









-
Plants are often subjected to biotic stresses, such as pathogenic bacterial infections, during their growth and development, seriously affecting yield and quality. Chrysanthemum morifolium (chrysanthemum) is one of the top ten traditional flowers in China. However, many diseases affect chrysanthemum development and growth, including black spot disease caused by Alternaria alternata. Black spot disease severely affects chrysanthemum plants or cuttings and may cause chlorosis and necrosis of leaves. Alternaria fungi can produce pectinase and cellulase enzymes that degrade plant tissues, such as plant cell walls, thereby aiding the invasion and establishment of other disease-causing pathogenic fungi[1,2]. Currently, chemical control is the primary control method for biotic stresses; however, it can harm plants, the environment, and humans. Therefore, using molecular biology techniques to develop new disease-resistant varieties is one of the most effective ways to control diseases and insect pests.
Plants have evolved a complex immune system to defend themselves against pathogenic infections through long-term development. When a pathogen invades a plant, it triggers a series of immune responses, and plants recognize pathogen-related molecular patterns through pattern recognition receptors on the cell surfaces (pathogen-related molecular pattern-triggered immunity [PTI])[3,4]. PTI is the first line of defense induced in response to pathogen invasion. However, pathogenic bacteria evolve and secrete a series of toxic effectors to disrupt the plant's PTI defense response mechanism or use the effector to mimic substances produced by plants to facilitate pathogen infection and reproduction in plants to suppress PTI immunity[5]. Simultaneously, plants will configure various types of immune receptor proteins containing nucleotide binding domains and leucine repeat rich domains based on the PTI immunity to monitor and inhibit the activity of infected pathogenic factors, an immunity type called effector-triggered immune response (ETI)[6]. PTI and ETI are primary immune defense mechanisms against plant pathogenic infections.
Reactive oxygen species (ROS) are a class of oxygen-containing molecules with more active chemical properties than oxygen. Various regions produce and remove ROS in plants, and each region has an antioxidant system that regulates the balance between local oxidation and reduction[7]. In contrast, ROS can act as a signaling molecule to induce an immune response to pathogens. Furthermore, ROS can be toxic to plants, causing damage to plant cells. ROS is produced in the chloroplasts, mitochondria, peroxisomes, and ectoplasts. Furthermore, the superoxide anion generates H2O2 under the action of superoxide dismutase (SOD). ROS production in ectoplasts may play a major role in biological and abiotic stresses[8]. Respiratory burst oxidase homolog (RBOH) enzymes are the most important extracellular (ROS)-related enzymes. Expression of the RBOHD gene containing a mutation in the bik1-specific phosphorylation site of the rbohD mutant inhibits excessive oxidative damage, and the mutant plants are more susceptible to pathogens, indicating that RBOHD and bik1-mediated phosphorylation contribute to plant immunity[9]. The sensor kinase protein MoOsm1 in rice can maintain ROS balance by phosphorylation and dephosphorylation transformation, thereby improving the resistance of rice to rice blasts[10]. The RbohA and RbohB expressions are upregulated to resist pathogen infection during invasion by Magnaporthe oryzae and powdery mildew (Blumeria graminis f. sp. hordei.) in rice and barley, respectively. Furthermore, antisense strains of the RBOH gene render plants susceptible to diseases[11,12].
Plants generate different hormone signaling pathways to resist pathogen infection. Jasmonic acid (JA) is an important lipid hormone in plant immunity against necrotrophic pathogens. JA can be catalyzed to form methyl jasmonate (MeJA) and other polymers and metabolites, and it can form derivatives through alkylation, glycosylation, or when combined with amino acids[13]. The role of JA signaling pathway genes in resistance to biotic stresses has been widely reported. For example, deletion of the GhOPR9 gene increased the susceptibility of cotton to Verticillium dahliae[14]. Overexpression of VvAOS and VvCOI genes can significantly reduce the hyphal area of Botrytis cinerea after infecting grapes[15]. Inhibition of the LOX gene expression in tobacco increased susceptibility to Phytophthora nicotianae infection[16]. LOX, COI, and OPR genes in wheat are expressed in response to aphid invasion. Downregulation of COI1 and JAZ in cotton increased susceptibility to aphid invasion[17,18]. A previous study reported that JA signal expression correlates with temperature. Low temperatures reduce the resistance of rice to M. oryzae and inhibit JA biosynthesis and signal transduction, increasing the susceptibility of rice to diseases[19].
The NAC transcription factor family is one of the largest transcription regulatory factors in plants and exists in various plant species, such as Arabidopsis thaliana (Arabidopsis), rice, tobacco, and chrysanthemums[20−23]. Previous studies found a highly conserved amino acid sequence, named the NAC domain, at the N-terminus of the protein sequence encoded by the genes in Petunia hybrida (NAM), Arabidopsis (ATAF1/2), and CUC2 (cup-shaped cotyledon)[24,25]. The NAC is divided into five subdomains: A, B, C, D, and E. The B and E subdomains are not highly conserved, whereas the C and D are highly conserved and may be the primary structures involved in the DNA-binding process[26−28]. The NAC domain has a symmetric dimer structure different from the general double-helix structure. The C-terminus of NAC transcription factors contains a transcriptional activation domain that regulates transcription. The C-termini of NAC proteins are diverse; however, some contain partially conserved motifs. NAC TFs can be divided into typical NAC TFs and atypical NAC TFs considering the characteristics of their C-terminal structure[29,30]. The atypical NAC TFs contain conserved motif fragments or C-terminal deletions. NAC transcription factors with the same motif fragments are similar in their physical structure and the biological functions they perform[31].
The biological functions of the NAC transcription factor family mainly regulate plant growth and development, plant response to abiotic and biotic stresses, regulation of plant secondary growth, and hormone signal transduction[32]. ATAF1 transcription factor in Arabidopsis is a negative regulator of the plant defense response to infection by biotrophic fungi and bacterial pathogens. According to a previous study, overexpression of ATAF1 exhibited higher sensitivity to B. cinerea[33]. Overexpression of OsNAC6 exhibited significant effects on dehydration, high salt stress, and tolerance to M. oryzae in rice[34]. Plants overexpressing OsNAC111, OsNAC066, and OsNAC58 had increased resistance to M. oryzae, Xanthomonas oryzae causing rice bacterial blight, and the sensitivity of rice to X. oryzae, respectively[35−37]. Furthermore, the mOsNAC60 mutation increased the sensitivity of rice to M. oryzae[35−37]. GbNAC1 positively regulates verticillium wilt resistance in cotton[38]. The regulation of pathogen invasion by NAC transcription factors often triggers plant defense responses through hormone-signaling molecules. For example, AtNAC019/AtNAC055, RIM1, and TaNAC6s regulate B. Cinerea, Rice dwarf virus, and B. graminis f. sp. tritici, respectively, via the JA signaling pathways[39−41]. Significant progress has been made in research on the biological functions of NAC genes in model plants. However, there are few studies on the functions of NAC family genes in chrysanthemums and no reports on the regulatory processes of biotic stress.
Therefore, the present study aimed to investigate the regulatory mechanisms of CmNAC083 in response to black spot infection in chrysanthemums. We cloned the CmNAC083 gene and determined its response to A. alternata infection. Furthermore, we evaluated the effect of CmNAC083 on black spot disease resistance of the genetically transformed chrysanthemum 'Jinba.' Through RNA sequencing, we found that CmNAC083 enhances the regulatory mechanism of chrysanthemum resistance to black spot disease. The present study lays a theoretical foundation for analyzing the role of NAC family genes in the resistance of chrysanthemums to black spot disease, and provides primary information for the breeding of new resistant varieties.
-
The chrysanthemum 'Jinba' used in the present study was obtained from the Chrysanthemum Germplasm Resources Conservation Center of the Nanjing Agricultural University, Nanjing, China. The material was grown under long sunshine (16 h light/ 8 h darkness) with temperature maintained at 25 °C and 70% relative humidity. Wild-type (WT) and transgenic materials were cut into a hole plate for 2 weeks and then transplanted into a small black square for cultivation for 2 weeks before treatment. Nicotiana Benthamiana was grown under long sunshine (16 h light / 8 h darkness), with temperature maintained at 25 °C and relative humidity of 70%.
