









-
Eggplant (Solanum melongena L.) is a highly nutritional and economically valued vegetable that is widely cultured worldwide. The fruit color of eggplant is a key factor in determining their commerciality, and is an important criterion in influencing consumer choice. Anthocyanins are water-soluble pigments giving the eggplant fruits their purple color, play significant roles in plant life, and contribute to various human health benefits[1,2].
However, the biosynthesis of anthocyanins is strongly modulated by environmental conditions, including light and temperature[3−6]. Both low light and high temperature conditions severely inhibit anthocyanin biosynthesis in eggplant fruit peel, leading to compromised fruit quality[3,7−9]. A series of studies have focused on identifying the molecular mechanisms of the light regulated anthocyanin biosynthesis in plants[10,11]. Recently, Li et al.[9] reported that exogenous methyl-jasmonate (MeJA) application can effectively rescue poor peel coloration under low light conditions, and the underlying regulatory relationship between jasmonate and light signaling was also revealed. However, the underlying molecular mechanisms of high temperature stress-regulated anthocyanin biosynthesis in eggplant are still not well understood. Whether jasmonic acid (JA) also modulates high-temperature-suppressed anthocyanin biosynthesis and the molecular mechanisms involved remain unclear.
Jasmonic acid (JA) and its derived compound 'jasmonates' are plant endogenous hormones that play pivotal roles in plant growth and developmental processes, and participate in abiotic and biotic stress response and resistance in plants, including high temperature, low light, salinity, drought, and phytopathogenic microorganisms[12−14]. Moreover, JA has also been reported as an important positive anthocyanin biosynthesis regulator[15,16]. The underlying molecular mechanisms of JA regulating anthocyanin biosynthesis have been reported. The JA receptor COI1 (CORONATINE-INSENSITIVE PROTEINI) and JAZ repressors (JASMONATE ZIM-DOMAIN) are both core interfaces of JA-regulated anthocyanin biosynthesis[15,16].
JAZ proteins have been identified as critical negative regulatory factors in the JA signal transduction pathway for regulating plant responses and anthocyanin biosynthesis[17,18]. Under normal conditions, the JA content in plants is low, the JAZ proteins bind to the JA signaling regulators to repress the transcription of downstream response genes. When the JA content in plants increased, JAZ proteins could be degraded by the 26S proteasome based on the interaction with COI1, and releasing the JA signaling regulators[18−20].
The JAZ protein family has been identified and characterized in numerous plant species, such as tomato[21], tobacco[22], Chinese cabbage[23], and potato[24]. The JAZ protein family is one of the subfamily of TIFY characterized by the TIFY domain at the N-terminus and a Jas domain (CCT_2) at the C-terminus[25]. Additionally, the response, resistance, or application range of the JAZ gene family in different stress or secondary metabolites were analyzed, including abiotic stresses such as drought[26], and salt[27], biotic stresses such as gray leaf spots[21], Ralstonia solanacearum[22], Plasmodiophora brassicae[28], and secondary metabolite pathways such as ginkgolide biosynthesis[29], and gypenoside biosynthesis[30]. Moreover, JAZ not only responds to JA, but also responds to ABA and SA[31,32]. Overall, JAZ proteins have been widely identified in plants, but little is known about the role of the JAZ family in high-temperature stress-regulated anthocyanin biosynthesis in eggplant.
Here, a genome-wide identification and systematic characterization of the JAZ family were performed in eggplants. A total of 16 SmJAZ proteins were identified, and gene structure, and evolutionary relationships among eggplants, Arabidopsis thaliana, and tomato were analyzed. Their expression patterns of SmJAZs responding to light or high temperature in eggplant peel were also analyzed and SmJAZ9 were selected. Sublocation and the functional role of the SmJAZ9 were analyzed. The molecular mechanisms of SmJAZ9 underlying anthocyanin biosynthesis were also explored. This study improves our understanding of JAZ in light and temperature regulating anthocyanin biosynthesis and provides several potential genes for eggplant breeding.
-
The genome sequence information of eggplant was downloaded from SGN (
https://solgenomics.net/ )[33]. To identify the SmJAZ gene family members in eggplant, Hidden Markov Model (HMM) search and blastp methods were used. Firstly, the HMM files for the TIFY domain (PF06200) and Jas domain (PF09425) involved in JAZ proteins were downloaded from the Pfam database (www.ebi.ac.uk/interpro/entry/pfam ). Then the 'simple hmm search' programme in TBtools software was used to screen the genes with the two domains from the eggplant protein sequence database. Additionally, the 18 AtJAZ protein sequences obtained from the Arabidopsis Information Resource website (www.arabidopsis.org ) served as queries to perform BLASTP searches against the eggplant genome (e-value < 10−5). Finally, the cross-sectional results from the two different approaches were considered as the total SmJAZ genes. Similarly, the SlJAZ gene family in Solanum lycopersicum were also identified.Bioinformatics analysis of SmJAZs
-
A neighborhood phylogenetic tree was constructed using MEGA7.0 with 1,000 bootstrap replicates, incorporating homologous sequences from Arabidopsis and tomato. Gene exon-intron organization was visualized via GSDS 2.0. Conserved functional motifs were identified using NCBI-CD search analysis. The 2,000 bp promoter sequences upstream of the start codon (ATG) of the SmJAZs gene were extracted to predict cis-acting elements and their potential related functions using the PlantCARE website. The genomic collinearity patterns of SmJAZ family genes in eggplant were investigated through replication event analysis using MCScanX software[34]. This computational approach enabled the identification of both intragenomic duplication events and cross-species syntenic relationships. The resultant data were subsequently visualized through dual-platform processing with TBtools.
High-temperature and dark treatments on eggplant
-
Fruits at 6 d post-anthesis (DPA) were selected for the experiment. High-temperature treatment was implemented in a controlled artificial climate chamber. Two regimes were applied: Normal temperature (NT): A diurnal cycle of 18–28 °C, with temperature gradually increasing from 18 to 28 °C between 04:00 and 11:00, maintained at 28 °C from 11:00 to 14:00, gradually decreasing to 18 °C from 14:00 to 20:00, and held at 18 °C from 20:00 to 04:00; High temperature (HT): A diurnal cycle of 28–38 °C, with temperature rising from 28 to 38 °C between 04:00 and 11:00, sustained at 38 °C from 11:00 to 14:00, gradually declining to 28 °C from 14:00 to 20:00, and maintained at 28 °C from 20:00 to 04:00. The peel adjacent to the calyx (1–2 cm region) was collected at three time points: immediately before treatment initiation (0 d, baseline control), and after 1 d and 7 d of continuous high-temperature exposure. The dark treatment was conducted on the eggplant grown naturally in a sunlit greenhouse. To establish differential light exposure, the peel under the calyx region shielded from light was set as the dark treatment, while the middle region peel of fruit exposed to light was set as the light control. Sampling was performed between 10:00 and 11:00 to minimize diurnal metabolic variation. The above samples were flash-frozen in liquid nitrogen, and stored at −80 °C for RNA-seq analysis.
