









-
Grape (Vitis. spp) is one of the most appreciated and extensively planted fruit crops, boosting economic significance and nutritional benefits. The growth, development, and ripening process of grape berry involve a complex interplay of biochemical and physiological events, following a typical biphasic sigmoidal profile that encompasses three different phases[1]. Phytohormones intricately regulate grape berry development and ripening. Notably, abscisic acid (ABA), ethylene, and BRs emerge as pivotal promoters in the ripening process, while auxin, cytokinin, and gibberellin are proposed as ripening inhibitors[1,2]. In grape, the level of endogenous BRs increased markedly at the initiation stage of berry ripening. Conversely, treatments with brassinazole (Brz), a BR synthesis inhibitor, significantly delay berry véraison and ripening process[3], suggesting a crucial role of BRs in grape berry ripening process.
BRs represent natural steroid-like hormones in the plant kingdom that play fundamental roles in various processes of plant growth and development, such as cell division and elongation, plant architecture, vascular differentiation, photomorphogenesis, flowering, senescence, tolerance to biotic and abiotic stresses[4−6]. More than seventy different BR compounds and their derivatives have been verified from the plantae, of which brassinolide (BL) and castasterone (CS) are emerging as the most actively and prevalently class of BR compounds[6,7]. BL and CS arise from the conversion of campesterol (CR), a prominent plant sterol, through two parallel routes known as C-6 oxidation pathways at the early and late phases, respectively[6,8]. A cytochrome P450 monooxygenase, denoted as DWF4, plays a pivotal role in this biosynthetic cascade by catalyzing multiple C-22 hydroxylation steps. This enzyme, recognized as a rate-limiting factor, significantly influences the BR biosynthesis route[9]. In Arabidopsis, the dwf4 mutant exhibits a dwarf phenotype, attributed to impaired BRs biosynthesis, but hydroxylated BRs can restore the defective phenotypes from the dwf4 mutant of Arabidopsis[9,10]. In autotetraploid apple, the reduction in BR levels and the dwarf phenotype are speculated to result from the significant down-regulation of MdDWF4 expression[11].
The signaling pathways of BRs are initiated through a binding of BRs with a leucine-rich repeat (LRR) receptor-like kinase known as BRASSINOSTEROID-INSENSITIVE 1 (BRI1), which operates in conjunction with its co-receptor BRI1-associated receptor kinase (BAK1)[12]. For example, the Arabidopsis bri1 mutant exhibits elevated endogenous BR levels[13] and manifests multiple developmental deficiencies, including a profound reduction in height, notably dark green, thick leaves, male sterility, and diminished apical dominance[14]. Subsequently, this signaling cascade culminates in the activation of a group of transcription factors, including BRASSINAZOLE RESISTANT 1 (BZR1)[15] and BRI1-EMSSUPPRESSOR 1 (BES1)[16]. BZR1 and BES1 not only directly regulate the transcription of numerous BR-responsive genes governing various plant growth and development processes but also contribute to feedback inhibition by binding directly to the promoter regions of BR biosynthetic-related genes, thereby inhibiting their transcription levels[17]. This regulation occurs through the modulation of light reactions, cell-wall-related metabolism, and plant hormone pathways[18].
In grapes, several previous investigations have confirmed the positive impact of BRs on the berry ripening process. The exogenous spray of epibrassinolide (EBR) not only extensively induced the accumulation of soluble sugars, anthocyanins, proanthocyanidins, and other phenolics but also promoted the enzyme activities and gene expression associated with anthocyanin and sugar biosynthetic pathways[19,20]. However, there are limited reports regarding the interrelationship between BRs-induced ripening and gene regulatory networks in grapes. Herein, this study has further confirmed the functions of BRs in promoting berry ripening and accelerating secondary metabolite accumulation. Transcriptomic analysis unveiled the regulatory networks governing berry ripening organs and the cross-talk among hormones in response to EBR treatment. The present results not only enhance understanding related to the roles of BRs during the regulation network of the ripening process in berry but also provide an excellent tool for improving the berry quality in commercial vineyards.
-
Six-year-old Muscat Hamburg grape (Vitis vinifera L.) was considered as the experimental material, which was grown on a grape farm at Qingdao Agricultural University, Qingdao, China. Berries were soaked with deionized water (control) containing a trace amount of ethanol (< 0.1%) and 0.5 mg·L−1 EBR (Sigma catalog no. E1641)[21] with 0.05% Tween 20 at one week before véraison (eight weeks after full bloom), respectively. The stock solution of EBR was prepared by dissolving its compound in 1 mL ethanol (98%), and the control stock solution only contained 1 mL ethanol (98%) but without EBR. One hundred berries were randomly harvested at 0, 5, 10, and 15 d after EBR application from three biological replicates. Thirty berries at each sampling date were selected for the physiology and biochemical indices as per the previous method[22], including longitudinal diameter (mm), transverse diameter (mm), fresh berry weight (g), content of total soluble solids (Brix°), content of titratable acidity (%). These left berries were frozen in liquid nitrogen and kept at −80 °C for the next analysis.
Determination of endogenous BRs level
-
The levels of endogenous BRs were quantified using the method described by Xu et al.[23]. One gram of each of the berry skin samples was ground and homogenized in PBS extraction solution. The extracts were then centrifuged at 12,000 rpm for 30 min, and the supernatant subsequently was collected and stored at 4 °C until enzyme-linked immunosorbent assays (ELISAs), which are manufactured by Shanghai Yanqi Biotechnology Co., Ltd (Shanghai, China).