CmNAC083 cloning and sequence analysis
-
In the present study, total RNA was extracted from chrysanthemum 'Jinba' leaves using the RNA isolation kit (Huayueyang, Beijing, China) according to the manufacturer's protocol. The PrimeScript reverse transcription kit (Takara Bio, Tokyo, Japan) was used to reverse-transcribe RNA into cDNA. The CmNAC083 sequence was downloaded from the chrysanthemum genome database to design primers (CmNAC083-ORF-F: 5′-ATGACAAAGCTAAATTTCTTCG-3′; CmNAC083-ORF-R: 5'-CTA-TAAACTATTCACTTCATCTTCA-3′). The open reading frame (ORF) of CmNAC083 was amplified, and the polymerase chain reaction (PCR) products were inserted into the Blunt plasmid of the cloning vector (Vazyme, Nanjing, China) for sequencing. The molecular weight and isoelectric point of CmNAC083 were predicted using the SWISS-PROT database (
https://web.expasy.org/compu-tepi/ ). The amino acid sequences of the AtNAC family members were downloaded from the Arabidopsis Information Resource website (www.arabidopsis.org/Blast/index.jsp ). The protein homologs of the CmNAC083 and AtNACs were compared using DNAMAN 9 software (Lynnon Biosoft Bioinformatic Solutions, San Ramon, CA, USA), and a phylogenetic tree was constructed using Molecular Evolutionary Genetics Analysis (MEGA) 5.1 software (MEGA, Mumbai, India) using the neighborhood connection method and 1,000 guided repeats. AtNAC genes includes the following: AtNAC002 (AT1G01720); AtNAC019 (AT1G52890); AtNAC041 (AT2G33480); AtNAC047 (AT3G04070); AtNAC055 (AT3G15500); AtNAC056 (AT3G15510); AtNAC059 (AT3G29035); AtNAC081 (AT5G08790); AtNAC083 (AT5G13180); AtNAC092 (AT5G39610); AtNAC100 (AT5G61430) genes.Prediction of CmNAC083 subcellular localization
-
The subcellular location of CmNAC083 was predicted using PlantPLOC (
www.csbio.sjtu.edu.cn/bioinf/plant-multi ). The CmNAC083 ORF sequence was amplified with CmNAC083-R4-F/R primers (CmNAC083-R4-F: 5′-TCCCCCGGGATGACAAAGC-TAAATTTCTTCG-3′; CmNAC083-R4-R: 5′-CCATCGATTAAACTATTCACTTCATCTTCAT-3′), in which an SmaI recognition site was included at the 5′ end of the forward primer, and a ClaI recognition site was included at the 5′ end of the Reverse primer. After double-digestion purification, the amplified product was inserted into the double-digested pORE-R4 and the vector contained the CaMV2 × 35S promoter-driven reporter green fluorescent protein (GFP) gene. P35S::CmNAC083-GFP (pORE-R4-CmNAC083) and p35S:: GFP (pORE-R4) were transformed into Agrobacterium strains using an electric shock. The constructed fusion protein was introduced into tobacco epidermal cells through needle injection for infection, considering the Agrobacterium infiltration method. After culturing in the dark for 24 h, the material was stored for 48 h under long-term sunshine conditions. GFP activity of the material was observed and imaged using a laser scanning confocal microscope (Carl-Zeiss, Munich, Germany).CmNAC083 transcriptional activation activity
-
The transcriptional activity of CmNAC083 was examined using the yeast two-hybrid system. The CmNAC083 ORF sequence was amplified using CmNAC083-BD-F/R primers (CmNAC083-BD-F: 5′-GGAATTCATGACAAAGCTAAATTTCTTCG-3′; CmNAC083-BD-R: 5′-CGGATCCGCTATAAACTATTCACTTCATCT-3′), and then digested using EcoRI and BamHI restriction endonuclease and introduced into the pGBKT7 vector to construct pGBKT7-CmNAC083. The plasmids pGBKT7-CmNAC083, pCL1 (positive control), and pGBKT7 (negative control) were transformed into Saccharomyces cerevisiae strain Y2HGold (Takara Bio, Tokyo, Japan) according to the manufacturer's instructions. The transformed strains containing pGBKT7-CmNAC083 or pGBKT7 were grown on the synthetic-defined (SD)/-tryptophan (Trp) medium, and the transformed strains containing pCL1 were grown on the SD/-Leucine medium. The transformed yeast cells were grown in SD/-Histidine-Adenine medium with X-α-Gal and SD/-Histidine-Adenine medium. Next, the transformed yeast cells were cultured at 30 °C for 3–4 d. Finally, the blueness of the yeast strain was observed. The present study also used the transient transformation of Arabidopsis protoplasts to verify the CmNAC083 transcriptional activity. The CmNAC083 ORF sequence was amplified using CmNAC083-pENTR1A-forward and -reverse primers (CmNAC083-pENTR1A-F: 5′-CGGGATCCGGATGACAAAGCTAAATTTCTTCGAA-3′; CmNAC083-pENTR1A-R: 5′-ATAAGAATGCGGCCGCGATAAACTATTCACTTCA-3′), and was digested using BamHI and NotI restriction endonuclease and then was introduced into the pENTR1A vector to construct pENTR1A-CmNAC083. The pENTR1A-CmNAC083 plasmid was recombined using the GAL4-DB plasmid using the LR recombination method. GAL4-DB-ARF5 and the empty vector GAL4-DB were used as the positive and negative controls. GAL4-LUC was used as the reporter co-transformation plasmid. High-concentration plasmids were extracted for protoplast transformation using a Plasmid Midi Kit (Qiagen, Hilden, Germany). The extraction and transformation methods for Arabidopsis protoplasts were according to Wang et al.[42]. After incubation for 16–18 h overnight, 20 mM sodium fluorescein (Sigma-Aldrich, Mannheim, Germany) was added to each sample. The indicated luciferase (LUC) activity was measured using a GloMax enzyme marker (GloMaxR20/20; Promega, Madison, WI, USA), and a CCD camera (Tanon 5200; Tanon Science, Shanghai, China) was used to capture the images.
A. alternata inoculation and disease severity assessment
-
A. alternata F20 was cultured on potato glucose agar medium (Sigma-Aldrich) at 28 °C in darkness for 4–5 d. An appropriate amount of bacterial cake was picked from the growth edge of the bacteria in the culture medium and put into the potato glucose broth medium (PDB [Sigma-Aldrich]) for shaking propagation for 24 h at 28 °C. Next, the cultivated F20 was broken, transferred into the new PDB shaking propagation for 24 h at 28 °C, and used for inoculation treatment. Next, 1 mL of mycelia from the culture medium was collected to form a ball and inoculated on the back of the third fully expanded leaf using a toothpick. Each leaf was inoculated twice. Each line was inoculated with nine strains, and the experiment was independently repeated in triplicate. The inoculated seedlings were placed in a storage box and cultured for 48 h in a long sunshine (16 h light / 8 h dark) light incubator at 28 °C and a relative humidity of 70%. ImageJ software (National Institutes of Health and the Laboratory for Optical and Computational Instrumentation, University of Wisconsin, WI, USA) was used to measure the diseased area of the inoculated leaves 2 d after inoculation. Disease severity was evaluated by determining the average lesion area.
qRT-PCR analysis
-
Quantitative reverse-transcription polymerase chain reaction (qRT-PCR) was carried out on a LightCycle 96 real-time PCR system (Roche, Basel, Switzerland) using the SYBR Premium Ex Taq II kit (Takara Bio, Tokyo, Japan) according to the manufacturer's instructions. The qRT-PCR reaction procedure includes 40 cycles of pre-denaturation at 95 °C for 2 min, and at 95 °C for 15 s, 60 °C for 15 s, and 72 °C for 15 s. The qRT-PCR data were obtained from three biological and three technical replicates for each sample. The expression level was calculated using the 2−ΔΔCᴛ method[43]. CmEF1α (GenBank: KF305681) was used as an internal reference gene. The primer pairs used for the qRT-PCR analysis are presented in Supplemental Table S1.
Chrysanthemum genetic transformation
-
The construction method for the pORE-R4 vector is described above. The constructed fusion plasmid was transformed into Agrobacterium tumefaciens EHA105 strain using the electric shock method and then transferred into chrysanthemum 'Jinba' using the leaf-disk infection method[38]. After leaf disc sprouts and roots, the positive strains were identified at the DNA level using the primer pair pORE-R4-F/CmNAC083-R4-R (pORE-R4-F: 5′-GACGCACAATC-CCACTATCC-3′), and the relative expression level of CmNAC083 was determined using qRT-PCR analysis. Each sample was tested using three biological and three technical replicates. The transgenic chrysanthemum and WT tissue culture seedlings were cultured at 25 °C under long sunshine (16 h light / 8 h darkness) after growing in the hole plate for 2 weeks.