Analysis of SmJAZ gene expression patterns based on RNA-seq data
-
RNA quality was evaluated using the Agilent Bioanalyzer 2100 system (Agilent Technologies, CA, USA) with the RNA Nano 6000 Assay Kit. Following standard procedures, sequencing libraries were prepared and their quality was verified using the same platform. Transcript alignment was performed against reference sequences using Hisat2 (version 2.0.5), followed by transcript assembly with StringTie (v1.3.3b) in reference-guided mode. Gene expression quantification was calculated as FPKM (Fragments Per Kilobase per Million mapped reads), which normalizes for both sequencing depth and transcript length. Expression values were log2-transformed (FPKM) and visualized as heatmaps using MEV software (version 4.3.1).
Subcellular localization
-
The coding sequence of SmJAZ9 was PCR-amplified and cloned into the pHB vector fused with the GFP tag, followed by transformation into Agrobacterium GV3101. For transient expression assays, bacterial suspensions carrying either pHB-GFP (control) or pHB-SmJAZ9-GFP constructs were prepared in infiltration buffer (MS medium, pH 5.8) supplemented with 10 mM MES, 10 mM MgCl2, and 100 μM AS. These suspensions were pressure-infiltrated into 4-week-old N. benthamiana leaves. The GFP fluorescence were detected using a Laser confocal microscope (LEICA-LSM510 Meta).
Transient expression assay in eggplant peel
-
To silence SmJAZs genes, target-specific primers were designedusing the SGN VIGS Tool (
https://vigs.solgenomics.net/ ). Next, 150~300 bp fragments were subsequently cloned into the TRV2 vector. For infiltration, Agrobacterium cultures harboring either pHB-SmJAZ9 or empty vector, and TRV1 mixed with TRV2-SmJAZ9 or empty vector were injected into the following eggplant peels. The purple peel of a photodependent anthocyanin biosynthesis cultivar 'Shaokao eggplant' was used. To establish a light-responsive phenotypic contrast, developing fruits were subjected to 25-d post-anthesis light-deprivation treatment using black paper enclosures, generating etiolated peels. Post-infiltration specimens underwent 12-h dark adaptation before exposure to sub-saturating irradiance (60 μmol·m−2·s−1). Anthocyanin content and peel color were quantitatively assessed 72 h post-treatment using three biological replicates across three independent experimental iterations.Extraction and quantification of anthocyanins
-
Anthocyanins were extracted and the content of anthocyanins was quantified following the methods described in a previous study[35].
Yeast two-hybrid assay
-
For yeast two-hybrid analysis, the coding sequences of SmJAZ9 and SmMYC2 were cloned into pGBKT7 as bait constructs, while SmMYB113, SmMYC2, and SmCOI1 CDSs were inserted into pGADT7 as prey vectors. Empty vectors served as negative controls. After co-transforming bait and prey plasmids into Y2HGold yeast cells, transformants were first selected on SD/-Trp/-Leu (SD-TL) medium at 30 °C for 3−4 d, followed by screening on stringent SD/-Trp/-Leu/-His/-Ade (SD-TLHA) plates for 2−3 d under the same conditions.
BiFC assays
-
For BiFC analysis, the coding sequences of the SmJAZ9 and SmMYC2 were amplified and inserted into the pY2C vector, while the coding sequences of the SmMYB113, SmMYC2, and SmCOI1 were amplified and inserted into the pY2N vector. The recombinant constructs were transferred to Agrobacterium GV3101. The empty pY2N and pY2C vectors were used as a negative control. Agrobacterium cultures harboring both pY2C-genes and pY2C-genes (or empty vector controls) were resuspended in an infiltration buffer and co-infiltrated into 4-week-old N. benthamiana leaves. The YFP fluorescence were detected using a laser confocal microscope (LEICA-LSM510 Meta).
Firefly luciferase complementation imaging (LCI) assay
-
The coding sequences of SmJAZ9 and SmMYC2 were cloned into the the cLUC vector, while the coding sequences of the SmMYB113, SmMYC2, and SmCOI1 were cloned into the nLUC vector. The recombinant constructs were transferred to Agrobacterium GV3101. The empty cLUC and nLUC vectors were used as a negative control. Agrobacterium cultures harboring cLUC-genes and cLUC-genes were co-infiltrated into 4-week-old N. benthamiana leaves. The luminescence intensity was using a vivo plant imaging system (IVIS, Lumina II), and the firefly luciferase and renilla luciferase activities were tested by the Luciferase® Reporter Assay System (E1910, Promega, USA).
Dual-luciferase assay
-
The promoter fragments of SmCHS and SmDFR were cloned into pGreenII 0800-LUC vector, while the coding sequences of SmJAZ9, SmMYB113, and SmMYC2 was cloned into the pHB overexpression vector. All recombinant constructs were transferred to Agrobacterium GV3101. Agrobacterium cultures harboring LUC-prompters and pHB-genes were co-infiltrated into 4-week-old N. benthamiana leaves. The luminescence intensity was using a vivo plant imaging system (IVIS, Lumina II), and the firefly luciferase and renilla luciferase activities were tested by the Luciferase® Reporter Assay System (E1910, Promega, USA). All the primers used in this study are listed in Supplementary Table S1.
Statistical analysis
-
The statistical package SPSS 17.0 was utilized to perform an analysis of variance (ANOVA) coupled with Duncan's New Multiple Range test for significance analysis (p < 0.05).
-
A total of 16 SmJAZ genes were identified in eggplant based on the blastp results using Arabidopsis AtJAZ as queries, and the conserved TIFY and Jas domains of JAZ protein. The gene IDs and their corresponding names are provided in Supplementary Table S2. To classify the SmJAZ proteins, a phylogenetic tree was generated using JAZ protein sequences from Arabidopsis, tomato, and eggplant, based on sequence homology (Fig. 1). The JAZ proteins were categorized into five distinct subgroups. SmJAZ1, SmJAZ2, SmJAZ5, and SmJAZ6 were divided into one group, with 2−5 exons and 136−305 aa. SmJAZ3, SmJAZ4, and SmJAZ9 were divided into one group, with seven exons and 227−399 aa. SmTIFY4A and SmTIFY4B were divided into one group, with 8−9 exons and 349−374 aa. SmJAZ7.1, SmJAZ7.2, and SmJAZ8 were divided into one group, with three exons and 105−127 aa. SmTIFY1, SmTIFY2A, SmTIFY2B, and SmTIFY8 were divided into one group, with 6−15 exons and 326−607 aa (Fig. 1).
