'-end fragment, both of which amplified the target sequences (Supplemental Table S1). The two PCR products were diluted 100-fold and used as a template to generate the full-length sequence, which was subcloned into pMD19-T vector and thoroughly sequenced. The ent-KS gene had an ORF of 2454 bp and was identical to the unigene from transcriptome. It was deposited in GenBank and the accession number was MN696541.
Sequence analysis showed that IlKS encoded a protein of 817 amino acids. According to TargetP analysis, the protein sequence of IlKS N-terminal contained a section of transit peptides that were rich in hydrophobic amino acids like other KSs[9,33,34]. It had two conserved motifs of 'DDxxD' and 'NSE/DTE' like other typical KSs in higher plants (Fig. 2), which were involved in metal-dependent ionization of the isoprenyl diphosphate substrate. Compared with the KSs functionally annotated from other plants, IlKS shared 72% amino acid similarity with CsKS (C. sinensis ent-kaurene synthase). The phylogenetic tree also indicated that IlKS was most closely related to CsKS among 17 KSs (Fig. 3).

Figure 2. Amino acid sequence alignment of I. latifolia KS (IlKS) with representative KSs from other plants, including Scoparia dulcis KS (SdKS), Arabidopsis thaliana KS (AtKS), Oryza sativa KS (OsKS), Populus trichocarpa KS (PtTPS19). The two conserved domains (DDxxD, NSE/DTE) binding to the diphosphate moiety of the substrate by magnesium ions are indicated using red lines. Conserved sequence elements are highlighted, and the shading indicates the conservation levels.