Virus-induced gene silencing
-
A previously reported virus-based miRNA expression system[44−47] was used to silence CmNAC083 in chrysanthemum. Web MicroRNA Designer (
wmd3.weigelworld.org/ ) was used to design the miRNA sequences (TAAGTAATGTACCGCAAGCTC) for the target genes. RS300 plasmid was used as a template and the following primers (I miR-s: 5′-gaTAAGTAATGTACCGCAAGCTCtctctcttttgtattcc-3′; II miR-a: 5′-gaGAGCTTGCGGTACATTACTTAtcaaagaga-atcaatga-3′; III miR*s: 5′-gaGAACTTGCGGTACTTTACTTTtcacaggtcgtga-tatg-3′; IV miR*a: 5′-gaAAAGTAAAGTACCGCAAGTTCtctacatatatattcct-3′) were used to amplify amiRNA precursors. RS300R-CmNAC083 was digested using XbaI and KpnI restriction endonucleases, introduced into the cabbage leaf-curl gemini-virus vector (CaLCuV), and transformed into Agrobacterium tumefaciens strain GV3101. PCVB and CaLCuV (control) or CaLCuV-CmNAC083 were mixed evenly in 1:1 ratio (v/v), incubated with infiltration buffer at 28 °C for 3 h and then vacuumed (0.7 MPa) for 5–10 min. The plants were placed in the dark at 10 °C for 1–3 d and then transferred to a flowerpot containing a mixture of nutrient soil and vermiculite 1:3 (v/v), and grown under long sunshine conditions (16 h light/ 8 h darkness) for 3 weeks. The primer pair (II miR-a; CaLCuV-R: 5′-GCAACGGACAGATCCTATGTAAC-3′) was used to identify the positive strains of CaLCuV-CmNAC083 by removing the newly grown third leaf from the top[48]. Positive strains were screened for the determination of the CmNAC083 expression. Two independent experiments were conducted, and at least nine positive WT or transiently silenced strains were used for inoculation.Determination of physiological indexes
-
The contents of endogenous JA were measured using plant JA elisa kit from the Shanghai Yuanju Biotechnology Center (Shanghai, China). The anti-superoxide anions and SOD activities were measured using a test kit from the Nanjing Jiancheng Institute of Biological Engineering, Nanjing, China, according to the manufacturer's instructions. Diaminobenzidine (DAB) staining was performed after inoculation with WT and transgenic strains. First, 0.1 g DAB powder was dissolved in 100 mL of 10 mM phosphoric acid buffer (pH 7.8 [FUJIFILM Wako Pure Chemical Corporation, Tokyo, Japan]), and the pH was adjusted to 3.8. The leaves were completely immersed in the dye solution, placed on a shaking table, and slowly shaken in the dark for 8–12 h until brown spots were observed. The dyed leaves were fully immersed in 80% alcohol for 15 min in boiling water. The leaves were then completely immersed in 95% alcohol for 15 min in boiling water; this was repeated twice until the color of the tissue faded. The decolored tissue was completely soaked in anhydrous ethanol and stored in a refrigerator at 4 °C until imaging.
RNA extraction, transcriptome sequencing, and bioinformatics analysis
-
The third fully expanded leaf of WT and OE-CmNAC083 (OE-128, OE-156) was inoculated with A. alternata for 48 h to study the possible signaling pathway of CmNAC083 regulating black spot disease. Samples of the third fully expanded leaf after inoculation and those of the control group were subjected to RNA-sequencing analysis. Each sample contained three biological replicates. The transcriptome was sequenced using an Illumina sequencing platform (GeneDenovo Biotechnology Co., Ltd., Guangzhou, China). The functional annotation of single genes was based on the Kyoto Encyclopedia of Genes and Genomes (KEGG;
www.kegg.jp ) and Gene Ontology (GO;www.geneontology.org ). The fragments per kilobase of script per million fragments mapped (FPKM) method was used to calculate the expression level of a single gene [49]. Differential expression analysis was performed using DESeq2 software. Genes with a false detection rate (FDR) lower than 0.05 and a single gene with | log2 FoldChange | > 1 were considered differentially expressed genes (DEGs)[50]. FDR was obtained by correcting the p-value of the significant difference. Venn maps and KEGG pathway enrichment analyses were performed using the online Omic Intelligent tool (www.omicsmart.com ).Statistical analysis
-
SPSS v17.0 (SPSS Inc., Madison, IL, USA) was used for one-way analysis of variance in all statistical analyses, and Duncan's multiple range test was used to determine significant differences. Statistical significance was set at p < 0.05.
-
After inoculation of chrysanthemum 'Jinba' with A. alternata at different times, the samples were sent for high-throughput sequencing. The results revealed that the effect of A. alternata inoculation on CmNAC083 expression was significantly different (Fig. 1a). The CmNAC083 ORF sequence (Fig. 1b) was cloned to 660 bp (Supplemental Material 1). Phylogenetic tree analysis of the AtNACs homologous to CmNAC083 indicated that CmNAC083 exhibited the closest homologous relationship with AtNAC041 (AT2G33480) and AtNAC083 (AT5G13180) in Arabidopsis (Fig. 1c). CmNAC083- protein domain analysis revealed that its N-terminus contains a highly conserved domain of NAC family genes (Fig. 1d). The CmNAC083 expression at different stages of A. alternata infection was analyzed using qRT-PCR to explore whether CmNAC083 was involved in the infection of the Chrysanthemum black spot pathogen. CmNAC083 was significantly and continuously induced to express at 6 h and reached the expression peak at 48 h after inoculation with A. alternata. The CmNAC083 expression in the inoculated sample was 8.65-fold higher than that in the control sample (Fig. 1e), indicating that CmNAC083 may participate in the response to the chrysanthemum black spot disease. The qRT-PCR analysis of the CmNAC083 expression pattern in different tissues revealed that CmNAC083 expression was not tissue-specific in chrysanthemum 'Jinba'. However, the CmNAC083 expression level was higher in the stem and bud during the vegetative growth period than that in other tissues during the same period. Furthermore, CmNAC083 expression was higher in the stems and roots during the flowering stage (Fig. 1f & g).
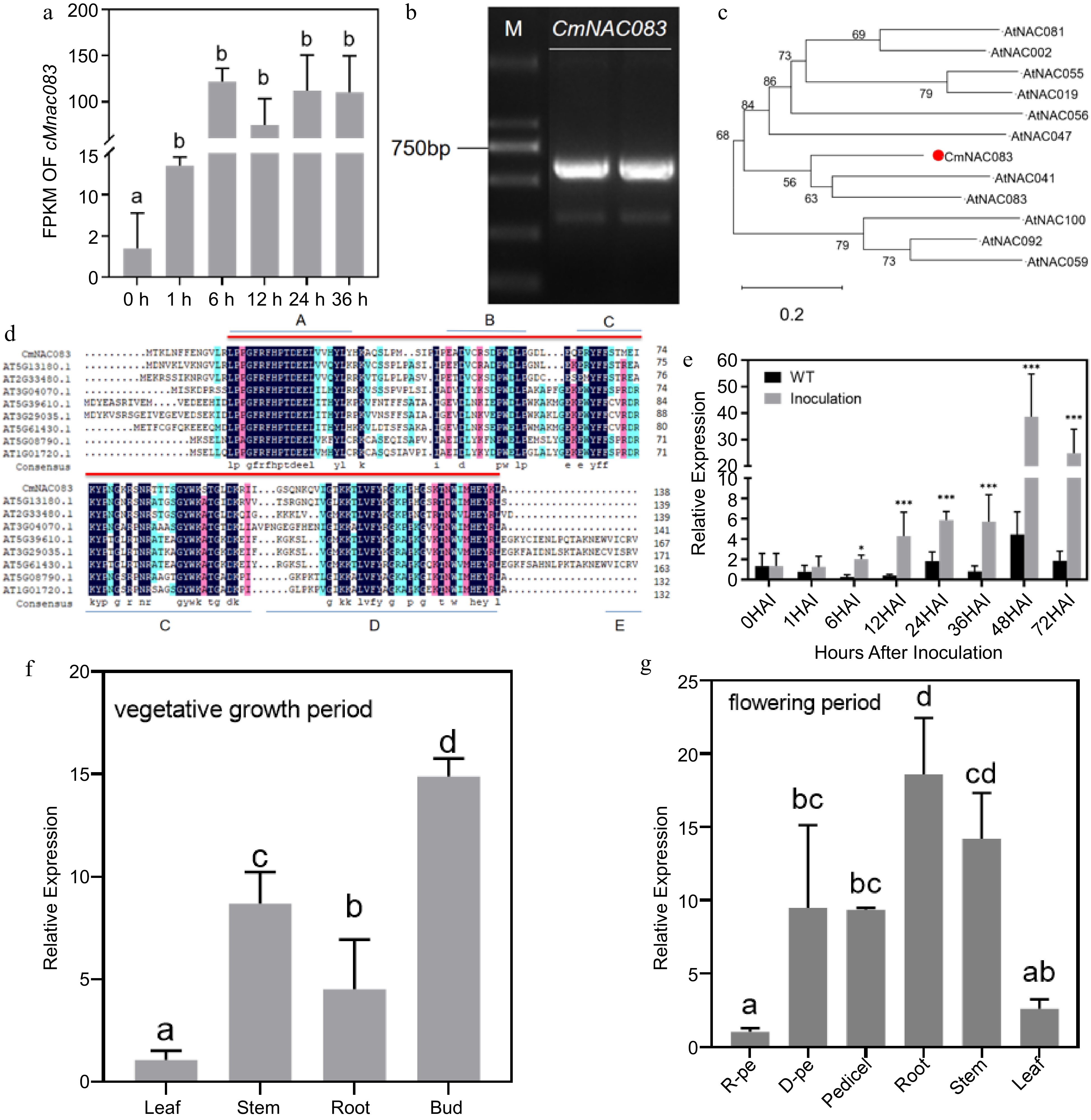
Figure 1.
