






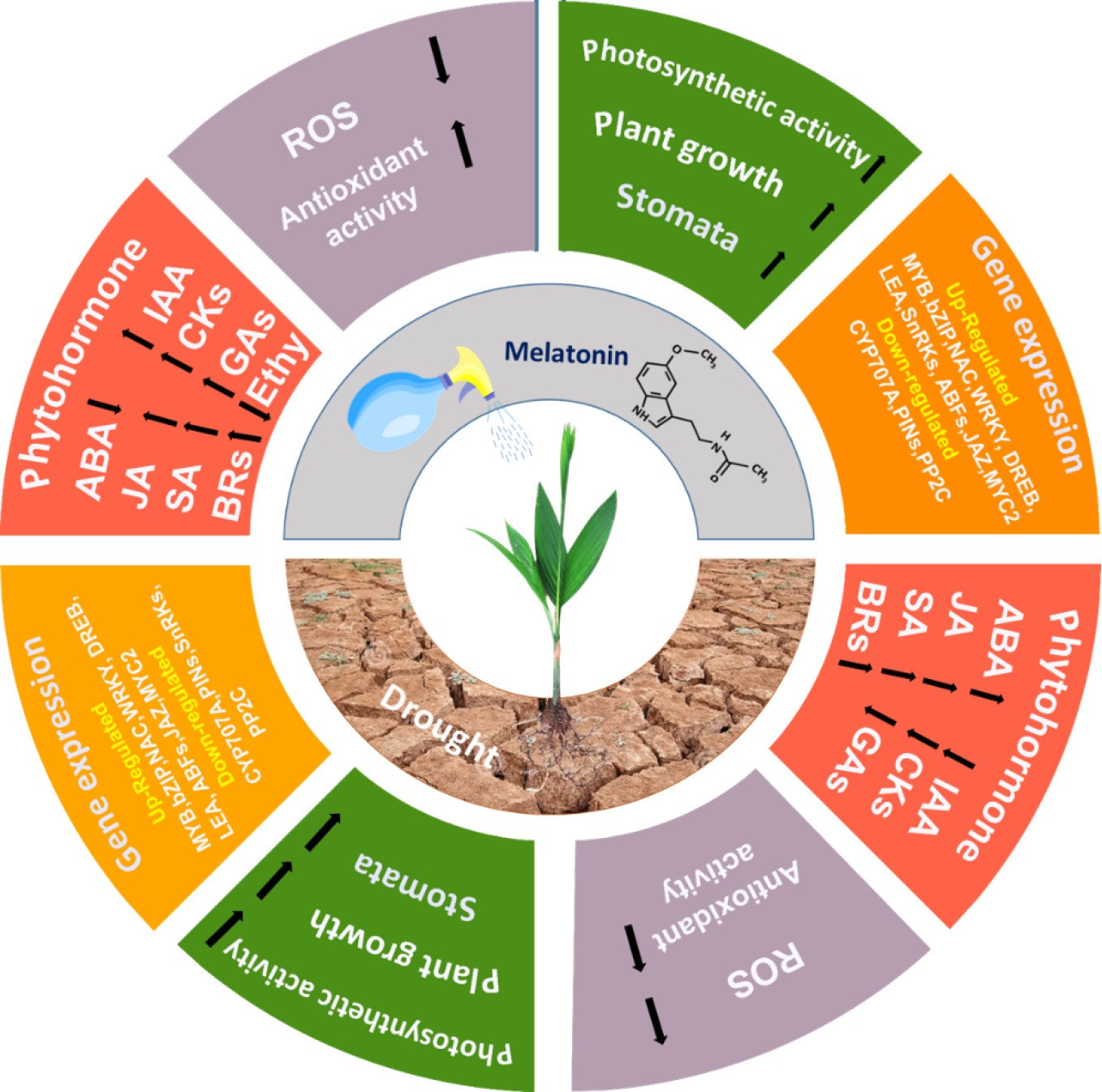



-
Drought stress (DS) significantly impairs plant growth and development by reducing photosynthesis, disrupting metabolism, and causing oxidative damage[1]. Approximately one-third of the world's land area is covered by deserts or semi-deserts, while other regions experience various degrees of drought throughout the plant-growing season[2]. Therefore, enhancing agricultural productivity in drought conditions is essential to meeting the growing global food demand.
Plants employ several physiological and biochemical defense mechanisms to counter drought stress. Key physiological adaptations include stomatal regulation, transcription factor activation, and abscisic acid (ABA) signaling to minimize water loss via transpiration[3,4]. Plants also activate antioxidant enzyme systems during stress to mitigate the detrimental effects of reactive oxygen species (ROS) and maintain metabolic equilibrium. Furthermore, plants produce secondary metabolites, such as soluble proteins, soluble sugars, flavonoids, and proline, which play crucial roles in osmoregulation and the modulation of reactive oxygen species (ROS) levels[1,2].
Exogenous applications of phytohormones, including melatonin (MT), regulate plant responses to DS by modulating physiological and biochemical processes[5]. MT, a substituted indoleamine derived from tryptophan (N-acetyl-5-methoxytryptamine), is naturally produced by nearly all organisms and plays vital roles in photosynthesis, flowering, root growth, and seed germination[6] Exogenous melatonin (Ex-MT) has been shown to enhance plant resistance to various abiotic stressors, such as drought, salinity, and heavy metal toxicity[6,7]. Research indicates that Ex-MT improves DS tolerance by regulating photosynthesis, ion homeostasis, antioxidant activity, and hormone interactions[8,9]. To mitigate stress damage, MT also influences critical metabolic pathways, including the MAPK pathway, flavonoid biosynthesis, glucose metabolism, and the ascorbate-glutathione cycle[10]. However, further research is required to fully elucidate melatonin's molecular role in improving drought tolerance.
Areca catechu L., a member of the Arecaceae family, possesses considerable economic and medicinal value. The plant is cultivated extensively in China, Thailand, Cambodia, Malaysia, Indonesia, India, and Pakistan[11]. It has also emerged as an important cash crop in Africa and Southeast Asia due to the presence of arecoline, a highly addictive alkaloid ranked fourth globally after alcohol, caffeine, and nicotine[11].
Drought stress remains a critical constraint on crop productivity, particularly in arid and semi-arid regions. A.catechu L., a widely cultivated economic crop, is highly sensitive to water deficits, leading to significant yield losses. While melatonin has emerged as a critical phytohormone in stress resistance, its specific molecular mechanisms in drought adaptation, particularly in A. catechu L., remain poorly understood. Previous studies on A. catechu have primarily focused on the physiological effects of melatonin (MT), such as its role in enhancing antioxidant capacity and photosynthesis[12−14]. However, the molecular and transcriptomic mechanisms underlying MT-induced drought resistance remain largely unexplored. This study investigates the physiological, biochemical, and molecular responses of A. catechu to melatonin treatment under drought stress, providing new insights into its application for improving crop resilience. This research examines key parameters, including enzyme activities, metabolite concentrations, photosynthetic performance, and gene expression profiles in plants subjected to drought stress with and without melatonin supplementation.
-
This research used two-leaf Areca catechu L. 'Reyan No. 1' seedlings supplied by the Coconut Research Institute of the Chinese Academy of Tropical Agricultural Sciences in Wenchang, Hainan province, China. The seedlings were grown in 12 cm × 12 cm plastic pots containing sterilized perlite. After being rinsed for two weeks with half-strength Hoagland's solution (pH = 6) for adaptive cultivation, the seedlings were treated with 200 μmol/L melatonin[15]. According to our preliminary experiments and prior research[16], the concentration of melatonin used could be the optimal level for reducing the effects of DS on the observed phenotype. The experiment was performed in Completely Randomized Design (CRD), with three biological replicates for each treatment. These replicates were utilized for phenotypic analysis, RNA sequencing, and subsequent molecular analyses to validate the RNA-seq findings. The experimental treatments were categorized into four groups: (1) CK: seedlings irrigated with regular water, serving as the control group; (2) D: seedlings exposed to drought stress (45% field capacity); (3) M: seedlings treated with melatonin; and (4) MD: seedlings subjected to drought stress following foliar spraying and irrigation with melatonin. Following experimental treatments, leaf samples were collected and immediately stored at −80 °C for further analyses.
Transcriptomic profiling using next-generation sequencing
de novo assembly
-
Total RNA was isolated from plant samples using an RNAprep Pure polysaccharide polyphenol plant total RNA extraction kit (DP441; TianGen; China) adhering to the manufacturer's guidelines. RNA concentration and purity were assessed by spectrophotometer (NanoDrop 2000). For RNA sequencing library construction, 1 μg of total RNA was used for each sample. The NEBNext UltraTM RNA Library Prep Kit for Illumina (NEB, USA) was used to create sequencing libraries, integrating distinct index codes for sample identification. Cluster generation was executed utilizing a cBot Cluster Creation System with the TruSeq PE Cluster Kit v4-cBot-HS (Illumina), per the company's guidelines. Subsequently, the prepared libraries were sequenced on an Illumina platform, generating paired-end reads. Raw data were subjected to quality control, removing reads comprising poly-N sequences, adapters, or low-quality bases. Retained clean reads were subjected to additional analysis, and their GC content, Q30, and Q20 sequence duplication levels were computed. Hisat2 software matched high-quality, clean reads to the reference genome sequence. We selected only readings exhibiting perfect matches or a single mismatch for further analysis and annotation following the reference genome.
