








-
Plant nutrient metabolism is generally achieved through the source-sink relationship, in which some organs function as source tissues and others function as sink tissues. Leaves act as source tissues and generate assimilation products through photosynthesis for later transport to sink tissues, such as tubers, fruits, and seeds[1]. As the main form of carbon storage, sugar functions as both an energy source and structural component in plants, which also affects gene expression, participates in various processes of plant growth, development and metabolism, and affects the response of plants to biological and abiotic stresses[2−4]. Carbohydrates produced by photosynthesis are transported to the cytoplasm in the form of triphosphate to be converted into hexose phosphate or sucrose, which are stored in vacuoles for later use[5]. Sucrose can be transported to other tissues to be converted into hexose or starch under the action of sucrose synthase and invertase[6], while excess photosynthates, which accumulate in chloroplasts during the day, are temporarily stored in the form of starch and converted to maltose and glucose at night[7]. Therefore, a strict sugar sensing and signaling system is essential for coordinating photosynthesis and carbon metabolism to maintain normal plant growth and development[8−10].
Among those sugars, glucose which is the most well-studied sugar molecule acting as a form of carbon metabolism and signaling plays a role in specific regulatory pathways. Glucose metabolism is one of the most fundamental processes in living organisms and exists throughout all kingdoms of life. Glucose is metabolized through glycolysis, gluconeogenesis, and pentose phosphate pathways. The glucose in Rosaceae plants is mainly converted from sucrose, with secondary contribution from sorbitol and can be used as a respiratory substrate to participate in glycolysis. Hexokinase (HXK), which can catalyze the metabolism of fructose and glucose, carries out one of the rate-limiting steps in glycolysis[11]. HXK can be divided into glucokinase (GLK) and fructokinase (FRK). The HXKs that prioritize glucose metabolism are known as GLKs, and have been found in germinating corn shield leaves[12], pea seeds (ATP: D-glucose 6-phosphotransferase, EC 2.7.1.2)[13,14], rice embryos[15], tomato fruits, and soybean nodules (EC. 2.7.1.1)[16,17]. Glucose participates in many complex metabolic processes that have been well-described over the past few decades. The effects of glucose signaling on fruit quality and fruit ripening, especially in Rosaceae plants, are discussed in this review.
-
Glucose not only plays a role in metabolic pathways, but also regulates plant growth through signal transduction. There are three main glucose signal transduction pathways in plants: HXK1-dependent glucose signaling; G-protein-coupled signaling associated with the AtRGS1 transduction pathway; and glycolysis dependent on SnRK1/TOR signal transduction pathways.
Sugar signaling is facilitated by HXKs, which play an important role in signal transduction[18]. Studies on different sugars, sugar analogues, and metabolic intermediates in mesophyll protoplast transient expression systems and phenotypic analysis of transgenic Arabidopsis have confirmed that HXK1 is a plant glucose sensor[19−21]. In previous studies, the decoupling of glucose metabolism and signal transduction was achieved by the construction and analysis of two inactive AtHXK1 alleles. A mutant of AtHXK1, S177A, has been shown to lack catalytic activity but retain glucose receptor activity, suggesting that the catalytic function of HXK1 is not coupled with signaling perception and transduction[22]. Glucose binds to AtHXK1, and the binding activity of AtHXK1 protein changes to initiate glucose signal transduction[23]. The localization of HXK protein may also play an important role in its function, since HXK1 is mainly located in the mitochondrial membrane, cytoplasm, and nucleus[24−27]. HXK1 can interact with vacuolar H+-ATPase B1 (VHA-B1) and the 19S regulatory particle of the proteasome subunit (RPT5B) in the nucleus to recruit transcription factors that bind directly to the promoter regions of the light-harvesting chlorophyll a/b-binding proteins CAB2 and CAB3 to regulate gene expression[24]. A cascade of glucose signaling through hormones has also been established by using sugar-insensitive mutants. HXK functions upstream of glucose insensitive 1 (GIN1) and abscisic acid (synthesis) 2 (ABA2) in the glucose signaling pathway[28]. Glucose insensitivity mutants of ABA3 (gin5 and gin6) inhibit the expression of AP2 transcription factor ABI4 in glucose signal transduction[29]. The glucose-insensitive sis4/gin1 mutant is an ABA2 allele that regulates the short chain dehydrogenase/reductase (SDR1) required for ABA synthesis[30]. Therefore, HXK1-dependent glucose signals regulate ABA synthesis by regulating ABA synthesis genes such as ABA1, ABA2, and ABA3, which regulate ABA signaling genes such as ABI8 and ABI5 to regulate plant growth and development[31]. Additionally, etr1-1 and ethylene insensitive mutant ein2 exhibit glucose hypersensitivity[28]. Glucose and ethylene antagonistically regulate protein stability of EIN3 through the Skp1-Cullin-F-box protein (SCF) ubiquitination pathway[32].