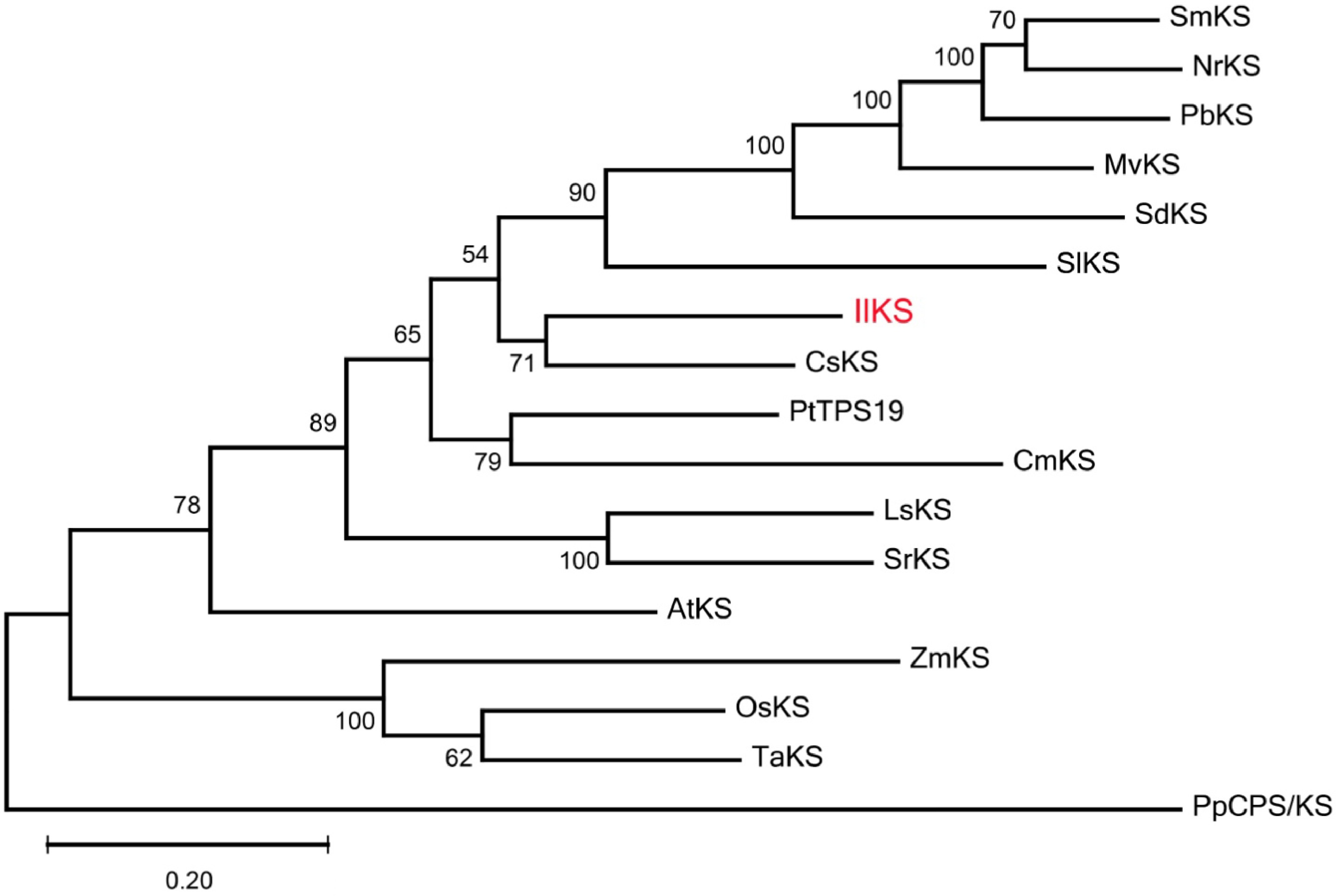
Figure 3. Phylogenetic tree of functionally characterized ent-kaurene synthases (KSs). The Maximum likelihood method was used to construct the tree with 500 bootstrap repetitions. Ent-kaurene synthase from I. latifolia (IlKS) is marked in red. The KSs from other plants used in the phylogenetic analysis are shown in Supplemental Table S2.
Functional characterization of IlKS
-
The IlKS gene after removal of transit peptide encoding region was inserted into the pET-28b vector to construct the expression vector pIlKS. After being induced, ent-kaurene was detected in the cells being co-transferred with two plasmids pSdGGSdeC and pSdKS, suggesting that the co-expression strategy in E. coli was a feasible way to test the activity of kaurene synthase (Supplemental Fig. S1). A new diterpene hydrocarbon product was detected in the cells containing pSdGGSdeC and pIlKS, while only ent-copalol was detected in the negative controls that only contained the plasmid pSdGGSdeC (Fig. 4). The mass spectrum and retention time were in accordance with the authentic standard of ent-kaurene, thus, it indicated that IlKS could convert ent-copalyl diphosphate to ent-kaurene. In addition, the trace ent-copalol in the cells containing pSdGGSdeC and pIlKS was the redundant substrate that was not catalyzed by IlKS.
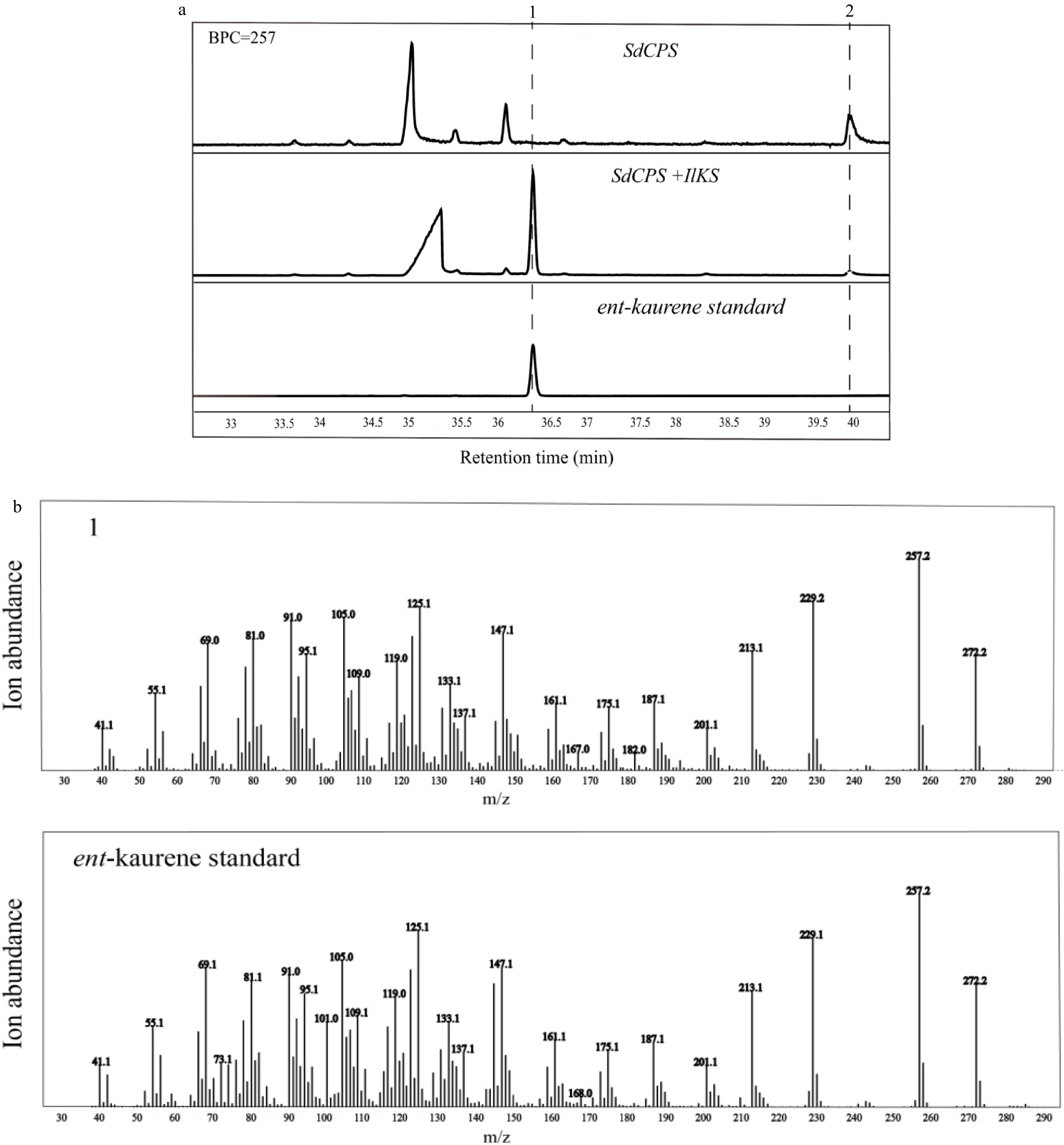
Figure 4. GC-MS analysis of diterpene products from hexane extracts of recombinant bacteria. (a) Selective ion chromatograms; (b) Mass spectra; 1, ent-kaurene; 2, ent-copalol.
