








-
Biological nitrogen fixation (BNF) contributes to 50% of the total annual usable nitrogen (N) of the biosphere[1−3]. The N molecule itself is highly inert; however, its fixation can generate compounds that contain reactive N (Nr), including NH3, NO, NO2, N2O, HNO3, and other organic N compounds[4]. Though N is abundantly present in the atmosphere, it is the most critical and limiting factor for ecosystem productivity[5,6], which is supported by BNF.
A group of bacteria and archaea named diazotrophs carry out BNF. A high percentage of total BNF is carried out via symbiotic N fixation in which the host plant provides photosynthetically fixed carbon to the symbiotic diazotrophs for their growth, while the host plant obtains the biologically fixed N[7]. Moreover, under specific conditions, non-symbiotic, free-living diazotrophs such as Burkholderia, Azotobacter, Klebsiella, Azospirillum, Bacillus, and Clostridium can also fix substantial amounts of N, which is known as free-living nitrogen fixation (FNF)[8,9]. In some cases, diazotrophs establish weak associations with lichens and cycads, including Ceratozamia Mexicana and some higher plants (e.g., conifers, cereals, and sugarcane), or colonize plant tissues and perform N fixation, which is considered associative N fixation[10,11]. Thus, FNF specifically is a form of BNF in which no obvious structures or symbiosis are established between plants and microbes. So far, there is no exact delineation between these forms of BNF[12]. In light of aforesaid definition, free-living N fixers include soil-dwelling associative bacteria (such as Cyanobacteria, Azotobacter, Klebsiella, and Pseudomonas); heterotrophic bacteria (such as Xanthomonas sp. and Phyllobacterium sp.[13]; anaerobic bacteria that inhabit reduced soil layers[9,14]; endophytic bacteria (such as Herbaspirillum sp., Azoarcus sp., and Gluconacetobacter diazotrophicus) that resides inside plant tissues; lichens (such as Lobaria pulmonaria and L. quercizans[15,16]; and species of archaeal domain (such as Methanosarcina barker and Methanococcus thermolithotrophicus[17]).
Recognizing the fact that free-living N fixers are growing, they require a large amount of carbon (C) and energy compounds to carry out their cellular processes[18,19]. Additionally, in symbiotic N-fixing bacteria, N fixation mainly occurs in non-growing cells, while in free-living N-fixers, fixation only occurs in growing cells. The N fixation rate of free-living fixers is consequently lower than symbiotic fixers that fix N in a static mode, returning the source of C[7]. Moreover, the lifetime of a free-living N fixing diazotroph is much shorter than a nodule that carries out N fixation activity for up to one month[19]. Although FNF is ubiquitous, research on FNF is largely lacking. Compared to symbiotic N fixation, FNF can potentially contribute to much higher levels of N in an ecosystem[12]. For example, studies on tropical forest systems identified that FNF occurring in the rhizosphere, litter, and on the decaying surfaces of logs, leaves, and plants contributed 6 kg N ha−1 yr−1 while 4.5 kg N ha−1 yr−1 via symbiotic N fixation[20].
Our review focuses on FNF, which is a global phenomenon of N addition to ecosystems. However, N inputs by FNF is difficult to estimate due to profound spatial and temporal variability in FNF[7]. We compiled and integrated information related to the biology and diversity of FNF organisms as well as factors that influence FNF. In addition, we discussed: 1) the contribution of FNF to the N budget of different ecosystems; 2) quantification of FNF using different methods; and 3) anthropogenic activities that affect FNF. It is hoped that our discussion deepens understanding of FNF functions across various ecosystems.