Cloning and expression analysis of CmNAC083. (a) Analysis of the fragments per kilobase of script per million fragments mapped of CmNAC083 at 0, 1, 6, 12, 24, and 36 h after inoculation with Alternaria alternata. (b) The results of electrophoresis of polymerase chain reaction products of the CmNAC083 clone Note: M: DL2000. (c) The phylogenetic analysis of CmNAC083 amino acid and Arabidopsis NAC family amino acid sequences. (d) Comparison of CmNAC083 amino acid sequence with the homologous protein in Arabidopsis. (e) CmNAC083 gene expression during A. alternata infection at different times. Wild-type not inoculated. The statistical analyses were performed using Student's test (* p < 0.05, *** p < 0.001). (f) CmNAC083 gene expression in different tissues during the vegetative growth period. (g) CmNAC083 gene expression in different tissues (R-pe, ray florets petals; D-pe, and disc florets petals) during the flowering period. Data are presented as the mean ± standard error of the mean from the three biological replications. Statistical analyses were performed using Duncan's test. Different lowercase letters represent a significant difference between treatments at (p < 0.05).
CmNAC083 as a transcription inhibitor
-
CmNAC083 was cloned into the p35S::GFP vector to confirm if it is a transcription factor. The fusion genes p35S::CmNAC083-GFP and p35S::GFP(control) were introduced into tobacco epidermal cells via Agrobacterium infection. The green fluorescence signal of the CmNAC083-GFP fusion protein was only expressed in the nucleus, whereas the control GFP protein was expressed in the nucleus and the cytoplasm (Fig. 2a), consistent with the results of the subcellular location of CmNAC083 predicted by the plantPLOC website. We performed a transcriptional activation validation experiment in yeast to detect the CmNAC083 transcriptional activity. The results revealed that CmNAC083 had no transcriptional activation (Fig. 2b). We further verified the CmNAC083 transcriptional activity through transient transformation experiments using Arabidopsis protoplasts. The recombinant plasmid obtained by introducing CmNAC083 into the GAL4-DB vector and the 5XGAL4-LUC plasmid were used as the effector and reporter genes, respectively (Fig. 2c). The GAL4-DB empty vector and GAL4-DB-ARF5 were used as negative and positive controls, respectively. Compared to the GAL4-DB empty vector and the positive control GAL4-DB-ARF5, the protoplast-transformed GAL4-DB-CmNAC083 had lower LUC activity (Fig. 2d & e), indicating that CmNAC083 may have transcriptional inhibition activity in plant cells and CmNAC083 may act as a transcription inhibitor in chrysanthemum.
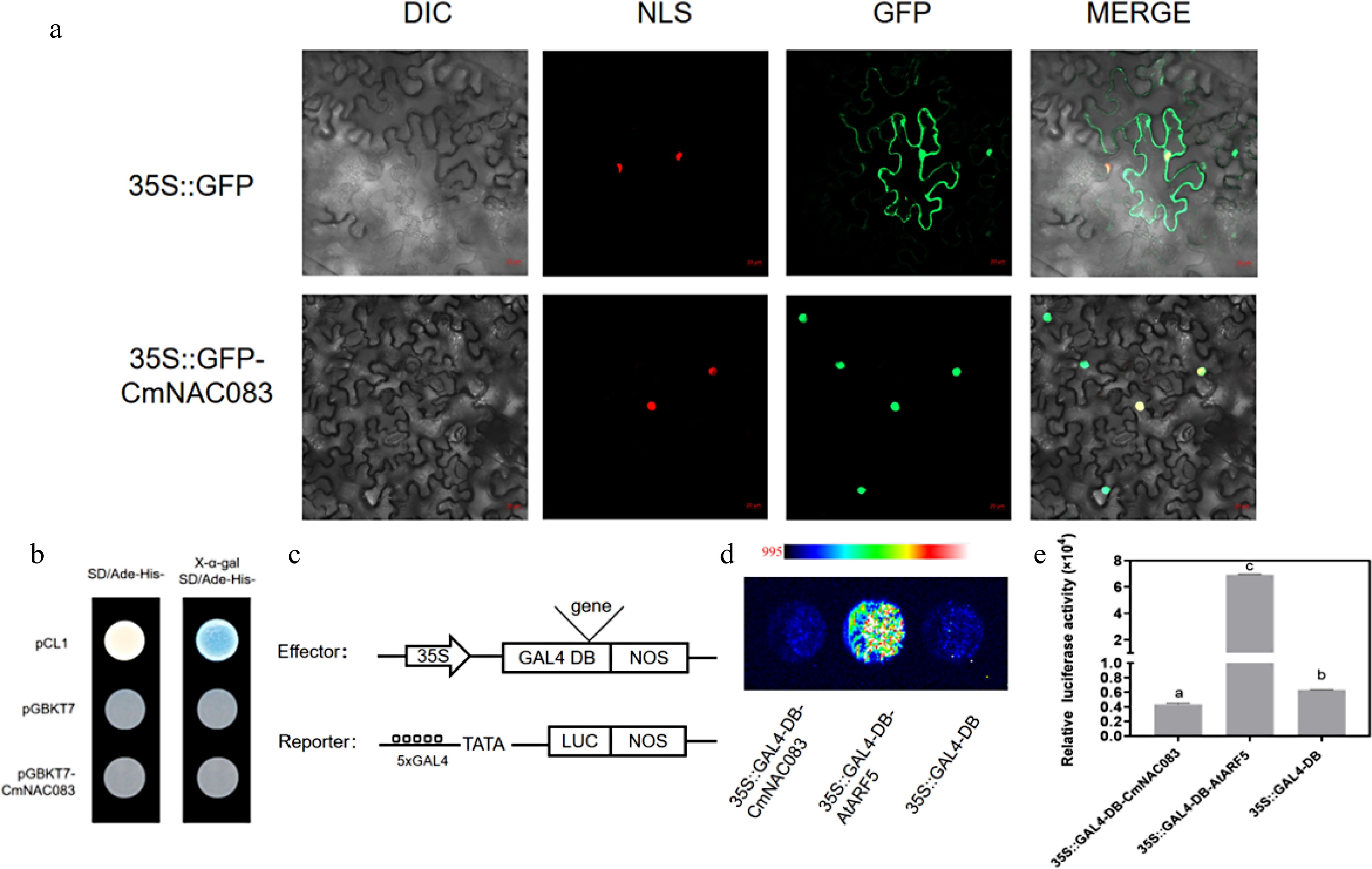
Figure 2.
CmNAC083 subcellular localization and transactivation analysis. (a) The subcellular localization of CmNAC083 in Nicotiana benthamiana cells. DIC, bright-field image; NLS, nuclear localization signal exhibited by the red fluorescent protein; GFP, green fluorescent protein; merged, overlay images. Scale bars = 20 µm. (b) CmNACO83 transcriptional activation analysis in yeast cells. Positive control: pCL1; negative control: pGBKT7. (c) Simple structure diagrams of the effector and reporter in the protoplast transfection experiment. (d) Fluorescence images of protoplasts after the addition of 20 mM fluorescein sodium. The gradual increase in fluorescence signal from blue to red. Positive control: ARF5; negative control: GAL4-DB. (e) Quantitation of luciferase intensity of each sample. Positive control: ARF5; negative control: GAL4-DB. Data are presented as the mean± standard error of the mean (n = 3). Significant differences were determined using Student's test at (*** p < 0.001).