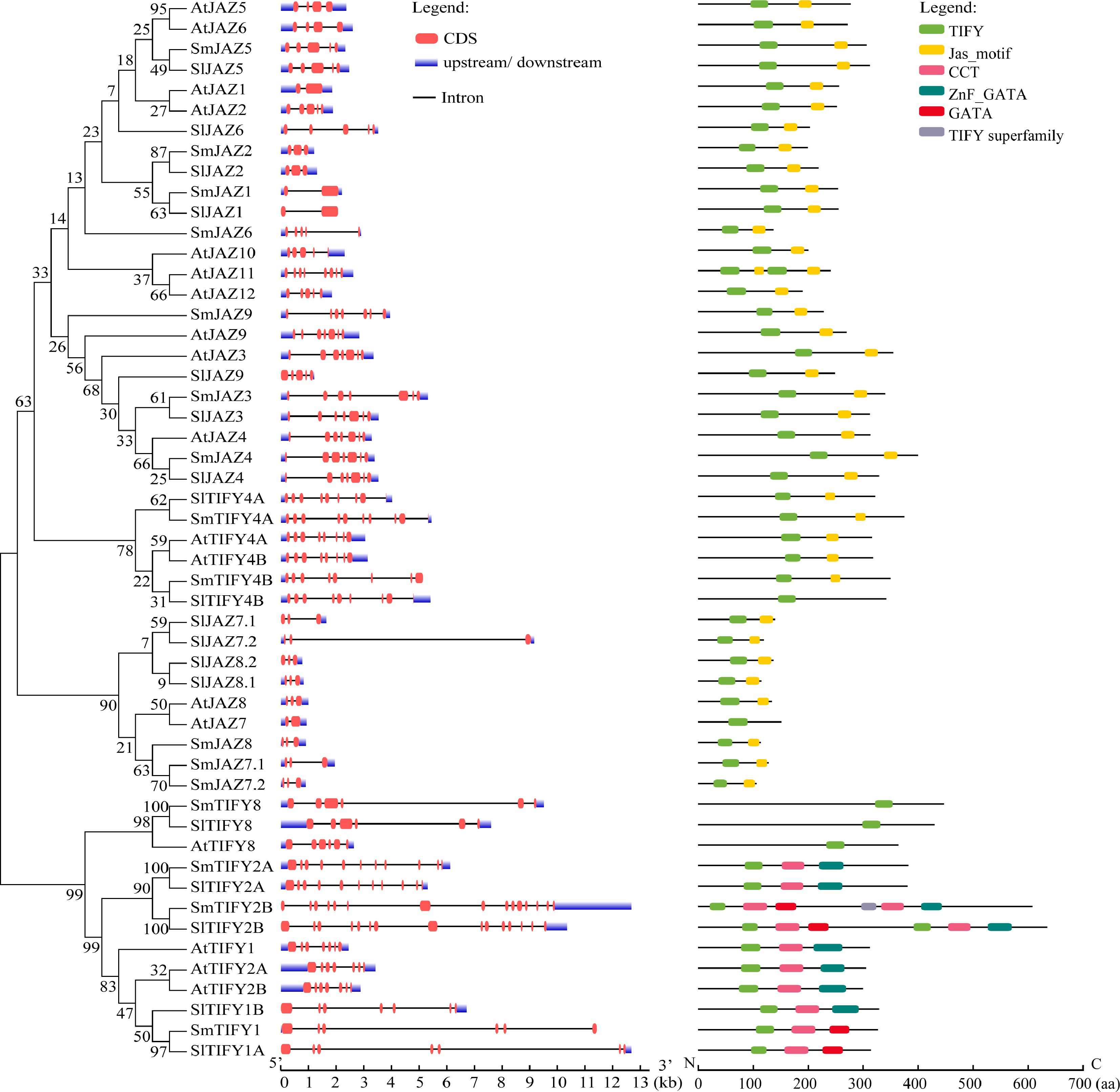
Figure 1.
Phylogenic (left), gene structure (middle), and protein conserved domain (right) analysis of JAZ proteins from eggplant, tomato, and Arabidopsis.
Chromosome distribution and evolutionary relationships of SmJAZ genes in eggplant
-
The 16 SmJAZ genes were distributed in nine chromosomes, with none genes found in chromosomes 2, 5, and 10 (Fig. 2a). To elucidate whether an intraspecific duplication event exists for the SmJAZ genes in eggplant and its evolutionary relationships among species, we performed comparative analyses within eggplant as well as with interspecific homologs from tomato, and Arabidopsis. In the eggplant genome, gene duplication events were found between SmJAZ1 and SmJAZ2, SmTIFY24A and SmTIFY4B, respectively (Fig. 2a). The results of syntenic analysis indicated that JAZ genes exhibited a stronger synteny among the three plant species. Seven SmJAZs were found collinearity in both Arabidopsis and tomato, and another five SmJAZs were found collinearity only in tomato (Fig. 2b), implying conserved protein function in terms of species evolution. However, in SmTIFY2A, SmJAZ7.1, SmJAZ7.2, and SmJAZ8 collinearity could not be found in both Arabidopsis and tomato, which may have a special function in eggplant.
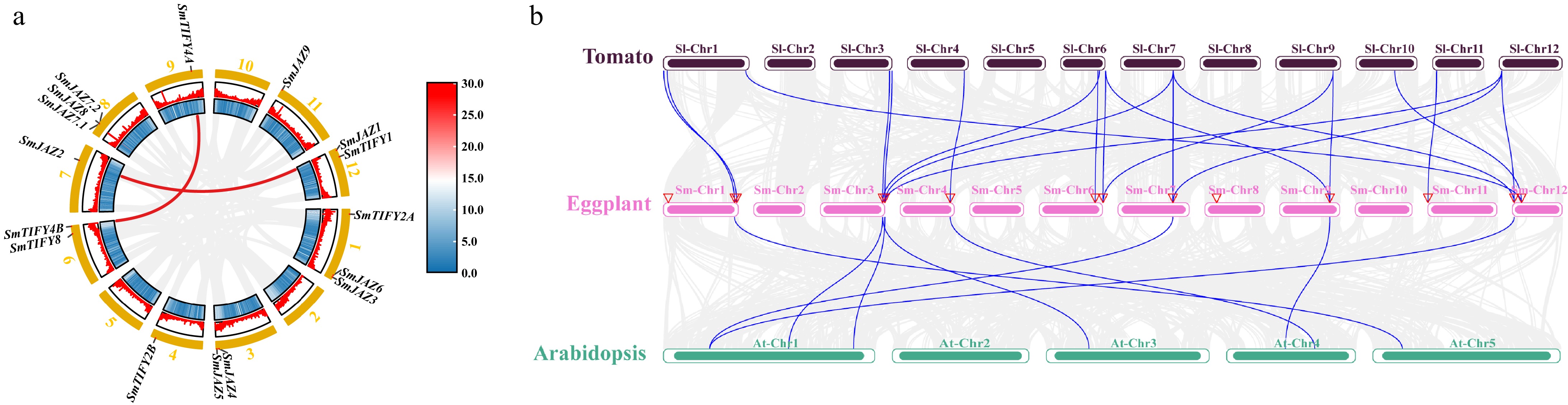
Figure 2.