Functional enrichment analysis of differentially expressed genes (DEGs)
-
DESeq2 software was used for differential expression analysis among two groups/conditions. p-values were modified using the Benjamini-Hochberg technique to regulate the false discovery rate. Genes exhibiting an estimated p-value < 0.01 were classified as differentially expressed. Furthermore, edgeR software was used to examine differences in differential expression between the two individual samples, using a strict threshold of Fold Change > 2 and false discovery rate FDR < 0.01 for significant differences in expression. To account for any gene length bias, the Wallenius non-central hyper-geometric distribution was included in the GO enrichment analysis of DEGs using the GOseq R tools[17]. Additionally, the statistical enrichment of DEGs in the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways was evaluated using the KOBAS tool[18]. Protein family and gene functions were determined using NCBI, Swiss-Prot, KO, and GO tools. Gene expression levels were quantified using FPKM values (fragments per kilobase of transcript per million mapped fragments).
Identification of co-expression modules and visualization of sub-networks associated with treatments
-
The weighted gene co-expression network analysis (WGCNA) was executed through the BMK Cloud online tool (
www.biocloud.net ). Cytoscape_v3.10.2 was used to view the top connected hub genes network among sub-modules with the highest centrality values within the network of DEGs.Quantification of melatonin content
-
Freeze-dried and ground plant extracts (1 g) were suspended in 10 mL of 50% methanol and subjected to ultrasonication for 20 min at room temperature. The homogenates underwent centrifugation for 30 min at 10,000 × g at 4 °C. Supernatants were moved to fresh vials filtered via a 0.22 μm filter, and methanol was vaporized through nitrogen gas. Concentrated samples were solubilized in 1 mL of methanol, filtered with a 0.22 μm filter paper, and preserved at 4 °C. A standard stock solution of melatonin (10 mg/mL) was prepared in the mobile phase and stored at −20 °C. The stock solution was diluted with mobile phase to create working solutions (0.01, 0.03, 0.05, 0.1, 1, 3, 5, and 10 μg/mL). Melatonin content was determined by a high-performance liquid chromatography (HPLC) system (e2695, Waters, USA) with a diode array detector (2998, Waters, USA), as described previously[19−21]. A 5 μL sample was injected into a C18 column (150 × 2.1 mm, 3.5 μm) (GL Science Inc., Japan). In a 70:30 volume ratio, mobile phases A and B contained 0.45% formic acid in water and acetonitrile. The flow rate was set to 0.5 mL/min and detected at 220 nm. Endogenous melatonin levels were calculated using a standard curve, recorded chromatograms, and peak areas were used to determine melatonin concentrations. Every experiment was done in triplicate, and the mean ± standard deviation was used to show the data.
Determination of physiological indexes
Quantification of proline content
-
The physiological indexes were determined following the manufacturer's recommendations using the Solarbio kit (Beijing, China). The proline content was determined using the sulfosalicylic acid/ninhydrin reaction method[22]. The frozen leaf sample (0.3 g) was suspended in 5 mL of 3% sulfosalicylic acid. After homogenization, the mixture was heated to 100 °C for 5 min and centrifuged at 4,500 × g for 10 min. A boiling water bath was used to incubate the mixture of 0.5 mL of supernatant, 2 mL of glacial acetic acid, 2 mL of acidified ninhydrin reagent, and 1.5 mL of water for 30 min. Five milliliters of toluene were added to the mixture once it had cooled to room temperature. Colored toluene fraction absorbance was measured at 520 nm using a microplate reader (TECAN, Switzerland). Proline (Sigma Aldrich, Germany) was our reference.
Determination of MDA and H2O2 contents
-
The malondialdehyde (MDA) concentration was quantified using the thiobarbituric acid (TBA) method, with some adjustments[23]. A frozen leaf (0.5 g) was mixed in 5 mL of 5% trichloroacetic acid (TCA), and the mixture was centrifuged at 10,000 × g for 15 min at 4 °C. One milliliter of supernatant was combined with 4 mL of 0.5% (w/v) thiobarbituric acid in 20% trichloroacetic acid. The reaction mixture was cooled to 4 °C after being incubated for 25 min at 90 °C. The sample was subsequently centrifuged at 4 °C for 15 min at 10,000 × g. The absorbance was measured at 532 and 600 nm, and MDA concentration was determined in μmol/g FW. The H2O2 concentrations were quantified using the potassium iodide (KI) assay method as defined by Velikova et al.[24]. Frozen leaf samples (0.5 g) were homogenized in 0.1% TCA and centrifuged for 15 min at 12,000 × g. To 0.5 mL of supernatant, 1 mL of 1 mM potassium iodide and 0.5 mL of 10 mM potassium phosphate buffer (pH 7.0) was added. The mixtures were measured at 390 nm for absorbance and the concentration in micromoles per gram of fresh weight (μmol/g FW) was calculated using an extinction coefficient (ε) of 0.28 mM/cm.
Measurement of antioxidant enzyme activities
-
The activities of several antioxidant enzymes were quantified using Solarbio assay kits following the company's protocol. The activity of superoxide dismutase (SOD; EC 1.15.1.1) was determined following the manufacturer's protocol for the SOD assay kit (Cat. No. BC0175) by measuring the absorbance at 560 nm. Peroxidase (POD; EC 1.11.1.7) activity was similarly determined using the POD assay kit (Cat. No. BC0095) with absorbance measured at 470 nm. Catalase (CAT; EC 1.11.1.6) activity was quantified using the CAT assay kit (Cat. No. BC0205), measuring absorbance at 240 nm. Finally, ascorbate peroxidase (APX; EC 1.11.1.11) activity was determined using the APX assay kit (Cat. No. BC0225) with absorbance measured at 265 nm[25].
Analysis of morphological indexes and biomass
Measurement of photosynthetic quantum yield and non-photochemical quenching
-
The completely extended first leaf from the top of each seedling was measured for chlorophyll fluorescence using a portable Dual-PAM-100 modulated fluorometer (WALZ, Germany). Each procedure used at least three plants and was replicated three times. Before measurements, all plants were subjected to darkness for over 30 min. The chlorophyll fluorescence kinetics curve was evaluated in the dual-emission mode (Fluo + P700). The saturating light pulse had an intensity of 7,500 μmol m–2 s–1 and lasted for 0.7 s. During this interval, we assessed the least fluorescence yield under dark-adapted conditions (F0), with all electrical gates in the open state. All electron transport pathways turned photosynthetic energy into heat and fluorescence during a 3-s saturating pulse of 3,000 μmol m−2 s−1. We measured the dark-adapted maximum fluorescence yield (Fm). Following this, the maximum fluorescence yield (Fm), minimum fluorescence yield (F0), and steady-state fluorescence yield (Fs) under light-adapted conditions were measured in Areca leaves exposed to an active light intensity of 1,000 μmol m−2 s−1. Using the equation Fv/Fm = (Fm – F0)/Fm, the maximal quantum yield of PSII photochemistry was calculated. Meanwhile, PSII photochemistry's effective quantum yield (ΦPSII) was determined using the formula (Fm' – Fs)/Fm'. The photochemical quenching coefficient (qP) was estimated using the equation qP = (Fm' – Fs)/(Fm' – F0'), whereas non-photochemical quenching (NPQ) was derived using the formula NPQ = (Fm – Fm')/Fm'. The electron transport rate (ETR) was determined using the equation ETR = ΦPSII × PPFD × 0.5 × 0.84, where 0.5 represents the fraction of excitation energy allocated to PSII, and 0.84 denotes the proportion of incident light absorbed by the leaves. Measurements of Fv/Fm, Fv0/Fm0, ETR, qP, qN, and NPQ were carried out and compared between the experimental and control groups.
Measurements of chlorophyll (a, b, ab) and carotenoid contents
-
Chlorophyll content and carotenoids were quantified as previously described with slight modifications[25]. Fresh leaf samples (0.5 g) were homogenized in 1 mL of 80% acetone and then stored in the dark until fully decolorized. The homogenate underwent centrifugation at 6,000 × g for 10 min at 4 °C. After removing the supernatant, spectrophotometers were used to evaluate the absorbance values for chlorophyll a, b, carotenoids (Car), and total chlorophyll content at 645, 663, and 440 nm, respectively.