Typical G protein signaling transduction pathways include heterotrimer G protein GPCRs, RGS proteins, downstream effector proteins, and other regulatory proteins. However, thus far no GPCRs have been found in plants. The major components of RGS1-dependent G protein signaling transduction pathways in Arabidopsis are Gα (AtGPA1), Gβ (AtAGB1), Gγ (AtAGG1), seven-times transmembrane protein (AtRGS1), and with no lysine (K) kinase (AtWNKs)[33]. In the absence of extracellular glucose stimulation, the Gα subunit and RGS1-Gβγ are in equilibrium, but when glucose stimulation occurs, more Gβγ dimers recruit AtWNK8 to phosphorylate AtRGS1. AtRGS1 then leaves the cell membrane through endocytosis by phosphorylation and releases Gα (AtGPA1) in a free state, leading to initiation of the G protein signaling pathway[34]. It has also been shown that AtWNK8 and AtWNK10 are active under high glucose concentrations, while AtWNK1 phosphorylated AtRGS1 at low concentrations[35].
Sugar signals can be translated by protein kinases, among which the target of rapamycin (TOR) is an important energy metabolism sensor that acts in conjunction with sucrose non-ferment related kinase 1 (SnRK1) to regulate cell homeostasis[36,37]. The glycolysis-dependent SnRK1/TOR signal transduction pathway is the only pathway that is dependent on glucose metabolism. Glucose can activate signaling by TOR, which is interconnected with regulatory-associated protein of TOR (RAPTOR) and lethal protein SEC13 protein 8 (LST8) to comprise the TOR complex 1 (TORC1). RAPTOR interacts with the N-terminus of TOR to promote recruitment of kinase substrates[38], while LST8 binds to TOR's C-terminal kinase domain to regulate substrate selectivity and adjust the activity of TORC[39]. In Arabidopsis, SnRK1 is encoded by three catalytic subunits: SnRK1.1, SnRK1.2, and SnRK1.3[40]. SnRK1s have been shown to be inactivated by sugar and play a central role in energy signaling in a similar manner to the homologous yeast Snf1 and mammalian AMPK proteins, but it is not associated with HXK1-dependent glucose signaling[40]. Sucrose/glucose signals affect the level of trehalose 6-phosphate (Tre6P), which directly binds to SnRK1.1 and reduces the phosphorylation of geminivirus Rep interacting kinase 1 (GRIK1)-SnRK1.1, thereby inhibiting SnRK1.1[41]. SnRK1.1 interacts with RAPTOR1B and phosphorylates it to inhibit TOR's activity[42]. The SnRK1/TOR signaling transduction pathway indirectly responds to glucose signaling by sensing energy levels in plants, of which glucose is a critical indicator[43]. SnRK1 responds to low energy states, while TOR positively regulates key biosynthesis processes and responds to high energy states[40,43,44].
-
Sugar signals, especially glucose, not only play important roles in the normal function of cells but also affect the quality of ornamental crops and fleshy fruits. Fruit color has an impact on the perception of fruit quality, and mostly depends on anthocyanin content. Studies have shown that sugar can induce the synthesis of anthocyanin in plants[45−48]. Anthocyanin is produced in the cytoplasm by the flavonoid pathway and then transported into vacuoles for storage. Phenylalanine can be catalyzed to form anthocyanin by phenylalanine ammonialyase (PAL), chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), dihydroflavonol reductase (DFR), anthocyanin synthase (ANS), and flavonoid 3-glucosyltransferase (UFGT)[49]. The glucose signal transduction pathway is dependent on HXK1 and can increase the accumulation of anthocyanin in grapes by inducing the expression of regulatory and structural genes, including VvUFGT and VvF3H[50,51]. Anthocyanin biosynthesis is also synergistically regulated by the MYB-bHLH-WD40 (MBW) complex[52,53]. MYB can be divided into R1/2-MYB, R2R3-MYB, and R3-MYB, among which R2R3-MYB is the largest MYB family in plants, which regulates anthocyanin synthesis and interacts with bHLH[54,55]. bHLH is the second largest transcription factor family in plants and is involved in plant growth and development processes, including hormone signaling and anthocyanin accumulation[54,56]. WD40 protein is a family of proteins containing 4−10 random WD repeat domains, and the core region is composed of 40 amino acid disabilities. Under the induction of glucose signaling, MdHXK1 stabilizes the members of MBW complex MdbHLH3 protein through phosphorylation, and MdbHLH3 could bind to G-box cis-acting element of MdMYB1 promoter to enhance the transcriptional activation of MdMYB1 and promote the accumulation of anthocyanin[26,57]. Furthermore, MdMYB1 binds to the promoter of GSTs to regulate anthocyanin accumulation, and GST can be induced by glucose signaling[58,59].
