









-
Tea, made from young shoots of Camellia sinensis(L.) O. Kuntze
, is one of the oldest and most widely consumed beverages in the world[1, 2]. Catechins are one of the three major characteristic-related secondary metabolites in tea leaves, which can comprise 12%−25% of the dry tea weight[3−6]. The accumulation of catechins in tea leaves significantly determines the final flavor characteristics, mouthfeel and health benefits of tea products[7−12]. For green tea, catechins are the main bitter- and astringent-tasting contributors of tea infusion[8, 9, 13]. After rolling, fermentation and piling, catechins can form theaflavin and thearubigins through a range of complicated biochemical reactions to determine the taste, color, and flavor of black or dark teas[14−16]. Previous studies have shown that environmental factors, including light, temperature and abiotic stresses can regulate the accumulation of catechins in fresh tea leaves[17−20]. Light is one of the most important environmental parameters that can regulate catechin accumulation in tea leaves[16, 21−24]. It has been well documented that shading, as a traditional practice has been commonly used in tea plantations, can effectively improve the quality of fresh leaves through the regulation of carbon and nitrogen metabolism and enhancement of chlorophyll accumulation for green tea production[25−27]. For Gyokuro and Matcha, shading before leaf picking is an essential process to reduce the phenol-ammonia ratio of tea leaves, and improve the freshness and umami taste of tea while reducing the bitterness and astringency[13, 28]. In addition to light intensity, different light wavelengths also have a significant regulatory effect on catechins in tea leaves[22, 29−31]. Previous studies showed that red and blue light can up-regulate the level of volatile terpenoids, and different light wavelengths can stimulate the expression of volatile substance synthesis genes in tea leaves[22, 30]. Ultraviolet light can regulate the biosynthesis of flavonoids in fresh tea leaves, therefore potentially affect the final flavor characteristics of tea products[24, 29, 32]. Furthermore, the Chinese chestnut-tea intercropping system can up-regulate the metabolite of green tea to improve the quality and the health effect of tea[33, 34]. Metabolic changes in flavonoids, amino acids and carbohydrates in tea leaves were potentially caused by the shading effects of chestnut trees[33, 34].
For tea production, years of experience and practices from tea farmers suggest that fresh leaves manually picked in the morning after dew evaporation makes the best tea with increased freshness and sweetness. In contrast, tea made of leaves picked in the afternoon with strong light exposure and relatively high temperature presents obvious bitterness and astringency. This indicates the diurnal environment potentially plays a role in metabolism regulation in fresh tea leaves, and these circadian rhythm effects directly lead to flavor changes of the final tea products. The diurnal environmental parameters in tea plantations such as light and temperature, as external regulators, together with the internal developmental regulation participate in the regulation of flavor-related metabolites in tea leaves[18, 35−37]. However, few studies have focused on the effects of circadian rhythm on major secondary metabolites in fresh tea leaves, and its potential effects on the final characteristic of tea products are unclear.