Measurement of relative water content (RWC)
-
RWC was determined by weighing fresh leaves from three plants instantly. The leaf samples were soaked for 12 h in distilled water to attain complete turgidity, following which their turgid weight (TW) was noted. The final dry weight was determined by drying the samples at 65 °C. RWC was calculated using the following equation[26]:
$ \mathrm{R}\mathrm{W}\mathrm{C}\left(\text{%}\right)=\dfrac{\mathrm{F}\mathrm{r}\mathrm{e}\mathrm{s}\mathrm{h}\;\mathrm{w}\mathrm{e}\mathrm{i}\mathrm{g}\mathrm{h}\mathrm{t}\;\left(\mathrm{F}\mathrm{W}\right)-\mathrm{D}\mathrm{r}\mathrm{y}\;\mathrm{w}\mathrm{e}\mathrm{i}\mathrm{g}\mathrm{h}\mathrm{t}\;\left(\mathrm{D}\mathrm{W}\right)}{\mathrm{T}\mathrm{u}\mathrm{r}\mathrm{g}\mathrm{i}\mathrm{d}\;\mathrm{w}\mathrm{e}\mathrm{i}\mathrm{g}\mathrm{h}\mathrm{t}\;\left(\mathrm{T}\mathrm{W}\right)-\mathrm{D}\mathrm{r}\mathrm{y}\;\mathrm{w}\mathrm{e}\mathrm{i}\mathrm{g}\mathrm{h}\mathrm{t}\;\left(\mathrm{D}\mathrm{W}\right)}\times 100 $ Real-time PCR verification
-
To validate the quality of the transcriptome data, 15 DEGs were randomly selected for qRT-PCR analysis. The plant RNA extraction kit (DP437, TIANGEN, Beijing, China) extracted total RNA from plant materials. RNA concentration and purity were evaluated using a NanoDrop 2000 spectrophotometer (KAIAO, Beijing, China). Reverse transcription was performed with the TIANScript RT Kit (Code No. KR104, TIANGEN, Beijing, China), and the qRT-PCR program was set up according to the manufacturer's instructions. Primer Premier 6.0 software was used to design pair primers with actin as a control primer (Supplementary Table S1). Each experiment included three biological and technical replicates.
Statistical analysis
-
Statistical significance (p < 0.05), degree of freedom (df), and F-values were determined using one-way analysis of variance (ANOVA) followed by Duncan's test in SPSS Statistics 26.0. The data was described using the mean ± standard deviation (SD) of the three biological replicates. Figures were created using GraphPad Prism 9.5. The effects were considered statistically significant when the p-value was less than 0.05. Statistical differences are denoted by lowercase letters on the mean values.
-
A. catechu seedlings exposed to water deficit conditions experienced notable morphological alterations. Over the 30-d DS period, a progressive decline in leaf and root water content resulted in visible yellowing of the leaf margins, followed by widespread yellowing of the leaves. These morphological changes indicated the negative impact of water scarcity on the seedlings' growth and development. Compared to non-treated controls, pretreatment with 200 μmol/L melatonin for 30 d did not significantly alter the morphology of leaves in treated plants. However, a prominent increase in root growth was observed in melatonin-treated plants (Fig. 1a). The concentrations of antioxidant enzymes, including SOD, POD, CAT, and APX, were analyzed in Areca leaves to determine the impact of MT on mitigating DS. The results showed that DS considerably elevated the levels of CAT (Fig. 1b), SOD (Fig. 1c), POD (Fig. 1d), and APX (Fig. 1e) in Areca leaves. The administration of melatonin during DS further elevated the levels of CAT, SOD, and APX in leaves, whereas the levels of POD decline. The levels of SOD and POD were not substantially elevated by melatonin treatment alone in the leaves. MDA and H2O2 are byproducts of cellular metabolism and signal a plant's ability to mitigate ROS under various stress conditions. Therefore, the concentrations of oxidant components, including MDA and H2O2, in Areca leaves were analyzed to determine the effect of MT on alleviating DS. The results demonstrated that DS significantly elevated MDA (Fig. 1j) and H2O2 (Fig. 1g) levels in Areca leaves.
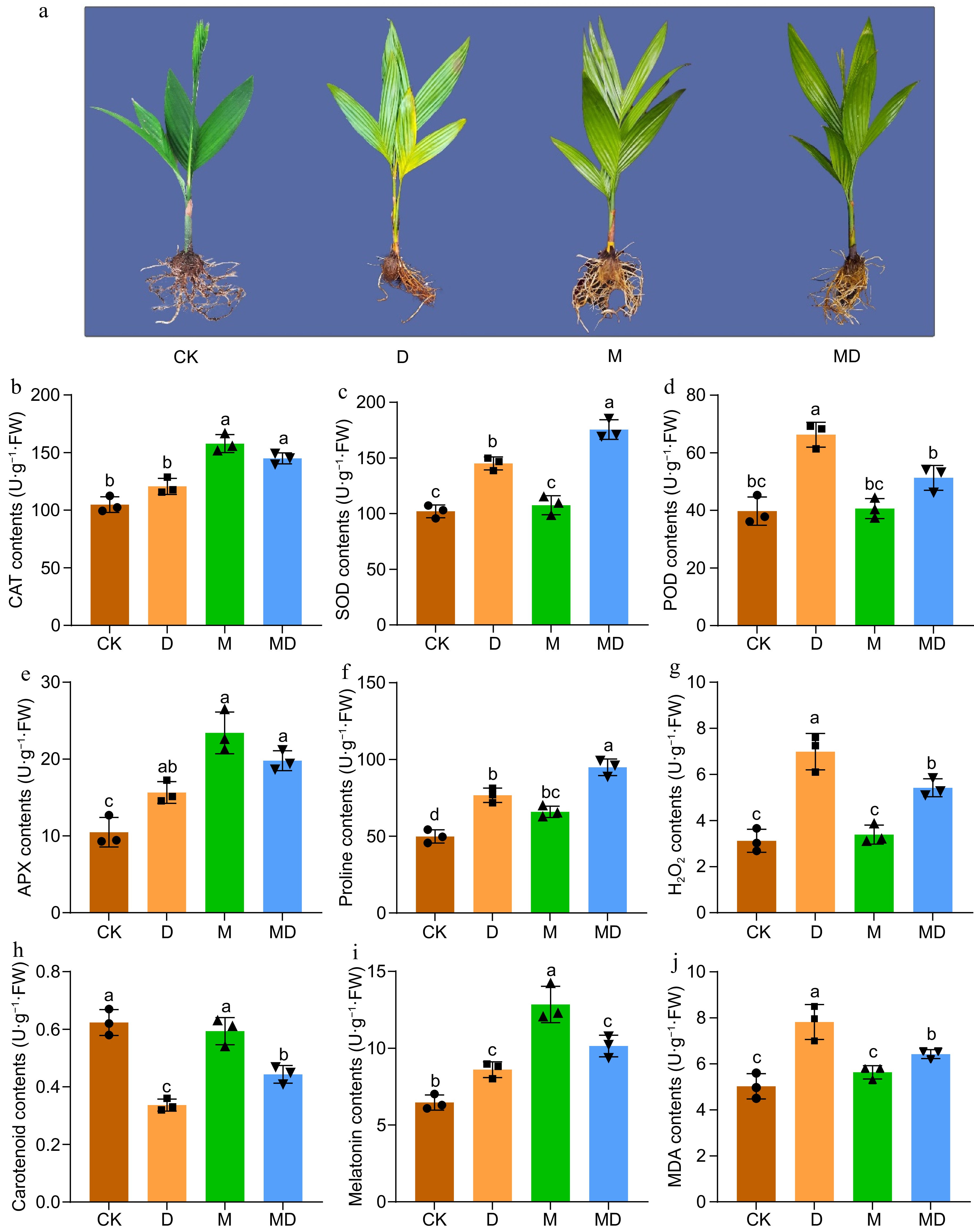
Figure 1.
This figure illustrates the phenotypic and biochemical responses of A. catechu seedlings subjected to drought stress (D) and exogenous melatonin (M) treatment following drought stress (MD). The experiment included four treatments: control (CK), melatonin (M), drought stress (D), and drought stress combined with melatonin (MD). (a) After a 30-d drought stress period, morphological observations were recorded. Subsequently, the activities of key antioxidant enzymes were assessed, including (b) catalase (CAT), (c) superoxide dismutase (SOD), (d) peroxidase (POD), and (e) ascorbate peroxidase (APX). Additionally, the levels of (f) proline, (g) hydrogen peroxide H2O2, (h) carotenoids, (i) endogenous melatonin, and (j) malondialdehyde (MDA) were measured. Each bar in the figure represents the mean ± standard deviation based on three biological replicates (n = 3). Statistically significant differences between treatments are denoted by different letters above the bars (p < 0.05), determined through Duncan's multiple range test. Treatment comparisons were performed using one-way ANOVA with a significance threshold of p < 0.05.
Furthermore, the contents of proline (Pro), carotenoids, and melatonin (Fig. 1f, h & i) were measured in Areca leaves. The results showed that proline and melatonin levels increased under DS and MT treatment compared to the control group. In contrast, the contents of carotenoids were lower under drought and post-melatonin DS treatment.
