








-
Pancreatic cancer (PC) is among the most lethal malignancies, and its mortality closely parallels its morbidity. PC is estimated to be the second leading cause of cancer deaths in the next decade, with incidence increasing rapidly[1,2]. Surgical pancreatectomy remains the only curative approach for PC. Unfortunately, the majority of patients are unresectable at the time of diagnosis due to extensive local spread and metastasis[3,4]. PC is an atypical disease whose symptoms are easily confused with other noncancerous gastrointestinal symptoms, including nausea, abdominal pain, weight loss, and jaundice, leading to later medical consultations. In addition, the pancreas is an easily overlooked organ (16 cm × 6 cm × 2 cm) hidden behind the stomach and duodenum, and pancreatic cancer holds few blood vessels compared to other gastrointestinal cancers, making PC insensitive to imaging detection, such as computed tomography (CT) and magnetic resonance imaging (MRI)[5]. Large-scale screening programs can effectively reduce the disease burden in various gastrointestinal cancers, including esophageal cancer and colorectal cancer. However, there are no noninvasive diagnostic methods that facilitate early diagnosis of population-based PC[6−9]. This fact sheds light on the necessity for discovering a novel powerful diagnostic approach with higher compliance for PC in the early stages of development.
The human body encompasses trillions of indigenous microbes, including eukaryota, bacteria, and archaea. Microbiota are a collection of microorganisms that live within/on the human body, and the human microbiome refers to the collective genome encoded by the microbiota[10,11]. Body sites such as the skin, oral cavity, stomach, bladder, vaginal cavity, nasal cavity, and lungs host rich microbial communities; among them, the gastrointestinal tract harbors the greatest density of microbiota, which means the gut microbiota receive the most attention. Bacterial analysis is the main focus when mentioning the microbiota unless otherwise specified. Recently, facilitated by the advancement of high-throughput 16S rRNA sequencing and whole-genome shotgun metagenomic technology, we can gain a glimpse into microbial diversity and abundance[12,13]. Accumulating studies have suggested that the human microbiota plays a vital role in human health, both in physiological and pathological conditions. Given that the microbiota is associated with a variety of diseases, its analysis may help in disease diagnosis, prognosis, prevention, and therapy[14−19]. Among several sample sources that can reflect the fluctuation of the human microbiota, such as intestinal mucosa and secretions, stool, and oral saliva are undoubtedly the most noninvasive and accessible means. It has been reported that diagnostic models based on discriminating bacterial species from oral or fecal samples show high sensitivity and specificity in various malignancies, including hepatocellular carcinoma and colorectal cancer[20−22]. Further studies reveal that dysbiosis of the gut microbiota participates in carcinogenesis via abnormal metabolites or immune modulation. There are also articles mentioning the relationship between gut microbiota and PC, which provides a basis for the diagnostic value of gut microbiota in PC. Farrell et al. compared oral microbial composition between PC patients and matched healthy controls via Human Oral Microbe Identification Microarray (HOMIM). They identified 16 species/clusters, which showed a significantly different frequency between the two groups[23]. Similarly, Michaud et al. compared antibodies against oral bacteria between 405 PC patients and 416 matched controls and showed that individuals expressing higher levels of antibodies against Porphyromonas gingivalis (P. gingivalis) have a twofold higher risk of PC than those with lower levels of antibodies[24]. In addition to HOMIM and antibodies, there are various methods to investigate the microbial composition, including the long-lasting gold standard, 16S rRNA sequencing[25]. Based on next-generation sequencing technology, several studies aimed to reveal the entirety of the genetic information contained in a sample from PC patients and looked to discriminate bacteria as a diagnostic biomarker for PC[26−40].
Considering the possibility of human microbiota serving as a noninvasive diagnostic tool in the screening of PC, we sought to systematically review the studies revealing different microbial compositions between PC patients and controls and the possible use of the models established on discriminated taxa for the early diagnosis of PC. Given the significant variations between different detection methods for the microbial community, this systematic review focuses only on articles using 16S rRNA sequencing.
-
The aim of this systematic review was to collect all original research articles that evaluated the possibility of gut microbiota, measured by 16S rRNA sequencing, as a noninvasive prognostic tool for PC. The research was performed in accordance with procedures recommended by the Cochrane Collaboration.
Search strategy
-
A comprehensive literature search for eligible articles was performed in PubMed, Cochrane Library, Embase, and Web of Science up to 20 November, 2024. The search approach followed the combinations of the terms 'pancreatic cancer', 'pancreatic ductal adenocarcinoma', 'pancreatic adenocarcinoma', 'gut', 'fecal', 'stool', 'oral', 'salivary', 'microbial', 'microbiome', and 'microbiota'. Relevant reviews were screened to seek missing studies. Titles and abstracts were screened to remove duplicates. This systematic review was limited to English-language articles only.
Inclusion and exclusion criteria
-
The articles that met all following terms were included in this review: assessed the gut microbial community in oral or fecal samples from PC patients compared with healthy controls, used 16S rRNA sequencing, and found several discriminated bacteria as outcomes. Studies were removed if they were published as letters or case reports, as they failed to provide enough data for the present review. Studies examining fungi or viruses in PC patients were excluded because they did not share the same sequencing method with bacteria. Articles focused on specific bacterial species, such as Helicobacter pylori, were also removed because several meta-analyses related to this topic have been published. The reviewers screened the studies based on the inclusion criteria independently, and disagreements were discussed to resolve the differences.