CmNAC083 overexpression increases chrysanthemum resistance to A. alternata
-
Overexpression transgenic lines (OE-CmNAC083) driven by the 35S promoter were obtained through chrysanthemum genetic transformation to study the role of CmNAC083 in response to A. alternata infection in chrysanthemums. Transgenic lines were used to study the effect of CmNAC083 on Chrysanthemum resistance to black spot disease. qRT-PCR confirmed the transcript abundance of CmNAC083 in the transgenic lines. The CmNAC083 expression in the OE-128 lines was 11.6-fold higher than that in the WT, and the expression of CmNAC083 in the OE-156 lines was 29.4 times as high as that in the WT (Fig. 3a). The results of the effect of A. alternata inoculation treatment on transgenic plants revealed that OE-CmNAC083 plants were resistant to A. alternata. The disease severity was lower in the overexpressing strains than that in the WT (Fig. 3b). The lesion area of CmNAC083 overexpression strains was significantly smaller than that of WT (Fig. 3c). The average lesion area of the WT plants was 0.946 cm2, whereas that of the overexpression lines OE-128 and OE-156 were 0.282 and 0.283 cm2, respectively, which was 0.3-fold that of the WT plant. There were no significant differences in root length and plant height between the overexpression lines and WT plants (Supplemental Fig. S1, Supplemental Table S2).
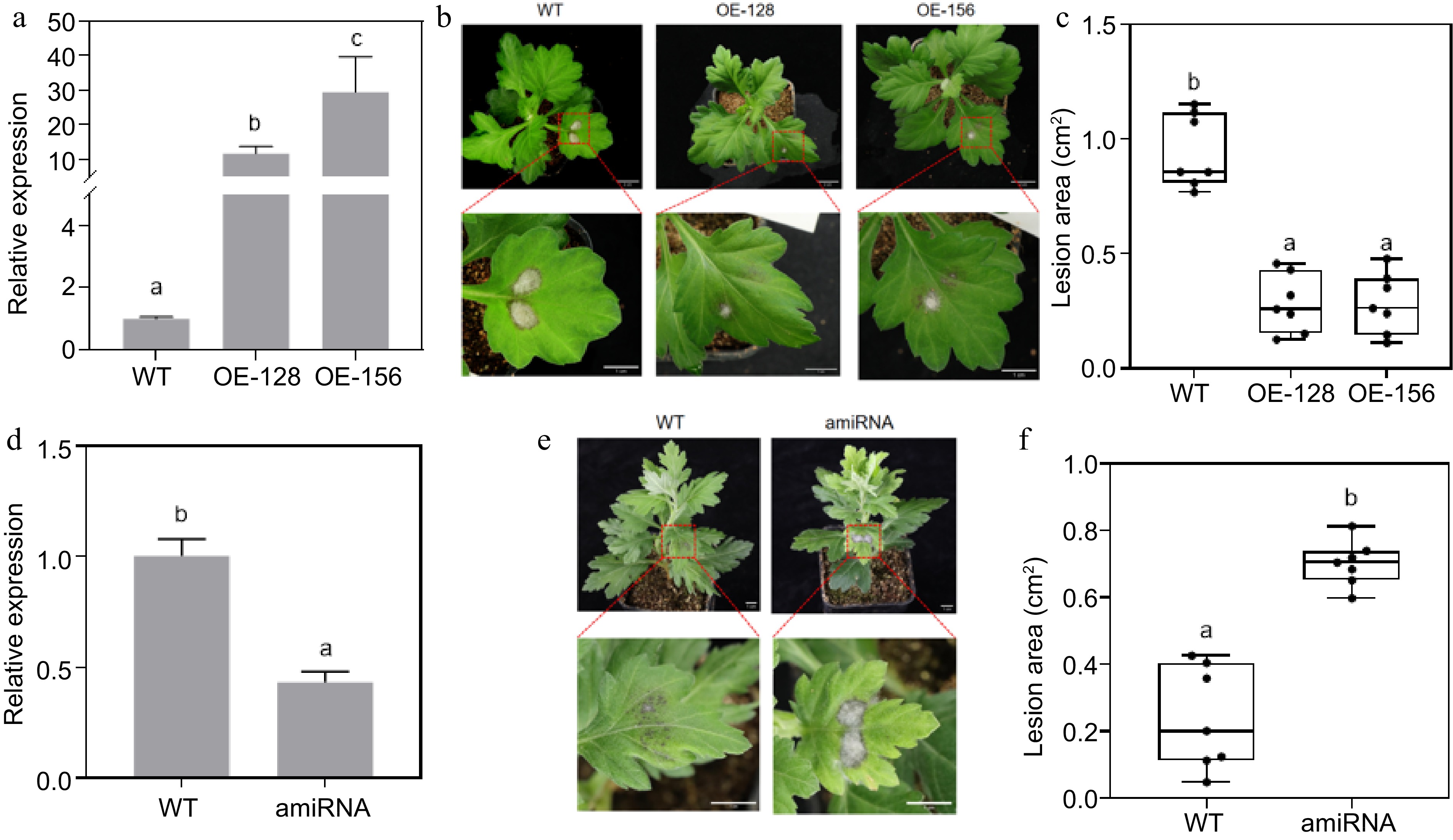
Figure 3.
Resistance analysis of CmNAC083 overexpression and transient silencing lines. (a) Identification of CmNAC083 overexpression transgenic lines at the RNA level. (b) Plant phenotype observation of transgenic lines and wild-type (WT) plants (1 month). Overall and drawings scale bars were = 2 cm and = 1 cm, respectively. (c) Area of diseased spots of the WT and transgenic lines inoculated with Alternaria alternata. (d) Identification of CmNAC083 instantaneous silencing transgenic lines at the RNA level. (e) Plant phenotype observation of WT-CaLCuV and instantaneous silencing transgenic lines (2 months). Scale bars = 1 cm. (f) Area of diseased spots of the WT-CaLCuV and instantaneous silencing transgenic lines inoculated with A. alternata. WT: WT-CaLCuV; amiRNA: WT-CaLCuV- CmNAC083. Data represent the mean ± standard error of the mean from three biological replications. Statistical analyses were performed using Duncan's test, and different lowercase letters represent a significant difference between treatments at (p < 0.05).
A modified CaLCuV was used to silence the CmNAC083 gene instantaneously through infection using Agrobacterium to determine the role of CmNAC083 in response to A. alternata infection. The transcriptome abundance of the CmNAC083 gene in the silenced lines was significantly lower than that in the WT plants (Fig. 3d). The CmNAC083 gene silencing resulted in a susceptible phenotype (Fig. 3e). The spot area of the CmNAC083 transient silenced strain was significantly increased compared with that of the control WT-CaLCuV (Fig. 3f). The average lesion area of the control WT-CaLCuV was 0.204 cm2, whereas that of the instantly silenced strain was 0.731 cm2, which was 3.6-fold that of the control. Overall, the results indicated that CmNAC083 conferred resistance to A. alternata infection in chrysanthemums.
Role of CmNAC083 in regulating black spot disease in chrysanthemums
-
One-month-old transgenic seedlings (OE-128, OE-156) and WT seedlings were inoculated with A. alternata for 48 h to determine the possible regulatory mechanism of CmNAC083 affecting the infection of black spot pathogens. DEGs were analyzed by comparing the control group with the WT and overexpression groups. The sequencing results revealed that there were 519 DEGs (456 upregulated and 63 downregulated) in WT compared with those of OE-128 and 803 DEGs (648 upregulated and 155 downregulated) in WT compared with those of OE-156 (Fig. 4a). After inoculation with A. alternata, 8,926 genes (5,366 upregulated and 3,560 downregulated) were differentially expressed in WT plants (WT-vs-WT-Aa). Furthermore, 10,505 genes (5,708 upregulated and 4,797 down-regulated) and 8,732 genes (4,613 upregulated and 4,119 downregulated) were differentially expressed in transgenic lines OE-128 (OE-128-vs-OE-128-Aa) and OE-156 (OE-156-vs-OE-156-Aa), respectively. After inoculation with A. alternata, 1,230 genes (840 upregulated and 390 downregulated) of the overexpression lines OE-128 and WT (WT-Aa-vs-OE-128-Aa) were differentially expressed, and 1,049 genes (617 upregulated and 432 downregulated) of the overexpression lines OE-156 and WT (WT-Aa-vs-OE-156-Aa) were differentially expressed. Additionally, 320 DEGs were found in WT-vs-OE-128 and WT-vs-OE-156 pairwise comparisons (Fig. 4b). DEG analysis results of the control and treatment groups of the WT and overexpression strains are presented in Supplemental Fig. S2. The results revealed 48 (40 upregulated and eight downregulated) of the 519 (456 upregulated and 63 downregulated) genes differentially expressed in the WT and the overexpression line OE-128 were induced by the inoculation with A. alternata. Furthermore, 240 (163 upregulated and 77 downregulated) of the 803 (648 upregulated and 155 downregulated) genes differentially expressed in the WT and OE-156 were induced by inoculation with A. alternata, indicating that CmNAC083 affects the expression of genes related to disease response.