The chromosome location, segmental duplication, and synteny analysis of SmJAZ genes. (a) Chromosome distribution and segmental duplication of the 16 SmJAZ genes in 12 chromosomes. Genes linked with a line are pairs of segmentally duplicated genes. (b) Syntenic relationships of JAZ genes among eggplant, tomato, and Arabidopsis. The JAZ gene pairs between different species are highlighted with different colored lines.
Correlationship analysis of the expression patterns of SmJAZ genes and anthocyanin biosynthesis-related genes in eggplant peel in response to high temperature and darkness
-
Both high-temperature (HT) and darkness treatments significantly reduced anthocyanin accumulation in eggplant fruit peels (Fig. 3a, b). The anthocyanin content was decreased by 28.7% and 72.3% under HT treatments for 7 and 14 d (Fig. 3c). Under darkness, none anthocyanins were biosynthesized (Fig. 3d). RNA-seq data further demonstrated that the expression levels of anthocyanin biosynthesis structural genes (SmCHS, SmCHI, SmF3H, SmF3'5'H, SmDFR, SmANS, Sm3GT, and Sm5GT) and regulatory genes (SmMYB113 and SmTT8) were suppressed under both stress conditions (Fig. 3e). Notably, darkness treatment resulted in significant transcriptional repression, with all the anthocyanin biosynthesis related genes exhibiting significant downregulation (|log2(fold change)| > 1, p-value < 0.05). However, high-temperature treatment resulted in moderate transcriptional repression with the log2(fold change) value ranging from −0.34 to −1.9 (p-value < 0.05). Correlation analysis between the expression patterns of SmJAZ genes and anthocyanin biosynthesis-related genes identified that SmJAZ9 exhibited strong co-expression patterns with all the anthocyanin biosynthesis-related genes under both stress conditions (Fig. 3e). Further screening based on differential expression thresholds (|log2(fold change)| > 1, p-value < 0.05), SmJAZ9 were identified as differentially expression genes (DEG). Therefore, we speculated that SmJAZ9 played an important role in both light and high-temperature regulating anthocyanin biosynthesis in eggplant peel.
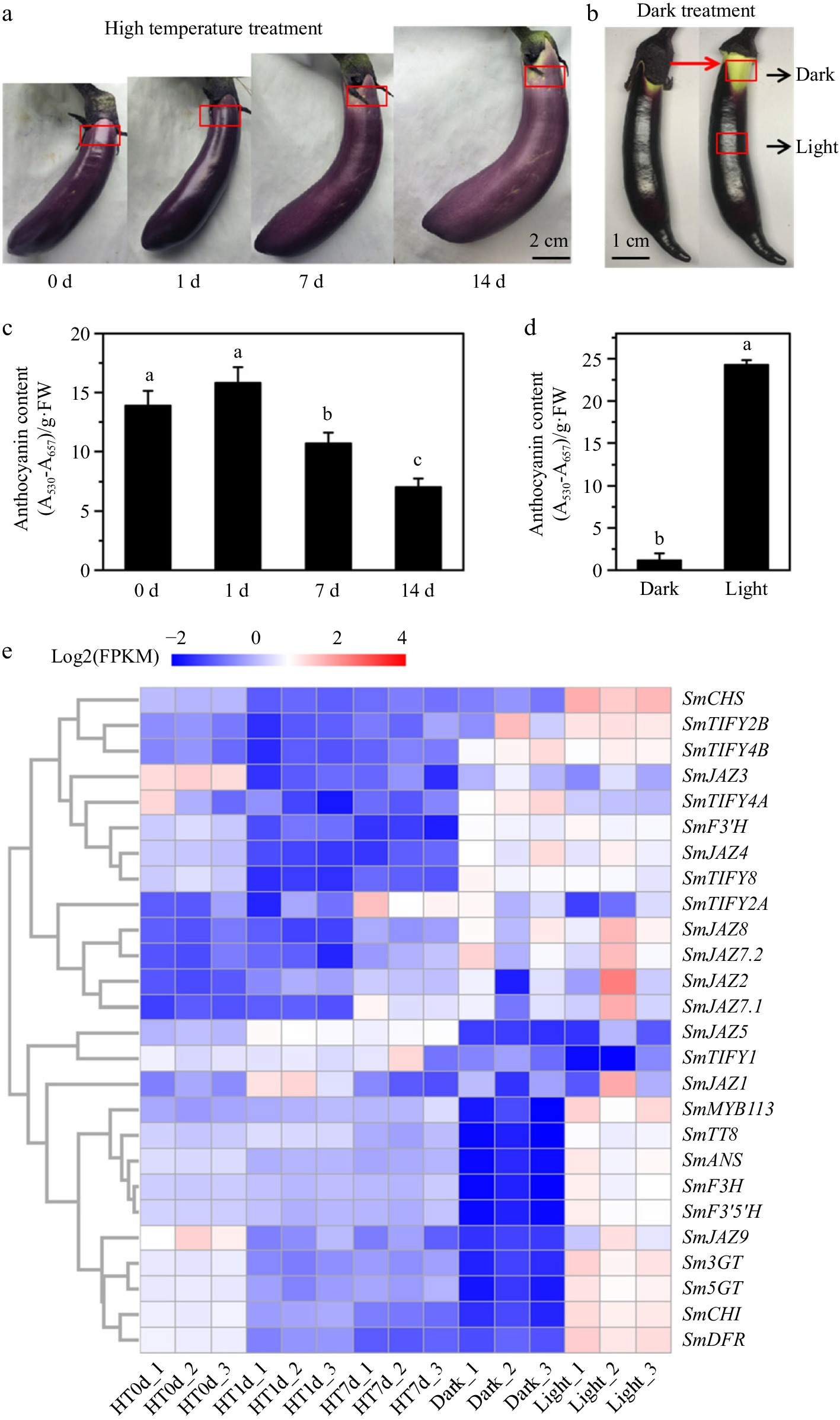
Figure 3.