Quantification analysis of soluble sugars and organic acids
-
The concentrations of soluble sugar (fructose and glucose) and organic acid (malic acid, tartaric acid, and citric acid) were determined by high-performance liquid chromatography (HPLC) as a previous method with some modifications[24]. Six hundred mg of berry samples were ground and extracted with 1.5 mL of 80% ethanol. These samples were vortexed and centrifuged at 12,000 rpm for 30 min, and then the extract was filtered through a 0.22 μm water filter for injection. The next analysis was manipulated by the liquid chromatography-mass spectrometry (LC-MS) system, which is a combination of Waters Acquity I-Class PLUS ultra-high performance liquid tandem Waters (Shanghai, China) and an AB Sciex Qtrap 6500+ high-sensitivity mass spectrometer. The chromatographic conditions of soluble sugars were as follows: Prevail Carbohydrate ES 5μ column (4.6 mm × 100 mm, 5 μm); mobile phase: acetonitrile/water, 80/20 (v/v); flow rate: 1.0 mL/min; column temperature: 50 °C; injection volume: 20 μL. Furthermore, the chromatographic conditions of organic acids were as follows: Discovery C18 column (4.6 mm × 250 mm, 5 μm); mobile phase: 50 mM K2HPO4 solution (pH adjusted to 2.4 with phosphoric acid); flow rate: 0.5 mL/min; column temperature: 30 °C; injection volume: 20 μL.
Quantification analysis of anthocyanin
-
Grape anthocyanins were extracted using the methanol containing 0.1% HCl (v/v). The components and contents of anthocyanin were determined by LC-MS according to the previous method[25], and the LC-MS system used has been mentioned above. The chromatographic conditions were as follows: aqueous mobile phase: 0.1% formic acid in water; organic mobile phase: 0.1% formic acid in acetonitrile; flow rate: 0.4 mL/min; injection volume: 2 μL. All mass spectrometry (MS) were equipped with turbo ion spray sources in both positive and negative ion modes, with a selected mass range of 50−1,200 m/z. The ionization parameters were set as the previous method[22]. Malvidin-3-O-glucoside (Extrasynthese, Genay, France) was used as the external standard for the quantitation of anthocyanins. Furthermore, the total anthocyanin contents of strawberry fruits were measured according to the pH differential method.
Measurement of fruit firmness and cell wall components
-
Firmness was measured by using a Texture Analyzer (TA. XT, Stable Micro Systems, Surrey, UK) with a 2 mm diameter penetration probe (needle P/2). The penetration test of the whole grape berry (with skin) was performed at 1 mm/s for 6 mm after contacting the surface of the flesh, and the results were expressed in N. Protopectin, water-soluble pectin (WSP) and cellulose contents were measured using the test kits (Suzhou Comin Biotechnology Co., Ltd. Suzhou, China) according to the manufacturer's instructions.
RNA extraction, cDNA library construction and sequencing
-
The CTAB method was used to total RNA isolation from berry skins. RNA integrity and concentration were measured by using a Bioanalyzer 2100 system (Agilent Technologies, CA, USA) and a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, CA, USA), respectively. Then, 3 μg total RNA was used to prepare a cDNA library using the NEBNext® Ultra™ RNA Library Prep Kit for Illumina® (NEB, MA, USA). All the library preparations were sequenced by an Illumina Hiseq platform (Hiseq 4000, 150PE) at Novogene Bioinformatics Technology Co. Ltd (Beijing, China).
Transcriptome analysis
-
Clean reads were aligned to the grape reference genome (
www.grapegenomics.com/pages/PN40024 ) by HISAT 2.0. The gene expression level was presented by using the fragments per kilobase of exon model per million mapped fragments (FPKM) values. Differentially expressed genes (DEGs) between BR-treated and control samples (two biological replicates per sample) were identified by the DESeq 1.8.3 package. The resulting p values were adjusted using the Benjamini-Hochberg approach for controlling the false discovery rate (FDR). The DEGs were collected by the |log2(fold change)| ≥ 1 and FDR < 0.05 as select standard. Furthermore, GO enrichment analysis of DEGs was conducted using the GOseq R package, and KEGG pathway analysis was implemented using KOBAS software. The function and biological pathway with a q-value ≤ 0.05 were considered significantly enriched.Quantitative RT-PCR validation
-
Twelve DEGs were chosen to validate the results of transcriptome sequencing by using qRT-PCR. Total RNA was extracted by the CTAB method from every single sample, and 1 μg of total RNA was used for cDNA synthesis by using PrimeScript™ RTase (TaKaRa Biotechnology, Dalian, China) according to the manufacturer's instruction. qRT-PCR was implemented by using SYBR®Premix Ex Taq™ (TaKaRa Biotechnology, Dalian, China) with the Bio-Rad iQ5 thermocycler (Bio-Rad, CA, USA). The grape Actin gene (AY680701) was used as the reference gene for normalization, and all primers pairs were shown in Supplementary Table S1. All experiments were implemented with three independent biological replicates for every sample. The relative expression values were calculated by the 2−ΔΔCᴛ method.
Transient overexpression of VvDWF4 using the strawberry fruit
-
The full-length coding sequence (CDS) encoding VvDWF4 (VIT_04s0023g01630) was cloned into a pRI101 binary vector to generate the plant expression constructs. The construct was introduced into the Agrobacterium tumefaciens strain GV3101 and then transformed into the fruits (as previously described[26]) of strawberry (Fragaria × ananassa Duch. L. cv. Albion) plants which were grown under the greenhouse at 25 °C, approximately 70% relatively humidity. Strawberry fruit infiltrated with an empty PRI101 vector was used as the control. The primers for construction were listed in Supplementary Table S1.
Statistical analysis
-
Statistical analysis was carried out with SPSS software ver. 22.0 (IBM, NY, USA). Data were expressed as means ± standard deviation (SD). Comparisons between groups were carried out by one-way ANOVA corrected with Tukey's multiple comparison test at a significance level of p < 0.05.