Data extraction and quality assessment
-
Two reviewers independently retrieved the articles. The basic information extracted from the articles was summarized as: study characteristics, population characteristics, and methodologic characteristics. The study characteristics were as follows: first author, publication year, country, publication type, and study type. The population and methodologic characteristics were displayed in terms of sample size, age, sex, body mass index, smoking status, sample source, temperature for storage, measurement method, diversity assessment, and administration of antibiotics or probiotics. To minimize biases, a quality assessment was conducted following the Newcastle–Ottawa Scale (NOS)[41], which is based on three components: the selection of study groups, comparability, and ascertainment of exposure/outcome. Two independent reviewers scored the NOS on major aspects of risk and applicability assessment.
-
The search process is displayed in a PRISMA flow diagram, as shown in Fig. 1. A total of 80 papers were retrieved from PubMed, the Cochrane Library, Embase, and the Web of Science. Following a thorough screening of their full titles and abstracts, 18 reviews or meta-analyses were excluded. The full-text articles were further examined and those that did not meet all the inclusion criteria were excluded. Fourty-seven papers were excluded due to the following reasons: use of invasive methods to obtain samples (n = 5), and not being human studies (n = 10); lack of controls (3); not pancreatic cancer (26); other detection methods (3). Finally, 15 further studies were ultimately included.
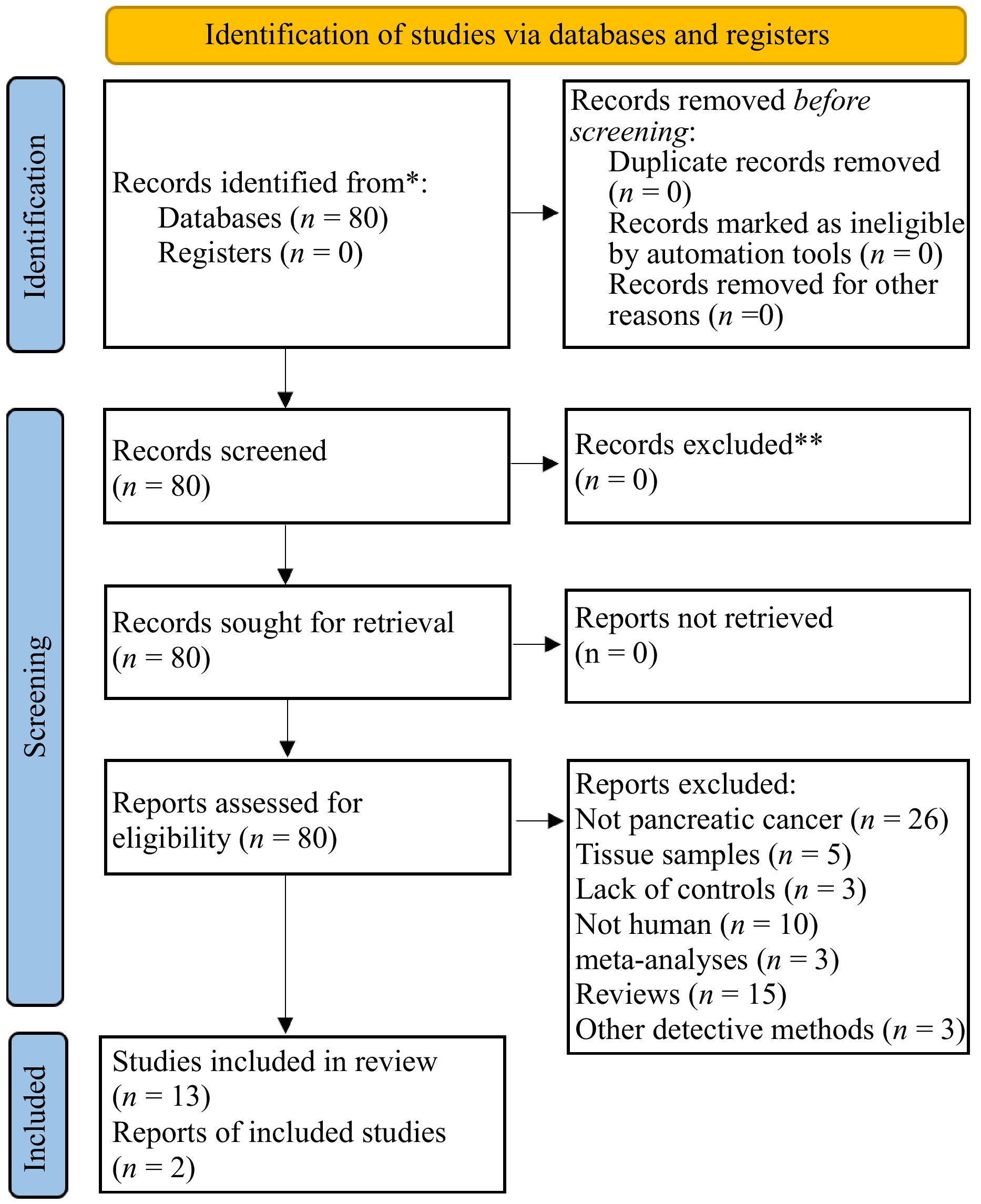
Figure 1.
Flowchart.
Study characteristics
-
Table 1 displays the characteristics of the 15 studies included in our review published between 2013 and 2024. Among the 15 studies, five originated from China, four were from the USA, another two were from Israel and Japan, and one was from Iran and Germany. The majority of studies were published as journal articles, and only two were conference proceedings. Only one of the studies was designed as a prospective study, while the remaining studies were case–control studies. The NOS score ranged from 6 to 8 in the included studies.
Table 1. Study characteristics.