Effects of high-temperature (HT), and darkness treatments on the colour, anthocyanin content, and gene expression level in the eggplant peel. (a) The peel color of eggplants after high-temperature treatment for 0, 1, 7, and 14 d. (b) The peel color of eggplant under the calyx region shielded from light, and the middle region peel of fruit exposed to light. (c), (d) The anthocyanin contents in the eggplant peel marked with a red rectangle. Data are means ± SD (n = 3 biological replicates). Different letters indicate significant differences (p < 0.05). (e) Hierarchical clustering analysis of SmJAZ genes and anthocyanin biosynthesis-related genes based on the RNA-seq data. Each row represents the log2 (FPKM) of a gene.
SmJAZ9 acts as a nuclear-localized positive regulator in anthocyanin biosynthesis
-
Subcellular localization analysis found that SmJAZ9 is localized in the nucleus, which showed clear nuclear fluorescence signals co-localizing with the nuclear marker gene (Fig. 4a). Functional validation via transient overexpression and silencing assays in eggplant peels revealed a direct regulatory role of SmJAZ9 in anthocyanin biosynthesis. Overexpression of SmJAZ9 significantly enhanced the peel color, with anthocyanin content increasing 1.6 fold compared to the empty vector control (Fig. 4b, c). Conversely, silencing SmJAZ9 using virus-induced gene silencing (VIGS) resulted in visibly lighter peel coloration and a 47.7% reduction in anthocyanin accumulation (Fig. 4b, d).
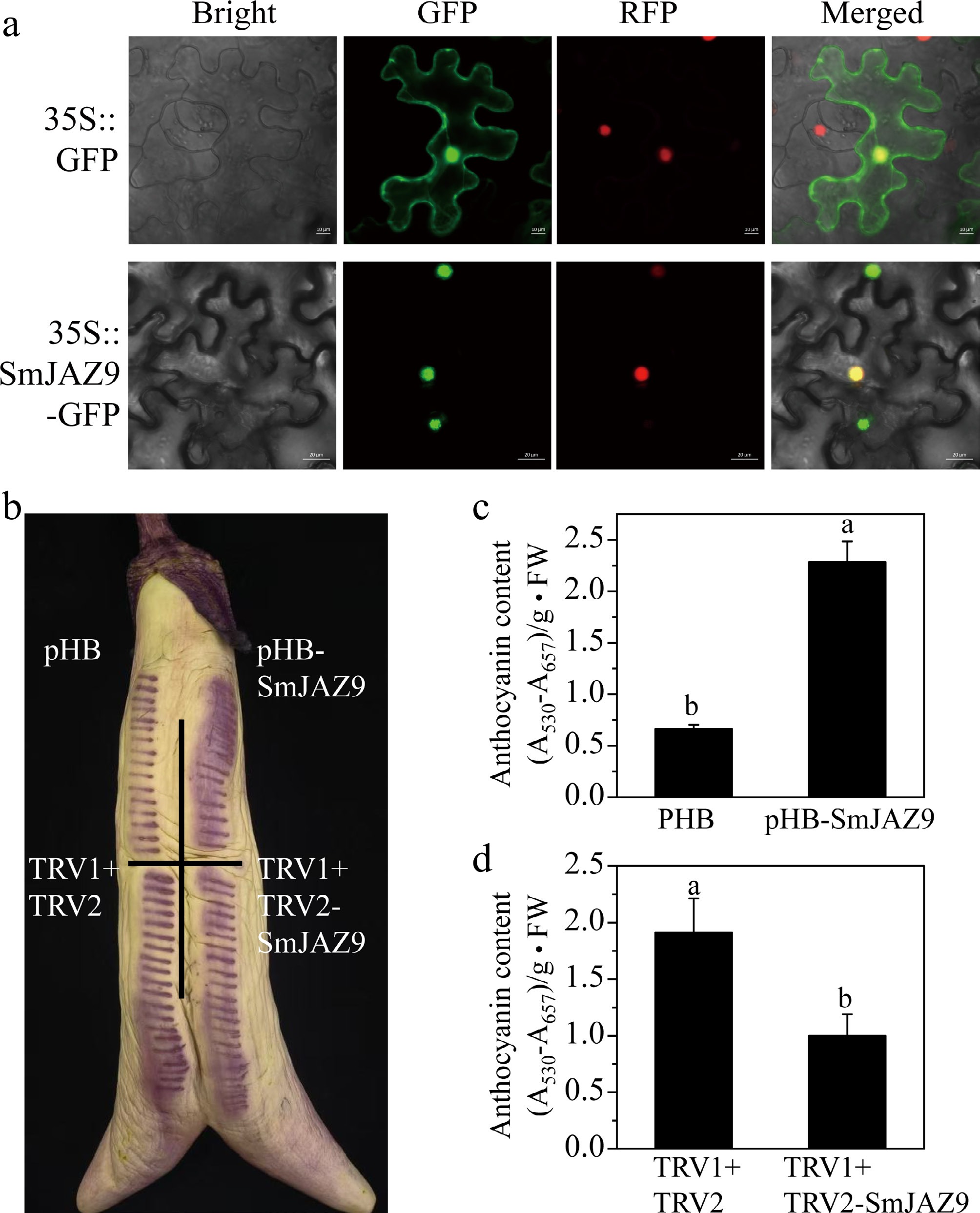
Figure 4.
Subcellular localization and functional analysis of SmJAZ9. (a) SmJAZ9 protein was located in the nucleus of tobacco cells. The 35S::GFP construct was used as the control. Bright: bright field; GFP: GFP fluorescence; RFP: RFP fluorescence. HTR12 (AT1G01370) was used as a nuclear-localized marker protein[36]; Merged: merged overlay of bright, GFP, and RFP. (b) Transient overexpression and RNAi assays in eggplant peel. (c), (d) The relative anthocyanin content in the infiltrated eggplant peel. Data are means ± SD (n = 3 biological replicates). Different letters indicate significant differences (p < 0.05).
SmJAZ9 interacted with SmMYB113 and promoted the transcriptional activity of SmMYB113 on the promoters of SmCHS and SmDFR
-
Given the absence of transcriptional activation activity in JAZ proteins, we hypothesized that SmJAZ9 might regulate anthocyanin biosynthesis through protein-protein interactions with transcriptional regulators (SmMYB113 and SmTT8). Yeast two-hybrid (Y2H) assays demonstrated that SmJAZ9 could interact with SmMYB113, while no interaction was detected with SmTT8 (Fig. 5a). This interaction was further validated in vivo using luciferase complementation (LCI), and bimolecular fluorescence complementation (BiFC) assays (Fig. 5b, c). Significant luciferase activity was detected for the SmMYB113-SmJAZ9 pair, and distinct fluorescent signals were observed in nuclei co-expressing pY2N-SmMYB113 and pY2C-SmJAZ9 (Fig. 5c), confirming their direct interaction.