This study aims to analyze the effects of circadian rhythm on catechin accumulation patterns in tender leaves of three tea cultivars with different phenotypes, 'Huangkui' ('HK'; light-sensitive etiolated cultivar)[38], 'Longjing43' ('LJ43'; green cultivar, small leaf size) and 'Fuding Dabaicha' ('FD'; green cultivar, medium leaf size). Total catechin content and major individual catechins in tender tea leaves were measured during a 48-h-period using high-performance liquid chromatography (HPLC) to explore the effects of circadian rhythm on catechin accumulation in tender leaves of tea plants. These findings provide new insights into our understanding of circadian rhythm regulation on the flavor-important secondary metabolites in tea leaves, which also provide suggestions for leaf harvesting in tea production.
-
'HK', 'LJ43' and 'FD' cultivars were used to analyze the circadian rhythm effects on catechin accumulation in this study. 'HK' which is suggested to be a light-sensitive albino cultivar with etiolated tender leaves and shoots with the onset of budburst was used as a representative of albino tea cultivars in this study[38]. 'LJ43' was selected to present ordinary green tea cultivars with small leaves, while 'FD' was used as a representative for green tea cultivars with medium leaf size. The environmental parameters and chlorophylls were measured (Fig. 1 & 2). In 2017, the second tender leaves were collected every 2 h during a 48-h-period from the 16th to the 18th of September when a new round of bud burst occurred (Fig. 2a). Consistent with their phenotypes, 'FD' leaves showed the highest chlorophyll accumulation (Chl a and Chl b), followed by 'LJ43' and then 'HK' (Fig. 2b). The ratio of Chl a and Chl b was also analyzed among the three tea cultivars (Fig. 2c). The ratio of Chl a and Chl b in 'HK' leaves was 5.45, and was significantly higher than that in 'LJ43' (3.50) and 'FD' (3.32).
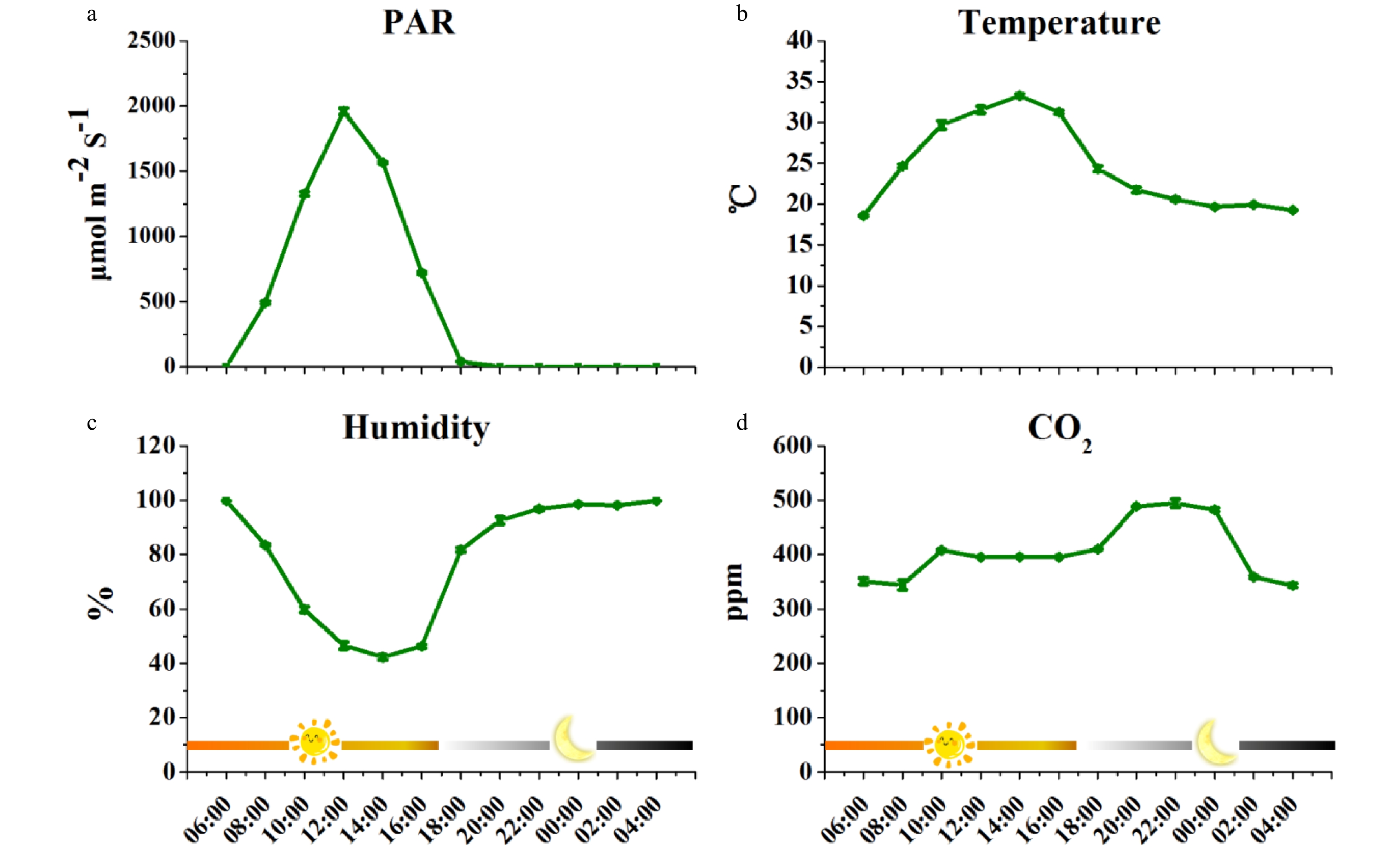
Figure 1. The changes of environmental factors during the sampling period in tea plantation. (a) PAR changes during the sampling period; (b) Temperature changes during the sampling period; (c) Humidity changes during the sampling period. (d) CO2 changes during the sampling period. Data were successively collected every 2 h during the 48-h-sampling period from the 16th to the 18th of September 2017 (n = 6). PAR, Photosynthetic Actively Radiation.
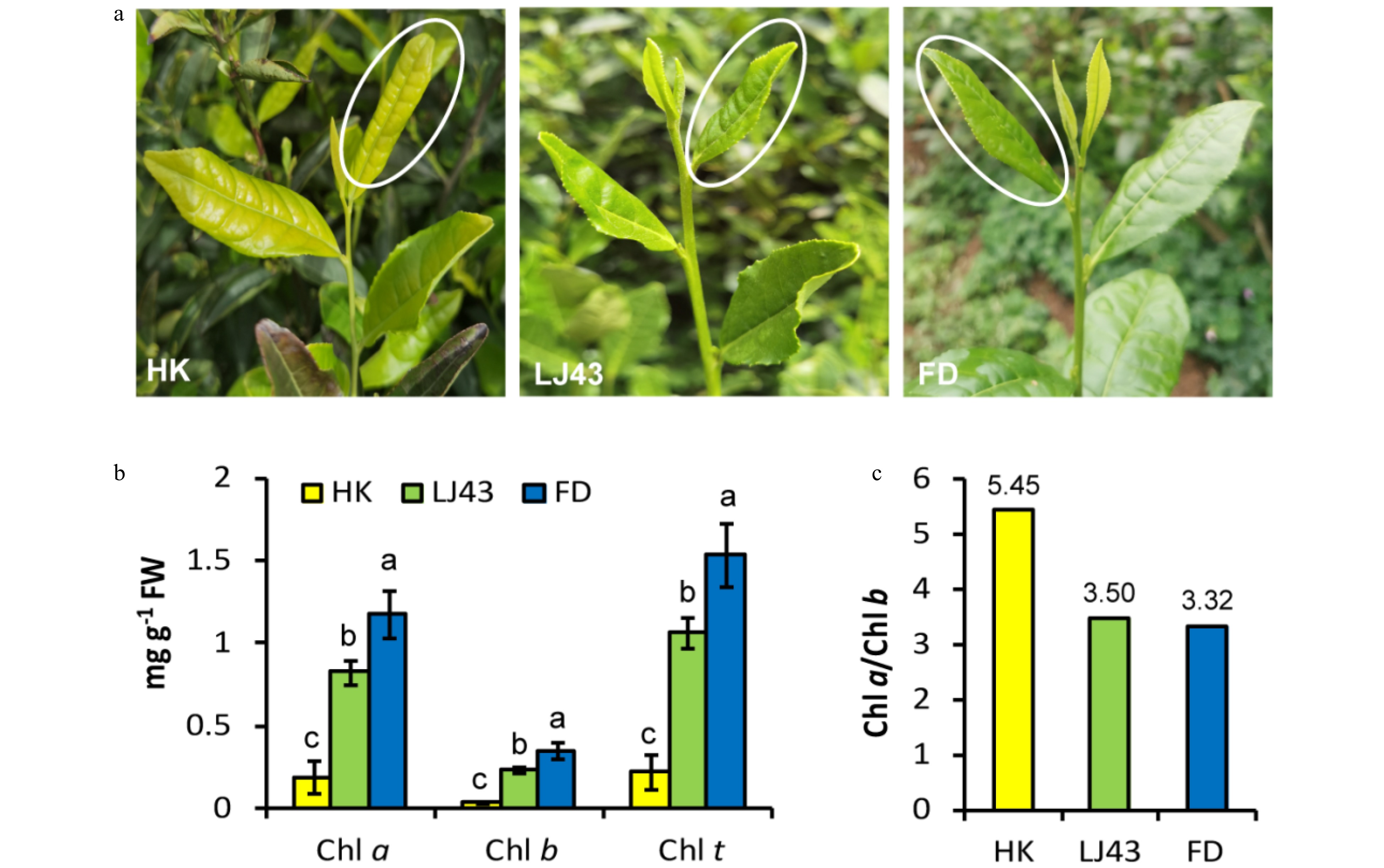
Figure 2. Phenotypes and chlorophyll content in tender leaves of three tea cultivars. (a) Tender leaves of three tea cultivars were used in this study; (b) Chlorophyll content in tender leaves of three tea cultivars; (c) The ratio of Chl a to Chl b in tender leaves of three tea cultivars. 'HK', 'Huangkui'; 'LJ43', 'Longjing43'; 'FD', 'Fuding Dabaicha'; Chl a, chlorophyll a; Chl b, chlorophyll b; Chl t, total chlorophylls. Error bars represent the SE (n = 3). Letters (a, b, c) show significant differences and grouping information according to one-way ANOVA and a Fisher's LSD test at the 5% level.