-
The exogenous BRs treatment remarkably facilitated ripening process of the berries, encompassing berry coloration, sugar accumulation, and organic acid degradation (Fig. 1). Within 15 d of BRs treatment, the berry clusters exhibited full coloration, whereas the control groups showed significantly poor color development compared to the BRs-treated berries (Fig. 1a). To further validate the correlation between endogenous BRs content and berry ripening, the BRs concentrations were detected. The results displayed that the content in BRs was obviously increased at 5 and 10 d after treatment. However, there is no significant difference found at 15 d between the control and treated fruit, suggesting BRs might be a potent inducer and started the cascade of events leading to ripening (Fig. 1b).
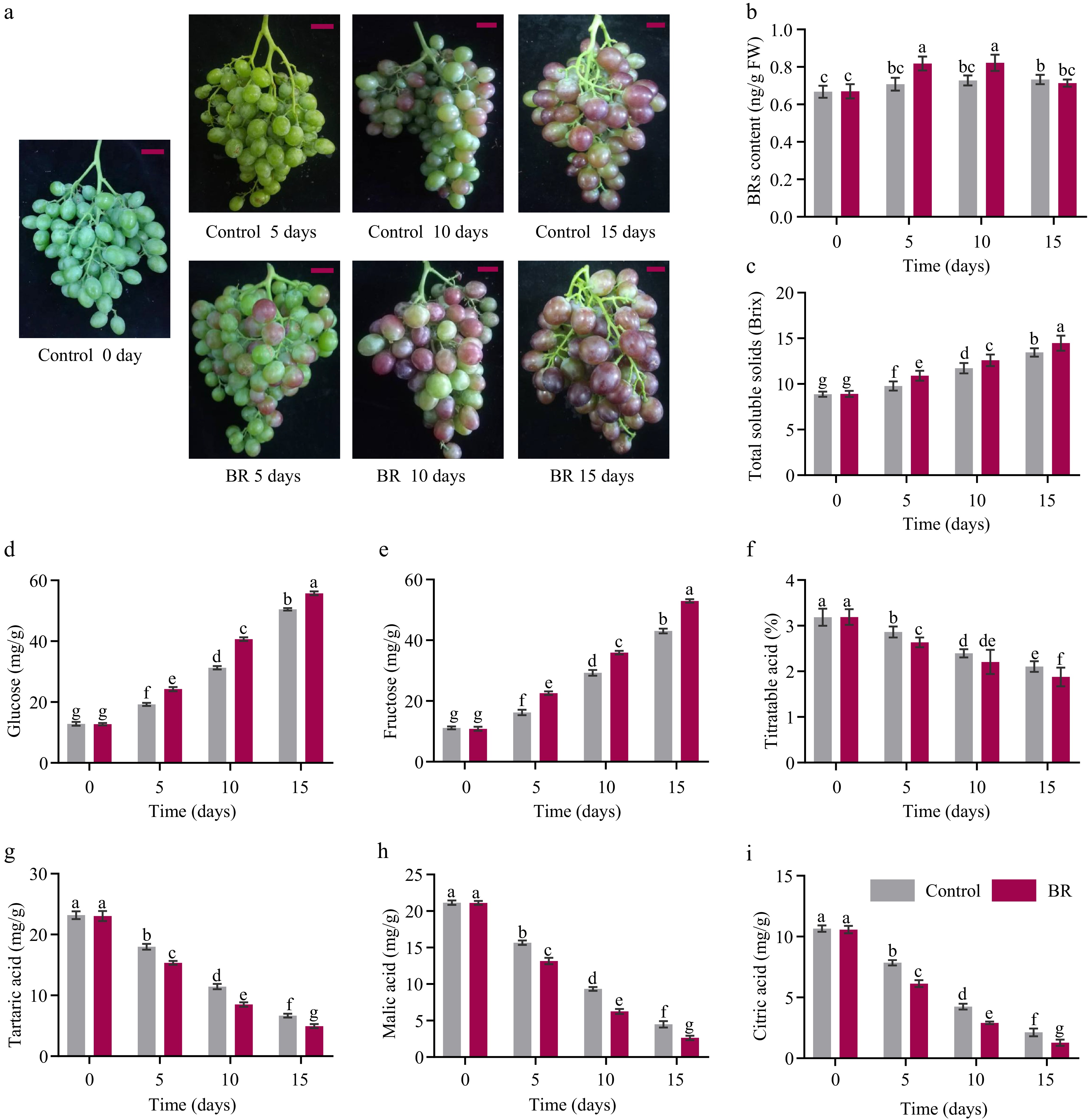
Figure 1.
The effect of exogenous 2,4-epibrassinolide (EBR) treatment on the berry ripening indexes of grape 'Muscat Hamburg'. (a) Color development and anthocyanin accumulation phenotypes under EBR treatment; The red bar indicates 5 cm, (b) BRs content, (c) total soluble solids, (d) glucose, (e) fructose, (f) titratable acid, (g) tartaric acid, (h) malic acid, and (i) citric acid were qualified post-EBR application. Data are expressed as the means ± standard deviation. Different letters indicate significant differences at p < 0.05 according to one-way ANOVA and Tukey's test.
Furthermore, BRs application significantly accelerated the accumulation of soluble solids, glucose and fructose (Fig. 1c−e), indicating the participation of BRs onto sugar accumulation. During the berry ripening process, the content of titratable acids in BRs-treated berries, including tartaric, malic, and citric acid, exhibited a significant reduction in comparison to the control group (Fig. 1f−i). All these results suggested that the appropriate level of epibrassinolide (EBR) treatment is an effective strategy for promoting the grape berry ripening process.