Ent-KS is required for GAs biosynthesis. The ga2-1 mutant (ent-KS deletion) of A. thaliana showed a nongerminating and extreme dwarf result[6]. Generally, KS is encoded by a single copy gene in plants like Arabidopsis[10]. However, in a few plants, the enzyme is encoded by multi-copy genes. For example, the rice KS subfamily contains 11 members, but interestingly, only OsKS1 participates in GAs biosynthesis, and the rest are responsible for the biosynthesis of secondary metabolites such as diterpenoid phytoalexins[35]. In I. latifolia, KS was also encoded by only one gene. It is inferred that the labdane-type diterpenes in I. latifolia are not as abundant as those in rice, corn and other Poaceae plants[15].
Transcript levels of KS gene in Ilex latifolia tissues
-
Gene expression of IlKS was detected in all tested tissues of I. latifolia, yet the transcript levels were much different. It exhibited the highest expression level in roots, which was approximately 10-fold higher than that found in leaves. Furthermore, the expression level in stems was also higher than that of the leaves. The expression levels in roots and stems were both extremely significantly different to that of leaves (p < 0.01) (Fig. 5). This result was consistent with the observations in S. dulcis and P. trichocarpa, whose KS genes also expressed the highest level in roots[3,36]. It is generally known that GAs are mainly biosynthesized in root, so there the transcription abundance of ent-KS gene was usually higher than other tissues in plants.