Total catechin accumulation pattern in three tea cultivars
-
To analyze the effects of circadian rhythm on catechin accumu lation in tea plants, catechins were extracted from second tender leaves of 'HK', 'LJ43' and 'FD' (Fig. 3). According to Principal Component Analysis (PCA), catechins from different tea cultivars were classed into three groups. This indicated a cultivar-specific regulation on catechin accumulation in tender tea leaves (Fig. 3a).
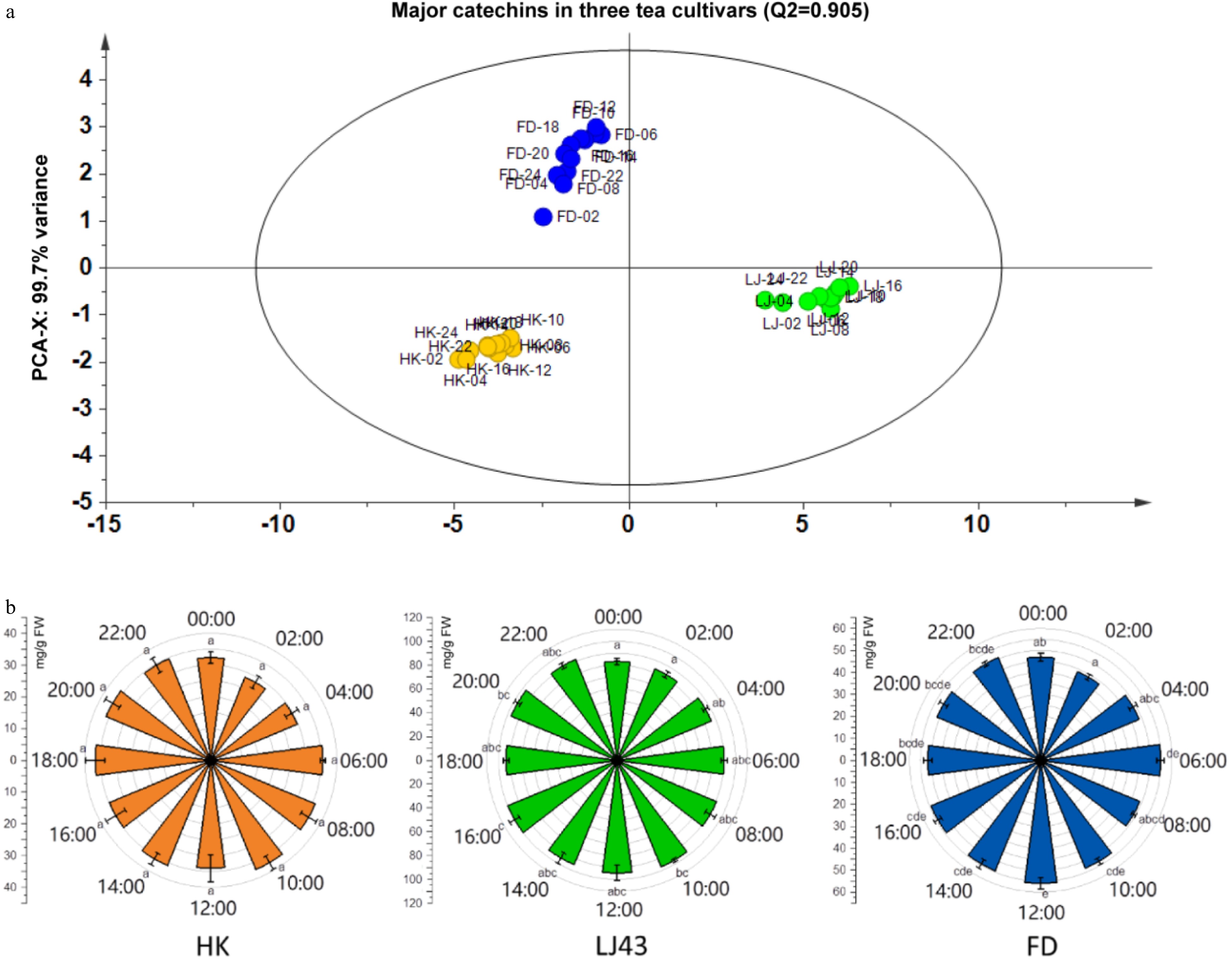
Figure 3. Cultivar-specific regulation and circadian rhythm effects on total catechins in tea plants. (a) PCA analysis of catechins in tender leaves of three tea cultivars; (b) The effects of flavonoids in tender leaves among three cultivars. 'HK', 'Huangkui'; 'LJ43', 'Longjing43'; 'FD', 'Fuding Dabaicha'. Error bars represent the standard error (n = 6). Letters (a, b, c, d, e) show significant differences and grouping information according to one-way ANOVA and a Fisher's LSD test at the 5% level.
The content of total catechins presented a circadian rhythm regulation in tender leaves of three tea cultivars (Fig. 3b). In total, relatively higher contents of catechins were detected during the daytime (from 06:00 to 18:00) when compared to that during the night. A clear pattern was observed in etiolated leaves of 'HK', compared with that in green leaves of 'LJ43' and 'FD' cultivars. Among all of the time points for sample collection, the lowest content of catechins were detected at 00:00−02:00 while the highest content of catechins were found in leaves at around 16:00 in all cultivars.
Individual catechin accumulation pattern in three tea cultivars
-
To investigate the effects of circadian rhythm on individual catechins in tea plants, major catechins which were highly accumulated in tea leaves of 'HK', 'LJ43' and 'FD' were analyzed using HPLC. These catechins included three non-gallated catechins including catechin (C), epicatechin (EC), gallocatechin (GC), epigallocatechin (EGC), as well as three gallated catechins including gallocatechin gallate (GCG), epicatechin gallate (ECG) and epigallocatechin gallate (EGCG) (Fig. 4−6).