-
The content of soluble sugar and organic acid has a strong effect on the quality of fleshy fruit[60,61]. Approximately 85% of the organic acids in apples is malate[62,63], which is a key intermediate in the tricarboxylic acid (TCA) cycle that enters mitochondria as a substrate for respiration[64−66]. Malate also regulates the pH of vacuoles to alter fruit acidity[67,68]. On the vacuole membrane, two different proton pumps, H+-ATPase (V-ATPase) and H+-pyrophosphatase (V-PPase), drive vacuole acidification by passing protons through the vacuole membrane and into the vacuole. In addition, a large number of secondary transporters and channels on the vacuole membrane are responsible for transporting malate from the cytoplasm to the vacuole[69−72]. The MBW complex is also involved in the determination of acidity and vacuolar pH[73]. Glucose signaling contributes to the stabilization of MdbHLH3 protein and transcriptional activation of MdMYB1[26,57]. MdMYB1 directly binds to the promoter of malate transporter V-ATPase subunit genes (MdVHA-B1, MdVHA-B2), a malate transporter (MdtDT) and a V-PPase gene (MdVHP1) to regulate fruit acidity[67]. Meanwhile, the interaction between MdbHLH3 and MdMYB1 could enhance the transcriptional activation of MdVHA-B1 and MdVHA-B2 by MdMYB1, which affects the expression of acid transporters Ma1 and MdtDT to regulate fruit acidity. In citrus, Noemi also encodes a bHLH transcription factor that is a major determinant of fruit acidity[74]. Many enzymes involved in malate synthesis and metabolism have also been identified, such as cytosolic NAD-dependent malate dehydrogenase (cyMDH) and phosphoenolpyruvate carboxylase (PEPC), which are key enzymes involved in malate synthesis, while cytosolic NADP-dependent malic enzyme (cyME) is an important enzyme in malate degradation[75−81]. During the early stages of fruit development, MdbHLH3 specifically regulates malate accumulation by directly binding the E-box cis-acting element of the MdcyMDH promoter independent of MYB transcription factors, which promotes the translocation of sugar signaling from source to sink[82].
-
Ethylene plays an important role in fruit ripening and senescence[83,84]. The accumulation of bright pigments in fruit is a result of the ethylene biosynthesis process, and many different components, such as sugar, are involved in the ripening process through interaction with ethylene biosynthesis in fleshy fruits[85−87]. The activated form of methionine, adenosine methionine (SAM), is converted by ACC synthase (ACS) to 1-aminocyclopropane-1-carboxylic acid (ACC), which is then oxidized by ACC oxidase (ACO) to form ethylene[88]. E3 ubiquitin ligase regulates ethylene levels through ubiquitination of multiple synthesis and degradation components[89,90]. The glucose-responsive E3 ligase MdPUB29 regulates fruit traits through ubiquitination of downstream genes[91]. In the presence of glucose signaling, the ubiquitination of MdbHLH3 by MdPUB29 is inhibited, which increases its stability. MdbHLH3 then binds to the promoters of ethylene synthesis related genes (MdACO1, MdACS1, and MdACS5A) to increase their transcription rate and promote fruit ripening[92]. MdbHLH3 also regulates ethylene synthesis by activating the ethylene precursor MdDEP1[93]. Interestingly, MdbHLH3 impacts anthocyanin accumulation, malate content, and ripening in fruits, which are all related to glucose signaling. This suggests that MdbHLH3 plays an important role in the regulation of sugar signaling during the growth and development of apple fruits.