Study Country Publication type Study type NOS score Fan et al., 2018[26] USA Journal article Prospective cohort 8 Torres et al., 2015[31] USA Journal article Controlled 8 Olson et al., 2017[29] USA Journal article Pilot 7 Lin et al., 2013[34] USA Conference proceedings Pilot 7 Lu et al., 2019[28] China Journal article Controlled 8 Vogtmann et al., 2019[32] Iran Journal article Controlled 8 Ren et al., 2017[30] China Journal article Controlled 8 Half et al., 2019[27] Israel Journal article Controlled 8 Half et al., 2015[33] Israel Conference proceedings Pilot 6 Kartal et al., 2022[35] Germany Journal article Case–control 6 Chen et al., 2023[36] China Journal article Controlled 7 Hashimoto et al., 2022[38] Japan Journal article Controlled 8 Sono et al., 2024[39] Japan Journal article Controlled 7 Zhao et al., 2024[37] China Journal article Controlled 8 Yang et al., 2023[40] China Journal article Controlled 6 NOS, Newcastle-Ottawa Scale. USA, United States of America. Population and methodologic characteristics
-
The population characteristics are described in Table 2. The sample size of the included studies ranged between 28 (13 PC) in the smallest study by Lin et al., and 732 (361 PC) in the largest study by Fan et al. To increase the specificity of the diagnostic taxa, four reviews included other pathological conditions, such as pancreatitis and nonalcoholic fatty liver disease, in the control group rather than healthy individuals only. The majority of journal articles were able to provide necessary demographic information such as age, gender, and body mass index. The sample type included an oral wash, saliva, tongue coat, and stool, which were all obtained by noninvasive methods. Half of the studies emphasized that samples were stored at −80 °C before analysis. It is recognized that the administration of antibiotics, probiotics, and prebiotics may affect gut microbiota. All the case–controlled journal articles limited antibiotic usage when enrolled. Smoking status, a well-known risk factor for PC, also plays a role in gut microbiota analysis, as mentioned in only four articles. Although all these articles adopted 16S rRNA sequencing, hypervariable regions vary with different papers including V3–V4, V4–V5, and V3–V5, while several articles did not mention the hypervariable regions they chose for analysis. Six articles assessed both α-diversity and β-diversity in the analysis, the Simpson index and the Shannon index were the most used as an outcome of α-diversity, and β-diversity is mainly represented by the principal coordinate analysis of weighted and unweighted UniFrac distances. The above description is shown in Table 3.
Table 2. Population characteristics.
Study Sample size Age % male BMI % smoking PC Control PC Control PC Control PC Control PC Control Fan et al., 2018[26] 361 371 68.5 68.3 57.1 57.1 − − 57.3 49.9 Torres et al., 2015[31] 8 100
(other disease 78 HC 22)71.1 60.7
(other cancers)75.0 50.0 − − − − Olson et al., 2017[29] 40 97
(IPMN 39; HC 58)< 70, 64.0%; ≥ 70, 35.0% < 70, 42.0%; ≥ 70, 59.0% (IPMN); < 70, 81.0%;
≥ 70, 19.0% (HC)53.0 56.0 (IPMN); 40.0 (HC) Normal 38.0%; abnormal 61.0% Normal 36.0%;
abnormal 64.0% (IPMN); normal 43.0%;
abnormal 57.0% (HC)44.0 46.0 (IPMN); 31.0 (HC) Lin et al., 2013[34] 13 15
(pancreatitis 3 HC 12)− − − − − − − − Lu et al., 2019[28] 30 25 50.8 ± 5.3 48.2 ± 6.0 70.0 80.0 22.5 ± 1.2 22.6 ± 1.6 − − Vogtmann et al., 2019[32] 273 285 < 70, 63.7%;
≥ 70, 36.3%< 70, 67.4%; ≥ 70, 32.6% 60.4 46.0 Normal 57.5%; abnormal 42.5% Normal 46.3%;
abnormal 53.7%30.5 25.6 Ren et al., 2017[30] 85 57 56.0 (33.0–78.0) 52.0 (43.0–67.0) 55.3 63.2 22.7 (19.5–26.0) 23.2 (18.5–27.1) − − Half et al., 2019[27] 30 35
(NAFLD 16; PCL 6 HC 13)68.9 ± 6.2 51.0 ± 10.8 (NAFLD);
66.0 ± 15.3 (PCL);
59.0 ± 8.7 (HC)53.3 75.0 (NAFLD);
83.3 (PCL); 46.2 (HC)− − − − Half et al., 2015[33] 15 15 − − − − − − − − Kartal et al., 2022[35] 57 79 (HC 50 CP 29) − − − − − − − − Chen et al., 2023[36] 40 54 (HC 50 CP 15) − − − − − − − − Hashimoto et al., 2022[38] 5 68 70.0−89.0 54.0 40.0 42.6 − − − − Sono et al., 2024[39] 30 18 63.7 63.0 53.3 66.7 22.0 24.5 36.7 88.9 Zhao et al., 2024[37] 29 9 67.6 ± 10.8 30.5 ± 6.8 58.6 33.3 22.4 ± 2.9 20.8 ± 1.5 − − Yang et al., 2023[40] 44 50 − − 47.7 − 22.4 ± 2.8 − − − /, no related information; BMI, body-mass index; HC, healthy control; IPMN, intraductal papillary mucinous neoplasms; NAFLD, non-alcoholic fatty liver disease; PCL, pre-cancerous lesions. Notes: * It was a prospective study including two large population-based cohorts whose BMI was described in Median or mean. Table 3. Methodologic characteristics.