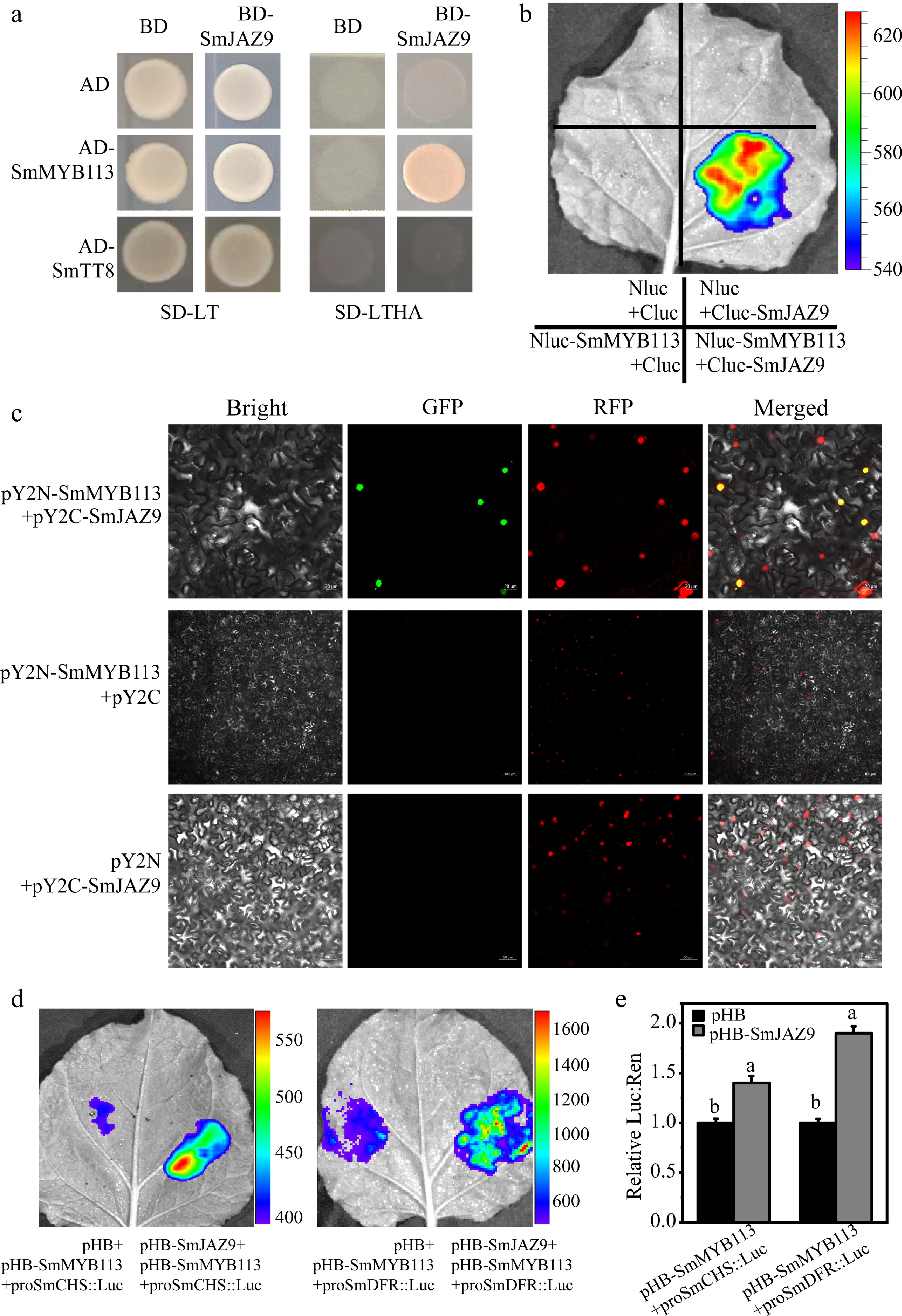
Figure 5.
SmJAZ9 interacted with SmMYB113 and enhanced the transcriptional activity of SmCHS and SmDFR promoters. (a) SmJAZ9 interacts with SmMYB113 but not SmTT8 in Y2H assays. SD-LT, SD medium without Trp/Leu; SD-LTHA, SD medium without Trp/Leu/His/Ade. (b) LCI assays results confirmed the interaction between SmJAZ9 and SmMYB113. (c) SmJAZ9 interacts with SmMYB113 in BiFC assays. (d), (e) Dual-luciferase assay analyze the activation of SmCHS and SmDFR promoters co-transfection with both SmJAZ9 and SmMYB113 or SmMYB113 only. Data are means ± SD (n = 3 biological replicates). Different letters indicate significant differences (p < 0.05).
To investigate the combined effects of SmMYB113-SmJAZ9 in activating the expression of SmCHS and SmDFR, which are the target genes of SmMYB113[37], dual-luciferase reporter assays were performed (Fig. 5d, e). The luminescence intensity and LUC activity in the site co-transfection of proSmCHS::LUC/proSmDFR::LUC with both SmJAZ9 and SmMYB113 was significantly enhanced comparing with that co-transfection of proSmCHS::LUC/proSmDFR::LUC only with SmMYB113. These results suggested that SmJAZ9 improved the anthocyanin biosynthesis through interacting with SmMYB113, which further promotes the transcriptional activity of SmCHS and SmDFR promoters.
SmJAZ9 interacted with SmMYC2, but not with SmCOI1
-
SmJAZ proteins are well known as negative regulators in the JA signal transduction pathway, which could interact with MYC2 transcription factors in the absence of JA and dynamically interact with COI1 upon JA perception. Phylogenetic analysis identified eggplant homologs of MYC2 and COI1 (Supplementary Fig. S1), respectively. Domain architecture analysis confirmed conserved functional motifs in both proteins: SmMYC2 retains the characteristic JID and bHLH domain required for DNA binding, while SmCOI1 preserves the F-box and leucine-rich repeat (LRR) domains critical for SCF complex assembly (Supplementary Figs S2 & S3).
Yeast two-hybrid (Y2H) assays revealed a direct interaction between SmJAZ9 and SmMYC2, as evidenced by growth on SD-LTHA medium. In contrast, no interaction was detected between SmJAZ9 and SmCOI1 (Fig. 6a). This interaction specificity was corroborated in planta using LCI and BiFC. Co-expression of pY2N-SmMYC2 and pY2C-SmJAZ9 in tobacco leaves produced strong nuclear fluorescence signals (Fig. 6b), whereas no signal was observed for SmJAZ9-SmCOI1 combinations. LCI assays showed significant luciferase activity when SmJAZ9-SmMYC2 pairs co-expressed, but no activity for negative controls (Fig. 6c). Based on a previous study[38], we found that the JAZ proteins that interact with COI1 contain a critical threonine (T), whereas the threonine (T) of SmJAZ9 has changed to a serine (S), which may be the result of SmJAZ9 does not interact with SmCOI1(Supplementary Fig. S4).