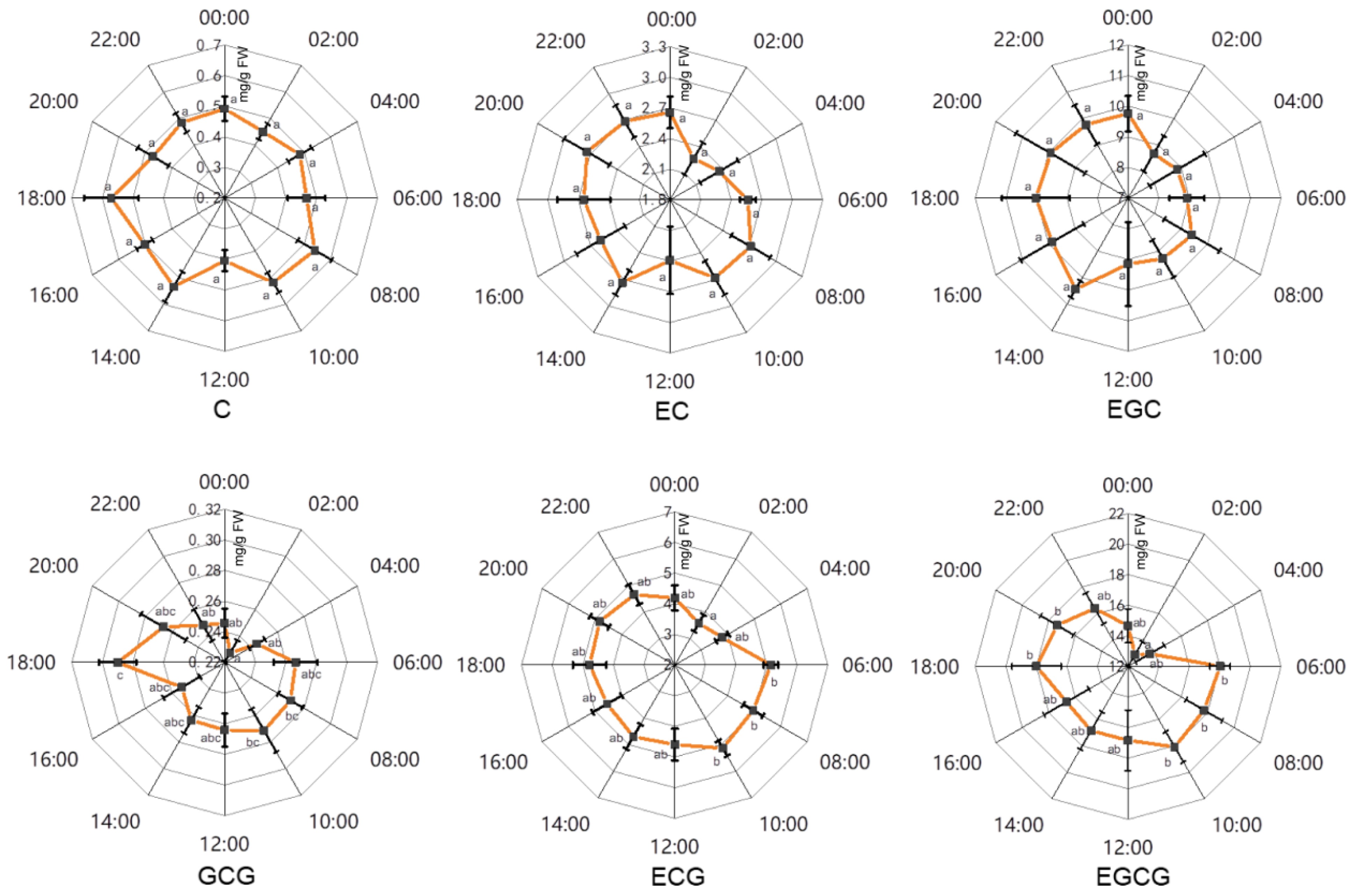
Figure 4. The effects of circadian rhythm on major catechins in tender leaves of 'HK'. C, catechin; EC, epicatechin; EGC, epigallocatechin; GCG, gallocatechin 3-O-gallate; ECG, epicatechin 3-O-gallate; EGCG, epigallocatechin 3-O-gallate. Error bars represent the standard error (n = 6). Letters (a, b, c) show significant differences and grouping information according to one-way ANOVA and a Fisher's LSD test at the 5% level.
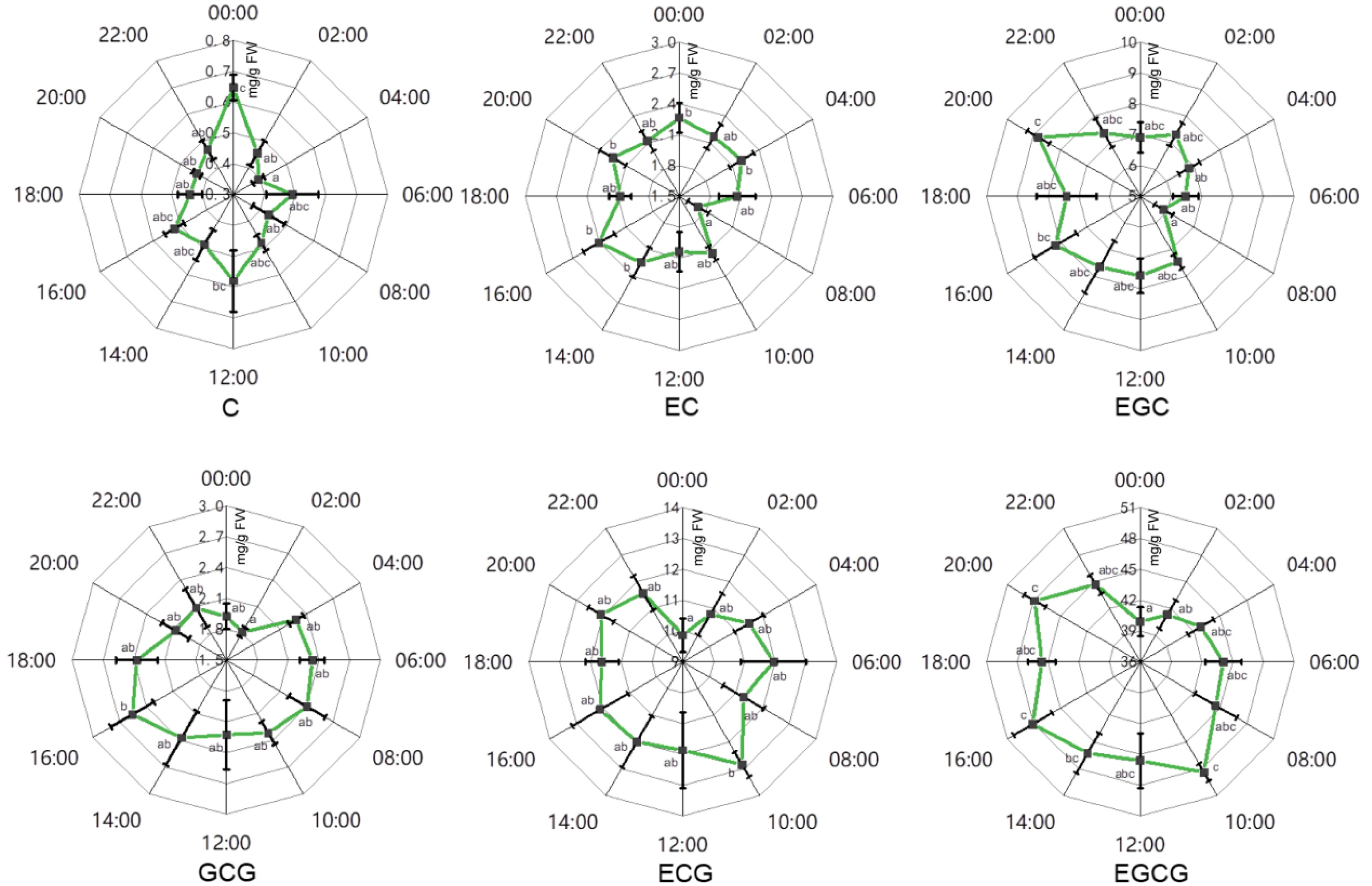
Figure 5. The effects of circadian rhythm on major catechins in tender leaves of 'LJ43'. C, catechin; EC, epicatechin; EGC, epigallocatechin; GCG, gallocatechin 3-O-gallate; ECG, epicatechin 3-O-gallate; EGCG, epigallocatechin 3-O-gallate. Error bars represent the standard error (n = 6). Letters (a, b, c) show significant differences and grouping information according to one-way ANOVA and a Fisher's LSD test at the 5% level.