BRs application promotes anthocyanin accumulation in grape
-
Anthocyanin composition and content were measured through the LC-MS system, leading to the identification and quantification of seven distinct anthocyanin compounds[27], including peonidin-3-O-glucoside (Pn-3-G), malvidin-3-O-glucoside (M-3-G), cyanidin-3-O-glucoside (C-3-g), petunidin-3-O-glucoside (Pt-3-G) and delphinidin-3-O-glucoside (D-3-G), peonidin-3-O-(6-p-coumaroyl)-glucoside (Pn-3-coum-G) and malvidin-3-O-(6-p-coumaroyl)-glucoside (M-3-coum-G). (Fig. 2). Application of exogenous BR resulted in a significant increase in total anthocyanins contents, spanning from 85.47 to 179.43 mg/kg and 274.91 to 385.44 mg/kg at 10 and 15 d post-BR treatment, respectively (Fig. 2h). Furthermore, the concentration of individual anthocyanins was also significantly enhanced due to BRs treatment. Among these, peonidin-3-O-glucoside (Pn-3-G), a 3'-substituted anthocyanin, stood as the highly abundant compound within the individual anthocyanin constituents of Muscat Hamburg, accounting for around 50.0% of the total anthocyanins level (Fig. 2a). Malvidin-3-O-glucoside (M-3-G), the key 3',5'-substituted anthocyanins, exhibited concentrations ranging from 19.61 to 31.83 mg/kg and 52.63 to 77.55 mg/kg at 10 and 15 d post-BR application, respectively (Fig. 2c). In addition, the contents of two p-coumaroyl anthocyanins, namely Pn-3-coum-G and M-3-coum-G, was notably lower than monoglucosidic anthocyanins. These findings robustly underscore the capacity of exogenous EBR application to trigger anthocyanin accumulation and promote berry coloration in grape.
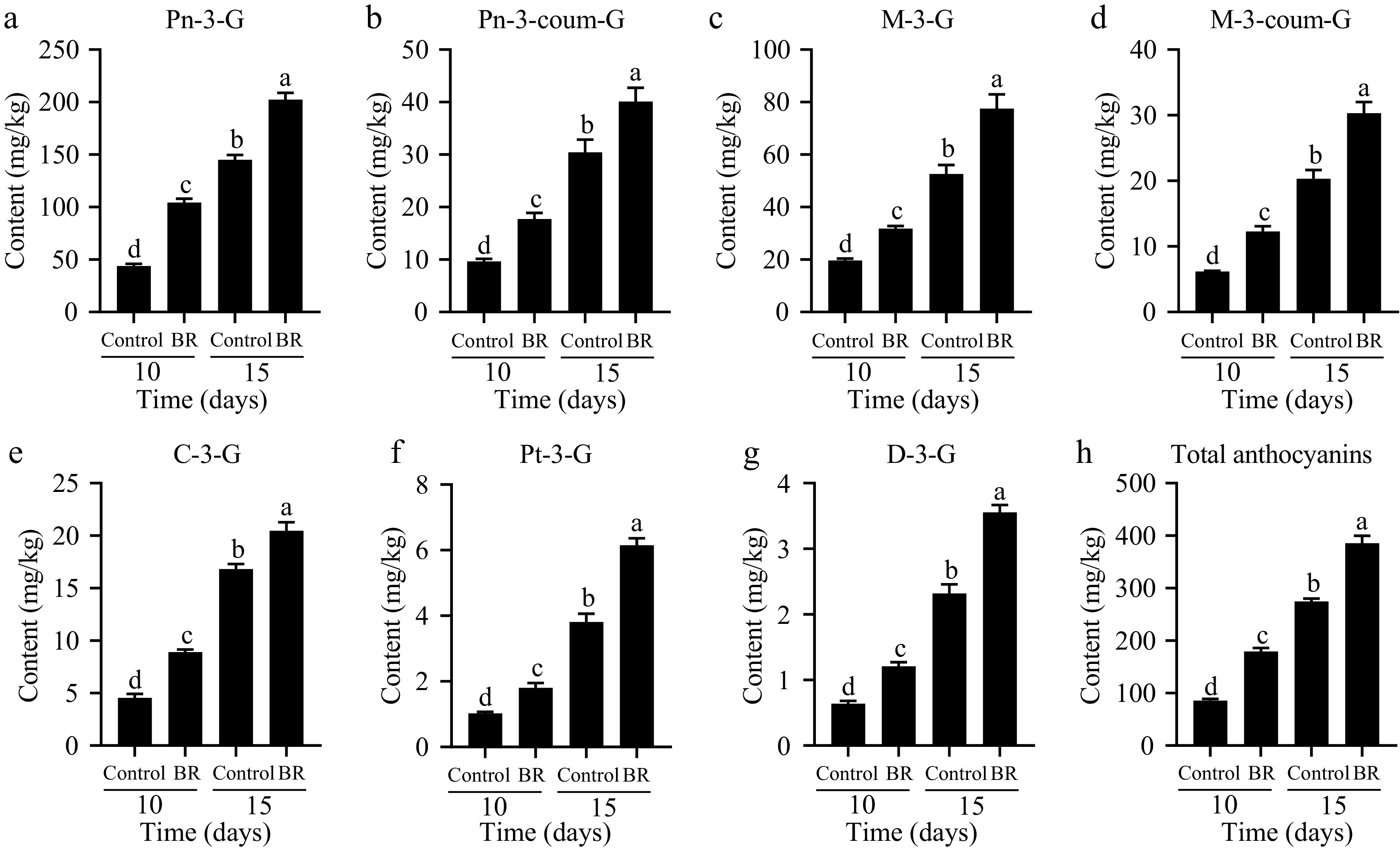
Figure 2.