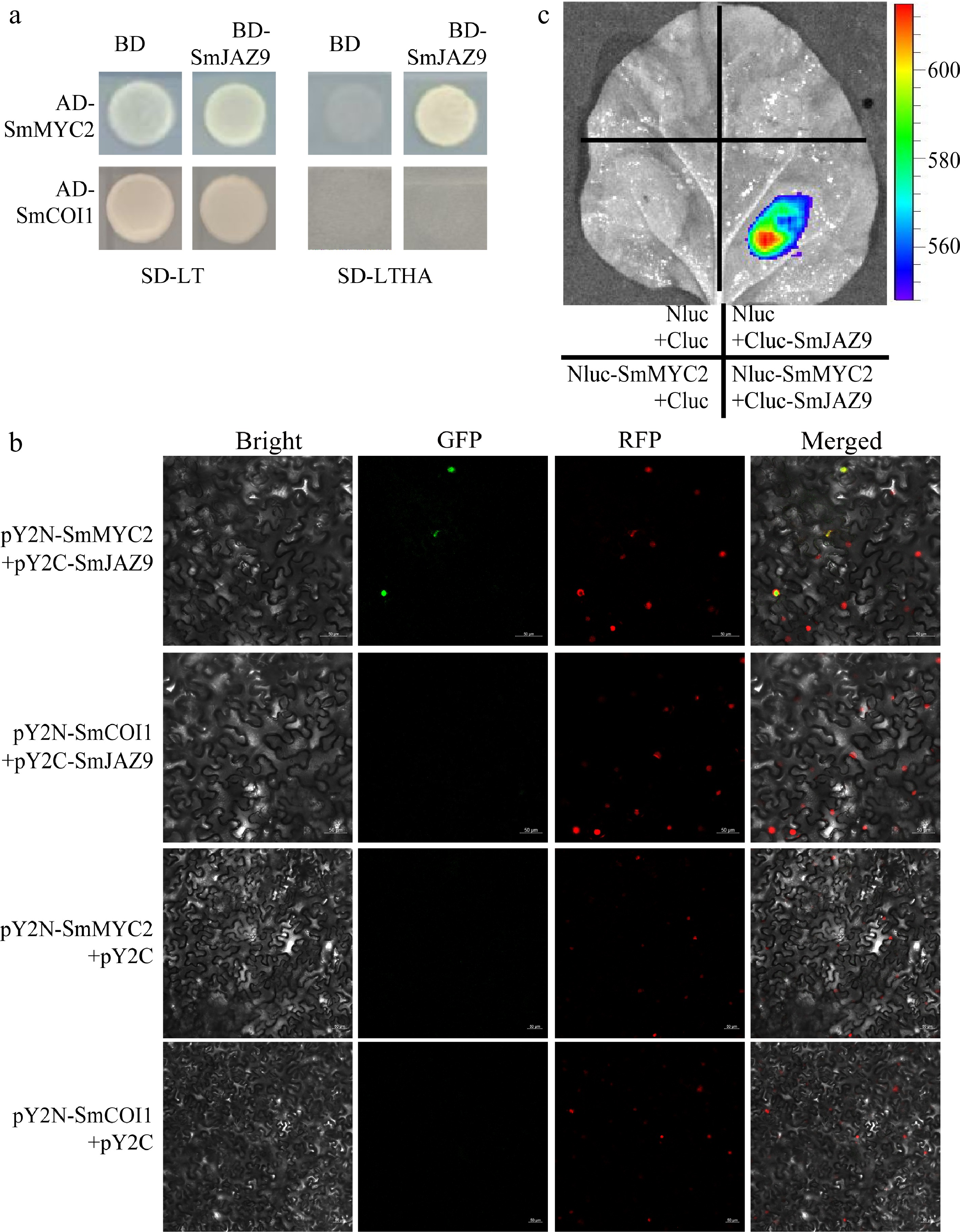
Figure 6.
SmJAZ9 interacted with SmMYC2 but not with SmCOI1. (a) SmJAZ9 interacts with SmMYC2 but not SmCOI1 in Y2H assays. (b) SmJAZ9 interacts with SmMYC2 but not SmCOI1 in BiFC assays. (c) LCI assays results confirmed the interaction between SmJAZ9 and SmMYC2.
-
Anthocyanin biosynthesis in eggplant, a key determinant of fruit commercial value, is tightly regulated by environmental stressors and plant hormones. For example, high temperature and low light (or dark) could repress anthocyanin biosynthesis, while low temperature and high light could promote anthocyanin biosynthesis[39]; JA is well known as a positive hormone in regulating anthocyanin biosynthesis[15,16]. Moreover, JA was also involved in abiotic stress responses, including high temperature and low light[9,40]. Classically, JAZ proteins repress JA responses by binding MYC2 transcription factors under low JA conditions, with subsequent COI1-mediated degradation upon JA perception[18−20]. JAZ proteins play crucial roles in regulating plant growth, development, and defense responses against biotic and abiotic stresses[41−43]. However, the role of JAZ proteins on anthocyanin biosynthesis under high-temperature or dark conditions remains unexplored.
The JAZ gene family has been identified in various plant species, including tobacco[44], tomato[21], Chinese cabbage[23], and potato[24]. In this study, 16 SmJAZ genes were identified in eggplant. Phylogenetic classification into five subgroups aligns with structural divergence in exon-intron architecture and protein lengths, suggesting functional specialization during evolution (Fig. 1). Notably, Group II (SmJAZ3/4/9) exhibits relatively conserved 7-exon structures, while Group IV (SmJAZ7.1/7.2/8) displays truncated 3-exon configurations. The chromosomal distribution pattern, particularly the absence of SmJAZs in chromosomes 2/5, correlates with tomato, implying selection against gene loss in these genomic compartments (Fig. 2). Collinearity analysis and gene duplication events revealed genetic divergence of SmJAZ genes during evolution, with eggplant showing a close phylogenetic relationship to tomato and Arabidopsis. HT stress significantly affected the expression level of SmJAZ1, SmJAZ7.1, SmJAZ7.2, and SmJAZ9, while only SmJAZ9 expression level was significantly changed by darkness stress (Fig. 2). Additionally, the anthocyanin content and the expression level of anthocyanin biosynthesis related genes were significantly repressed by both HT and dark stresses. Correlationship analysis also revealed that SmJAZ9 expression strongly correlated with anthocyanin biosynthesis-related genes under HT and darkness conditions (Fig. 3). Therefore, it can be speculated that SmJAZ9 may respond to HT and darkness stress, and participating in the regulation of anthocyanin accumulation in eggplant fruit peel.