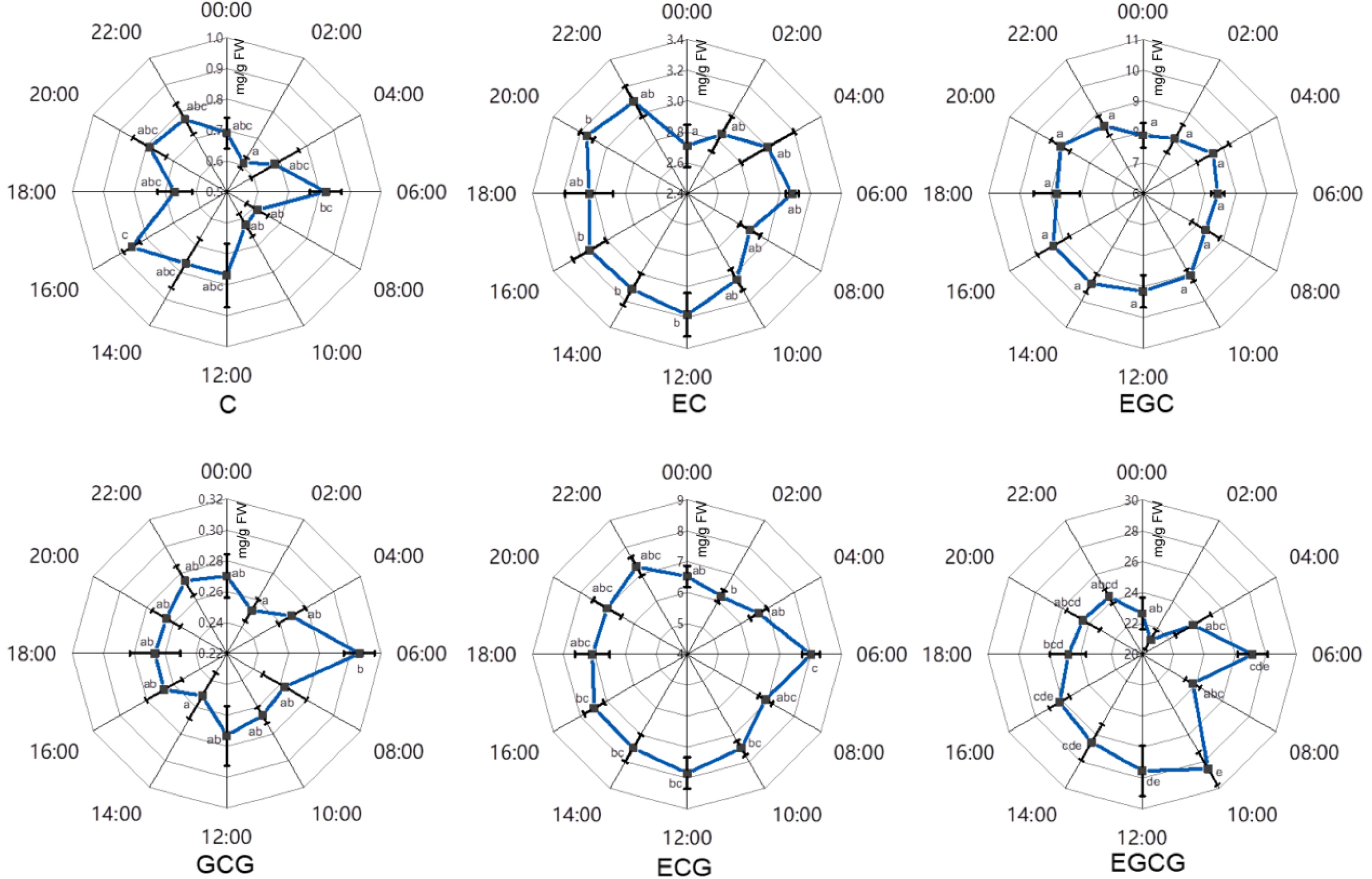
Figure 6. The effects of circadian rhythm on major catechins in tender leaves of 'FD'. C, catechin; EC, epicatechin; EGC, epigallocatechin; GCG, gallocatechin 3-O-gallate; ECG, epicatechin 3-O-gallate; EGCG, epigallocatechin 3-O-gallate. Error bars represent the standard error (n = 6). Letters (a, b, c, d, e) show significant differences and grouping information according to one-way ANOVA and a Fisher's LSD test at the 5% level.
In etiolated tender leaves of 'HK' (Fig. 4), an accumulation pattern was detected, with relatively lower content of catechins found at around 00:00−02:00 and higher content found during the daytime, especially for the gallated catechins (ECG, GCG and EGCG). Compared with the gallated catechins (ECG, GCG and EGCG), non-gallated catechins including EGC, C and EC showed a slight decrease at noon.
Consistent accumulation patterns were detected in tender leaves of 'LJ43' (Fig. 5). Catechins, especially gallated catechins presented higher content during the daytime when compared to nighttime. The lowest catechin content was detected at around 00:00−02:00, whilst higher content was detected in the afternoon around 16:00. Non-gallated catechins also showed a temporary decrease at around 8:00.
As showed in Fig. 6, 'FD' leaves contained relatively higher content of catechins during the daytime for EC, EGC, ECG and EGCG when compared with during the night. Also, temporary decreases were detected during the 00:00−02:00 period compared with other time points, especially for the contents of C, EC, GCG and EGCG.
Catechin composition in three tea cultivars
-
To analyze the effects of circadian rhythm on catechins in more detail, the composition of catechins was measured in tender leaves of 'LJ43' and 'FD' (Fig. 7). Based on the accumulation patterns of catechins shown earlier in the text, catechin composition at 02:00 and 16:00 were selected to present the composition changes of catechins in different cultivars during night and daytime, respectively. Clear differences were detected among the three cultivars, with the highest percentage of EGCG content recorded in 'LJ43' (≥ 64%) when compared to 'HK' (≥ 45%) and 'FD' (≥ 48%). In contrast, 'LJ43' presented the lowest percentage of EGC (around 11%) in comparison to that in 'HK' (≥ 29%) and 'FD' (≥ 17%). The highest percentage of GC was spotted in 'FD' (≥ 10%), while the same in 'HK' and 'FD' leaves was only around 1%.