The effect of exogenous 2,4-epibrassinolide (EBR) treatment on the content and constitute of anthocyanin during berry ripening process. (a) Pn-3-G, peonidin-3-O-glucoside. (b) Pn-3-coum-G, peonidin-3-O-(6-p-coumaroyl)-glucoside. (c) M-3-G, malvidin-3-O-glucoside. (d) M-3-coum-G, malvidin-3-O-(6-p-coumaroyl)-glucoside. (e) C-3-G, cyanidin-3-O-glucoside. (f) Pt-3-G, petunidin-3-O-glucoside. (g) D-3-G, delphinidin-3-O-glucoside. Data are shown as the means ± standard deviation of three biological replicate assays. Different letters indicate significant differences at p < 0.05 according to one-way ANOVA and Tukey's test.
Identification of DEGs between BRs-treated and non-treated groups during berry ripening
-
To identify DEGs in response to BRs, seven samples of BRs-treated and non-treated groups across four stages (0, 5, 10, and 15 d) were subjected to RNA sequencing, with each sample having two biological replicates. A total of 100.01 Gb of clean reads were acquired, with more than 90% of clean reads for every library successfully mapped to the grape reference genome (Supplementary Table S2), suggesting that sequencing data can accurately represent the differences among these samples in the transcription levels. Furthermore, the principal component analysis (PCA) demonstrated the combined effect of developmental stage and EBR treatment on the transcriptome differences among the samples (Supplementary Fig. S1). In total, 4,578, 3,397 genes and 3,569 DEGs were verified post-5, 10, and 15 d of EBR application, respectively (Supplementary Fig. S2). The KEGG analyses of the DEGs displayed that pathways related to the plant hormone signal transduction and phenylpropanoid biosynthesis were significantly enriched (q-value ≤ 0.001) at the véraison stage (10 d after EBR treatment) (Supplementary Fig. S3) and these pathways were essential for the anthocyanin synthesis and berry ripening. Subsequently, there were five main pathways in the berries at the ripening stage (15 d after EBR treatment), including carbon metabolism, biosynthesis of amino acids, phenylpropanoid biosynthesis, plant-pathogen interaction, and flavonoid biosynthesis (Supplementary Fig. S3). Taken together, these findings underscore that BRs treatment regulated a series of secondary metabolic pathways that underpin berry ripening at the transcription level.
BRs application influences the gene expression related to soluble sugar and organic acid metabolism
-
Herein, this study also found the differential expression of several genes related to the soluble sugar and organic acid metabolism under BR treatment. The expression of one sucrose synthase gene (VIT_11s0016g00470) which catalyzes the reversible reaction between sucrose and glucose, displayed nearly 10-fold lower at 15 d after BR application. Otherwise, one sucrose phosphate synthase (VIT_18s0089g00410) gene expressed five times higher than control fruits, and the other one sucrose phosphate synthase gene also showed nearly two times higher than control. In addition, BR application also significantly increased the expression of one starch synthase (VIT_16s0098g01780) (Supplementary Table S8).
Regarding to the organic acid metabolism-related genes, the expression levels of several genes like one NAD-dependent isocitrate dehydrogenase gene (NAD-ICDH: VIT_03s0038g03120), one fumarase gene (FUM: VIT_07s0005g00880), two succinate thiokinase gene (SAT: VIT_07s0005g03790 and VIT_17s0053g00120), one GDP-l-galactose pyrophosphatase gene (GalPP: VIT_10s0405g00030), one phosphoenolpyruvate carboxylase gene (PEPC: VIT_12s0028g02180), one UDP-glucose-dehydrogenase gene (UGDH: VIT_17s0000g06960), were found to be significantly inhibited at 15 d after BR application (Supplementary Table S8). In general, the transcription profiles of sugar and organic acid metabolism genes were also significantly altered by the BR treatment.
Exogenous BRs treatment strongly stimulates anthocyanin biosynthetic-related genes in grape
-
To further evaluate the BRs-mediated anthocyanin accumulation, we scrutinized the transcription profiles of anthocyanin biosynthetic genes. A group of 12 phenylpropanoid pathway genes, including nine phenylalanine ammonia-lyase (VvPAL), one cinnamate 4-hydroxylase (Vv4CH), and two 4-coumarate-CoA ligases (Vv4CL) showed increased expression at 10 d, followed by a decline in expression in 15 d post-EBR application (Fig. 3, Supplementary Table S3). BRs treatment notably enhanced the expression levels in a lot of flavonoid biosynthesis genes. For example, two chalcone synthases (VvCHS: VIT_05s0136g00260 and VIT_14s0068g00920), two flavanone 3-hydroxylase (VvF3H: VIT_04s0023g03370 and VIT_16s0098g00860), one flavonoid 3'-hydroxylase (VvF3'H: VIT_17s0000g07210), one flavonoid 3',5'-hydroxylase (VvF3'5'H: VIT_06s0009g03010), one flavonol synthase (VvFLS: VIT_18s0001g03430) and one leucoanthocyanidin dioxygenase (VvLDOX: VIT_02s0025g04720) exhibited increased expression in 5 d post-EBR application (Fig. 3, Supplementary Table S3). However, dihydroflavonol 4-reductase (VvDFR: VIT_18s0001g12800) shows no discernible response for the exogenous EBR treatment (Fig. 3, Supplementary Table S3).