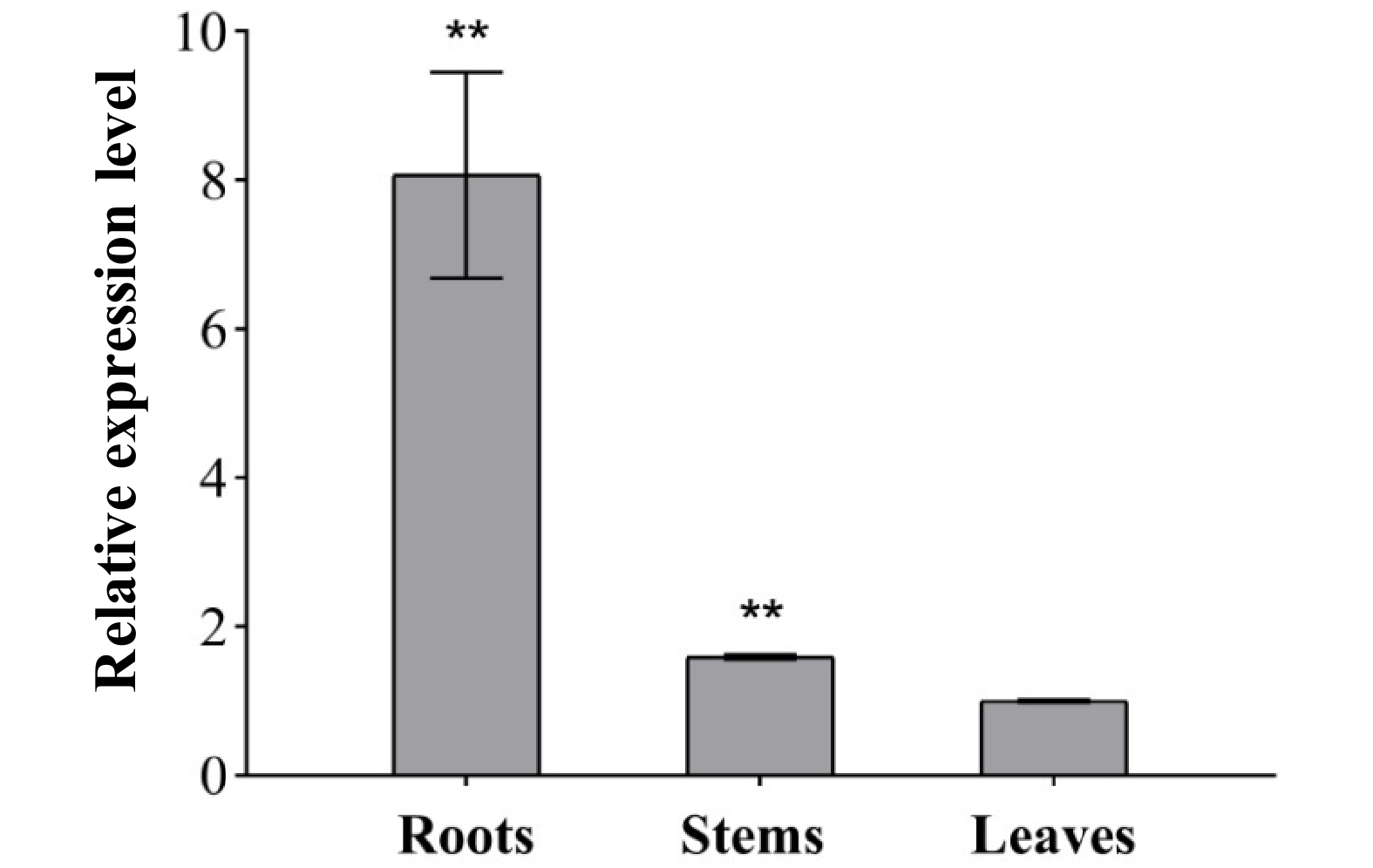
Figure 5. Relative transcript abundance of IlKS in roots, stems and leaves of I. latifolia. Transcript abundance was measured using qRT-PCR and normalized to the internal reference actin. The 2−ΔΔCT method was used to calculate the relative values. Asterisks indicate significant differences compared with the expression level in leaves. (** p < 0.01).
-
GAs is a diterpene phytohormone that plays a vital role in plant growth and development. The class I diTPS ent-kaurene synthase (KS) is the key enzyme catalyzing the cyclization reaction in GAs biosynthesis. The KS gene in I. latifolia was isolated and functionally confirmed that it could catalyze ent-CPP to form ent-kaurene. The gene exhibited the highest expression level in roots where GAs may be mainly biosynthesized. These findings shed light on further understanding GAs biosynthesis and its regulation function in I. latifolia.
-
The clonally propagated seedlings of I. latifolia Thunb were purchased from Wanchang Kuding tea farm (Hainan province, China), and planted in substrate and grown naturally in the field. Buds with two or three leaves were plucked on September 16th, 2019 and were used for transcriptome sequencing. Roots, stems and leaves were collected to examine the tissue-specific expression of IlKS. The harvested plant materials were frozen immediately in liquid nitrogen and stored at −80 ºC for subsequent experiments.