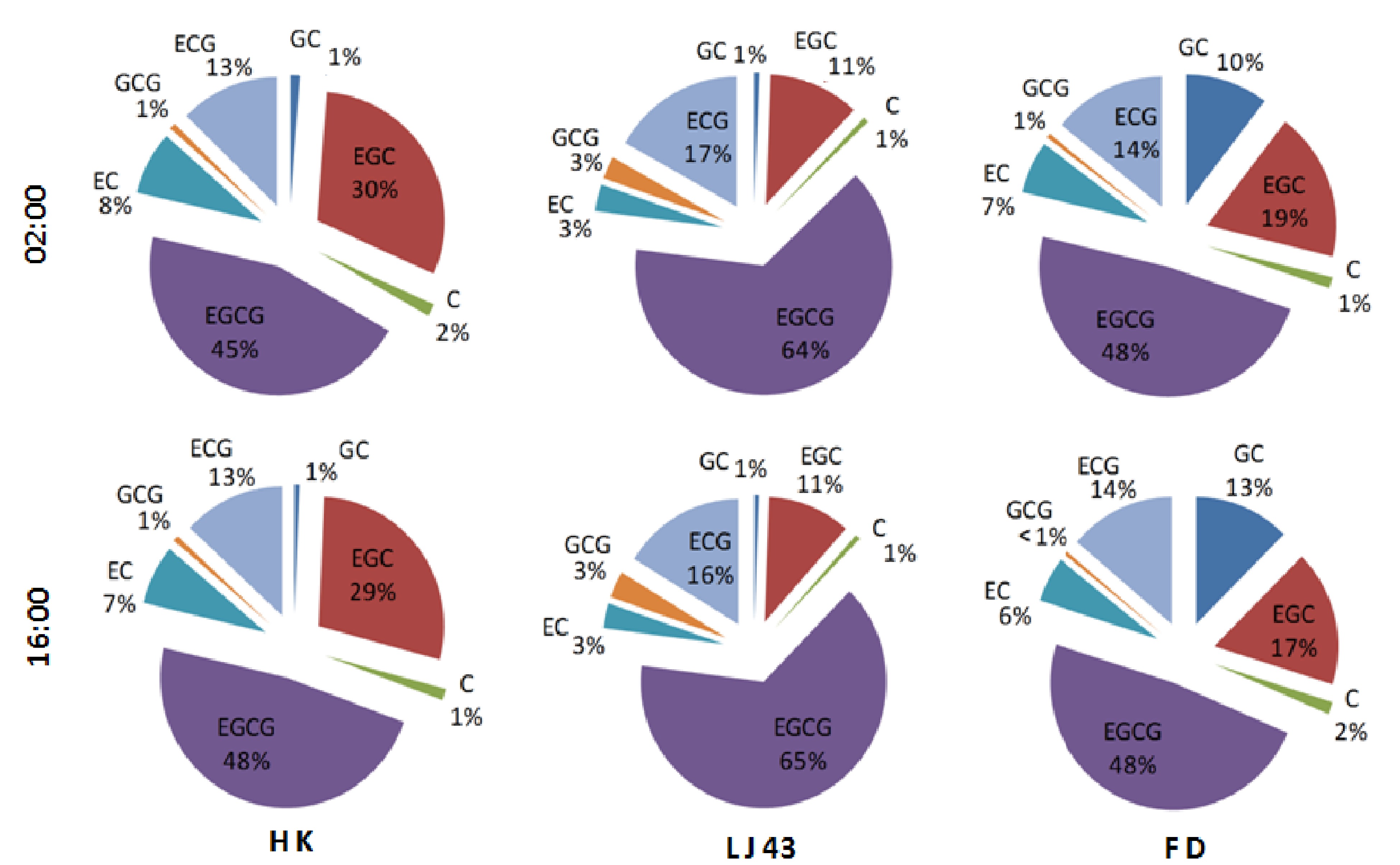
Figure 7. The composition of catechins in tender leaves at 02:00 and 16:00 among three tea cultivars. 'HK', 'Huangkui'; 'LJ43', 'Longjing43'; 'FD', 'Fuding Dabaicha'. ECG, epicatechin 3-O-gallate; EGC, epigallocatechin; C, catechin; GCG, gallocatechin 3-O-gallate; EC, epicatechin; EGCG, epigallocatechin 3-O-gallate.
In addition to cultivar-specific regulation on catechin composition, effects of circadian rhythm were also detected. Compared with leaves sampled at 02:00, higher levels of EGCG were found at 16:00 in both 'HK' and 'LJ43' cultivars. In contrast, decreases were detected in 'HK' and 'FD' with compositions of EC and EGC at 02:00 compared with that at 16:00. The compositions for GCG and ECG did not show significant differences between 02:00 and 16:00.
Correlation between catechin accumulation and environmental factors
-
To explore the effects of environmental factors on catechin accumulation in tea leaves, the correlation between catechins and major environmental factors was analyzed (Fig. 8). The PAR and temperature were positively correlated with most catechins, with the exception of GC content. In contrast, air humidity was negatively correlated with catechins but was positive with GC. Furthermore, positive correlations were detected between CO2 and C, GC, EC, EGC and EGCG, whereas a negative correlation was found between CO2 and GCG.