Role of melatonin in sustaining photosynthetic performance in A. catechu during drought
-
RWC and Photosystem II (PSII) activity are valuable indicators of plant water status and photosynthetic efficiency. They can provide insights into a plant's overall health and metabolic activity. To assess the effectiveness of Ex-MT in reducing DS, RWC, chlorophyll content, and photosynthetic parameters, we evaluated Areca seedlings treated with Ex-MT under drought conditions (Fig. 2). Our results demonstrate that melatonin pretreatment significantly increased RWC in plants compared to the control group. As DS progressed, RWC decreased across all groups. In contrast to the drought-stressed group, the melatonin-pretreated plants showed a considerably greater RWC, showing improved water retention capacity. The relative water content (RWC) decreased by as much as 41% during 30 d of water scarcity, whereas plants administered melatonin (MD) exhibited a substantial increase in RWC, elevating by 52%. The control group (CK) and melatonin (M) treated plants had elevated levels of relative water content (RWC) at 64% and 63%, respectively, after 30 d (Fig. 2a). Chlorophyll fluorescence parameters and chlorophyll contents analysis are powerful tools for measuring the photosynthetic performance of plants. Several variables were assessed and contrasted between the control and experimental groups, encompassing maximum chemical efficiency (Fv/Fm), photochemical efficiency of Photosystem II (Y(II)), photochemical chlorophyll fluorescence quenching (qP), non-photochemical quenching (NPQ), electron transport rate (ETR), and chlorophyll a, chlorophyll b, and the chlorophyll a/b ratio. As water scarcity increased, plants under DS showed a significant drop in chlorophyll concentrations. Nevertheless, these adverse effects were mitigated with melatonin administration. The results indicate that the maximum quantum efficiency of PSII (Fv/Fm) significantly decreased under drought conditions (D), reaching a value of 0.28 after 30 d of water deprivation. This substantial decline suggests severe impairment of PSII functionality due to DS. However, pretreatment with melatonin (MD) slightly mitigated the negative impact of drought, with Fv/Fm values rising to 0.33, demonstrating melatonin's role in enhancing plant resilience to drought. In contrast, no substantial changes were observed between the control group (CK) and the melatonin-treated group (M), with Fv/Fm values of 0.81 and 0.83, respectively (Fig. 2b). These findings suggest that under non-stress conditions, melatonin does not influence PSII efficiency. At the same time, it provides a protective effect under DS. Similarly, the PSII (Y(II)), qP, and ETR showed substantial declines under DS, decreasing to 0.06, 0.15, and 10, respectively, following 30 d of water deprivation. Pretreatment with melatonin resulted in a moderate improvement in (Y(II), raising its value to 0.12 after 30 d, indicating some recovery in PSII functionality (Fig. 2c). However, no significant change was observed in the electron qP and ETR, which remained constant at 0.16 and 11, suggesting that melatonin did not enhance the qP and ETR processes under prolonged drought conditions (Fig. 2d, f). The results demonstrate a progressive increase in non-photochemical quenching (NPQ) as the duration of the water deficit increased. After 30 d of withholding water, NPQ reached a peak of 0.50. Notably, the pre-melatonin effect on NPQ remained relatively low and did not decline following the 30-d DS period. In contrast, the NPQ values in the control and melatonin-only groups exhibited no significant change (Fig. 2e). Similar to other chlorophyll-related parameters, chlorophyll content decreased under DS, while pretreatment with melatonin increased chlorophyll levels. The concentrations of chlorophyll a, chlorophyll b, and the chlorophyll a/b ratio were substantially reduced under the drought situation, recording values of 0.32, 0.26, and 0.81, respectively, in contrast to the control group (CK), which displayed values of 0.76, 0.46, and 1.3. However, pretreatment with melatonin followed by drought stress (MD) improved chlorophyll levels, corresponding values of 0.57, 0.35, and 1.0. No significant changes were observed between the control and melatonin-only groups (CK vs M) (Fig. 2g−i).
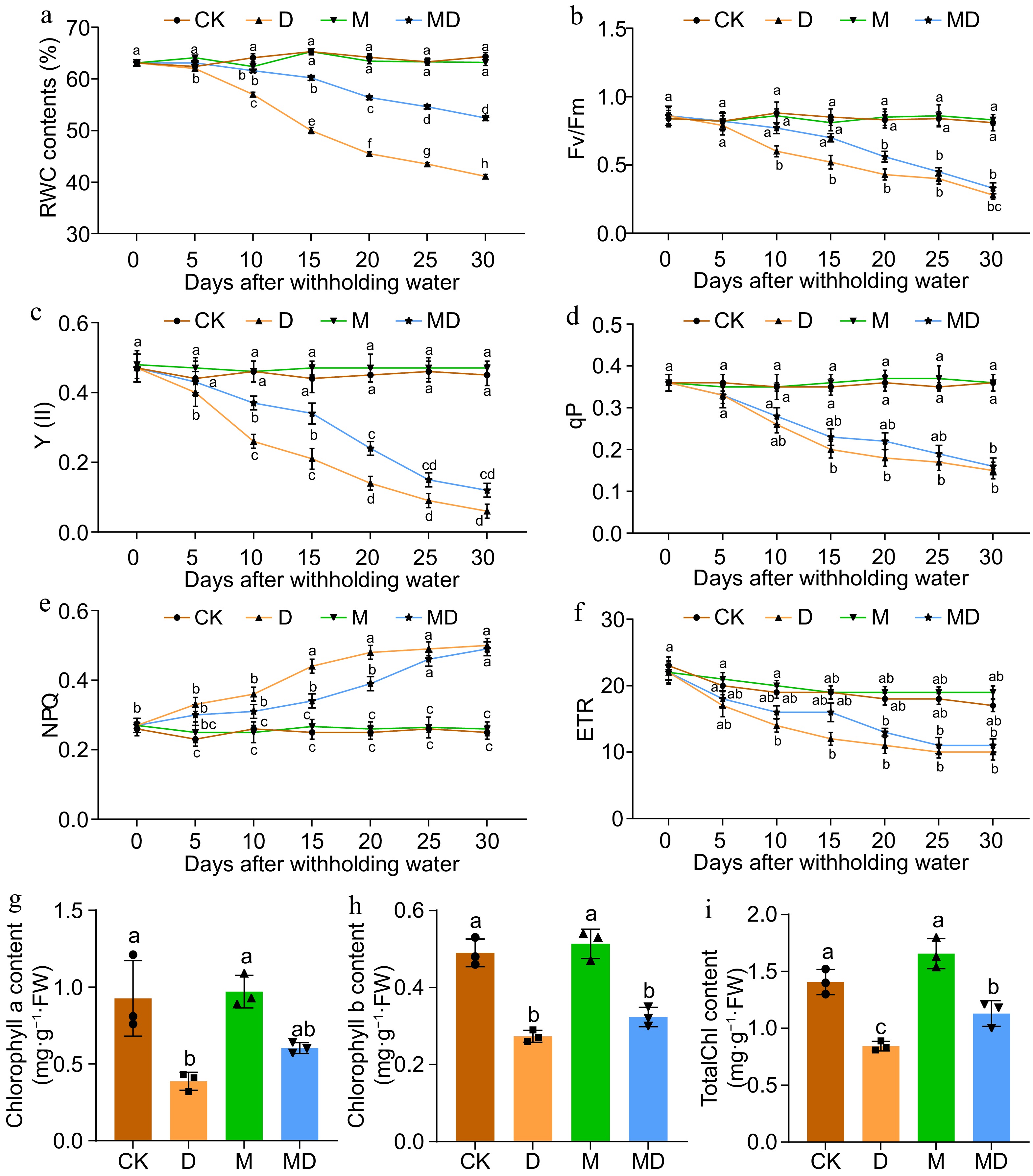
Figure 2.
This figure presents the impact of drought stress (D) and exogenous melatonin (M) treatment on photosynthetic efficiency and chlorophyll content in A.catechu seedlings. The experiment included four treatments: control (CK), melatonin (M), drought stress (D), and drought stress combined with melatonin (MD). The following parameters were measured: (a) relative water content (RWC), (b) maximum quantum yield of PSII photochemistry (Fv/Fm), (c) effective quantum yield of PSII photochemistry (Y(II)), (d) photochemical quenching (qP), (e) non-photochemical quenching (NPQ), (f) electron transport rate (ETR), (g) chlorophyll a content, (h) chlorophyll b content, and (i) total chlorophyll content. Each bar in the figure represents the mean ± standard deviation based on three biological replicates (n = 3). Statistically significant differences between treatments are denoted by different letters above the bars (p < 0.05), determined through Duncan's multiple range test. Treatment comparisons were performed using (a)−(f) two-way ANOVA, and (g)−(i) one-way ANOVA with a significance threshold of p < 0.05.
RNA sequencing analysis of A. catechu seedlings under normal drought stress and pretreatment with melatonin followed by drought stress imposition
-
After DS and melatonin pretreatment, RNA-seq transcriptome analysis was performed on all A. catechu seedling samples. Each sample yielded at least 5.71 GB of clean data (97.7%). The total repeatability and quality score of bases in each sample was 97.7% and 99.6% for Q30 and Q20 base percentages of clean data. Moreover, the GC content of all readings was above 45%. Additionally, clean reads of each sample were mapped using HISAT2[27] to a specified reference genome (Areca_catechu.customer_P417.genome.fa.) (Supplementary Table S2). StringTie[28] was used to build the mapped readings. In addition to the mapping findings, gene structure optimization analysis, alternative splicing prediction, and new gene discovery were conducted. The sample mapping ratios to the reference genome ranged from 94.08% to 97.21%. A total of 27,738 genes were successfully identified, including 3,341 annotated novel genes that were annotated with potential functions throughout this procedure (Supplementary Table S3). The mean sample-to-genome blast ratio was 97.52%, and the mean match rate was 86.56%. All compiled unigenes were queried against databases, including KEGG, COG, GO, KOG, Swiss-Prot, Pfam, NR, and eggNOG, for annotation purposes. In 33,823 annotated unigenes, 3,341 were identified as new genes in at least one database (Supplementary Table S4).