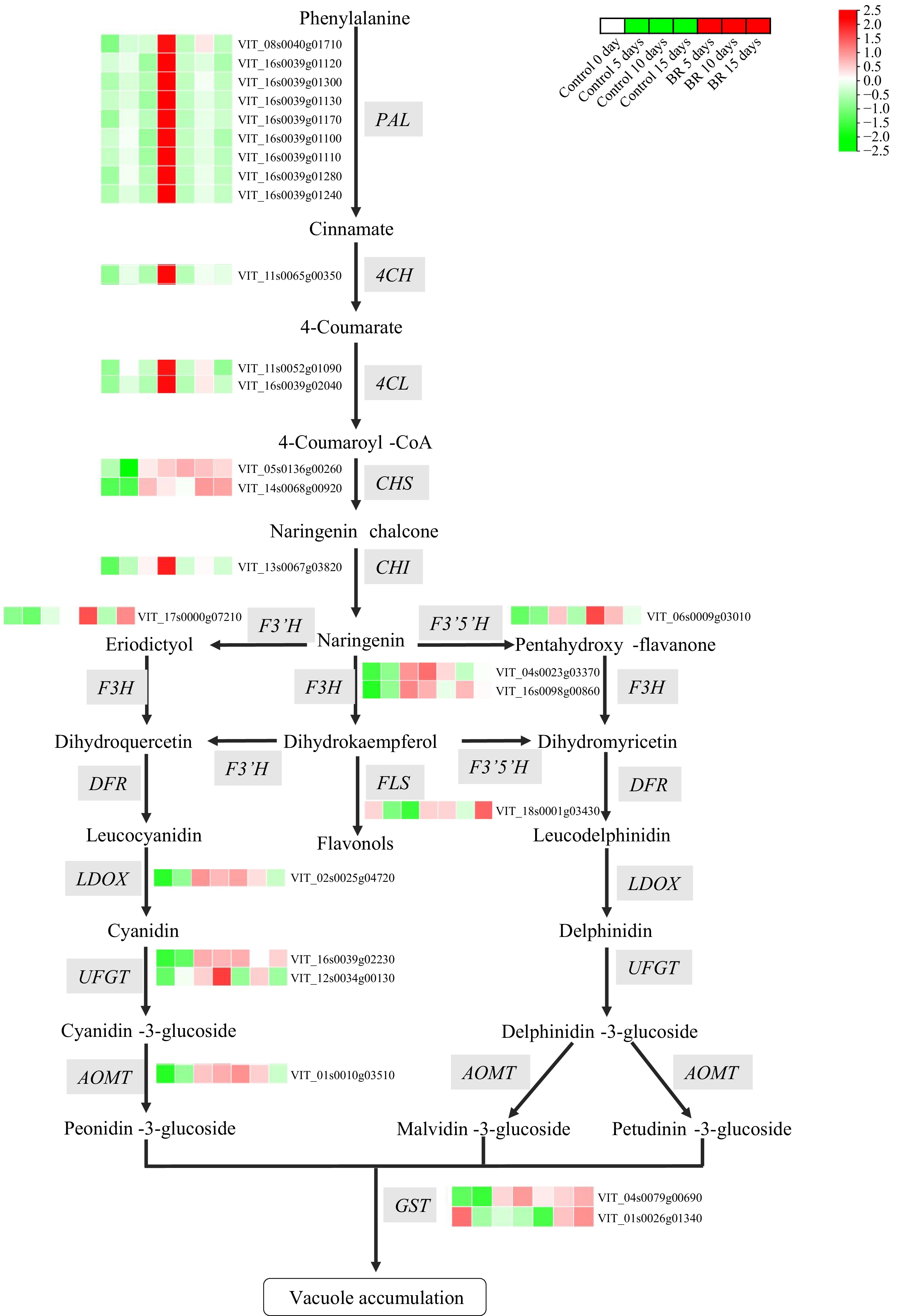
Figure 3.
The effect of exogenous 2,4-epibrassinolide (EBR) treatment on the anthocyanin biosynthetic pathway in grapevine berries. The heatmap was constructed based on relative log2 (FPKM) values obtained by RNA-Seq data during berry development. Green to red color indicates a gradual increase in gene expression level.
Furthermore, the transcription of UDP-glucose: flavonoid-3-O-glucosyltransferase (VvUFGT: VIT_16s0039g02230), which was involved in the final process of anthocyanin biosynthetic pathway, was robustly activated at 5 d after BRs treatment. In contrast, other VvUFGT (VIT_12s0034g00130) showed down-regulated expression at 5 and 15 d after treatment. An anthocyanin O-methyltransferase (VvAOMT: VIT_01s0010g03510, VIT_07s0031g00350, and VIT_03s0063g00140) involved in anthocyanin methylation and two glutathione-S-transferase (VvGST: VIT_04s0079g00690 and VIT_01s0026g01340) involved in anthocyanins transport, exhibited induced transcript level in response to BRs application compared to control berries (Fig. 3, Supplementary Table S3). These results suggested that BRs promoted skin coloring and anthocyanin accumulation in berry by inducing the transcript level of anthocyanin biosynthetic-related genes.
BRs application affects the gene expression of plant hormone biosynthesis and signaling
-
Plant hormones have crucial effects on berry development and ripening. At the transcriptomic analysis, a multitude of genes related to biosynthesis and signaling of different plant hormones, such as BRs, ethylene, abscisic acid (ABA), auxin (IAA), and cytokinin (CTK), were detected responding to EBR application (Fig. 4, Supplementary Table S4). For example, one steroid 22-alpha-hydroxylase (VvDWF4: VIT_04s0023g01630), one 3-epi-6-deoxocathasterone 23-monooxygenase (VvROT3: VIT_04s0023g02650), and one brassinosteroid-6-oxidase (VvBR6OX1: VIT_14s0083g01110) involved in BR biosynthesis were down-regulated under EBR treatment. In BR signaling pathway, three brassinosteroids insensitive 1-associated receptor kinase 1 (VvBAK1: VIT_12s0055g01160, VIT_12s0055g01280, VIT_12s0121g00300), one BES1/BZR1 homolog protein (VvBES1: VIT_10s0003g01790), and one BES1-interacting Myc-like protein 1 (VvBIM1: VIT_17s0000g04790) all demonstrated down-regulated expression patterns upon BR application (Fig. 4a, Supplementary Table S4), suggesting a loop regulation network for regulation in BR biosynthesis pathway.