-
Around 90 genera of diazotrophs possess the ability to fix atmospheric N[21]. Diazotrophs exhibit the following range of physiologies: they are strictly aerobic (such as Cyanobacteria and Azotobacter); facultatively anaerobic (such as Bacillus, Carnobacterium, Paenibacillus, Pseudomonas, gamma-proteobacteria, and Actinobacteria) or anaerobic (such as Clostridium); heterotrophic (such as Rhodobacter and Anabaena); phototrophic (such as Rhodobacter capsulatus); and chemolithotrophic (such as Leptospirillum ferrooxidans). Cyanobacteria (such as Cyanothece sp. and Synechococcus sp.) is prevalent in terrestrial ecosystems because of its ability to colonize matter in extreme environments like deserts and tundra[10,22]. Cyanobacteria can also grow on tree trunks in temperate or tropical forests as well as on moss carpets where they decompose litter and marine environments (e.g., Crocosphaera watsonii)[23]. Moreover, active N fixation by heterotrophic bacteria, such as Clostridium butyricum, Enterobacter agglomerons, and Klebsiella pneumoniae, has been reported in different habitats that even include Antarctica[24]. Lichens have also been reported to fix N in many ecosystems[25]. Data from Hawaiian forests reveal that the canopy features a higher rate of N fixation over the forest mainly because of lichen abundance in these locations[26,27]. Within the Archaea, N fixation has been found only in the methanogens. For example, Methanococcus thermolithotrophicus, a thermophilic lithotrophic methanogen, can fix N at 60 °C[17]. As free-living N-fixers have diverse natures, habitat, and distributions, considerable spatio-temporal variability exists at both biome and microbiome scales, which in turn results in significant variation in the overall rate of N fixation[27].
-
Two groups of nitrogenase in soil catalyze N fixation[28]. The first group consists of molybdenum (Mo) nitrogenase; vanadium (V) nitrogenase; and an iron (Fe) nitrogenase[29,30], which are oxygen sensitive and differ in kinetic parameters[29]. The second group contains a less-studied nitrogenase of the bacteria, Streptomyces Thermoautotrophicus, which is structurally very different from group 1 nitrogenases[31]. This enzyme is not sensitive to O2 and other known nitrogenase inhibitors[32]. X-ray studies have revealed that nitrogenase of the first group share a similar structure[13,33]. In general, nitrogenase is composed of two major components[34−36]. Component one is termed 'MoFe protein', which possesses unique subunits and cofactors. In MoFe protein, subunits α and β are combined as α–β. Between α–β, a prosthetic group termed the P-cluster is present (8Fe:7S P)[37,38]. The 7Fe: M: 9S: C: homocitrate cofactor (also known as M center) contains the centrally located Fe atoms that constitute the substrate binding site of the nitrogenase enzyme[39,40]. In addition, the small subunit, designated δ, is also present that stabilizes two tetramers[39]. The Fe protein is the second component of nitrogenase, with two subunits that are bridged by a 4Fe:4S cluster[40]. Each subunit of Fe protein has its own ATP binding site that reduces MoFe protein[41].
Mo-dependent nitrogenase is the predominant nitrogenase present in almost all free-living N fixing bacteria[28,42]. The predominate form of nitrogenases in archaea is Mo-nitrogenase, consisting of the same subunits as bacteria, excluding nitrogenase reductase, which in archaea is a homotetramer, unlike the homodimer found in bacteria[17]. Only in A. vinelandii are all three types of nitrogenase of the first group present[43, 5]. However, it has also been demonstrated that Mo-nitrogenase shows high efficiency at high temperatures, while alternative nitrogenases are more efficient at lower temperatures[18,44].