Identification of the ent-kaurene synthase gene by transcriptome sequencing
-
To identify the putative ent-kaurene synthase gene, leaves of I. latifolia were used to construct the transcriptome library, and to sequence the library using the next generation sequencing technique. Transcriptome sequencing was performed by Novogene Co., Ltd (Beijing, China) using Illumina Hiseq 2000 platform. The unigene database was searched using the N-terminal domain (PF01397) and the metal-binding domain (PF03936) by the HMMER (v3.2.1) method. After the candidate gene was identified, it was further confirmed by PCR cloning from cDNA of I. latifolia. Total RNAs were extracted using the Polysaccharide and Polyphenol Total RNA Isolation Kit (Tiangen Biotech Co., Ltd., Beijing, China) as per the manufacturer's instructions. RNA quality and concentration were assessed by a spectrophotometer (NanoDrop 2000c, Thermo Fisher Scientific, Waltham, MA, USA) and formaldehyde agarose gel electrophoresis. The first strand cDNA was synthesized by GoScript™ Reverse Transcriptase (Promega Corporation, USA) using primer AUP1, then the 3'-end sequence of IlKS gene was amplified with specific primer P2 and universal downstream joint primer AUP2 using KOD FX Neo polymerase (Toyobo Co., Ltd., Japan). The primers used for RACE are listed in Supplemental Table S1. The PCR program was set as follows: pre-denature at 94 ºC for 2 min, 35 cycles of denature at 98 ºC for 10 s, annealing at 58 °C for 30 s, extension at 68 ºC for 2 min, and a final extension at 72 ºC for 10 min. The PCR product was purified by DNA gel extraction (Takara MiniBEST Agarose Gel DNA Extraction Kit Ver 4.0, Dalian, China) and then cloned into pMD19-Tvector (Takara pMDTM19-T Vector Cloning Kit, Dalian, China) for sequencing (Tsingke, Hangzhou, China).
Database research and multiple sequences alignment
-
The encoding protein sequence of IlKS gene was obtained by ExPASy online (http://web.expasy.org/translate). Multiple sequences of other plant KSs were retrieved by Blastp from NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi), and the detailed information of all KSs are listed in Supplemental Table S2. Sequence alignment was carried out using the ClustalW program embedded in MAGE 6.0[37], and the phylogenetic tree was also constructed by MAGE 6.0 based on the Maximum likelihood (ML) method and Poisson correction model. The signal peptide and transmembrane region of KS were analyzed using the SignalP 4.1 Server (http://www.cbs.dtu.dk/services/SignalP) and TMHMM program online (http://www.cbs.dtu.dk/services/TMHMM), respectively. The transit peptide sequence was predicted using TargetP (http://www.cbs.dtu.dk/services/TargetP).
Protein expression and enzymatic activity determination of IlKS
-
The RNA and cDNA were prepared as described above. We constructed a co-expression strategy in E. coli to test the activity of kaurene synthase. As E. coli didn't typically produce GGPP, it was necessary to introduce a geranylgeranyl diphosphate synthase gene (SdGGPPS, S. dulcis, AB034250) into E. coli that could offer the substrate GGPP for ent-copalyl disphosphate synthase (CPS)[38]. Secondly, an ent-CPS (SdCPS, S. dulcis, AB169981) was also required to produce ent-CPP as the substrate for KS. Thus, they were assembled into a dual gene expression vector pACYA-Duet as SdGGPPS in multiple cloning site 2 (MCS2) and SdCPS in multiple cloning site 1 (MCS1)[36]. Finally, the IlKS without transit sequence was cloned using a pair of primers IlKS-F1 and IlKS-R2 (Supplemental Table S1) and digested with two restriction enzymes BamHI and NotI, and then was subcloned into the pET-28b vector. The ent-kaurene synthase from S. dulcis (SdKS, AEF33360) was also ligated into pET-28b to generate a pSdKS plasmid as the positive control.