Study Sample Temperature
for storageMeasurement
methodDiversity assessment Antibiotics Probiotic or
prebioticα-diversity β-diversity Fan et al., 2018[26] Oral wash −80 °C 16S V3–V4 Shannon, Simpson PCoA − − Torres et al., 2015[31] Saliva −80 °C 16S Chao1 ANOSIM Not in 2 wk − Olson et al., 2017[29] Saliva − 16S V4–V5 NP Shannon, Inverse Simpson NA Not in 30 d − Lin et al., 2013[34] Oral wash − 16S − − − − Lu et al., 2019[28] Tongue coat − 16S V3–V4 Shannon, Simpson, inverse Simpson, Obs, Chao 1, ACE PCoA Not in 8 wk Not in 8 wk Vogtmann et al., 2019[32] Saliva −70 °C 16S V4 Observed SVs, Shannon, Faith's PD PCoA − − Ren et al., 2017[30] Stool −80 °C 16S V3–V5 Shannon, Simpson, Chao 1 PCoA Not in 8 wk Not in 8 wk Half et al., 2019[27] Stool −80 °C 16S Shannon PCoA Not in 8 wk − Half et al., 2015[33] Stool − 16S − ANOSIM − − Kartal et al., 2022[35] Stool and saliva −80 °C 16S V4 Shannon, Simpson Unweighted TINA index − − Chen et al., 2023[36] Fecal and saliva −80 °C 16S V3–V4 Chao1, Shannon observed species, and PD whole tree PCoA Not in 4 wk Not in 4 wk Hashimoto et al., 2022[38] Stool and saliva −80 °C 16S Shannon PCoA Not in 6 months Not in 6 months Sono et al., 2024[39] Stool and saliva − 16S V3–V4 Observed features, Shannon BrayeCurtis dissimilarity QIIME 2 − − Zhao et al., 2024[37] Stool −80 °C 16S V3–V4 Chao 1 Acex, Shannon Simpson Sobs i Coverage Mothur software Qiime Not in 8 wk Not in 8 wk Yang et al., 2023[40] Stool −80 °C 16S Chao 1 Shannon PCoA − − −, no related information. d, day; wk, week. Bacterial detection
-
Bacterial taxonomic levels are described in Table 4. Combining studies that investigated samples from the oral cavity, six phyla were significantly different between PC patients and healthy controls, including Fusobacteria, Bacteroidetes, Firmicutes, Actinobacteria, Verrucomicrobia, and Proteobacteria. In addition, the abundance of Firmicutes and Proteobacteria was found to differ significantly in at least two studies, and they shared the same changing directions in different articles. However, there were some contradictory findings in the phyla Bacteroidetes and Fusobacteria. Fan et al. found that the phylum Fusobacteria was associated with decreased PC risk (OR = 0.94), while the phylum Bacteroidetes was associated with a higher risk of PC (OR = 1.01)[26]. Their findings were inconsistent with the results from Lu et al., whose findings suggested that PC patients presented a higher abundance of Fusobacteria and a lower abundance of Bacteroidetes than healthy controls[28]. However, the results from Fan et al. were in agreement with the findings from a European cohort containing 405 PC patients and 416 matched controls[24], in which higher antibody levels from Fusobacteria in prediagnosis blood were associated with reduced PC risk. At the genus level, 19 taxa were found to be significantly different between PC patients and healthy controls in five studies: Leptotrichia, Alloprevotella, Bacteroides, Porphyromonas, Aggregatibacter, Neisseria, Streptococcus, Haemophilus, Corynebacterium, Fusobacterium, Actinomyces, Rothia, Solobacterium, Oribacterium, Campylobacter, Atopobium, Parvimonas, Paraprevotella, and Lachnospiraceae G7. Among these identified genera, Bacteroides was found to be higher in PC, while Porphyromonas, Aggregatibacter, Neisseria, and Haemophilus were lower in PC when compared with controls in at least two articles. All five genera were found to change in the same direction between different studies. Nevertheless, there was an inconsistency between articles, such as the genus Leptotrichia that was found by Torres et al.[31] and by Lu et al.[28] to be more abundant in PC patients, while Fan et al.[26] found the genus Leptotrichia to be associated with decreased PC risk (OR = 0.87). This inconsistency may be explained by different experimental designs and various types of published articles.
Table 4. Discriminating taxa.