-
The N fixation gene, i.e., nif that encode for the nitrogenase, are an evolutionarily conserved set of genes in N fixing microorganisms. The nif genes have many subunits that encode for proteins, forming the structural subunits of the nitrogenase enzyme, the cofactors, and other regulatory proteins involved in N fixation. The three nif genes, nifH, nifD, and nifK, are highly conserved structural genes, and are grouped together in one operon in almost all N fixing microorganisms, barring some rhizobia species. The gene nifH encrypts for Fe-protein, and the nifD and nifK encode for Fe-Mo proteins (dinitrogenase), respectively[45]. The nifLA operon consists of nifA and nifL genes that regulate the expression of other nif genes. Various factors tightly regulate the transcription of nif genes, including levels of fixed N, carbon, ATP/ADP ratio, and oxygen concentration[45,46]. The nifA gene encodes for a binding protein. The RNA polymerase sigma factor σ5 is activated under N limiting conditions and activates the expression of other nif genes[47]. Moreover, NtrB–NtrC regulatory systems usually regulates nifA transcription[48,45]. nifL gene-encoding for the flavoprotein is activated under N excess and high redox status, inhibiting the expression of nifA and other nif genes. Another locus is called nifx, which is also supposed to work in a similar way to nifL genes in inhibiting nif expression[49]. Regulation of N fixation by nifL–nifA systems varies from organism to organism[45]. In Kallesbela pneumoniae, the N sensor protein glnB regulates NtrB–NtrC systems, which then control the expression of glnK, nifL, and nifA proteins[46,50]. Under N-limiting conditions, glnK is essential to stop nifL from hindering nifA. Conversely, in A. vinelandii, glnK is not required for nifL to inhibit nifA under conditions of N limitations[51−54]. For diazotrophs such as Azospirillum brasilense and Rhodospirillum rubrum, post-translational regulation of nitrogenase has been demonstrated[55−57]. In the presence of external sources of N such as ammonium, a regulatory protein, dinitrogenase reductase glycohydrolase (DraG), is activated, switching off nitrogenase enzymes. In N-limiting conditions, dinitrogenase reductase ADP-ribosyltransferase (DraT) activates nitrogenase activity. However, our knowledge about the detailed regulation mechanism of DraG-Drat remains limited[57].
The structural organization of nif genes also varies extensively within genomes of different N fixing organisms. In K. pneumoniae, 20 nif genes are organized into 9 transcriptional units that cluster together on the chromosome[58]. Similar nif gene organization is found in Azotobacter, Asospirillum, and Clostridium[40]. In cyanobacterium, Anabaena sp, strain PCC 7120, 14 nif genes have been recognized[59]. The nif genes in Anabena sp. are not organized into clusters or groups, but rather have been dispersed due to the insertion of intervening DNA elements[59,60]. The classification of the methanogen nif gene discloses some transformations when compared to bacteria[17]. In methanogens, novel homologues of the bacterial N sensor regulator glnB are located between nifH and nifD genes. Moreover, six nif genes (nifH, nifD, nifK, nifE, nifN, and nifX) and the glnB in a single operon are unique to methanogens[17].
-
Various studies have reported that symbiotic N fixation is a dominant source of N to the ecosystem[21,61] which can supply 150 kg N ha−1 yr−1 to an ecosystem. However, N supply by FNF may be as high as ~76 Tg N yr−1, exceeding the overall supply by lightning (5 Tg N yr−1) and industry-fixed N (120 Tg N yr−1)[12]. It has been shown that FNF contributes to a significant amount of N in ecosystems in the absence of symbionts[22]. For example, biological soil crusts with a variety of microbial communities may fix significant quantities of N in dryland ecosystems with scarce symbiotic N fixation[62−65]. FNF is the major source of N in temperate and boreal forests where abundant cyanobacteria are present in ground moss covering[66]. FNF provides major support to plant production in wetland ecosystems in Brazil where symbiotic N fixation is rare[8,67,68]. Additionally, FNF is the dominant source of N availability in temperate forests, temperate grasslands, and shrub lands, and has also been found to occur in deep soils, canopy soils, and canopy leaves[9,12,69−71]. FNF may be high at sites where N demand is high and vice versa. As FNF occurs under diverse and variable environmental conditions across a variety of substrates, predicting actual rates of FNF can be challenging. Studies attempting to estimate the rates of N fixation by symbiotic and free-living pathways are not from simultaneous assessments and are typically not from the same individual sites[12]. In fact, methodological constraints as well as significant spatial and temporal variations found in the process preclude us from developing conclusive estimates, potentially leading to an underestimation of global rates of FNF.