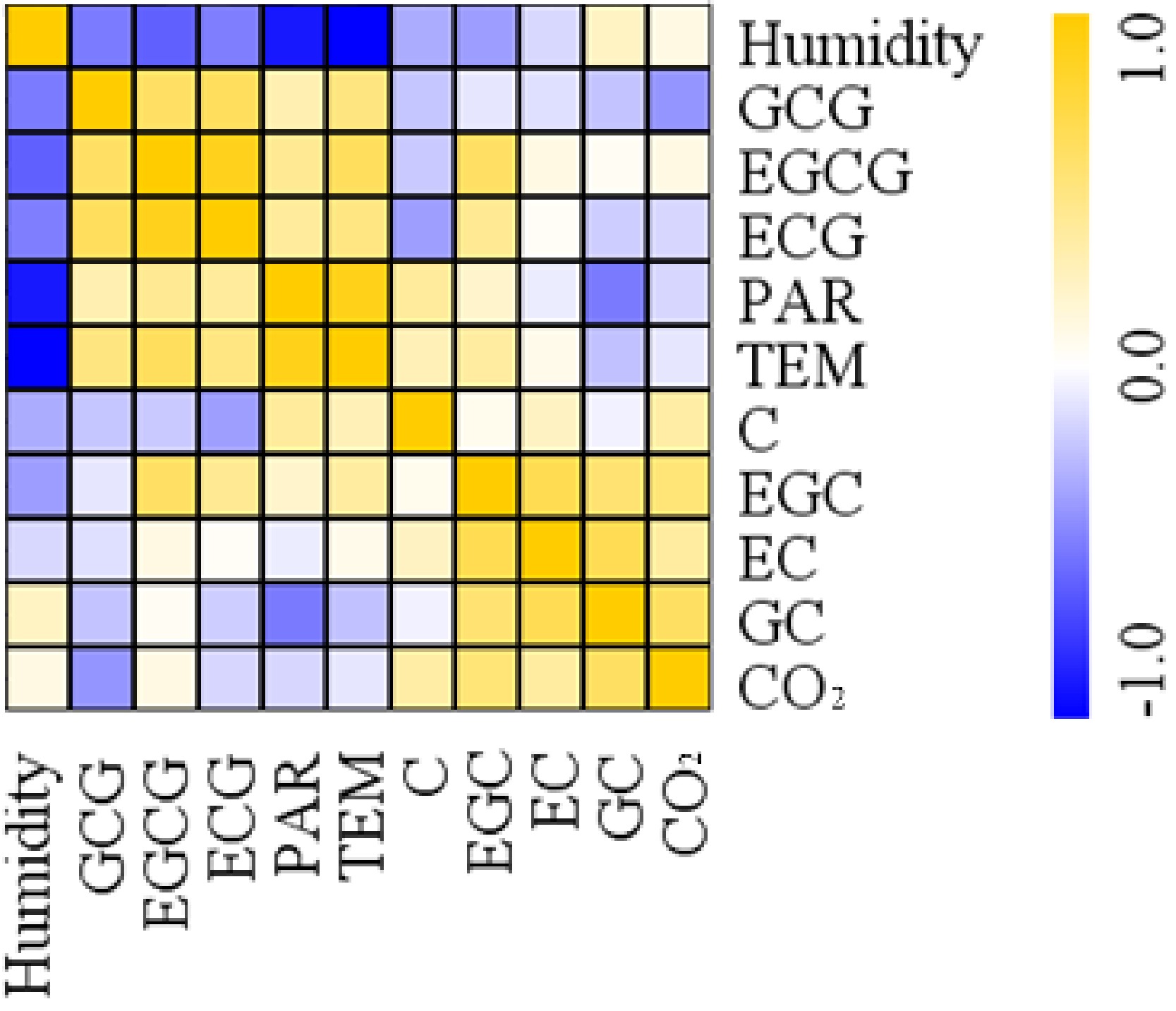
Figure 8. Correlation analysis between catechins and environmental factors. EGC, epigallocatechin; C, catechin; EC, epicatechin; GC, gallocatechin; ECG, epicatechin 3-O-gallate; GCG, gallocatechin 3-O-gallate; EGCG, epigallocatechin 3-O-gallate; PAR, Photosynthetic Actively Radiation; TEM, temperature.
Data availability
-
Original data in this study are available from Liulinlin@ahau.edu.cn.
-
Catechins are important bitter- and astringent-tasting substances for tea flavor characteristics which also contribute to the health benefits of tea products[8, 10]. Although many studies have shown that catechins are light-sensitive and can be regulated by the changes of both light intensity and light quality including shading decreasing light accumulation in tea plantation[23, 26, 31, 37], red/blue/green/UV light wavelengths modulated the composition of catechins in tea leaves[22, 24, 29, 30, 32]. This study, however, is the first study in tea plants (C. sinensis) that investigates the effects of circadian rhythm on catechins among different tea cultivars with regards to various environmental factors in tea plantations.
There are two factors involved in catechin accumulation in tea plants that were tested: a cultivar-specific regulation of catechin accumulation, and temporal modulation of day-night rhythm as an environmental aspect of regulation including light, temperature and humidity. In both total content and composition of catechins, a clear cultivar-specific difference was found in tender leaves among three tea cultivars (Fig. 3 & 7). Compared with the ordinary green tea cultivars 'LJ43' and 'FD', the etiolated tender leaves of 'HK' contained significantly lower content of chlorophylls and catechins (Fig. 2 & 3). These results are consistent with previous studies that showed relatively lower content of catechins compared to a higher accumulation of free amino acids found in the albino/etiolated leaves of albino tea cultivars, which resulted from the re-adjustment of the carbon and nitrogen mechanism according to their deficient chloroplast development and chlorophyll biosynthesis[38−43]. A previous study reported that the accumulation patterns of catechins and flavonol glycosides were mainly dependent on the genetic background of the cultivars rather than growth environments[44]. Consistent results were found in the composition of catechins in this study, the effects of circadian rhythm on catechin composition in tea leaves were negligible when compared with the cultivar-specific regulation (Fig. 7). A clear cultivar-specific difference was also found in catechin composition among tender leaves of the three tea cultivars. A higher composition of EGCG was detected in 'LJ43' (64%), compared with that in 'HK' and 'FD' leaves (about 48%; Fig. 7).
A circadian rhythm regulation was also detected in the catechin accumulation in tender leaves of the three tea cultivars. Both the total and major individual catechins showed a higher accumulation during the daytime (06:00-18:00) when compared with the night, especially during the period of 00:00-02:00 (Fig. 3−6). These results were predictable in that catechins, as products of carbon assimilation in tea leaves, accumulated significantly during the daytime due to high light exposure and active photosynthesis of tea plants[23, 37]. In contrast, respiration (presented as CO2) took place instead of photosynthesis during the night leading to a decrease of carbon (catechins) accumulation in tea leaves. These findings indicated that early morning, when the contents of bitter- and astringent-tasting catechins were relatively low, might be the best leaf plucking time for green tea making. In contrast, for the black or dark tea industry, fresh leaves which require higher contents of flavonoids to survive the fermentation process to form strong tastes and rich flavors, could be harvested during the late afternoon. Interestingly, previous studies have shown that the non-gallated catechins (C, EC and EGC) are light/UV-responsive when compared with the gallated catechins (GCG, ECG and EGCG) in tea leaves[21, 23, 26, 29]. However, in this study non-gallated catechins did not present more significant effects of circadian rhythm when compared with gallated catechins in tea leaves of the three cultivars (Fig. 4−6).
According to correlation analysis among major catechins and environmental factors, both light and temperature showed high positive correlation with catechin accumulation, whereas humidity presented the opposite pattern (Fig. 8). We presume that the changes in catechin accumulation were caused by a combined regulatory mechanism of both light and temperature. There is no doubt that light plays an essential role in regulating catechin biosynthesis in tea leaves[23, 26, 29]. A previous study of shading effects on tea plants has shown that lower temperature decreased flavonols and their glycosides but increased accumulation of flavan-3-ols and proanthocyanidins, and temperature could have a greater effect on the galloylation of catechins than light intensity[18]. Further studies are needed on the molecular regulatory mechanism of temperature on secondary metabolites in tea plants.