Study Bacteria taxonomic level Phylum Class Order Family Genus Fan et al., 2018[26] Bacteroidetes (↑) SR1[C-1] (↑) Fusobacteriales (↓) Leptotrichiaceae (↓) Alloprevotella, Porphyromonas gingivalis, and Aggregatibacter actinomycetemcomitans (↑) Fusobacteria (↓) Fusobacteria (↓) Leptotrichia (↓) Torres et al., 2015[31] Firmicutes (↑) Leptotrichia Bacteroides (↑) Proteobacteria (↓) Porphyromonas Aggregatibacter Neisseria (↓) Olson et al., 2017[29] Firmicutes (↑) Bacilli (↑) Lactobacillales (↑) Streptococcaceae (↑) Streptococcus (↑) Proteobacteria (↓) Gammaproteobacteria; Betaproteobacteria (↓) Pasteurellales Neisseriales (↓) Pasteurellaceae Neisseriaceae (↓) Haemophilus Neisseria (↓) Lin et al., 2013[34] Bacteroides (↑) Corynebacterium Aggregatibacter (↓) Lu et al., 2019[28] Firmicutes, Fusobacteria and Actinobacteria (↑) Leptotrichiaceae, Fusobacteriaceae, Actinomycetaceae, Lachnospiraceae, Micrococcaceae, Erysipelotrichaceae, and Campylobacteraceae (↑) Leptotrichia, Fusobacterium, Actinomyces, Rothia, Solobacterium, Oribacterium, Campylobacter, Atopobium, and Parvimonas (↑) Bacteroidetes (↓) Prevotellaceae, Pasteurellaceae, and Porphyromonadaceae (↓) Porphyromonas, Haemophilus, and Paraprevotella (↓) Vogtmann et al., 2019[32] Enterobacteriales (↑) Enterobacteriaceae, Bacteroidaceae, Staphylococcaceae (↑) Lachnospiraceae G7 (↑) Haemophilus (↓) Ren et al., 2017[30] Bacteroidetes (↑) Prevotella, Veillonella, Klebsiella, Selenomonas, Hallella, Enterobacter, and Cronobacter (↑) Firmicutes and Proteobacteria (↓) Gemmiger, Bifidobacterium, Coprococcus, Clostridium IV, Blautia, Flavonifractor, Anaerostipes, Butyricicoccus, and Dorea (↓) Half et al., 2019[27] Bacteroidetes (↑) Bacteroidia; Verrucomicrobiae; Clostridia Bacteroidales; Verrucomicrobiales; Clostridiales Porphyromonadaceae; Verrucomicrobiaceae; Clostridiaceae1 Odoribacter, Akkermansia (↑) Firmicutes (↓) Clostridiumsensustricto1 (↓) Half et al., 2015[33] Bacteroidetes Verrucomicrobia (↑) Sutterella, Veillonella, Bacteroides, Odoribacter, and Akkermansia (↑) Firmicutes and Actinobacteria (↓) Kartal et al., 2022[35] Veillonella atypica, Fusobacterium, nucleatum/hwasookii, Alloscardovia, omnicolens (↑) Romboutsia timonensis, Faecalibacterium, rausnitzii, Bacteroides, coprocola, Bifidobacterium, and bifidum (↓) Chen et al., 2023[36] Bacteroidetes (↑) Veillonella, Peptostreptococcus, Akkermansia, Parvimonas, Solobacterium, Olsenella, and Escherichia-Shigella (↑) Firmicutes (↑) Proteobacteria (↑) Verrucomicrobia (↑) Hashimoto et al., 2022[38] Actinomyces, Lactobacillus, Streptococcus, and Veillonella (↑) Anaerostipes (↓) Sono et al., 2024[39] Firmicutes (↑) Streptococcus (↑) Proteobacteria (↓) Neisseria (↓) Zhao et al., 2024[37] Moraella, Sphingomonas Oxalobacteriae Yang et al., 2023[40] Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria (↑) Streptococcus, Lactobacillus, and Bifidobacterium (↑) Based on nine studies focusing on fecal samples, six phyla differed between PC patients and controls. It seems that these nine articles came to some consensus at the phylum level, which all found that Bacteroidetes was significantly more abundant in PC patients than in healthy controls, while Firmicutes was less abundant in PC patients. Two of these articles were conducted by Half et al.[27], whose results were somewhat consistent in some genera, such as Odoribacter and Akkermansia. The comparison is mainly performed between articles from Ren et al.[30] and Half et al.[27] Unlike at the phylum level, there were extensive differences that distinguished PC patients from controls at the genus level between the papers from China and Israel, with almost no overlap. Ren et al. reported that PC patients had significantly higher levels of Prevotella, Veillonella, Klebsiella, Selenomonas, Hallella, Enterobacter, and Cronobacter, as well as lower levels of Gemmiger, Bifidobacterium, Coprococcus, Clostridium IV, Blautia, Flavonifractor, Anaerostipes, Butyricicoccus, and Dorea. The inconsistency is probably driven by a difference in methodology as well as population features, including host genetics and lifestyle.
Multibacterial models for the diagnosis of PC
-
Five articles established diagnostic models based on the discriminated bacteria derived from their analysis[27,28,30,36,40]. The area values under the receiver operator characteristic curve (AUC) in these five studies were very similar to each other, ranging from 0.802 to 0.927. The details are displayed in Table 5. Based on a sample from the tongue coat, Lu et al. developed a model based on the combination of Fusobacterium, Leptotrichia, and Porphyromonas, with high sensitivity (0.771) and specificity (0.786) to discriminate patients with PC from controls[28]. Ren et al.[30] and Half et al.[27] focused on fecal analysis and established a diagnostic model with values of 0.859 and 0.769 for sensitivity and 0.667 and 0.8 for specificity, respectively. Based on fecal flora, Chen et al.[36] and Yang et al.[40] developed diagnostic models with values of 0.916 and 0.927, respectively, but did not provide sensitivity and specificity.
Table 5. Diagnostic models.
Study Models Sensitivity Specificity AUC Lu et al., 2019[28] Fusobacterium, Leptotrichia, and Porphyromonas 0.771 0.786 0.802 Ren et al., 2017[30] Based on the 40 genera 0.859 0.667 0.842 Half et al., 2019[27] Based on discriminating taxa 0.769 0.8 0.825 Chen et al., 2023[36] Random forest − − 0.916 Yang et al., 2023[40] Random forest − − 0.927 AUC, area under the receiver operator characteristic curve. -
Fifteen studies examining the difference in the microbial community between PC patients and controls were included in the present systematic review. Although very limited comparisons were allowed due to the great heterogeneity between these studies, there existed an optimistic conclusion that microbial analysis holds the potential to provide a novel diagnostic tool for the early detection of PC.