-
A major factor that limits FNF is the O2 liability of nitrogenase enzymes[29,30,72]. As a mechanism to protect nitrogenase from O2, most obligate and facultative aerobes tend to increase their cellular respiration rate, which tends to lower O2 levels. Further, N-fixers have developed other mechanisms to avoid O2 toxicity by carrying out fixation in separate cellular compartments that spatially separate N fixation from photosynthesis[73,74] or a conformational change in nitrogenase, though this also affects fixation[75]. The O2 liability of nitrogenase confines N fixation to anaerobic or microaerobic environments, with the exception of A. vinelandii, which can fix N2 under entirely aerated conditions[76]. In anaerobic environments, rates of N fixation remain relatively high because there is no need to protect nitrogenase against O2, and the energetic cost of fixation is relatively low. However, the capacity to fix N by anaerobic and microaerophilic bacteria is constrained by the inefficiency of anaerobic metabolism. Indeed, O2 concentrations in environments are constant and thus hard to quantify[46,72,75]. An optimal O2 requirement cannot be defined for a mixed microbial community, where each diazotroph has a different O2 requirement.
Metal ions
-
Molybdenum (Mo) is central for the activity of nitrogenase enzyme; however, in some species it is replaced by a vanadium (V) or iron (Fe)[77−80]. In terrestrial ecosystems, where Mo is often limited in soil, a significant fraction of FNF is carried out by either vanadium nitrogenases or iron nitrogenases[15,81−83]. Bacteria also require other metals like K, Ca, and Mg to fix N. Deficiency of these metals affects the N-fixing efficiency of some FNF bacteria, as these act as cofactors in reactions[84].
Energy requirement
-
FNF require a continuous input of reducing source and ATP supply[85,86]. Generally, 16 molecules of ATP are needed to fix one molecule of N2[87]. The energy requirement of FNF varies with the organism, energy source, and environmental factors. For instance, photosynthetic organisms can attain the required energy by the natural process of photosynthesis. For heterotrophic microorganisms, required energy is solely obtained from the catabolic activities of the organic substrate. A lower C/N ratio in the substrate and presence of O2 is associated with decreasing capacity of FNF[87].
Nutrient availability
-
Nutrient limitations are related to enzymatic requirements and are therefore considered an overriding control over N fixation[88]. Low phosphorous (P) availability constrains the rate of FNF by affecting ATP and regulating nitrogenase enzyme[89−91]. One previous study suggested that FNF is ~27% higher in soils receiving P inputs over an unamended control[92]. Similar positive associations between P inputs and FNF have also been reported in tropical forest systems[12]. Wurzburger et al.[93] found that P availability can influence soil Mo availability and thus limit FNF in P-depleted soils. Carbon (C) are also known to limit N fixation[90, 94−96,97]. In Clostridium pasteurianum, FNF is inhibited by deficiency of C. Moreover, FNF is also affected by the C quality of the substrate. For example, acetate appears to impede nitrogenase activity of Azotobacter paspali[94−96,97].
The availability of inorganic N decreases the rate of FNF. Moreover, lower N/P ratio is associated with higher rates of FNF by soil-dwelling bacteria[98−101]. In contrast to the stimulatory effect observed with P fertilization, N fertilization has an inhibitory effect on N fixation[102]. The rate of FNF is higher under N-limiting conditions[72]. Under severe N-limiting conditions, microorganisms meet their N demands through fixing atmospheric N via obtaining exogenous N or by the re-allocation of endogenous N[103]. Exogeneous sources of N in the form ammonium, glutamine, and histidine are known to decrease the FNF rate in soils[104]. Research has shown that ammonium and glutamine act as inhibitors of nitrogenase enzyme synthesis through regulation of nifA gene expression. Nitrate can also decrease FNF in A. brasilense, while in C. watsonii no inhibition was observed[105].
Edaphic factors
-
Temperature has been shown to inhibit nitrogenase activity and thus restrain FNF, with maximum rates occurring at ~26 °C, depending on species and ecosystem[106]. Insufficient soil moisture might limit the capacity of diazotrophs for N fixation due to altering O2 availability. Soil texture can affect O2 concentrations and the rate of substrate diffusion in soils[107]. Soils with high clay content are estimated to have high N fixation rates due to low O2 pressure on the diazotrophs[107]. Moreover, soil pH has an influence on the abundance and diversity of diazotrophs[108,109]. Although soils contaminated with heavy metals have depressive effects on rhizosphere microbial communities[110], their effects on free-living N fixers and the rate of FNF are poorly understood. Land use adoptions also alter the diversity of free-living bacteria[111,112]. The abundance and diversity of free-living N fixers have drastically changed alongside the deforestation of the Amazon rainforest[113,114]. Additionally, plants can alter FNF rates via various indirect mechanisms[115−117]. For example, Cotta et al.[118] mentioned that planting genetically modified maize significantly changes the abundance of N-fixing bacterial and archaeal communities.