Analysis of differentially regulated genes
-
Fragments per kilobase of exon per million fragments mapped (FPKM) values were used to quantify gene expression level. A substantial positive correlation (r > 0.90) between replicates was found in the correlation analysis of the nine samples, showing five different groups and excellent repeatability and reliability of the transcriptome data (Supplementary Fig. S1). A strict criterion was implemented, requiring a fold change (FC) > 2, FDR < 0.01, and a nucleotide length ≥ 200 bp to identify differentially expressed genes (DEGs). Pairwise sample comparisons revealed 10,145 differentially expressed genes (DEGs) (Supplementary Table S5). A total of 2,394 up-regulated and 2,245 down-regulated DEGs were expressed among CK_vs_D and 702 up-regulated and 939 down-regulated DEGs were noted among CK_vs_MD. Similarly, 1,856 up-regulated and 2,009 down-regulated DEGs were observed among D_vs_MD (Fig. 3a). Among the DEGs, the number of up-regulated genes was higher in the DS condition, while a smaller number of DEGs were expressed among control and MD treatments. Additionally, the Venn diagram shows 279 (4.6%) common DEGs differentially expressed among the three comparison categories (Fig. 3b).
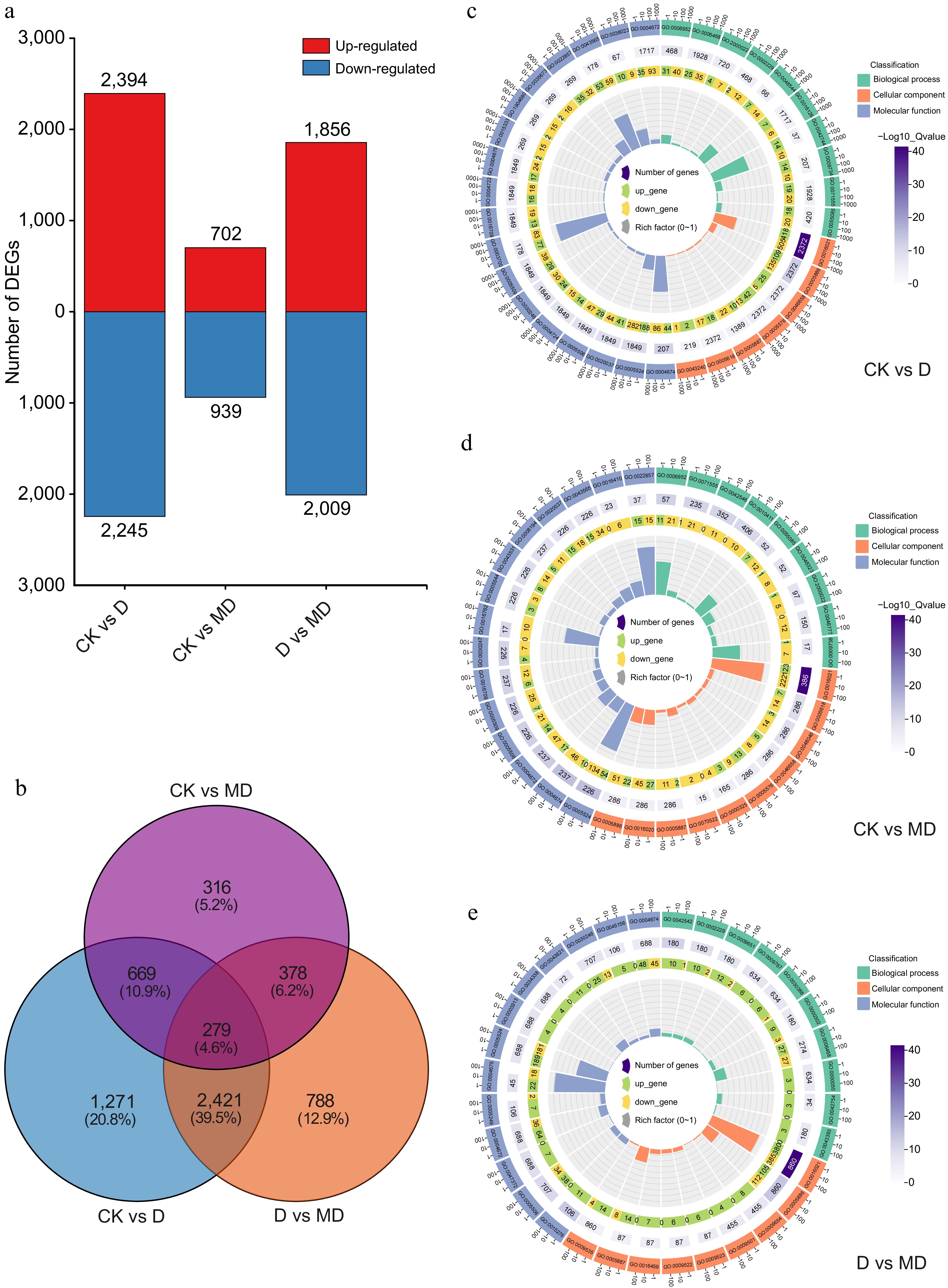
Figure 3.
Overview of differentially expressed genes (DEGs) and GO enrichment analysis in response to drought stresses and melatonin treatment in A. catechu. (a) Common DEGs under DS and melatonin treatment conditions. (b) Commonly expressed genes in a comparison Venn diagram. (c) - (e) Top GO term enriched DEGs analysis found via comparing the D vs MD, CK vs D, and CK vs D comparison.
GO and KEGG pathway enrichment analysis of DEGs
-
GO enrichment analysis was conducted to elucidate the function of these differentially expressed genes (DEGs). DEGs were divided into biological processes, cellular components, and molecular functions based on their predicted roles. GO analysis in DEGs revealed an enrichment in many GO pathways when comparing CK and D samples. Among the biological processes, the top GO terms were defense response (GO:0006952), protein phosphorylation (GO:0006468), regulation of jasmonic acid mediated signaling pathway (GO:2000022), hydrogen peroxide catabolic process (GO:0042744), and auxin-activated signaling pathway (GO:0009734) were the most enriched terms (Fig. 3c & Supplementary Table S6). The analysis of cellular components revealed that the integral component of the membrane (GO:0016021), plasma membrane, and its components (GO:0005886; GO:0046658; GO:0005887) were the most enriched terms (Fig. 3c & Supplementary Table S7). Similarly, the most enriched GO terms in molecular functions were protein serine/threonine kinase activity, ATP binding, heme and Iron binding (GO:0004674; GO:0005524; GO:0020037; GO:0005506) (Fig. 3c & Supplementary Table S8). The top biological processes GO enrichment DEGs pathways between CK vs DM were defense response (GO:0006952), transmembrane transport (GO:0055085), and regulation of the jasmonic acid mediated signaling pathway (GO:2000022) (Fig. 3d & Supplementary Table S9), the top GO cellular components DEGs pathways were components of plasma and cell membrane, while the most enriched GO terms in molecular functions were protein serine/threonine kinase activity, ATP and Iron binding (GO:0005524; GO:0005506), and protein kinase activity (GO:0004672) (Fig. 3d, Supplementary Tables S10 & S11). We further compared the GO pathways enriching most DEGs between D and MD. The GO results indicated that protein phosphorylation (GO:0006468), response to hydrogen peroxide (GO:0042542), defense response (GO:0006952), an integral component of the membrane and plasma membrane, heme, and iron ion binding and DNA−binding transcription factor activity (GO:0003700) were the top biological, cellular, and molecular enhanced processes (Fig. 3e, Supplementary Tables S12−S14). KEGG enrichment pathway analysis showed that the Plant hormone signal transduction pathway (ko04075) was highly enriched across all three circumstances (Fig. 4a). The top 20 KEGG enriched pathways results of CK vs D showed that plant hormone signal transduction (ko04075), MAPK signaling pathway (ko04016), starch and sucrose metabolism (ko00500), and Flavonoid biosynthesis (ko00941) were mainly enriched under DS conditions (Supplementary Fig. S2 & Supplementary Table S15). The top enrichment pathways in the CK vs DM group were the MAPK signaling pathway (ko04016), plant hormone signal transduction (ko04075), phenylpropanoid biosynthesis (ko00940), plant-pathogen interaction (ko04626), and starch and sucrose metabolism (ko00500) pathways were the most enriched pathways (Supplementary Fig. S3 & Supplementary Table S16). Similarly, in the D vs DM group, the top enrichment pathways were Circadian rhythm – plant (ko04712), Plant hormone signal transduction (ko04075), MAPK signaling pathway (ko04016), as well as starch and sucrose metabolism (ko00500) pathway (Supplementary Fig. S4 & Supplementary Table S17).