-
Glucose signaling can be transmitted through HXK, G-protein-coupled signaling associated with the AtRGS1 transduction pathway, and glycolysis-dependent SnRK/TOR pathways. Glucose participates in the regulation of fruit quality through signal transduction and metabolism under low glucose condition. Under high glucose conditions (e.g. 6% glucose), glucose is primarily used as a signaling molecule to initiate biological processes[26]. In this review, we summarized a regulatory network in which glucose signaling regulates fruit quality (Fig. 1). We found that the regulation of fruit quality and ripening by glucose signaling is mostly dependent on the HXK1 pathway and the glucose metabolism pathway, with several other pathways directly or indirectly related to HXK1. Therefore, whether the regulation of glucose signaling on fruit quality and ripening depends on G protein signaling pathway remains to be explored. MdPUB29 responds to glucose signals and is independent of HXK1, while MdbHLH3 is stabilized in HXK1-dependent glucose signaling pathways. In the glucose signal transduction process involving MdPUB29, MdbHLH3 is ubiquitinated and degraded, which indicates that the glucose signal transduction by MdPUB29 is distinct. MdPUB29 can trigger reactive oxidant species (ROS) responses and ROS can activate TOR[94,95]. Therefore, MdPUB29 might respond to glucose signals through the TOR/SnRK1 pathway. On the other hand, recent studies have shown that MdHXK1 can regulate the disease resistance of apples and MdPUB29 also regulates the disease resistance of plants[96]. Therefore, glucose signaling is involved in the regulation of plant disease resistance. Whether glucose signaling could coordinate and regulate the synthesis of salicylic acid through MdHXK1 and MdPUB29, thus affecting the disease resistance of plants, remains to be studied.
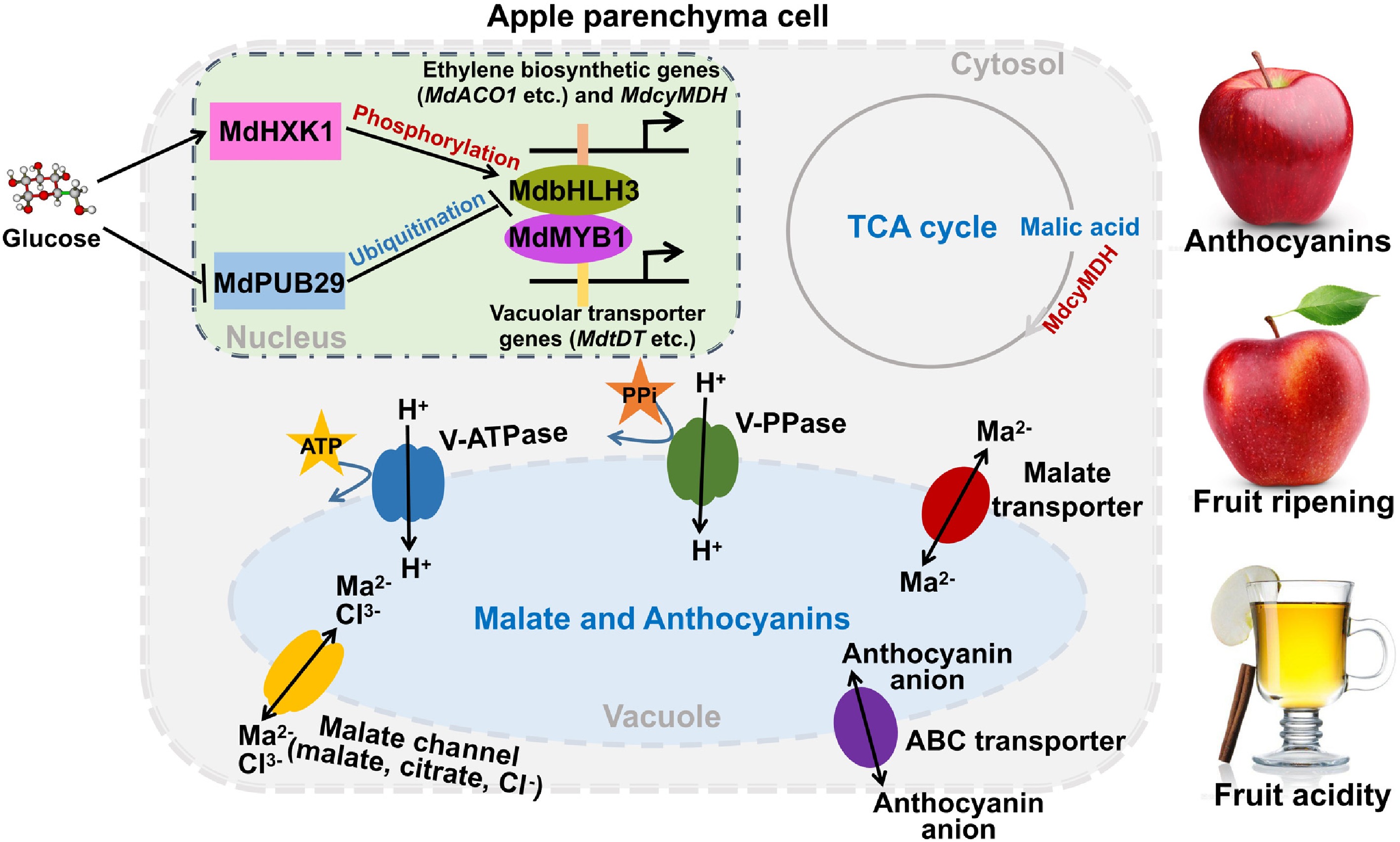
Figure 1. Glucose signaling stabilizes MdbHLH3 by promoting the phosphorylation of MdbHLH3 by MdHXK1 and inhibiting MdPUB29 ubiquitination of MdbHLH3. This process can enhance the transcriptional activation of MdbHLH3 on ethylene synthesis-related genes, such as MdACO1, to regulate fruit ripening and enhance the transcriptional activation of MdbHLH3 on MdcyMDH to regulate fruit acidity through the TCA cycle. Additionally, the interaction between MdbHLH3 and MdMYB1 could enhance the transcriptional activation of MdVHA-B1 and MdVHA-B2 by MdMYB1, which affects the expression of acid transporters Ma1 and MdtDT to regulate fruit acidity. MdMYB1 can also affect fruit color through the transcriptional regulation of genes related to anthocyanin synthesis.