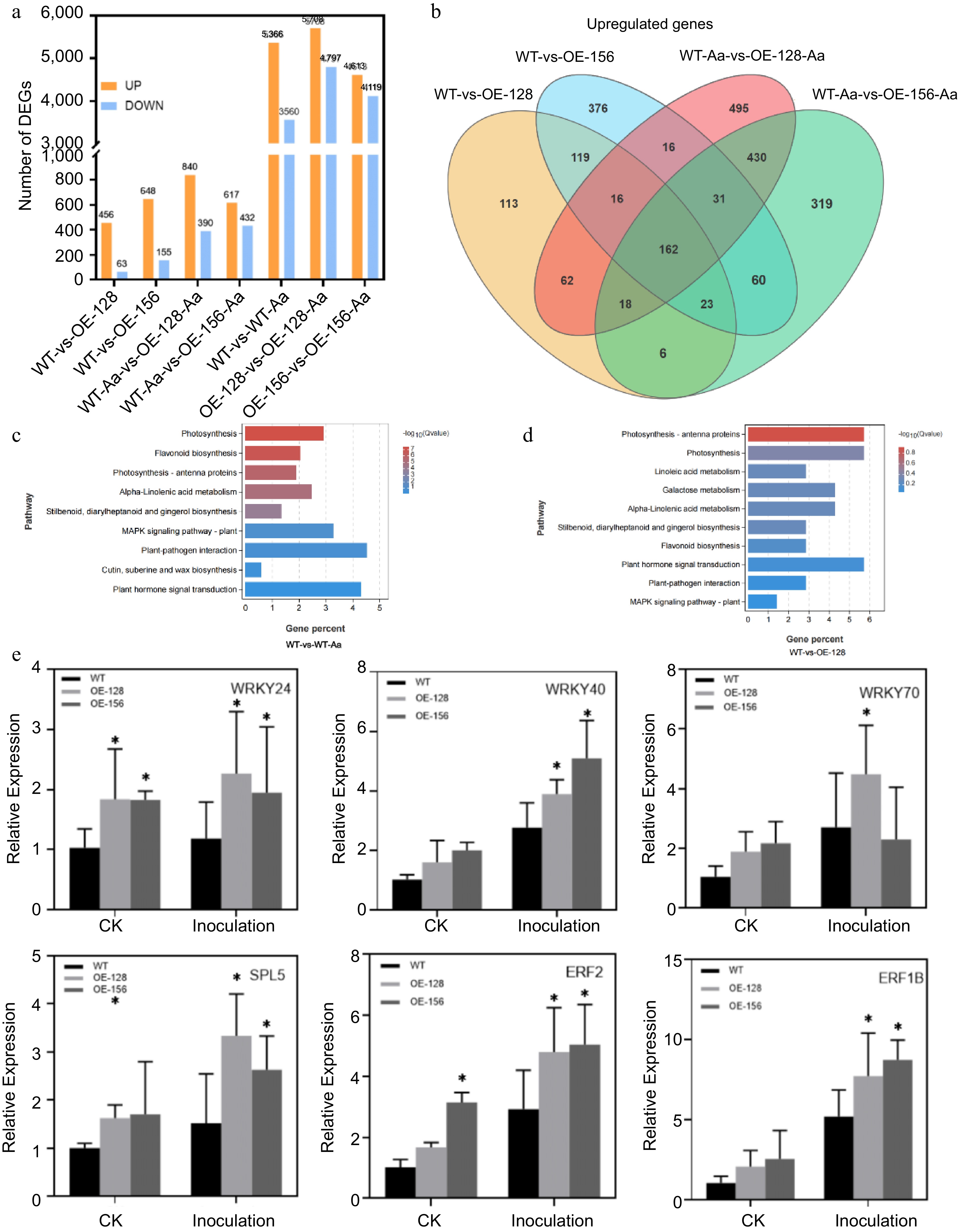
Figure 4.
Analysis of CmNAC083 transcriptome sequencing data. (a) The number of upregulated or downregulated genes in seven different comparison groups. (b) The Venn map reveals the overlap of differentially expressed genes among WT-vs-OE-128, WT-vs-OE-156, WT-Aa-vs-OE-128-Aa, and WT-Aa-vs-OE-156-Aa. (c) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of the differentially expressed genes in two comparisons (WT-vs-WT-Aa, WT-vs-OE-128) of the WT and overexpression transgenic lines under contrast and inoculation. (e) Verification of six differentially expressed transcription factor genes between WT and overexpression strains. Data are presented as the mean ± standard error of the mean from the three biological replications. Statistical analyses were performed using Duncan's test. Different lowercase letters represent a significant difference at (p < 0.05).
KEGG enrichment analysis was performed on the differential genes obtained from the seven comparative groups (WT-vs-OE-128, WT-vs-OE-156, WT-vs-WT-Aa, WT-Aa-vs-OE-128-Aa, WT-Aa-vs-OE-156-Aa, OE-128-vs-OE-128-Aa, and OE-156-vs-OE-156-Aa). The results exhibited that plant hormone signal transduction, plant-pathogen interaction, mitogen-activated protein kinase signal pathway-plant, alpha-linolenic acid metabolism, and other signaling pathways related to disease resistance were significantly enriched (Fig. 4c & d, Supplemental Fig. S3). Six differentially expressed transcription factors were randomly selected, and the transcriptome data were validated using qRT-PCR (Fig. 4e, Supplemental Table S3). The quantitative results obtained were consistent with the transcriptome analysis results, indicating the reliability of the transcriptome data.
Most significantly DEGs were enriched in the linolenic acid metabolism pathway and pathways related to disease resistance. Linolenic acid is a substrate in JA synthesis, and many transcription factors regulate JA biosynthesis. The JA response signaling pathway was also found in the plant hormone signal transduction pathway (Fig. 5a). The expression of several JA-related genes was detected in overexpression lines and WT plants using qRT-PCR. The results revealed that the expression of genes increased after inoculation with A. alternata, and the expression in the overexpression lines was significantly higher than that in the WT after inoculation (Fig. 5b). Then we measured endogenous JA content of the overexpression lines and the WT. The results revealed that the endogenous JA content of the overexpressing strains were higher than those of the WT after treatment with A. alternata (Fig. 5c). Additionally, we found that RBOHD, the key gene encoding respiratory burst oxidase in the ROS pathway of the plant-pathogen interaction signaling pathway, was different between the WT and overexpression strains before and after inoculation with A. alternata (Fig. 6a & b). DAB staining was performed on the overexpressing strains and WT plants after inoculation with A. alternata. The results revealed that the staining degree and range of the WT were deeper and larger, respectively, than that of the overexpression lines after inoculation (Fig. 6c). Furthermore, we measured anti-superoxide anion and SOD activities of the overexpression lines and the WT. The results revealed that the anti-superoxide anion and SOD activities of the overexpressing strains were higher than those of the WT after treatment with A. alternata (Fig. 6d & e).
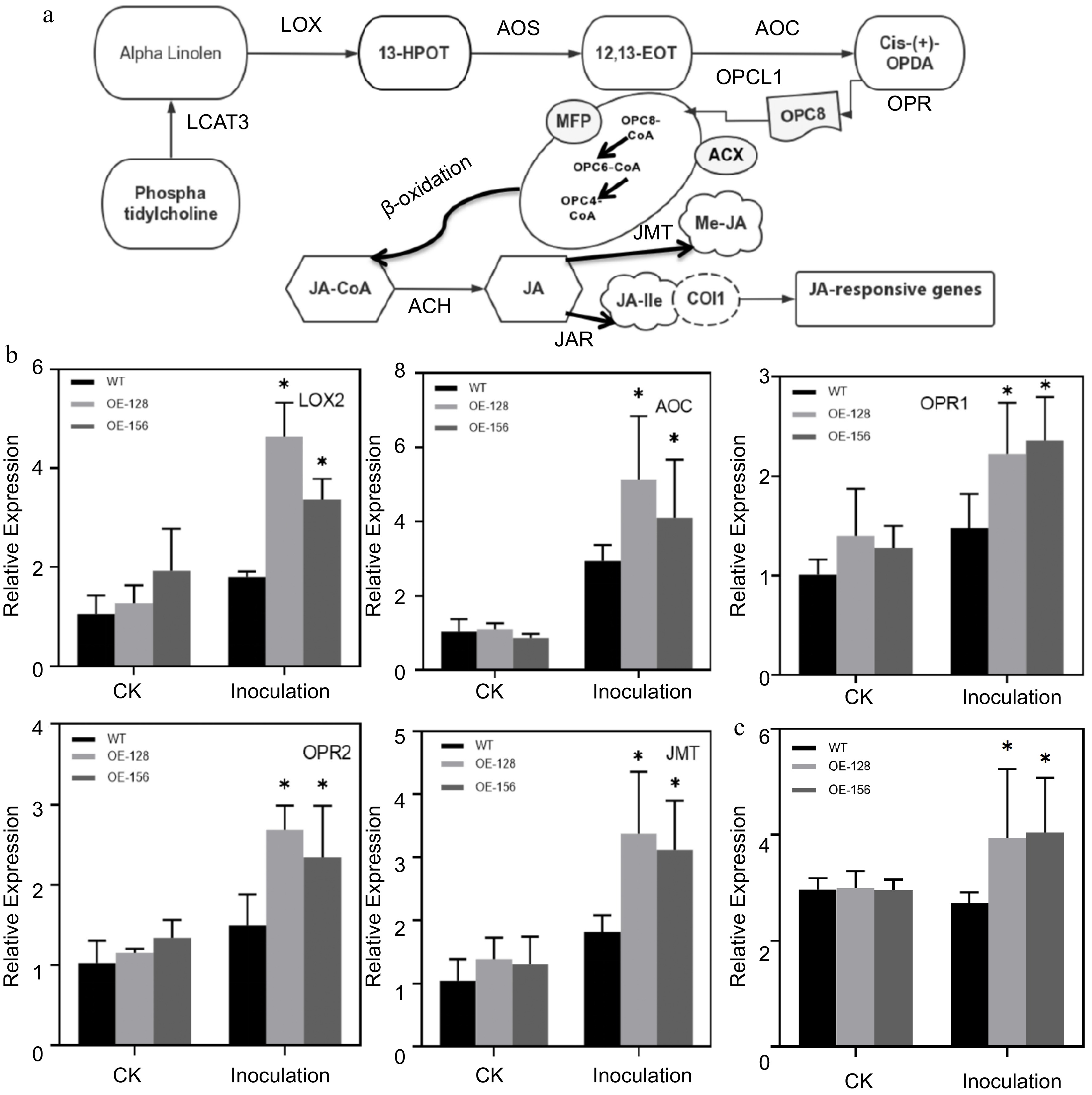
Figure 5.