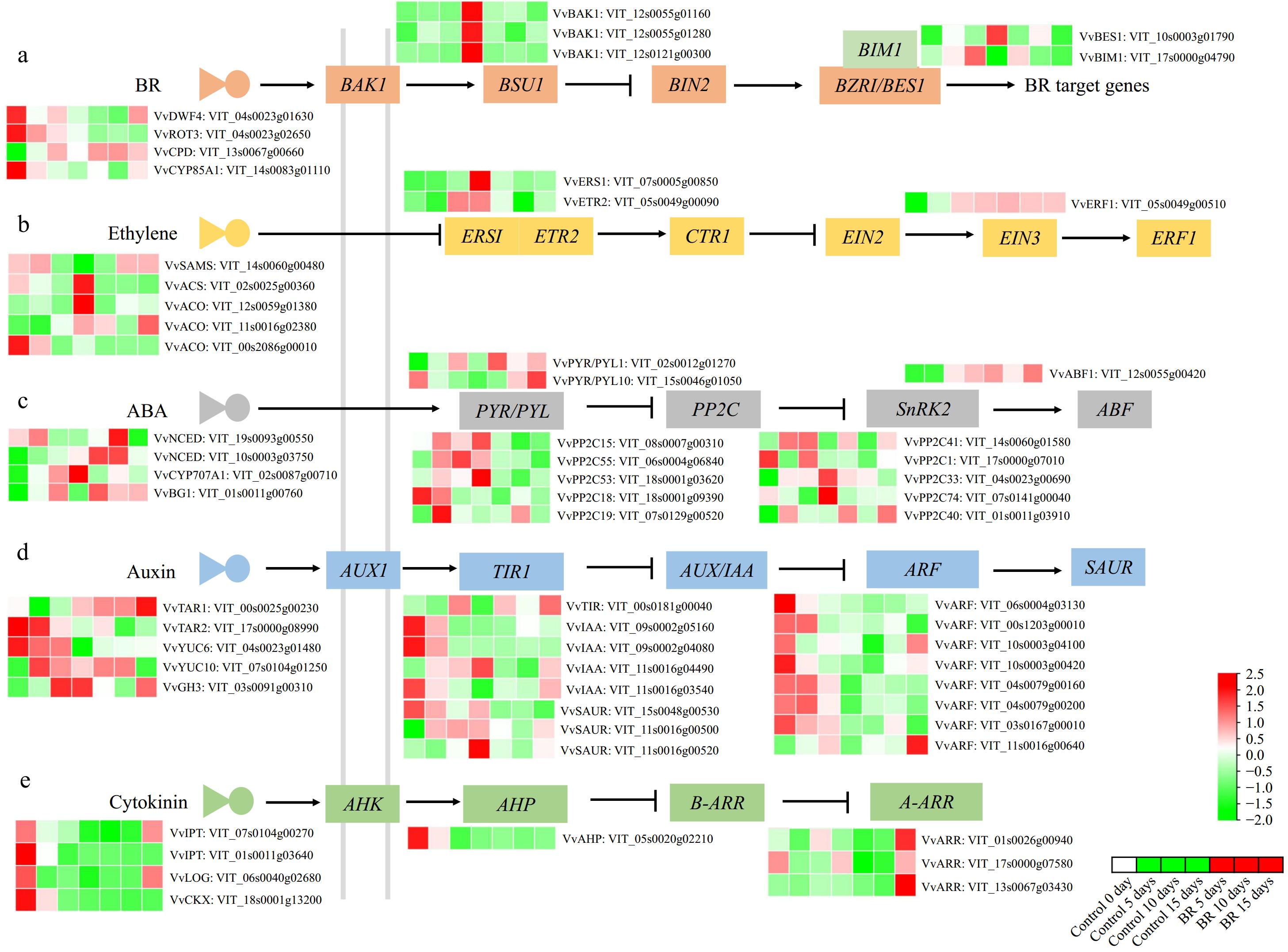
Figure 4.
The effect of exogenous 2,4-epibrassinolide (EBR) treatment on the phytohormones biosynthesis and signaling pathways in grapevine berries. (a) BRs, (b) ethylene, (c) abscisic acid (ABA), (d) auxin, and (e) cytokinin biosynthesis and signaling-related genes responded to EBR application, respectively. The heatmap was constructed based on relative log2 (FPKM) values obtained by RNA-Seq data during berry development. Green to red color indicates a gradual increase in gene expression level.
EBR application evoked pronounced effects on the genes associated with the ABA biosynthetic and signaling pathways. EBR treatment highly induced the transcript level of ABA biosynthesis-related gene (VvNCED: VIT_10s0003g03750) while concurrently leading to a decrease in the transcript level of ABA metabolism-related gene (VvCYP707A1: VIT_02s0087g00710) (Fig. 4c, Supplementary Table S4). In ABA signaling pathway, a notable reduction was observed in 9 VvPP2Cs, known as negative regulators of ABA. Among them, VvPP2C15 (VIT_08s0007g00310) and VvPP2C55 (VIT_06s0004g06840) consistently exhibited down-regulation at all time points. Another three ABA signaling genes, including two VvPYR/PYL (VIT_02s0012g01270 and VIT_15s0046g01050) and one VvABF1 (VIT_12s0055g00420), manifested increased expression following exogenous BRs treatment (Fig. 4c, Supplementary Table S4). Furthermore, two ethylene receptors, ethylene response sensor 1 (VvERS1: VIT_07s0005g00850) and ethylene response 2 (VvETR2: VIT_05s0049g00090), recognized as negative regulators of ethylene responses, exhibited reduced expression under BRs treatment when compared with control berries. However, ethylene response factor 1 (VvERF1: VIT_05s0049g00510) was up-regulated in expression (Fig. 4b, Supplementary Table S4).