EIN3 is expressed in fruit and can respond to sugar signals. Glucose inhibits EIN3 expression and thus inhibits ethylene response genes that in turn inhibit ethylene signal transduction[97], slowing the release of ethylene and the ripening of fruit. Therefore, glucose can inhibit the ubiquitination of MdbHLH3 by inhibiting the activity of MdPUB29, which induces the expression of ethylene synthesis related genes, promoting ethylene release and fruit ripening. On the other hand, glucose signaling can inhibit the expression of EIN3 to reduce the expression of ethylene responsive genes to inhibit fruit ripening. These two pathways work together to maintain the stability of ethylene in the later stages of fruit development, highlighting the critical role that glucose signaling plays in ethylene synthesis and transduction. This complex interaction may be the reason why ethylene release does not significantly change from the late stage of fruit development to ripening.
MdbHLH3 impacts fruit color, acidity, and ripening pathways and is regulated by glucose signaling, suggesting that it plays a pivotal role in glucose signaling transmission and responses. A total of 13 single nucleotide polymorphisms (SNPs) related to fruit taste are located in the MdbHLH3 gene in the apple GDDH13 V1.0 genome database (www.rosaceae.org)[98], indicating that MdbHLH3 may be responsible for much of the different tastes present among apple varieties. Markers near MdbHLH3 could therefore potentially be utilized for breeding different fruit qualities.
The taste of fruit depends on the ratio of sugar to acid, and glucose signaling affects fruit acidity. Ma1 and tDT are acid transporters found on the vacuolar membrane that play a significant role in the regulation of fruit acidity. Thus far, the impact of glucose signaling on Ma1 has not been reported, although it may play a yet undiscovered role. More research is needed to better understand the impact of glucose signaling on Ma1, as well as many other genes, in order to better understand its role in the regulation of fruit acidity.
- The authors would like to thank TopEdit (www.topeditsci.com) for linguistic assistance during the preparation of this manuscript. This project was supported by grants from the National Key Research and Development Program of China (2018YFD1000200); the National Natural Science Foundation of China (32122080, 31972375, 31902049); and Shandong Province (ZR2020YQ25).
- The authors declare that they have no conflict of interest.
- Copyright: © 2022 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang C, Zhao Y, Sun C, Hu D. 2022. Deciphering the impact of glucose signaling on fruit quality. Fruit Research 2: 3 doi: 10.48130/FruRes-2022-0003

Deciphering the impact of glucose signaling on fruit quality
- Received: 05 November 2021
- Accepted: 27 January 2022
- Published online: 22 February 2022
Abstract: Glucose is a preferred source of carbon and energy for plants. In addition to metabolic functions, glucose is a well-known signaling molecule that regulates plant growth and development through multiple pathways. In this review, the mechanisms by which glucose signaling regulates the accumulation of sugars and organic acids, as well as the ripening of fleshy fruit, are examined. An analysis of these complex molecular networks demonstrates the impact of glucose signal perception on fruit quality.
-
Key words:
- Glucose /
- Sugars /
- Organic acids /
- Ripening /
- Fruit quality