-
FNF is a ubiquitous phenomenon of terrestrial ecosystems occurring on plant surfaces, litter, decaying logs, and in soils[12]. Cyanobacteria are the dominant N-fixers in terrestrial ecosystem because of their ability to colonize extreme environments ranging from deserts to tundra[23]. The N inputs by FNF to the total N budget of an ecosystem can vary due to ecosystem and community-level patterns of bacteria. According to Reed et al.[12], FNF contributes to 1−20 kg N ha−1 yr−1 at the biome scale, and symbiotic N fixation reaches as high 150 kg N ha−1 yr−1. The rate of N fixation by heterotrophic bacteria alone ranges from 1 to 5 kg ha−1 yr−1 for decomposing litter in different ecosystems, while the rate of Cyanobacteria ranges between 1 and 10 kg ha−1 yr−1[94,119]. The estimated range of N fixed by FNF is > 10 kg N ha−1 yr−1 for tropical rain forests[120,121], 4 kg N ha−1 yr−1 for desert ecosystems, and > 2 kg N ha−1 yr−1 for tundra and boreal forests[122, 66]. Research has revealed that FNF bacteria add 2 kg N ha−1 yr−1 in the soils of semiarid grasslands in California. Although these rates of FNF are far below symbiotic N fixation, FNF could nonetheless supply a meaningful amount of N to the total N budget of a particular ecosystem in the long term.
FNF in agroecosystems
-
Contributions to total N needed in agricultural systems might not be primarily fulfilled by FNF. However, it can fulfill a substantial amount of the total N requirement of crops[62,123]. Studies have shown that N fixed by free-living Cyanobacteria in rice paddies is ~12−33 kg N ha−1 in a cropping season[124], ~25−150 kg N ha−1 yr−1 in sugarcane[8,68], 20 kg ha−1 yr−1 in wheat, < 10 kg N ha−1 yr−1 in extensive tropical savannas used for grazing[119,125], and < 5 kg N ha−1 yr−1 in croplands other than leguminous crops[126]. The agronomic effects of FNF on crops have provided useful model-systems for rhizosphere studies, plant-microbe interactions, and environmentally sound agricultural cultivation. Sustainable agriculture requires smarter utilization of FNF, which can offer a sound means of reducing external N input; therefore, future research into agroecological sustainability should focus on FNF.
FNF in aquatic ecosystems
-
N fixation in aquatic ecosystems also cannot be overlooked[127]. Microorganisms with the capability to fix N have been detected across various aquatic environments[100,128,129]. In these environments, cyanobacteria are the dominant diazotrophs[130]. Previous studies reported N fixation and diazotrophs in lakes, but these studies neglected to highlight the contribution of FNF. For example, MacGregor et al.[131] detected N fixation below the photic zone in oligotrophic Lake Michigan, but actual rates remain unknown. A study conducted in an oligotrophic, hypersaline, meromictic lake in California showed that anaerobic bacteria possessing significant potential for N fixation were present across all depths[132]. In the deep water of Lake Michigan, N fixation was also found to be an important source of N[131]. A study conducted in Lake George in New York State disclosed that a diverse group of diazotrophs, including unicellular and filamentous cyanobateria, alpha- and gamma-proteobacteria, and an undefined group of bacteria were found in the lake. Again, the actual rate of N fixation was not measured[133].
On the other hand, in oceanic areas, a significant portion of N fixation is interceded by cyanobacteria and anaerobic bacteria[134,135]. Montoya et al.[136] showed that in the tropical North, N fixation supplies a substantial amount of N. Similar phenomena of N fixation exist in the Mediterranean Sea[137]; however, no past studies targeted FNF contributions in oceanic environment.