-
Catechins are major bitter- and astringent-tasting secondary metabolites in tea leaves, which are light-responsive and important contributors to tea flavor characteristics and health benefits. To date, the effects of circadian rhythm on catechin accumulation patterns are poorly understood. Here we reported that temporal effects of the circadian rhythm were involved in the catechin regulation in tea plants. Compared with the cultivar-specific regulation on catechin accumulation, the changes in catechin formation patterns caused by environmental factors were conserved across three tea cultivars. As the first report on circadian rhythm effects on catechin accumulation in tea plants, we believe these findings provide new insights into our understanding of circadian rhythm regulation on the flavor-important secondary metabolites in tea leaves and also contribute to leaf plucking and processing improvements for the tea industry.
-
Samples were collected from the research tea plantation of Anhui Agricultural University (N31°.55', E117°.12', +24m; Tea Plant Cultivar and Germplasm Resource Garden, Anhui Province, China)[21]. Three vegetatively propagated 10-year old tea cultivars, including 'Huangkui' ('HK', etiolated cultivar), 'Longjing43' ('LJ43', green cultivar) and 'Fuding Dabaicha' ('FD', green cultivar), which had been maintained at dimensions of 1.4 m wide, 0.5 m within the row and 1.5 m tall plants from the soil surface, were used in this study. The second developing leaves from tender shoots were collected successively every 2 h during a 48-h-period from the 16th to 18th of September in 2017 at the onset of new bud bursts. For each sampling, the second developing leaf was collected from the tea plants, frozen immediately in liquid nitrogen, and stored at −80 °C for further use.
Environmental parameter measurement
-
Environmental parameters were measured as previously reported to monitor the growth conditions of tea plants during the experimental period (Fig. 1)[21], including the Photosynthetic Activity Radiation (PAR; Light Scout® Quantum Light Meters, Item#3415F, Spectrum Technology® Inc. USA), temperature, humidity and the content of CO2 around tea leaves (TEMP/RH/CO2 hand-held meter, catalog#3440, Spectrum Technology® Inc. USA). PAR increased from 06:00 at sunrise and reached the highest point at around noon (Fig. 1a). Consistently, the temperature increased as PAR increased but decreased during the night (Fig. 1b). In contrast, humidity showed an opposite pattern: as the PAR and temperature rose at sunrise and gradually increased, the humidity dropped but as the temperature dropped at sunset, the humidity escalated (Fig. 1c). CO2 was detected to monitor the respiration of tea plants during the diurnal cycle (Fig. 1d). Data showed the CO2 concentration reached its maximum at about 22:00 when tea plants could not maintain photosynthesis without PAR during the night.
Chlorophyll determination
-
Chlorophyll determination was carried out according to the method previously published[27]. Tea leaves were crushed in liquid nitrogen and chlorophyll extracted overnight in the extraction solvent (5% acetone in 95% ethanol, v/v). After centrifuging at 10,000 g for 5 min, the absorbance of the extract was measured at 663 nm and 645 nm by an Ultrospec 2100 pro spectrophotometer (GE Healthcare Biosciences). Chlorophyll a and b were calculated by the formula: Chl a = 12.7A663 − 2.69A645; Chl b = 22.9A645 − 4.68A663[45]. At least three independent biological replicates were measured for each sample.
Catechin analysis
-
Catechins were extracted and analyzed according to a previous study[21]. Freeze-dried leaf powder (100 mg) in 3 mL of 80% methanol was subjected to ultrasonic sonication for 10 min at 4 °C. After centrifugation (13,000 rpm, 10 min), the extract was collected and the residues were re-extracted twice with 1 mL of methanol. The supernatants were combined and filtered by 0.22 μm organic membrane and collected for HPLC analysis.
For HPLC analysis, the extracts were analyzed using a Waters 2695 HPLC system and a Waters 2489 ultraviolet-visible detector, with a reversed-phase C18 column (Phenomenex 250 mm × 4.6 mm, 5 μm) at a flow rate of 1.0 mL·min−1. The flow rate, detection wavelength, column temperature, and injection volume were set at 1.0 mL min−1, 278 nm, 25 °C, and 10 μL, respectively. The separation used solvent A (0.2% acetic acid in water, v/v) and solvent B (100% methanol) with the following gradient: 0 min, 94% A, 6% B; 4 min, 94% A, 6% B; 16 min, 86% A, 14% B; 22 min, 85% A, 15% B; 32 min, 82% A, 18% B; 37 min, 71% A, 29% B; 45 min, 55% A, 45% B; 50 min, 55% A, 45% B; 51 min, 94% A, 6% B; 60 min, 94% A, 6% B. C, EC, GC, EGC, EGCG, GCG and ECG were used as standards (Sigma Chemical Company, USA).
Statistical analysis
-
Data were obtained from three independent biological replicates. Statistical analysis was conducted using Minitab 17.0 statistical software (Minitab Inc., United Kingdom). One-way analysis of variance (ANOVA) with grouping information and Fisher's least significant difference (LSD) test at the 5% level was used.
- This work was financially supported by the National Natural Science Foundation of China (32072623, 31700611 ) and the Natural Science Foundation of Anhui Province-Outstanding Youth Project (2008085J18). We would like to acknowledge Prof. Tao Xia and Prof. Liping Gao for sharing flavonol standards and LC-MS/MS methods. Thanks to students Dandan Sun and Binbin Cai for sample collections.
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang X, Zhu W, Cheng X, Lu Z, Liu X, et al. 2021. The effects of circadian rhythm on catechin accumulation in tea leaves. Beverage Plant Research 1: 8 doi: 10.48130/BPR-2021-0008

The effects of circadian rhythm on catechin accumulation in tea leaves
- Received: 29 July 2021
- Accepted: 24 August 2021
- Published online: 02 September 2021
Abstract: As the major bitter- and astringent-tasting components in tea leaves, catechins play an essential role in determining flavor characteristics and health benefits of tea products. This study analyzed the circadian rhythm effects on the accumulation pattern of catechins in tender leaves of three tea cultivars, 'Huangkui' (HK), 'Longjing43' (LJ43) and 'Fuding Dabaicha' (FD). There were two major factors involved in the catechin regulation in tea plants: genetic background of the cultivar-specific regulation, and temporal effects of the circadian rhythm. Etiolated leaves of 'HK' contained lower levels of catechins than 'LJ43' and 'FD'. The composition of EGCG in total catechins was significantly higher in 'LJ43' leaves (65%) when compared with that in 'HK' and 'FD' (45%). The accumulation of individual catechins increased during the daytime but decreased overnight, especially over the time period of 22:00−02:00. The correlation analysis between catechins and environmental factors indicated catechin accumulation in tea leaves potentially resulted from a combined regulation of light and temperature. These findings provide new insights into our understanding of circadian rhythm regulation on flavor-important secondary metabolites in tea leaves.
-
Key words:
- Circadian rhythm /
- Camellia sinensis /
- Catechins /
- Light /
- Environmental factors