Sources heterogeneity
-
It has been reported that the majority of humans may share a 'core microbiota'[42], which has evolved with humans for thousands of years and participates in physiological activities such as nutrient absorption and immune regulation. It is also undoubted that human microbiota may be affected by various external factors, including geographic location, ethnicity, lifestyle, and medications[11,12]. Differences in host location have been reported to be the strongest phenotypic determinant with microbiota variations in humans with similar ancestry[43]. Therefore, the geographical differences of the included articles contribute to inevitable heterogeneity in this systematic review. In addition, there are mutual regulations between microbiota and smoking status or diabetes[14,44], which are also recognized as risk factors for PC. There were limited included articles that described the conditions of smoking or diabetes in detail. Finally, the administration of antibiotics, probiotics, and prebiotics has a crucial impact on the microbiota. Although five journal articles excluded individuals who had taken antibiotics within an indicated period, the majority of the articles failed to provide information about probiotic or prebiotic issues. The sample size, a highly relevant factor in statistical strength, varies from 28 to 732 people in the papers included in this report. Compared with small sample sizes, studies with large sample sizes potentially represent better credibility in the results. Among the nine studies, there was only one prospective evaluation based on two large cohorts. The variations in study design and sample size also led to heterogeneity between articles.
The present research focused on studies that investigated the microbial community based on 16S rRNA sequencing, whose protocols are divergent between articles. The variations in sample handling, variable regions, sequencing depth, and sequencing platform resulted in significant methodological heterogeneity in the present review.
Porphyromonas gingivalis and Leptotrichia
-
In addition to 16S rRNA sequencing, detection methods for microbes include plasma antibody analysis, enzyme-linked immunosorbent assay, quantitative polymerase chain reaction, and microbe identification microarray[45−47]. Combining the results of different techniques and populations, there are two promising microbes involved in PC development, P. gingivalis and Leptotrichia. Periodontal disease is an acknowledged risk factor for PC[48]. Furthermore, P. gingivalis is a major pathogen responsible for periodontal disease[49], and it was found to be increased in PC by multiple articles. Torres et al.[31] and Fan et al.[26] found that P. gingivalis differed significantly between PC and controls. Moreover, Michaud et al. found that a higher level of antibodies against P. gingivalis was associated with an increased risk of PC. Further studies have revealed that P. gingivalis promotes PC carcinogenesis by producing PAD enzymes, which induce mutations in P53 or KRAS or initiate Toll-like receptor (TLR) signaling pathways, which are critical in PC development[47,50]. Fan et al.[26], Torres et al.[31], and Lu et al.[28] found that Leptotrichia differed between PC and controls despite inconsistent directions. Leptotrichia, known as an opportunistic pathogen involved in periodontal disease, is associated with a decreased risk of PC by Fan et al.[26], and Torres et al.[31] are regarded as a protective bacterium for PC[51,52]. In addition, Torres et al. suggested a relatively high ratio of Leptotrichia to Porphyromonas in PC patients. However, further investigation is needed to reveal their role in pancreatic cancer development[31].
P. gingivalis and Leptotrichia in relation to PC
-
P. gingivalis and Leptotrichia, along with their metabolic products, contribute to the development of pancreatic cancer through multiple mechanisms. Here are the possible mechanisms: P. gingivalis can induce inflammation by releasing lipopolysaccharides (LPS) and other virulence factors, thereby triggering an immune response. This includes the activation of Toll-like receptors (TLRs) on pancreatic cells and immune cells, leading to the secretion of pro-inflammatory cytokines (such as IL-1β, IL-6, TNF-α)[53]. P. gingivalis and Leptotrichia produce short-chain fatty acids (SCFAs) and other metabolites during carbohydrate fermentation. While some SCFAs can be protective, elevated levels may contribute to the activation of pro-inflammatory pathways[54]. Enzymes produced by P. gingivalis (such as gingipains) and Leptotrichia can degrade extracellular matrix components and epithelial cells. Damage inflicted on pancreatic tissues can stimulate further inflammation and repair processes[55]. Both bacteria can modulate the adaptive immune response, leading to reduced efficacy in tumor cell recognition. For example, P. gingivalis can induce the polarization of macrophages toward an M2 phenotype, which is immunosuppressive, and stimulate regulatory T cells (Tregs)[56].
Bacterial models for diagnosis
-
A single bacterium fails to show powerful diagnostic ability in early PC detection, and obvious diagnostic models based on multiple bacteria have better performance. As reported in the study by Zheng et al.[20], the multibacterial model based on 30 optimal microbial markers identified from East China can also discriminate hepatocellular carcinoma independently in Northwest China and Central China. The AUC was 80.64% (95% CI 74.47% to 86.80%) for early HCC diagnosis in the discovery phase, which was relatively stable in two validation phases, with values of 76.80% (95% CI 67.90% to 85.70%) and 79.20% (95% CI 67.40% to 90.90%) between early HCC and controls[20]. Yu et al. established a diagnostic model from fecal metagenomes, which can distinguish colorectal cancer from controls with an AUC of 84% in China. Moreover, the AUC values are 72% and 77% when validated in France and Australia, respectively[57]. These studies suggest the possibility of universal bacterial models in discerning gastrointestinal cancers from healthy controls even with microbial variations from geographical and population differences. The analysis by Torres et al. of salivary microbial profiles supports prior work suggesting that salivary microbial communities of patients diagnosed with pancreatic cancer are distinguishable from salivary microbial communities of healthy patients or patients with other diseases, including non-pancreatic cancers[31]. At the phylum level, pancreatic cancer patients tended to have higher proportions of Firmicutes and lower proportions of Proteobacteria. The most striking difference between the microbial profiles of pancreatic cancer patients and other patient groups was in the ratio of the bacterial genera Leptotrichia and Porphyromonas. The LP ratio had been identified as a potential biomarker from a preliminary analysis and an analysis of the full dataset found significantly higher LP ratio in pancreatic cancer patient saliva than in other patients. Based on a sample from the tongue coat, Lu et al.[26] developed a model based on the combination of Fusobacterium, Leptotrichia, and Porphyromonas, with high sensitivity (0.771) and specificity (0.786) to discriminate patients with PC from controls. Zhao's study indicated that pancreatic cancer can significantly increase the species richness and diversity of gut microbes in patients[37]. The dominant species of gut microbes in pancreatic cancer and healthy people are Bacteroides and Lachnospira. The study by Yang et al. demonstrated significant differences in intestinal microbiome composition between PC patients and healthy people, and found that the intestinal microbial richness of PC patients was higher, and the Streptococcus content was significantly increased[40]. Through LEfSe, RF analysis and verified by ROC curve, it was found that it had important discrimination ability in the PC group.