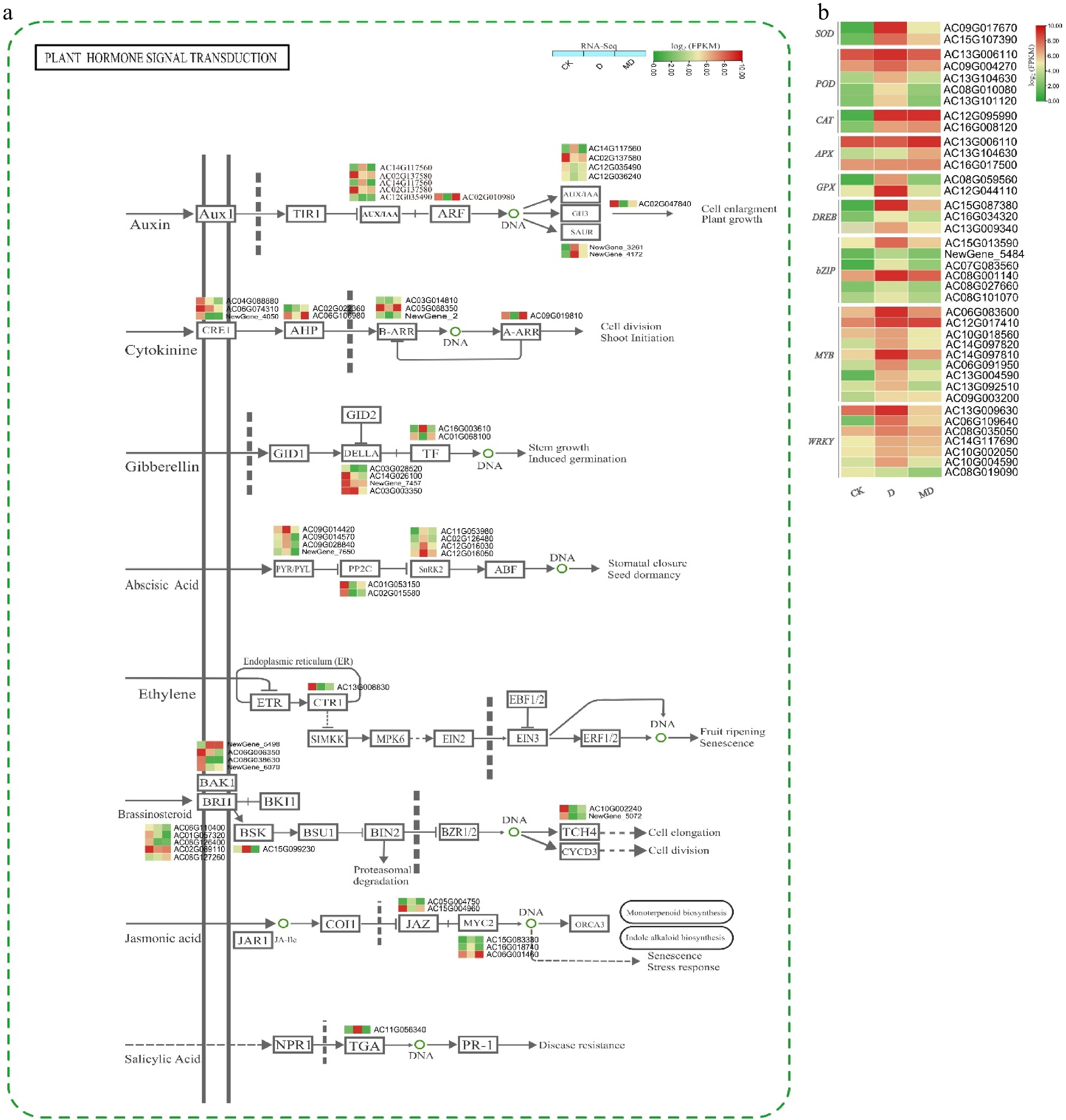
Figure 4.
Expression profiles of hormone signaling and ROS/TF DEGs in A. catechu during drought and melatonin treatment. (a) Key DEGs shaping hormonal signal transduction in A. catechu. (b) The heat map of DEGs related to ROS and TF. The heat map illustrates the fold change derived from log2FPKM values of DEGs. C, control; D, drought; MD, pre-melatonin + drought stress. The colors red and green indicate up-regulated and down-regulated genes, respectively. Values denote mean ± SE (n = 3). The figure adopted using KEGG online data base[29].
DEGs under plant hormones and signal transduction pathway
-
According to the GO and KEGG enrichment analyses, DS and MD treatments significantly influenced plant hormones and the signal transduction pathway (Fig. 4a). The results highlighted significant alterations in the expression of genes related to these pathways, providing insights into how plants manage stress under different conditions. The differential expression analysis revealed that under DS, 157 differentially expressed genes (DEGs) were identified, while under MD conditions, 137 DEGs were detected. These DEGs are crucial for understanding the plant's response mechanisms to water scarcity and the effects of melatonin pretreatment. Drought stresses and melatonin pretreatment significantly altered gene expression in hormone synthesis or signaling. Many genes were linked to auxin, brassinosteroids (BR), cytokinins (CKs), ABA, ethylene (ET), jasmonic acid (JA), and salicylic acid. MT and DS significantly influence the regulation of the ABA signaling pathway. This includes genes encoding ABA receptors (e.g., PYR/PYL), protein phosphatases (e.g., PP2Cs), ABA signaling and stress responses genes, i.e., Serine/threonine protein kinases family SNF1-related protein kinase 2 (SNRK2) and transcription factors (e.g., ABF/AREB).
Expression of DEGs related to transcription factors (TFs) and ROS
-
A comprehensive analysis of transcription factors (TFs) associated with DS response in A. catechu revealed 3289 TFs belonging to 207 families. Among these, 956 genes were differentially expressed (DEGs) (Supplementary Fig. S5 & Supplementary Tables S18, S19). Several TF families responded substantially to DS, including MYB, NAC, bHLH, WRKY, C2H2, and bZIP. Notably, most TFs were upregulated in melatonin-treated and drought-stressed plants, suggesting their involvement in stress-responsive pathways mediated by melatonin signaling. There were 110 differentially expressed genes (DEGs) linked to ROS were identified. MT and DS synergistically induced the expression of antioxidant enzymes. Both treatments significantly upregulated the expression of SOD, POD, CAT, APX, and GPX (Fig. 4b).
WGCNA-based co-expression network construction
-
This research used WGCNA to conduct a co-expression network analysis of all differentially expressed genes (DEGs) to comprehensively understand the gene regulation network related to growth and phytochemical parameters (Fig. 5). Differentially expressed genes (DEGs) were clustered into six distinct co-expression modules: MEgreen, MEblack, MEblue, MEbrown, MEmagenta, and MEturquoise (Fig. 5a). The gene numbers of six modules revealed that MEmagenta and MEturquoise have the highest number of expressed DEGs (167 and 1303 DEGs, respectively), while MEblue and MEblack have shown downregulated DEGs. Most of the physiochemical parameters were positively correlated with DEG expression in MEmagenta and MEturquosie except for total chlorophyll content. The higher expressed modules (MEturquoise and MEmagenta) DEGs were drowned by Cytoscape (Fig. 5c, d).

Figure 5.
Module trait weighted gene co-expression network analysis (WGCNA). (a) Eigengene adjacency heatmap for each module, illustrating the correlation among the modules. Each column denotes distinct processing conditions. Each row represents a unique gene of the module. The Pearson correlation coefficient describes the relationship between the two variables in each cell, with the p-value provided in parenthesis. The cell color spectrum varies from red, signifying a robust positive association, to blue, denoting a strong negative correlation. (b) Eigengenes modules hierarchical clustering dendrogram; Genes fit into one of six modules. The top image shows the sequence diagram. The colorful gene modules and their bases are shown at the bottom of the figure. (c) Hub-gene clustering network of MEturquoise module. (d) Hub-gene clustering network of MEmagenta. Genes in red are at the core and have strong interactions, whereas genes in orange are near the periphery.
Moreover, the Plant hormone signal transduction (ko04075) and flavonoid biosynthesis (ko00941) pathways were significantly enriched in MEmagenta. In contrast, the Plant hormone signal transduction (ko04075) and starch and sucrose metabolism (ko00500) were the highly enriched pathways in MEturquoise modules (Supplementary Tables S20, S21). Furthermore, several transcription factors were analyzed inside these modules, with the majority associated with WRKY, NAC, MYB, bHLH, and bZIP families. Subsequently, the two highly expressed modules were chosen for the hub gene correlation network based on signed module membership (MM) values and trait significance (TS) to identify a strong connection among hub genes. The degree of connection between gene pairs varies significantly within a gene co-expression module. We only kept the edges highly connected to the hub genes and have the most incredible weights to improve the display of these networks. This allowed us to focus on the most important linkages.
Validation of DEGs via qRT-PCR analysis
-
The randomly selected genes included essential melatonin pathway genes (ASMT), phytohormone-related genes (NCED, PYR/PYL, SNRK2 ), Flavonid biosynthesis pathway gene (CHS), and key ROS (SOD, POD, CAT, APX, GPX) and TFs (WRKY1, bHLH, MYB, bZIP) related gene response to DS. The expression trends observed by qRT-PCR analysis were highly correlated (Pearson correlation > 0.9) with the RNA-Seq results for most genes (Fig. 6). This consistency between qRT-PCR and RNA-Seq data validates the reliability of the transcriptome analysis.