-
Acetylene reduction has been used to quantify FNF, which follows the principle of no specificity of nitrogenase. The reduction of acetylene is carried out by nitrogenase, which produces ethylene. Then, ethylene production is quantified by gas chromatography coupled with a flame ionization detector. Finally, the acetylene reduction rate is converted into a rate of reduction of N2. About one mole of N2 is reduced by three moles of acetylene. During N fixation, hydrogen evolution, which involves electron transfer, occurs. The general conversion factor is 4:1[3,138] and can vary across natural environments[139,140]. Acetylene reduction assay has several limitations: 1) during acetylene reduction assay, microorganisms may become N-deprived and can perhaps reduce more acetylene[138]; 2) acetylene is not suitable for some microbes like methanotrophic bacteria because they cannot tolerate acetylene due to interference with methanogenesis that can cause starvation or death of the microbe[141]; and 3) three different types of nitrogenases vary in their affinity for the substrate acetylene, consequently producing different acetylene reduction rates that could lead to the misinterpretation of FNF rates in a natural system.
15N-tracer method
-
The 15N-tracer method has become a widely adopted method in modern research[6]. This method utilizes the 15N stable-isotope that occurs naturally in small amounts. The FNF organism is given 15N2, and the incorporated isotope in the particulate organic N is quantified via stable-isotope mass spectrometer[142]. This method is more reliable relative to the acetylene reduction method, as this directly measures N fixation[142]. However, the 15N-tracer method purely quantifies net N fixation. Thus, biased results may occur where N fixation is vigorous, and there is only partial release of the fixed N by microorganisms[143].
Stable isotope abundance method
-
The natural abundance of stable isotopes (14N and 15N) is another method that can be used to quantify the FNF of N. 14N is a lighter isotope that is preferred over 15N in chemical reactions, as the bonds of the 14N are easily broken, leading to substrates featuring greater enrichment of 15N[144,145]. For N2, the δ15N value is normally 0%. The δ15N value of an organism approaching 0% indicates a fixation of N. Many aquatic environments like lakes and oceans facilitate this phenomenon[146,147]. The stable isotope abundance method is not usually suggested because it misses volumetric N fixation rate, and in many cases, the N sources may not be precisely known.
Gene markers
-
The nif genes have been widely used as a marker for studying the identification, quantification, diversity, and phylogenies of bacteria in various environments[148−152]. PCR-based amplification and sequencing of nifH DNA have been used for detecting non-cultural, soil-borne microorganisms[153]. Many correlation studies revealed the effects of environmental factors, plant genotypes, and nutritional availability on the diversity and abundance of the nifH gene[148,154]. Although the nifH gene is highly conserved, it is not necessarily a true indicator of actual N fixation because these genes are prone to mutate and may ultimately be excluded from the genome[155,156]. Finally, three types of nitrogenases are produced by separate genes, i.e., nifH encodes for Mo-nitrogenase, anfH for Fe-nitrogenase and vnfH for V-nitrogenase; therefore, measurements based solely on nifH abundance may lead to biased interpretations.