Analysis of the involvement of JA signaling pathway in Alternaria alternata infection. (a) Simplified diagram of the JA signaling pathway. (b) Quantitative analysis of the JA Signaling pathway genes between WT and overexpression strains. Data are presented as the mean ± standard error of the mean from the three biological replications. Statistical analyses were performed using Duncan's test. Different lowercase letters represent a significant difference between treatments at (p < 0.05). (c) Determination of endogenous JA content after inoculating with A. alternata.
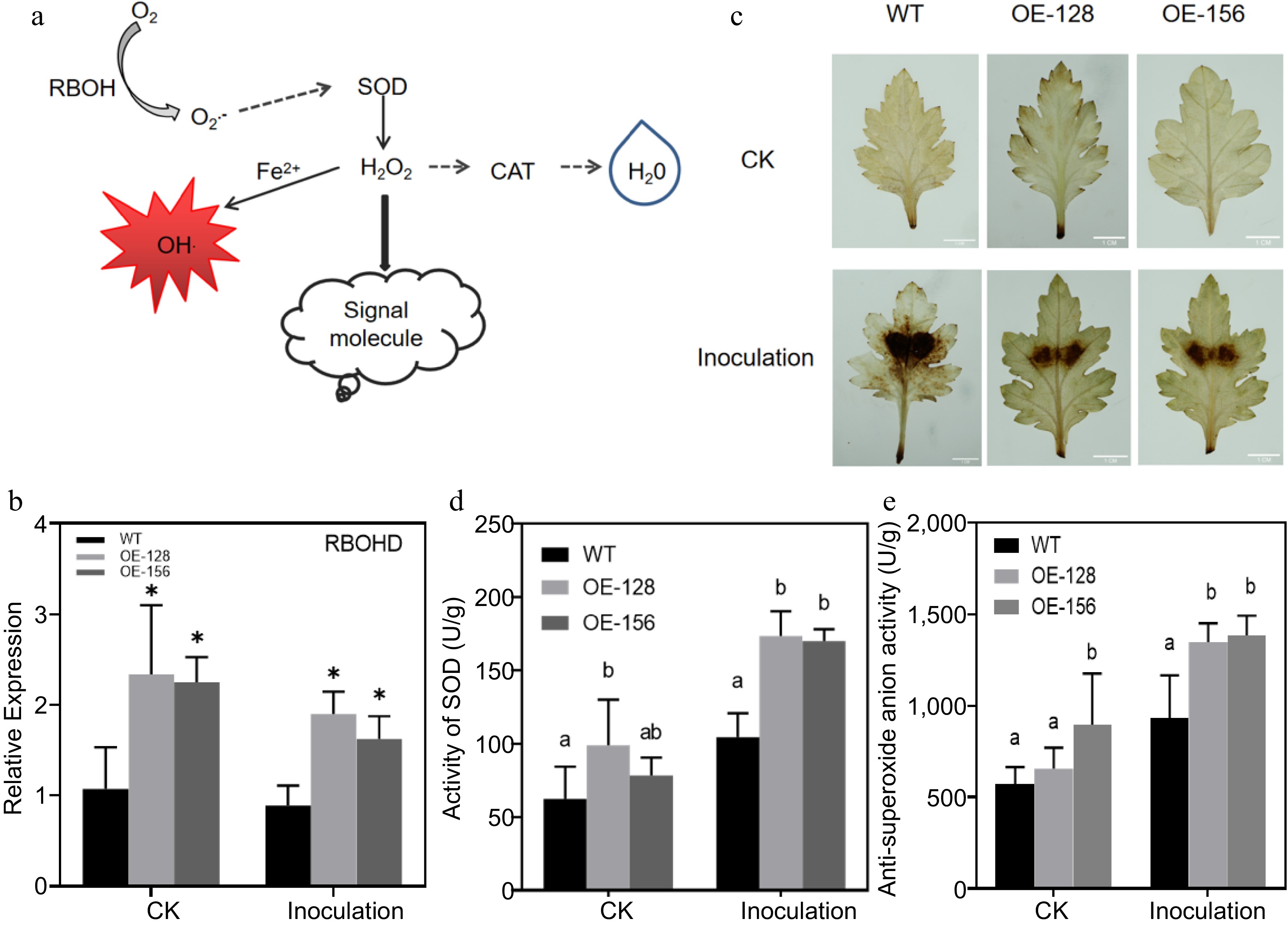
Figure 6.
Analysis of the role of the ROS signaling pathway in Alternaria alternata infection. (a) Simplified diagram of the ROS signal pathway. (b) Quantitative analysis of CmRBOHD between the wild-type (WT) and overexpression strains. (c) Diaminobenzidine staining for the transgenic and WT plants inoculated with A. alternata. (d) Determination of the superoxide dismutase activity in transgenic and WT plants inoculated with A. alternata. (e) Determination of anti-superoxide anion activity for the transgenic and WT plants inoculated with A. alternata. Values represent the mean ± standard error of the mean from three biological replications. Statistical analyses were performed using Duncan's test. Different lowercase letters represent a significant difference between treatments at (p < 0.05).
-
Abiotic stresses, such as diseases and insect pest invasion, affect the growth and development of plants. Chrysanthemum black spot is one of the fungal diseases of economic importance that seriously affects the ornamental value of chrysanthemums. Previous studies have revealed that the NAC transcription factor family genes play an important role in plant resistance to different pathogens.
In the present study, WT plants were inoculated with A. alternata, and high-throughput sequencing was performed. There was a significant difference among the WT plants in the expression of one NAC family member, the TF CmNAC083. Furthermore, we isolated and identified the CmNAC083 gene from the chrysanthemum 'Jinba' leaves and analyzed its protein structure, and the N-terminal had a conserved domain of the NAC family. Phylogenetic tree analysis of the NAC gene in Arabidopsis revealed that CmNAC083 has close homology with AtNAC041 and AtNAC083. In Arabidopsis, the interaction between AtNAC083 and AtNAC030 regulates the synthesis of xylem vessels synthesis of xylem vessels[51,52], whereas AtNAC041, bZIP1, and MYB46 directly regulate the expression of the cellulose synthase gene CSLA9[53]. However, the effects of the NAC083 gene on disease resistance mechanisms in different species have not yet been reported. We found that CmNAC083 was located in the nucleus through subcellular localization in tobacco, providing the first evidence that CmNAC083 is a transcription factor. The results of yeast transcriptional activation revealed that CmNAC083 does not exhibit transcriptional activation activity. We further verified the CmNAC083 transcriptional activity through the transient transformation of Arabidopsis protoplasts and found that CmNAC083 had transcriptional inhibitory activity. Therefore, we conclude that CmNAC083 is a transcription inhibitor that may affect disease resistance by directly binding to downstream genes and negatively regulating their expression. However, the exact underlying regulatory mechanisms require further investigation. Quantitative analysis of different tissues revealed that the CmNAC083 expression was higher in the stem and bud during the vegetative growth period and in the stem and root during the flowering period. This could be related to the regulation of xylem vessel development by AtNAC083 in Arabidopsis[54,55]. Xylem vessel development in the stems and roots at the flowering stage is relatively advanced, increasing the CmNAC083 expression. Inoculation with A. alternata to WT plants at different time points induced the continuous expression of CmNAC083, indicating that CmNAC083 responds to A. alternata infection. Therefore, we constructed the CmNAC083 overexpression lines and found that the overexpression conferred resistance to A. alternata, and the disease severity of the transgenic lines was significantly lower than that of the WT. We transiently silenced CmNAC083 to determine the resistance phenotype. The results revealed that CmNAC083 had contrasting phenotypes before and after silencing, consistent with the finding that CmNAC083 improves the resistance to black spot disease in chrysanthemums.