The plasmids pIlKS carrying IlKS gene with pSdGGSdeC containing SdGGPPS and SdCPS were co-transferred into E. coli (strain C43) by heat shock at 42 ºC for 1 min. Then the cells were injected into 1mL terrific broth (TB) medium supplemented with 1% glucose and shaking for 24 h at 37 °C, 220 rpm. The following day, the cell culture fluid was inoculated into fresh 50 mL TB liquid medium and cultured under the same conditions. When the OD value at 600 nm reached 0.4, the incubation temperature was shifted down to 16 ºC and shaking for 1 h. The final concentration of 0.5 mM of isopropyl-β-D-thiogalactopyranoside (IPTG, 50 mg/mL, Tiangen, Ltd., Beijing, China) and 1 mM MgCl2 were added to the culture, followed by continued incubation with shaking at 16 °C for 72 h. After incubation, the induced cultures were extracted with equal volume of n-hexane twice, and the micelles were disintegrated by EtOH. The supernatant was collected and concentrated to 1 mL by N2-blowing. The n-hexane extracts were analyzed using a gas chromatograph mass spectrometer (GC-MS, Agilent 7890B-7000C, Agilent Technologies. Inc. California, USA) using a HP-5MS (30 m × 0.25 mm, 1909IS-433UI, Agilent Technologies. Inc. California, USA). Each sample (2 μL) was injected at 250 °C in a splitless mode. After holding the samples for 3 min at 50 °C, the oven temperature was increased at 5 °C per min to 250 °C and held for 5 min. The flow rate of helium carrier gas was set at 1.9 mL/min. All MS data was collected from 40 to 400 m/z[39].
Tissue-specific gene expression profile of IlKS
-
Total RNAs were extracted from roots, stems and leaves of I. latifolia. The cDNAs were obtained as described above and then diluted to 200 ng/μL as the templates. Real-time PCR was carried out using TB GreenTM Premix Ex TaqTM (TliR NaseH Plus) (TaKaRa Bio Inc) in a Roche Light Cycler 480-II real-time thermal cycler (Roche, Switzerland). The Actin gene identified from the transcriptome was used for normalization. The primers for qRT-PCR were shown in Supplemental Table S1 and Actin gene identified from the transcriptome was used as the internal control. Data for the relative quantity was calculated using the 2−ΔΔCT method[40].
- This work was supported by the National Natural Science Foundation of China (31470693, 31100503), the Natural Science Foundation of Zhejiang Province (LY18C160006), the Central Public-interest Scientific Institution Basal Research Fund (1610212018004, 1610212016017), the Science and Technology Innovation Project of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP-2014-TRICAAS), and the Key Project of International Science & Technology Cooperation, National Key Research and Development Program of China (2017YFE0107500). We thank Dr. Xiaoling Gu for collecting Ilex latifolia samples and taking pictures.
- The authors declare that they have no conflict of interest.
- Supplemental Table S1 The primers used in this study.
- Supplemental Table S2 The detail information of KSs used in phylogenetic analysis .
- Supplemental Fig. S1 The biochemical activity of SdKS. A, Selective ion chromatograms (BPC = 257); B, Mass spectra; 1, ent-kaurene; 2, ent-copalol.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yang M, Liu G, Yamamura Y, Fu J. 2021. Identification and functional characterization of ent-kaurene synthase gene in Ilex latifolia. Beverage Plant Research 1: 7 doi: 10.48130/BPR-2021-0007

Identification and functional characterization of ent-kaurene synthase gene in Ilex latifolia
- Received: 10 June 2021
- Accepted: 24 August 2021
- Published online: 30 August 2021
Abstract: Diterpenes are the most complex and abundant plant metabolites, some of which play significant roles in both primary and secondary metabolism. Ent-kaurene synthase is the key enzyme for gibberellins (GAs) biosynthesis, which may act as a catalyst in the formation of ent-kaurene, the precursor for GAs, by cyclization or rearrangement of the substrate ent-copalyl diphosphate (ent-CPP). Ilex latifolia Thunb (Aquifoliaceae) is a widely distributed Chinese plant whose leaves are used to process a popular Chinese bitter tea named 'Kudingcha', which has anti-microbial, anti-oxidant and anti-inflammatory properties. Here, we isolated a diterpene synthase gene from the leaf transcriptome of I. latifolia, and further identified its biosynthesis activity by in vitro enzymatic testing. The heterologous expressed ent-kaurene synthase of I. latifolia (IlKS) in E. coli could catalyze ent-copalyl diphosphate (ent-CPP) to form ent-kaurene. Tissue-specific expression indicated that IlKS had the highest transcript level in roots which is maybe the major location of ent-kaurene biosynthesis. This study would help us to determine diterpenoid metabolism and GAs biosynthesis in I. latifolia, to better understand the regulation function of GAs in growth and development.
-
Key words:
- Ilex latifolia /
- ent-kaurene synthase /
- KS /
- gibberellin biosynthesis