At the same time, none of them performed independent validation in different geographic regions, which limits the reliability of the multibacterial model. Nevertheless, these studies indicate that a noninvasive strategy for the early diagnosis of PC may be achieved by microbiota-targeted biomarkers, which need further investigation in a larger population and cross-regional validation.
Mechanisms by which microbiota are associated with PC
-
The association between the microbiota and PC is widely recognized. However, whether it is a causal relationship and the mechanisms contributing to these observations remain elusive. Emerging studies have moved into the field of elucidating mechanisms by which the microbiota influences the initiation and progression of PC.
Intratumoral microbiota
-
According to the present studies, the microbiota may impact PC development through local and/or remote means. The pancreas, which used to be considered a sterile organ, harbors its own microbial environment, at least under pathological conditions[58]. Considering that the pancreas gland is directly connected to the gastrointestinal tract via a pancreatic duct in the duodenum, it appears possible that some microorganisms directly translocate to the pancreatic parenchyma through the pancreatic duct. The relatively higher prevalence of PCH (pancreatic head carcinoma) than PCB (pancreatic body and tail carcinoma) may provide feasible evidence for this hypothesis due to the closer anatomical location of the pancreatic head to the duodenum[59,60]. In addition, oral gavage of Bifidobacterium pseudolongum leads to the presence of the bacterium in the pancreas in a mouse model of PC[61]. Another alternative hypothesis is explaining how microbiota gain access to the pancreas; it is based on mesenteric lymph and/or mesenteric venous drainage[62]. The gut microbiota is an important component of the gut barrier, and disturbance of the microbiota may induce dysfunction of the gut barrier. The microbes may migrate into pancreatic parenchyma as a consequence of intestinal barrier damage and lymphatic dysfunction, which is caused by dysbiosis. The pancreatic microbiota, especially the intratumoral microbiota, may affect the prognosis and therapeutic efficacy of pancreatic cancer. Riquelme et al. compared tissue microbiota between long-term survival and short-term survival PC patients by 16S rRNA sequencing and found a significant difference in diversity and dominant taxa between them. Patients with a higher abundance of Pseudoxanthomonas-, Streptomyces-, Saccharopolyspora-, Bacillus clausii in tumor tissues presented with long survival, suggesting the importance of pancreatic microbiota in prognosis prediction[63]. Geller et al. found the presence of γ-proteobacteria in pancreatic tumors, probably induced via gemcitabine resistance, owing to gemcitabine degradation by γ-proteobacteria products, such as cytidine deaminase[64]. In addition, Pushalkar et al. also suggested that the intratumoral microbiota influences the response to immunotherapy[61]. These findings indicate the potential of microbial modulation as adjuvant therapy in PC treatment.
Chronic inflammation
-
It is well-established that chronic inflammation is paramount for PC initiation and progression[65]. Chronic pancreatitis, a typical condition of pancreatic inflammation, as well as diabetes and obesity, which may cause systematic inflammation, are risk factors for PC. The microbiota plays a critical role in the development of chronic pancreatitis[66], diabetes, and obesity, by which microbes indirectly impact PC tumorigenesis. Furthermore, an increased abundance of LPS-producing bacteria has been observed in a variety of gastrointestinal malignancies, including pancreatic cancer. Ren et al. found that LPS-generating bacteria, such as Prevotella, Hallella, and Enterobacter, were enriched in fecal samples from PC patients[30]. Lipopolysaccharide (LPS), a major microbe-specific molecular compound localized in the outer membrane of gram-negative bacteria may be recognized by Toll-like receptors (TLRs), which are expressed in immune cells. The combination of LPS and TLR activates multiple downstream proinflammatory pathways, including the NF-κB/MAPK signaling pathway, which plays an important role in inflammation[67−69]. The microbiota exerts a carcinogenic impact by maintaining an inflammatory environment.
Immunity
-
The immunosuppressive microenvironment is an essential feature of pancreatic cancer. Recently, immune therapy of PC has attracted much attention despite the limited response thus far[61]. The microbes have a dual impact on malignancies. On the one hand, bacterial ablation improves the efficacy of checkpoint-targeted immunotherapy, which suggests the immune-suppressive effect of microbiota; on the other hand, antibiotic administration limits the effectiveness of PD-1 therapy, which indicates that microbes may provoke cancer immunity[70−73]. Although an intact gut microbiota is essential for the maturation of the immune system, the absence of microbiota may lead to hypoplastic lymphoid organs and immune cells. Some microbes are responsible for innate and adaptive immune suppression in tumorigenesis by the interaction between LPS and TLRs. The binding and activation of TLRs, including TLR7, TLR9, and TLR5[61,74,75] , via bacterial products maintain an immune-suppressive microenvironment by T cells and another lymphocyte inactivation as a consequence of the macrophage M2-like phenotype. Riquelme et al. found that the gut microbiome can affect immune cell infiltration in pancreatic tumors, and microbiota from long-term surviving patients may induce a strong antitumor immune response[63].