The JAZ genes are generally considered negative regulators of anthocyanin biosynthesis. For example, CmJAZ7 can interact with CmMYB6/7 to negatively regulate anthocyanin biosynthesis in chrysanthemums[45]; MdJAZ2 interacts with MdHIR protein to inhibit anthocyanin biosynthesis in apple[46]; Anthocyanin accumulation after the JAZ4 mutation indicates that JAZ4 negatively regulates anthocyanin synthesis in Arabidopsis[47]. However, transient expression assays in eggplant peel demonstrated that SmJAZ9 positively regulates anthocyanin accumulation (Fig. 4), which is a novel function of JAZ proteins in anthocyanin biosynthesis. SmMYB113 and SmTT8 have been previously demonstrated as key regulators promoting anthocyanin biosynthesis[1,48,49]. Existing studies have established that MYB and bHLH transcription factors can form protein complexes with JAZ family genes[16,50,51]. Our results suggested that SmJAZ9 could interact with SmMYB113, while no interaction was detected with SmTT8 (Fig. 5). Previous research has shown that SmMYB113 could directly bind and activate the promoters of key anthocyanin biosynthetic genes, SmCHS and SmDFR[37,52]. In this study, we found that the SmJAZ9-SmMYB113 interaction enhances the expression of both SmDFR and SmCHS.
Different from other JAZ proteins, SmJAZ9 acts as a positive regulator in anthocyanin biosynthesis. To further analyze in depth the reasons why SmJAZ9 functions differently from other JAZs, we explored the interaction relationship between SmJAZ9 and SmMYC2/SmCOI1. However, unlike canonical JA-COI1-JAZ-MYC2 signaling, SmJAZ9 exhibited unconventional behavior: it interacted with SmMYC2 but failed to associate with SmCOI1 (Fig. 6). These results suggest that the degradation of SmJAZ9 may be decoupled from the perception of JA, thereby favoring alternative regulatory pathways. In other plants, some of the JAZ proteins could not interact with COI1. For example, only six ZmJAZ proteins were found to interact with ZmCOI1a, while the remaining 13 ZmJAZ proteins could not interact with ZmCOI1a in maize[53]; Eight of the 12 SlJAZ proteins could not interact with SlCOI1 in tomato[54]; Seven of the 11 GpJAZ proteins could not interact with GpCOI1 in Gynostemma pentaphyllum[30].
-
Our study identified 16 SmJAZ genes in eggplant, which were classified into five subfamilies with conserved domains and exons. Collinearity analysis revealed a close evolutionary relationship between the JAZ genes of eggplant and tomato. Correlationship analysis of RNA-seq data demonstrated that SmJAZ9 both responds to HT and darkness stresses and participates in the regulation of anthocyanin accumulation in eggplant fruit peel. Furthermore, we found that SmJAZ9 could interact with SmMYB113, further promoting the transcriptional activity of SmCHS and SmDFR promoters. Additionally, the degradation of SmJAZ9 may be decoupled from the perception of JA based on the absence of SmJAZ9 and SmCOI1 interaction.
This work was supported by the National Key Research and Development Program (2023YFD2300702), the National Natural Sciences Foundations of China (Grant No. 32172556), the Shandong Provincial Key Research and Development Program (2021LZGC018), the Special Fund of Vegetable Industrial Techonolgy System of Shandong Province (SDAIT-05-11), the Tai'an Agricultural Breeding Program (2023NYLZ02), and the Grand Science and Technology Special Project of Zhejiang Province (2021C02065).
-
The authors confirm contribution to the paper as follows: study conception and design: Li J, Yang F, Ji T, Gu D; data collection: Li J, Wang Y, Wang H, Xin Y, Yuan M; analysis and interpretation of results: Li J, Wang Y; manuscript preparation: Li J, Wang Y; manuscript review and editing: Li J, Wang Y; manuscript revision and data re-analysis: Li J, Yang F. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files. The raw data of the sequences used during the current study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 All the primers used in the study.
- Supplementary Table S2 The gene ID of SmJAZ genes in egplant genome database.
- Supplementary Fig. S1 The phylogenetic relationships of the MYC2 (a) and COI1 (b) proteins among different plant species.
- Supplementary Fig. S2 Alignment of SmMYC2 with the COI1 proteins from the other plants.
- Supplementary Fig. S3 Amino acid sequence comparison of SmCOI1 with the COI1 proteins from the other plants.
- Supplementary Fig. S4 Amino acid sequence comparison of SmJAZ9 with the other JAZs in the same clade.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Y, Wang H, Xin Y, Yuan M, Ji T, et al. 2025. Genome-wide identification of SmJAZ gene family in eggplant and functional mechanism analysis of SmJAZ9 modulating high temperature and darkness stress-regulated anthocyanin biosynthesis. Vegetable Research 5: e018 doi: 10.48130/vegres-0025-0019 

Genome-wide identification of SmJAZ gene family in eggplant and functional mechanism analysis of SmJAZ9 modulating high temperature and darkness stress-regulated anthocyanin biosynthesis
- Received: 16 April 2025
- Revised: 20 May 2025
- Accepted: 23 May 2025
- Published online: 09 June 2025
Abstract: The biosynthesis of anthocyanin in plants is regulated by temperature, light, jasmonic acid (JA), among others. JASMONATE ZIM-DOMAIN (JAZ) proteins are critical repressors of the jasmonic acid (JA) pathway, the role of JAZ proteins on anthocyanin biosynthesis under high-temperature or dark conditions remains unexplored. Here, we identified 16 SmJAZ genes in eggplant and characterized their evolutionary relationships, gene structures, and high-temperature (HT) and darkness stress-responsive expression patterns. Both the HT and darkness treatments significantly inhibit anthocyanin biosynthesis in eggplant peel. Correlation analysis revealed that the SmJAZ9 expression strongly correlated with anthocyanin biosynthesis-related genes under HT and darkness conditions. Transient expression assay in eggplant peels demonstrated that nuclear-localized SmJAZ9 is a positive regulator, enhancing anthocyanin biosynthesis via interaction with the transcription factor SmMYB113, thereby promoting SmCHS and SmDFR transcription. Additionally, SmJAZ9 interacted with SmMYC2 but not SmCOI1, suggesting a JA signaling-independent regulatory mechanism under HT and darkness conditions. This study revealed a novel JAZ-mediated regulatory module integrating JA signaling, environmental stress responses, and anthocyanin biosynthesis, providing potential targets for improving eggplant fruit quality.
-
Key words:
- Eggplant /
- Anthocyanin /
- JAZ proteins /
- High-temperature /
- Darkness