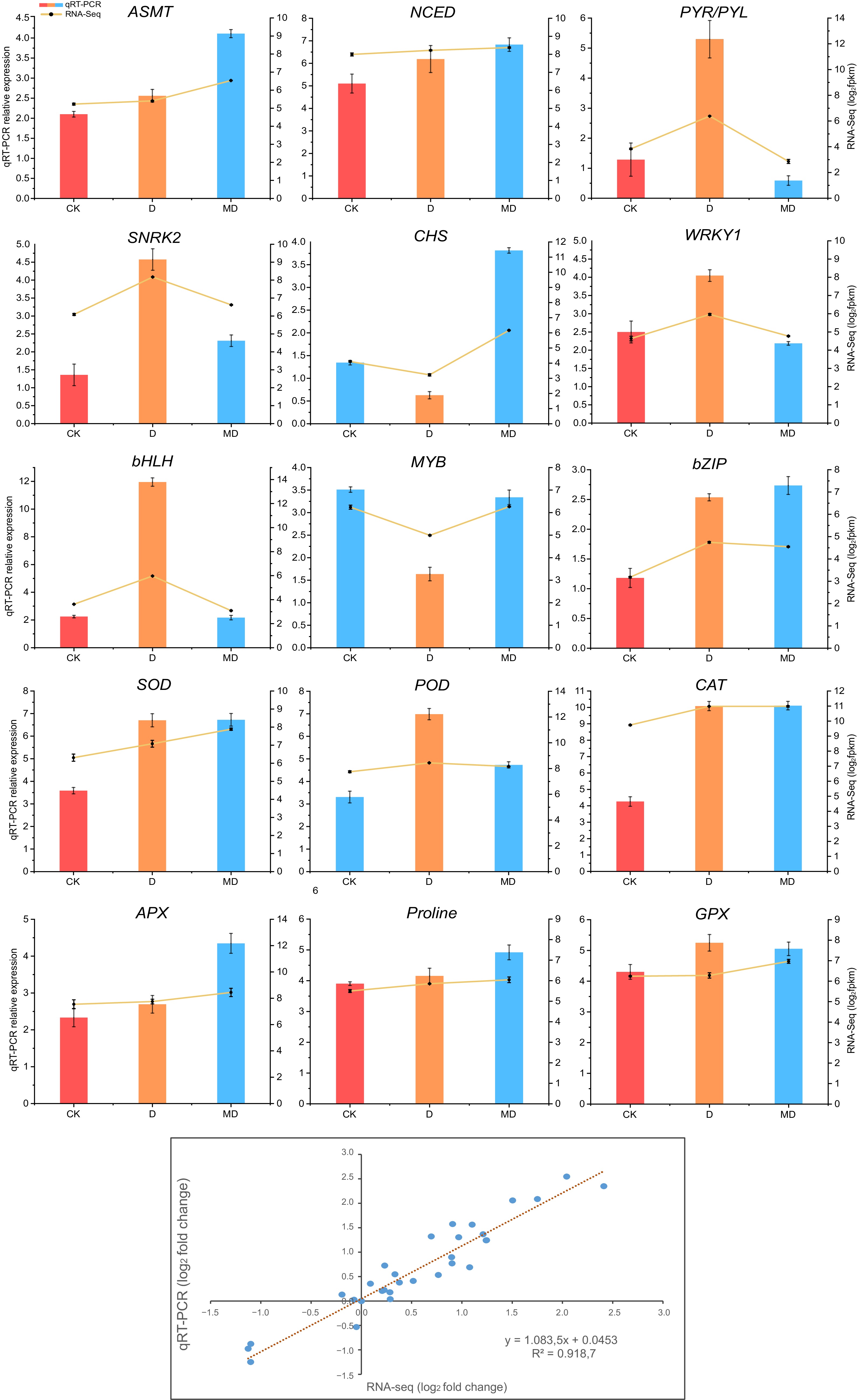
Figure 6.
Validation of differentially expressed genes associated with phytohormones, ROS, and transcription factors by RT-qPCR, followed by RNA-seq data. Analysis of correlation between RNA-seq and qRT-PCR methodologies. Log2 fold changes of RNA-seq data (x-axis) are displayed against log2 fold changes of qRT-PCR data (y-axis). The error bars represent the standard error of three biological replicates mean ± SE (n = 3). C, control; D, drought; MD, pre-melatonin + drought stress.
-
Drought stress is widely known to trigger a series of physiological, molecular, and biochemical changes in plants[30]. Previous studies have demonstrated that exogenous melatonin, a phytohormone with antioxidant properties, plays a key role in mitigating drought-induced oxidative stress in several crops like maize[31], rice[8,32], cotton[33], soybean[34,35], perennial ryegrass[36], sorghum[37], wheat[38], etc. However, the specific molecular mechanisms by which melatonin influences drought tolerance in A. catechu remain insufficiently understood. This study addresses this gap by investigating the effects of exogenous melatonin (Ex-MT) on A. catechu under drought stress at morphological, physiological, biochemical, and transcriptomic levels.
Drought disrupts plant hormone balance, affecting growth and development. Melatonin regulates physiological functions and boosts plant vigor during stress. Our results revealed that compared to Ck, the melatonin-treated plants exhibited higher stem diameters, root biomass, and longer roots. (Fig. 1a). These results suggest that melatonin enhances root system architecture, promoting better water and nutrient uptake under drought conditions consistant with the prior study which evidence carbon uptake through improved photosynthetic performance and enhanced root development[39,40].
Moreover, under abiotic stresses, plants activate enzymatic and non-enzymatic defense mechanisms, maintaining cellular homeostasis and resisting abiotic stress[41]. Our findings show that Ex-MT significantly increased antioxidant enzyme activities, reducing H2O2 and MDA levels in melatonin-treated plants (Fig. 1), these results align with prior research identifying melatonin's role in mitigating oxidative stress by enhancing antioxidant activity and reducing ROS-induced damage[39,42]. However, plants synthesize endogenous melatonin and proline, which act as osmoprotectants by maintaining cellular turgor under stress and promoting the synthesis of essential proteins involved in stress response mechanisms[43]. The melatonin pretreatment in our results was shown to improve proline and melatonin accumulation in Areca seedlings subjected to DS, indicating its potential to reduce the detrimental impacts of drought (Fig. 1f), consistent with previous findings, which describe suitable MT dosage enhanced both proline and endogenous melatonin content under drought conditions[16,31,44].
Additionally, DS triggers stomatal closure to minimize water loss, but this response also limits respiration and photosynthesis[45]. In contrast, studies have shown that melatonin enhances stomatal conductance, relative water content (RWC), and net photosynthetic rate under drought conditions[23,46]. Our findings confirm that melatonin administration improved RWC in A. catechu seedlings (Fig. 2a), likely by regulating stomatal closure, thus balancing water retention with gas exchange. This aligns with previous research showing that melatonin mitigates water loss through stomatal regulation and increases RWC[47,48]. Plant PSII photosynthetic capability and energy conversion efficiency are measured by chlorophyll fluorescence[49]. Several investigations have indicated that acute or long-term water shortage leads to photoinhibition in the PSII system[31,46]. In our study, drought-stressed A. catechu plants exhibited significant decreases in Fv/Fm, Y(II), and ETR, along with increased non-photochemical quenching (NPQ) (Fig. 2), indicating severe PSII impairment. Melatonin treatment (Ex-MT) significantly mitigated these effects, as treated plants demonstrated higher Fv/Fm, Y(II), and ETR levels, with reduced NPQ compared to untreated drought-stressed plants (Fig. 2). These improvements in PSII performance suggest that melatonin protects the photosynthetic machinery from drought-induced photoinhibition, consistent with findings in other crops such as maize and rice[42,50,51]. Furthermore, our findings demonstrated that melatonin treatment mitigates the detrimental effects of DS, enhancing chlorophyll and carotenoid levels in comparison to control plants exposed to DS (Fig. 2). These findings are inconsistent with studies demonstrating melatonin's ability to prevent chlorophyll degradation and maintain photosynthetic efficiency under drought conditions[52].
RNA-Seq provides a global transcriptome profile, gives insights into molecular regulation in response stresses, and helps plant biologists to expand new insights into molecular mechanisms and responses to biotic and abiotic events.
Our RNA-sequencing analysis revealed that Ex-MTtreatment modulates gene expression patterns in A. catechu, particularly those associated with secondary metabolites, plant hormone signaling, and transcription factors. Differentially expressed genes analysis showed a significant upregulation of stress-responsive pathways under DS and MT treatment (Fig. 3a, b). These findings indicate that MT enhances stress tolerance by activating essential metabolic and hormonal pathways[53,54]. Moreover, WGCNA analysis with RNA-Seq data effectively identifies essential genes and their networks associated with specific traits, such as drought stress[55]. Our WGCNA analysis identified two critical modules, MEmagenta and MEturquoise, strongly related to drought-tolerance traits in A. catechu (Fig. 5a). These modules exhibited a positive correlation with the expression of DEGs for most physiochemical parameters.
The WGCNA modules along with KEGG pathway enrichment analysis indicating the enrichment of key pathways like Flavonoid Biosynthesis, MAPK, and Plant Hormone Signal Transduction, highlighting the importance of hormonal regulation and secondary metabolism in stress adaptation.