Plants respond to pathogens through various large-scale transcriptional changes, including increased synthesis of stress hormones when infected by pathogens. Plant hormone signaling pathways activate the co-expression of many defense response genes, thereby activating plant defense response[56,57]. Analysis of the transcriptome data revealed that the expression of some JA biosynthesis genes (LOX1, LOX2, AOC, OPR1, OPR2, and JMT) in the overexpression lines were significantly higher than those in the WT after inoculation with A. alternata. JA generally plays a positive regulatory role in plant disease resistance. Knockout of the GhOPR9 gene increased sensitivity to V. dahlia in cotton[13], where low temperatures inhibit JA biosynthesis and signal transduction and reduce rice resistance to rice blasts[18]. We measured the endogenous JA content of the overexpression lines and the WT after treatment with A. alternata. The results revealed that inoculation with A. alternata significantly increased the endogenous JA content of the overexpressing strains, consistent with the increased expression of JA biosynthetic genes observed above. Plants infected with pathogens can accumulate ROS to resist infection. When ROS accumulates to a certain limit, it activates the enzyme defense system of the plant, increased SOD enzyme activity, and removes excess ROS from the plant tissue[58,59]. The respiratory burst oxidase encoded by the RBOH gene is the primary source of ROS production in plants, and plants resist the invasion of pathogens by regulating ROS content. In a previous study, the expressions of RbohA and RbohB genes were upregulated, improving the rice resistance to M. oryzae infection[55]. Through transcriptome analysis, we found that the key gene for the ROS pathway, RBOHD, was more highly expressed in the overexpression line than that in the WT before and after inoculation with A. alternata, which may provide an immune effect for plants against pathogenic bacterial infection at the early growth stage. However, when ROS exceeds the range that a plant can withstand, it can cause irreversible mechanical damage[60]. DAB staining can be used to locate the tissue producing H2O2 in plants. After inoculation, the staining degree and range of the WT were deeper and larger than that of the overexpression lines, respectively, probably owing to the overexpression lines undergoing less mechanical damage after inoculation with the pathogen. The results of anti-superoxide anion and SOD enzyme activities in the overexpression lines and WT plants revealed that their activities in the WT plants after inoculation were significantly lower than those in the overexpression lines, indicating that the SOD activity of the overexpression lines increased after inoculation to remove excess ROS and anti-superoxide anion activity was enhanced, maintaining the ROS balance in the plant, and causing less mechanical damage. Overall, CmNAC083 overexpression affected the expression of JA synthesis and ROS regulation pathway genes in response to A. alternata infection.
We speculate that CmNAC083 may indirectly regulate JA biosynthesis and ROS outbreaks by inhibiting the expression of downstream JA and ROS signaling negative regulators as a transcriptional repressor, and regulates the defense response of chrysanthemums to black spot disease through the coordination of these two biological pathways. We conducted a preliminary study on the expression pattern of CmNAC083 under A. alternata treatment and its positive regulatory role in chrysanthemum black spot disease. Furthermore, we provided a theoretical basis for analyzing the mechanism of chrysanthemum resistance to black spot disease.
-
In conclusion, we isolated and identified the NAC transcription factor CmNAC083 from the chrysanthemum 'Jinba' leaves which conferred resistance to A. Alternata infection. The present preliminary study revealed that the regulatory effect could be related to the JA biosynthesis and ROS pathways. Although the specific CmNAC083 molecular regulatory mechanism in chrysanthemum black spots still needs to be studied, the present study provides a theoretical basis for improving the breeding of chrysanthemums for resistance to black spot disease.
-
The authors confirm contribution to the paper as follows: study conception and design: Fang W, Liu Y; data collection: Huang G, Dong B; analysis and interpretation of results: Jiang J, Chen S, Huang G, Dong B; draft manuscript preparation: Huang G, Liu Y. All authors reviewed the results and approved the final version of the manuscript.
-
Available data and materials can be obtained from the corresponding author according to reasonable requirements.
This work was supported by the National Key Research and Development Program of China (2020YFE0202900); Jiangsu Agriculture Science and Technology Innovation Fund (cx(20)1001); the Program for Key Research and Development, Jiangsu, China (BE2022417); the National Natural Science Foundation of China (32171854); the 'JBGS' Project of Seed Industry Revitalization in Jiangsu Province (JBGS(2021)094).
-
The authors declare that they have no conflict of interest. Jiafu Jiang and Sumei Chen are the Editorial Board members of Ornamental Plant Research who were blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of these Editorial Board members and their research groups.
-
# These authors contributed equally: Gan Huang, Boxiao Dong
- Supplemental Fig. S1 15 days root length and 45 days plant height of wild type and transgenic lines.Scale bars = 2cm.
- Supplemental Fig. S2 Venn diagram representing the numbers of upregulated and downregulated genes. the numbers of upregulated(a) and downregulated(b) genes in four comparisons of the WT and transgenic lines of OE-128 under contrast and inoculation.the numbers of upregulated(c) and downregulated(d) genes in four comparisons of the WT and transgenic lines of OE-156 under contrast and inoculation. The numbers in photo indicated the number of genes that were upregulated or downregulated. OE-128 and OE156, different overexpression lines; Aa, inoculated with A. alternata.
- Supplemental Fig. S3 KEGG pathway analysis of DEGs in five comparisons of the WT and overexpression transgenic lines under contrast and inoculation. (a)WT-vs-OE-156. (b)WT-Aa-vs-OE-156- Aa. (c)OE-156-vs-OE-156-Aa. (d)OE-128-vs-OE-128-Aa. (e)WT-Aa-vs-OE-128-Aa.
- Supplemental Material 1 Genetic CDNA sequence and protein sequence of CmNAC083.
- Supplemental Table S1 Statistics of plant height and root length of transgenic lines and wild type.
- Supplemental Table S2 List of primer sequences used for qRT-PCR in this study.
- Supplemental Fig. S3
- Supplemental Table S3 Differential expression genes between CmNAC083 overexpression strains and WT.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Huang G, Dong B, Jiang J, Chen S, Fang W, et al. 2023. CmNAC083 regulates resistance to Alternaria alternata via reactive oxygen species and jasmonic acid signaling pathways in Chrysanthemum morifolium. Ornamental Plant Research 3:16 doi: 10.48130/OPR-2023-0016 

CmNAC083 regulates resistance to Alternaria alternata via reactive oxygen species and jasmonic acid signaling pathways in Chrysanthemum morifolium
- Received: 07 May 2023
- Accepted: 15 September 2023
- Published online: 27 September 2023
Abstract: Chrysanthemum morifolium is an ornamental plant of economic importance. Chrysanthemum black spot is caused by Alternaria fungi. Members of the NAC family regulate plant biological and abiotic stress responses. We screened and confirmed the target gene, CmNAC083, by transcriptome sequencing after inoculation with Alternaria fungi. In the present study, the CmNAC083 gene was cloned from the chrysanthemum 'Jinba' leaves, and its protein structure was analyzed. CmNAC083 exhibits transcriptional inhibitory activity, and a tobacco subcellular localization experiment revealed that CmNAC083 is found in the nucleus. Compared with other tissues, CmNAC083 was highly expressed in the stem and bud during the vegetative growth period and in the stems and roots during flowering. Inoculation with Alternaria alternata induced sustained high expression of CmNAC083, reaching its peak at 48 h after infection with the black spot pathogen. Overexpression of CmNAC083 can enhance the resistance of chrysanthemums to black spot disease, and its transient silencing presents a susceptible phenotype. The results of RNA sequencing results revealed that the transcription levels of jasmonic acid (JA) biosynthesis, response signal pathway, and reactive oxygen species (ROS) pathway genes in the CmNAC083 overexpression lines were upregulated compared with those in the wild-type (WT) plants. Furthermore, the overexpressing strains exhibited a higher endogenous JA content than WT after treatment with A. alternata. Overall, CmNAC083 overexpression positively enhances chrysanthemum resistance to black spot disease by regulating A. alternata infection by upregulating the transcription levels of the JA synthesis and ROS pathways.
-
Key words:
- CmNAC083 /
- Alternaria alternata /
- ROS /
- Chrysanthemum /
- JA /
- Black spot disease