Metabolism
-
Diabetes and obesity, as the systemic inflammatory status mentioned above, are also the two most common metabolic diseases worldwide. Considering the essential role of gut microbiota in the digestion and metabolism of nutrients, the gut microbiota is regarded as an intermediate key linking metabolic disorders and pancreatic cancer. Ren et al. found that the abundance of butyrate-producing bacteria, including Coprococcus, Clostridium IV, Blautia, and Flavonifractor, decreased in the stool of PC patients, suggesting that gut microbiota may affect PC development through metabolites[30]. Butyrate is a short-chain fatty acid (SCFA) derived from the fermentation of fiber by gut microbiota and has shown an antitumor effect in various malignancies. Butyrate can help maintain the integrity of the gut barrier by providing an energy source for intestinal epithelial cells and inhibiting microbe translocation by gut leakage[76,77]. In addition, butyrate plays an essential role in multiple cancerous activities by regulating epigenetic processes, including proliferation, differentiation, apoptosis, and invasion.
Some specific microorganisms in PC
-
It is estimated that approximately 10-20% of malignancies are attributed to infectious factors. Several specific microbes have been identified as carcinogenic pathogens, such as Helicobacter pylori in gastric cancer and HPV in cervical cancer. However, no specific pathogen has been found to be the causative agent for PC[78,79]. Accumulating epidemiological studies suggest a close relationship between H. pylori, hepatitis virus, and PC despite still controversy between studies[80−83]. In addition, some articles highlight the importance of fungi in the carcinogenesis of PC. Aykut et al. found that pancreatic cancer tissue harbors 3,000-fold increased fungi compared with healthy tissue, especially Malassezia spp. Administration of amphotericin B to eliminate fungi may delay the progression and invasion of PC in mouse models[84]. Further research suggested that glycans on the fungal wall may bind and activate mannose-binding lectin (MBL) and then drive the complement system cascade, which is necessary for oncogenic progression.
From IPMN to PC
-
The development of pancreatic cancer is a long-term process that develops through several histopathological facets, including the following: chronic inflammation, pancreatic intraepithelial neoplasia, and finally pancreatic cancer[85]. The microbiota may play a key role in the entire process from chronic pancreatitis to precancerous conditions, even to pancreatic cancer. A qualified diagnostic model can distinguish PC patients from confusing diseases such as other gastrointestinal malignancies rather than healthy individuals only. Four studies included in this review have considered this and recruited pancreatitis, NAFLD, and IPMN patients as controls[27,29,31]. Although independent articles explain the relationship between chronic pancreatitis or IPMN and microbiota, a comparative model from a homogeneous region and population is still needed to establish a more qualified diagnostic model for pancreatic cancer with higher specificity.
The present study had several limitations. First some of the sample sizes of the articles included were small, with only 13 individuals in some disease groups, which is mainly due to the low rate of pancreatic cancer incidence. Secondly, not all medical records had randomized controls, furthermore, the specimen sites studied in the individual articles differed, which may have led to a bias in the measured flora.
-
This systematic-based review on limited existing human studies, supporting the possibility of microbiota analysis, searched for a useful noninvasive diagnostic tool in PC detection. The variations in study design and population, methodological characteristics between studies, as well as the influence of region, drugs, and diet on the microbiota led to heterogeneity in this systematic review. Therefore, future studies with larger sample sizes and controlled covariates are needed for a better understanding of the relationship between microbiota and PC, as well as several different diseases.
-
Not applicable.
-
The authors confirm contribution to the paper as follows: draft manuscript preparation: Zhang Y, Du H; manuscript reviewed and editing: Chen H. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yanyan Zhang, Huang Du
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang Y, Du H, Chen H. 2024. 16S rRNA-based analysis of microbiota as a promising noninvasive biomarker for pancreatic cancer diagnostic tool: a systematic review. Gastrointestinal Tumors 11: e004 doi: 10.48130/git-0024-0003 

16S rRNA-based analysis of microbiota as a promising noninvasive biomarker for pancreatic cancer diagnostic tool: a systematic review
- Received: 03 September 2024
- Revised: 26 November 2024
- Accepted: 12 December 2024
- Published online: 27 December 2024
Abstract: Pancreatic cancer (PC) is a severe disease with no effective early detection methods. New evidence links microbiota to PC, hinting at its diagnostic potential. Despite no current clinical microbiota biomarkers for PC, a systematic review was conducted to assess microbiota's role in noninvasive PC detection. A systematic review analyzing 16S rRNA sequencing data on pancreatic cancer-related microbiota was conducted. The search on PubMed, Cochrane Library, Embase, and Web of Science conducted on 20 November, 2024, aimed to find studies linking PC with microbiota in noninvasive samples. The analysis centered on the differences in microbial taxa between PC patients and healthy controls. Fifteen studies were included, with 10 focusing on oral, and nine on fecal microbiota. There was significant variation in six bacterial phyla (Fusobacteria, Bacteroidetes, Firmicutes, Actinobacteria, Verrucomicrobia, and Proteobacteria ) between pancreatic cancer patients and controls. While genus-level findings varied, some genera like Leptotrichia showed diagnostic potential. Three diagnostic models using these taxa showed good sensitivity and specificity for pancreatic cancer. The studies suggest a link between microbiota and pancreatic cancer, though no single bacterium stands out as most relevant. Diagnostic models using multiple taxa show promise for early detection of PC. More extensive, well-designed studies are needed to establish microbiota's role in pancreatic cancer screening.
-
Key words:
- Pancreatic /
- Cancer /
- Microbiota /
- Diagnostic /
- Reviews