The enrichment of Flavonoid Biosynthesis Pathways in the MEmagenta and MEturquoise WGCNA modules highlight the significance of secondary metabolites in combating drought-induced oxidative damage. Flavonoids are well-documented antioxidants that mitigate ROS accumulation, and their biosynthesis under Ex-MT treatment underscores the protective metabolic adaptations in A. catechu. This adaptive mechanism is also supported by the upregulation of DEGs linked to antioxidant enzymes such as CAT, SOD, and POD[56]. The activation of the MAPK Signaling Pathway offers another layer of insight into the molecular mechanisms underlying drought tolerance. MAPK cascades are crucial for transducing stress signals and initiating transcriptional responses. The upregulation of MAPK-related DEGs (Fig. 4) in our study suggests that Ex-MT primes the plant's stress signaling network, enabling a more robust response to environmental challenges. The Plant Hormone Signal Transduction Pathway illustrates melatonin's role in modulating hormonal regulation, including auxin (IAA), gibberellin (GA), and abscisic acid (ABA) (Fig. 4a). ABA, in particular, plays a pivotal role in drought stress responses, primarily by regulating stomatal closure and reactive oxygen species (ROS) production[57,58].
However, our data revealed that melatonin mitigates excessive ABA accumulation by upregulating PP2C, a negative regulator of ABA biosynthesis, thereby alleviating oxidative stress and improving photosynthetic efficiency[59−61]. This regulation is consistent with prior studies in crops like maize, oats, and Medicago truncatula, where melatonin similarly reduced ABA-induced stress responses[60,62,63]. Furthermore, Ex-MT reversed drought-induced overexpression of SnRK2 genes, promoting better stomatal regulation and physiological recovery. These results underscore melatonin's ability to fine-tune ABA signaling and reduce the negative impact of excessive ABA accumulation. A similar previous study also showed that SnRK2 genes are up-regulated during DS while Ex-MT supplementation decreased the ABA content and its biosynthesis gene expressions in tomato and Arabidopsis[64,65]. Melatonin also significantly modulated the expression of critical transcription factors (TFs) involved in stress responses, such as AP2/ERF, NAC, WRKY, MYB, and bZIP (Fig. 4b, Supplementary Fig. S5, Supplementary Table S18). These TF families are well-known for promoting the expression of genes linked to abiotic stress tolerance[66]. Notably, the upregulation of DREB, NAC, and TIFY TFs in melatonin-treated seedlings highlights its role in preparing plants for environmental challenges. These findings align with studies on Arabidopsis and tomato, where melatonin pre-treatment improved drought resilience through TF regulation[67,68].
Overall, the present study provides strong evidence that exogenous melatonin can enhance DS in A. catechu by improving antioxidant activity, photosynthetic performance, and regulating metabolic pathways. However, this leaves open questions regarding melatonin's effectiveness under multiple concurrent stressors, such as heat, salinity, or heavy metal exposure to A. catechu. Additionally, the relatively short duration of the experiment limits our understanding of the potential long-term impacts and sustainability of melatonin application. Future research should aim to refine melatonin-based strategies by determining optimal application rates, assessing long-term impacts across varying environmental conditions, and evaluating its effectiveness in crops beyond A. catechu to enhance resilience against environmental stress.
-
In summary, the results of this study provide evidence of Ex-MT regulating physiology and gene expression of A. catechu under water deficit conditions. Melatonin significantly increased RWC, chlorophyll, photosynthetic rate, and antioxidant activity to mitigate DS-induced damage. Furthermore, melatonin regulates key metabolic pathways such as MAPK, plant hormonal pathway, flavonoid, isoflavonoid, and steroid biosynthesis pathways. DS and Ex-MT enhance endogenous hormone levels and significantly regulate downstream transcription factors, including AP2/ERF-ERF, WRKY, GOX, TIFY, MYB, and EIN. These findings give significant insights for developing melatonin-based strategies to protect plants from drought and propose a putative mechanism through which melatonin confers protection against abiotic stress in A. catechu.
We thank BMKCloud (www.biocloud.net) for gene expression analysis assistance in this manuscript. We also thanks the National Natural Science Foundation of China (31960064) and The Hainan Normal University Talent Research Start-up Fund Project Funding (HSZK-KYQD-202421) for funding this project.
-
The authors confirm contribution to the paper as follows: project design and supervision: Yinglang W, Guangzhen Z; data collection: Ali A; interpretation of results: Yi QJ; draft manuscript preparation: Khan NM; Statistical analysis: Khan WU. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available on request from the corresponding author.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0009)
-
Received 21 November 2024; Accepted 6 February 2025; Published online 15 April 2025
-
This study presents the first report of the effect of exogenous melatonin at the physiological, morphological, and molecular level in A. catechu L., under drought stress.
Exogenous melatonin treatment significantly improved drought tolerance in A. catechu seedlings, as evidenced by increased relative water content (RWC) (52%), total chlorophyll concentration (19%), and Photosystem II (PSII) activity.
Melatonin enhanced antioxidant enzyme activities and reduced levels of reactive oxygen species (ROS) in A. catechu.
RNA-seq and WGCNA analyses revealed that melatonin modulates key hormonal signaling pathways (including ABA, IAA, GA, CK, JA), and secondary metabolism pathways, particularly flavonoid biosynthesis.
-
# Authors contributed equally: Noor Muhammad Khan, Yiqi Jiang
- Supplementary Table S1 Gene-specific primers used for qRT-PCR.
- Supplementary Table S2 RNA-seq transcriptome clean data.
- Supplementary Table S3 Annotated novel genes.
- Supplementary Table S4 All annotated genes.
- Supplementary Table S5 DEG number.
- Supplementary Table S6 Biological process GO enrichment analysis CK vs D.
- Supplementary Table S7 Cellular process GO enrichment analysis CK vs D.
- Supplementary Table S8 Molecular process GO enrichment analysis CK vs D.
- Supplementary Table S9 Biological process GO enrichment analysis CK vs MD.
- Supplementary Table S10 Cellular process GO enrichment analysis CK vs MD.
- Supplementary Table S11 Molecular process GO enrichment analysis CK vs MD.
- Supplementary Table S12 Biological process GO enrichment analysis D vs MD.
- Supplementary Table S13 Cellular process GO enrichment analysis D vs MD.
- Supplementary Table S14 Molecular process GO enrichment analysis D vs MD.
- Supplementary Table S15 KEGG pathway enrichment analysis CK vs D.
- Supplementary Table S16 KEGG pathway enrichment analysis CK vs MD.
- Supplementary Table S17 KEGG pathway enrichment analysis Dvs MD.
- Supplementary Table S18 Total transcription factors (TFs) identified.
- Supplementary Table S19 DEGs transcription factors (TFs) identified.
- Supplementary Table S20 KEGG enrichment analysis of Magenta modules-related genes.
- Supplementary Table S21 KEGG enrichment analysis of Turquoise modules-related genes.
- Supplementary Fig. S1 Pearson correlation matrix of all RNA-seq samples under control, drought, and melatonin + drought treatments.
- Supplementary Fig. S2 KEGG pathway enrichment analysis of differentially expressed genes (DEGs) between control (CK) and drought (D) conditions.
- Supplementary Fig. S3 KEGG pathway enrichment analysis of differentially expressed genes (DEGs) between control (CK) and melatonin + drought (MD) conditions.
- Supplementary Fig. S4 KEGG pathway enrichment analysis of differentially expressed genes (DEGs) between drought (D) and melatonin + drought (MD) conditions.
- Supplementary Fig. S5 Transcription factor families and protein kinase subfamilies identified in the study.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Khan NM, Jiang Y, Ali A, Khan WU, Zhou G, et al. 2025. Role of melatonin in mitigating drought stress in Areca catechu L.: insights from integrated physiological, biochemical, and transcriptomic analysis. Tropical Plants 4: e016 doi: 10.48130/tp-0025-0009 

Role of melatonin in mitigating drought stress in Areca catechu L.: insights from integrated physiological, biochemical, and transcriptomic analysis
- Received: 21 November 2024
- Revised: 26 January 2025
- Accepted: 06 February 2025
- Published online: 15 April 2025
Abstract: Melatonin is widely recognized for its role in enhancing plant stress tolerance; however, its potential for mitigating drought stress in Areca catechu L. remains underexplored. This study evaluates the application of exogenous melatonin in enhancing drought tolerance in A. catechu L. through comprehensive physiological, biochemical, and transcriptomic analyses. Exogenous melatonin application significantly improved relative water content by 52%, total chlorophyll concentration by 19%, and Photosystem II (PSII) activity under drought conditions. Additionally, melatonin enhanced antioxidative defense systems, improving reactive oxygen species (ROS) detoxification. RNA-seq analysis revealed 2,394 up-regulated and 2,245 down-regulated genes under drought stress compared to control conditions. Exogenous melatonin treatment modulated gene expression, with 702 genes up-regulated and 939 down-regulated compared to untreated drought-stressed plants. Enrichment analyses and weighted gene co-expression network analysis (WGCNA) revealed that exogenous melatonin significantly activated critical pathways, including plant hormone signal transduction (ko04075), flavonoid biosynthesis (ko00941), MAPK signaling (ko04016), and starch and sucrose metabolism (ko00500). Collectively, these findings provide a molecular framework for utilizing exogenous melatonin to provide a drought-resistance in A. catechu.
-
Key words:
- Areca catechu /
- Drought stress /
- Exogenous melatonin /
- Transcriptome /
- Plant hormone