-
The productivity and dynamics of any ecosystem are limited by the supply of biologically available N[157,158]. It is well known that anthropogenic activities are constantly altering the global N cycle[159]. Over-use of fertilizer in agriculture is a factor that has approximately doubled the rate of N input into terrestrial ecosystems[160]. This has resulted in the acidification of soils as well as the unbalancing and loss of soil nutrients that are essential for the maintenance of soil fertility[161]. The estimated total production of N from synthetic fertilizer is 120 Tg N yr−1, which represents the largest contribution of Nr to the atmosphere. Furthermore, the wide cultivation of leguminous crops contributes an additional 60 Tg N yr−1. N in oxidized forms (NO and NO2) as a byproduct from automobile combustion and industrial emissions adds an additional 40 Tg N yr−1 to the atmosphere. The estimated anthropogenic production of Nr is therefore 180 (± 20) Tg N yr−1[4,160,162]. The emission of Nr by natural as well as anthropogenic activities is expected to increase in the future (Fig. 1). It is also reasonable to expect that with global population growth the demand for food and feed will increase the production of reduced N in the form of N fertilizers that would require greater fossil fuels combustion. The future emission of NOx is highly uncertain, and it is expected to increase alongside industrial developments until abated by pollution controls. According to Fowler et al.[160] the N input to ecosystems via natural and anthropogenic fixation will increase in the future (Fig. 2). Increasing N availability generally reduces the biological diversity of ecosystems[163], which in turn affects the rates of various biological processes. The increasing level of N in soils is known to inhibit the growth of symbiotic as well as free-living organisms[164]. Nitrogen addition has obvious effects on the C pool of the ecosystem[158]. Excessive nitrogen deposition favors the uptake and storage of carbon. This high C sequestration will demand high N input, which will likely be insufficient in the future if N addition as N2 fixation is controlled by resources that do not contain N.[22,165]. Therefore, there is a need to increase BNF to meet the N supply required to balance the increased C. The expanded utilization of BNF will reduce the dependence on synthetic N, which in turn will reduce the negative effects on biogeochemical cycles. FNF may compensate N losses through denitrification, increasing soil phosphate levels by altering phosphatase activity, and increasing the net primary productivity of terrestrial ecosystems[12,91].

Figure 1. Anthropogenic contribution of Nr to the atmosphere[160].
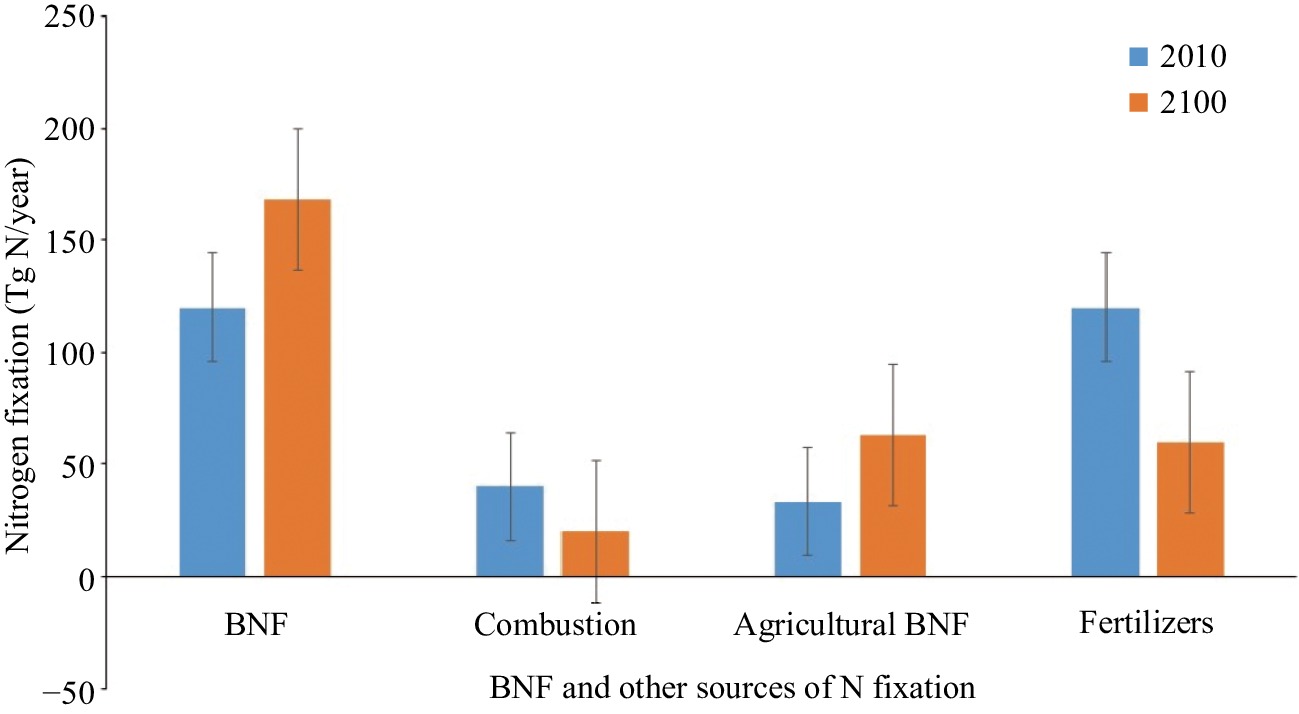
Figure 2. Changes in N fixation in the coming future[160].
-
It is now established that anthropogenic activities have increased the total global N deposition. This has destabilized biogeochemical cycles, including the N cycle. However, to accurately address the effects of anthropogenic activities on the N cycle, we need a detailed understanding of the FNF process. Overall, available research has concluded that FNF is a ubiquitous phenomenon across ecosystems that contributes significantly to the N inputs of a range of ecosystems. Based on knowledge gaps, we suggest that future research in the following mentioned areas will increase our understanding of FNF:
The crucial need is to resolve uncertainties in observing N2 fixation rate per unit area along with spatial distribution of the N fixing bacterial species. Previously, authors have reported global N fixation across various ecosystems; however, most of these studies were unable to investigate several ecosystems[12,119,162]. Recently, research has produced more accurate findings regarding cultivated and uncultivated lands, using data from different sources[126,162]. Additionally, none of the studies take into account the constraints that FNF bacteria encounter in their life cycle. In fact, methodological constraints, spatiotemporal variation of the process, and complex interactions between plants, microbes, and environments do not allow us to develop conclusive estimates for overall fixation rates in nature and/or agroecosystems. We suggest that site- and situation-specific research is necessary to obtain real estimates on a granular scale. This can be achieved by wisely designed models that consider the dynamics, heterogeneity, influences, and existing constrains to N fixation by free-living N-fixing bacteria. Thus, we propose a framework for gaining a better understanding of FNF (Fig. 3). Another essential issue in need of clarification is the ecology of FNF bacteria. We urgently need to deepen our understanding of the numerous factors, including biotic and abiotic, that affect the diversity and abundance of FNF bacteria in the rhizosphere, which in turn influence the rate of N fixation.

Figure 3. Framework for expanded utilization of FNF.
- Our work was funded by the Key Project from the Ministry of Sciences and Technology of China (No. 2017YFC0505100). We are grateful to the Chinese Academy of Sciences for CAS-President's International Fellowship Initiative (CAS-PIFI), Fellowship (Grant No. 2019PC0011 and 2021PB00094).
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Khan S, Nadir S, Iqbal, Xu J, Gui H, et al. 2021. Towards a comprehensive understanding of free-living nitrogen fixation. Circular Agricultural Systems 1: 13 doi: 10.48130/CAS-2021-0013

Towards a comprehensive understanding of free-living nitrogen fixation
- Received: 08 August 2021
- Accepted: 27 October 2021
- Published online: 30 November 2021
Abstract: Free-living nitrogen fixation (FNF) is a ubiquitous phenomenon that plays a modest role in the (N) economy of an ecosystem. However, sampling difficulties, methodological constraints and environmental controls have presented challenges for predicting the actual rate of FNF. Therefore, a deeper understanding of the accuracy to design models that consider dynamics, heterogeneity, influences, and other limitations is needed. This review presents an overview of the biology and diversity of microorganisms related to FNF as well as various ecological controls that influence these microorganisms. We also discussed contributions of FNF to the N input of various ecosystems. Overall, previous research has shown that considerable spatiotemporal variability exists in microbial types at both biome and microbiome scales, resulting in significant variation in FNF. Beyond this, rate of FNF is controlled by certain factors, such oxygen and metal ion availability, source of energy and soil nutrients, temperature, and pH. Empirical evidence increasingly indicates a significant contribution of FNF to N inputs in natural, agricultural, and aquatic ecosystems. It is inferred from this review that for the expanded exploitation of biological nitrogen fixation (BNF), we must pay additional attention to FNF because it occupies a central role within the process. Finally, we propose a framework for the quantification of FNF alongside a suite of recommendations that would deepen our understanding of FNF.