Four auxin synthesis genes, including one tryptophan aminotransferase related (VvTAR2: VIT_17s0000g08990), two VvYUCs (VIT_04s0023g01480 and VIT_07s0104g01250), and one IAA-amido synthetases (VvGH3: VIT_03s0091g00310) showed significant down-regulation in response to BRs treatment. Only one VvTAR1 (VIT_00s0225g00230) was obviously up-regulated in all samples. Furthermore, the transcription levels of 13 genes related to auxin, including three Auxin/Indole-3-acetic acid-inducible proteins (VvIAAs), seven auxin response factors (VvARFs), and three auxin-responsive SAUR signaling, underwent significant attenuation following BRs treatment (Fig. 4d, Supplementary Table S4). In the CTK biosynthesis and metabolism pathway, two isopentenyltransferases (VvIPTs: VIT_07s0104g00270 and VIT_01s0011g03640) and one cytokinin oxidase gene (VvCKX: VIT_18s0001g13200) showed decreased expression in 5 d post-BRs application (Fig. 4e, Supplementary Table S4). Additionally, one histidine-containing phosphotransfer protein (VvAHP: VIT_05s0020g02210), the positive regulator of CTK signaling, expression of auxin and CTK biosynthetic and signaling pathway genes.
BRs application enhances berry softening through regulating the transcription of cell wall-related genes
-
The effect of BRs in grape berry softening was thoroughly investigated, including the evaluation of berry firmness, protopectin content, soluble pectin content, and cellulose content. BRs application significantly accelerated the decline in berry firmness and promoted berry softness (Fig. 5a). The cellulose and protopectin contents were lower in BRs-treated samples compared with the control group (Fig. 5b & c), while the water-soluble pectin (WSP) content was higher at all time points (Fig. 5d). These results showed that BRs treatment accelerated berry softening and cell wall degradation by decreasing the cellulose and protopectin content, and increasing the WSP content.
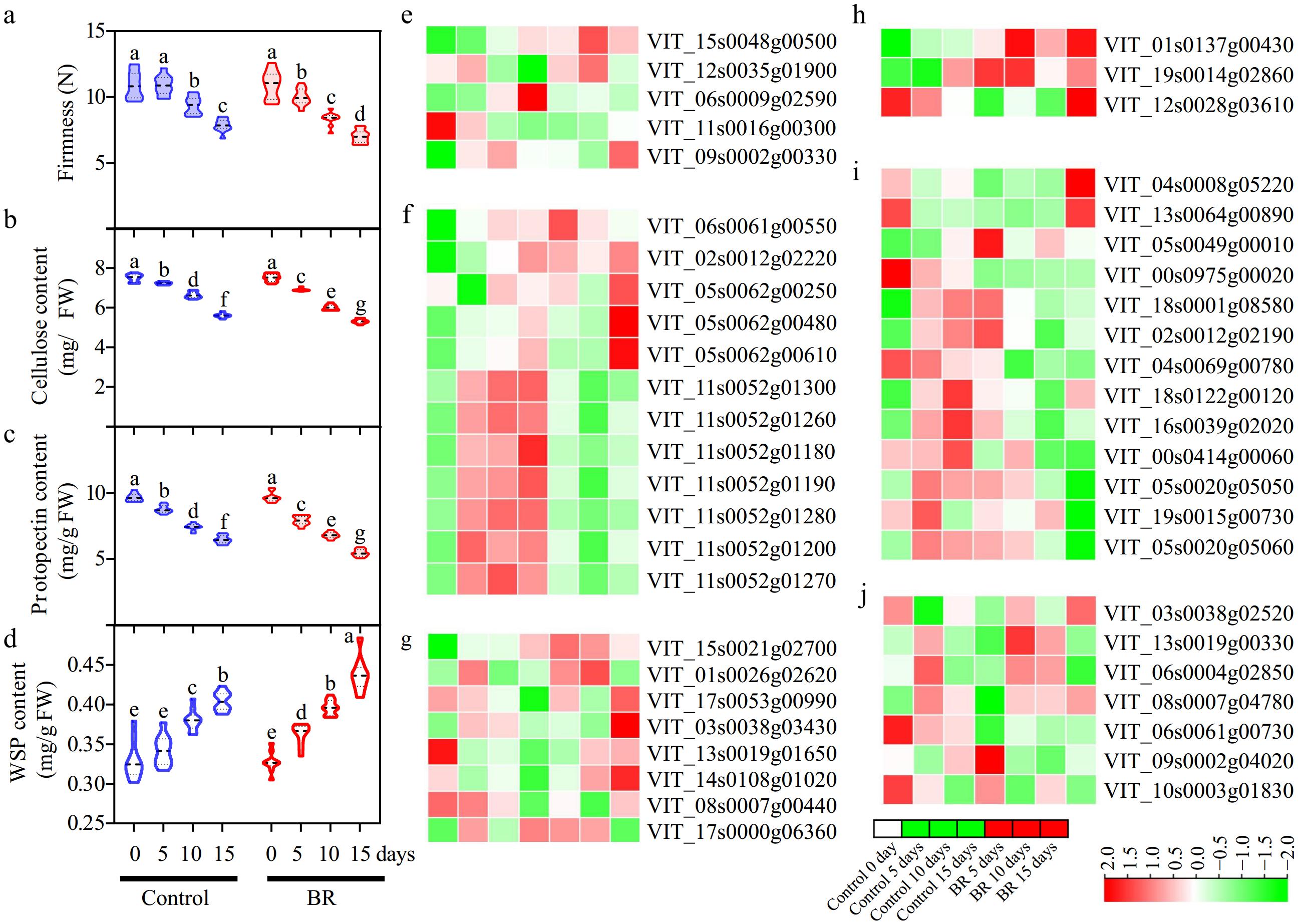
Figure 5.
The effect of exogenous 2,4-epibrassinolide (EBR) treatment on the cell wall components and the expression of genes related to the degrading enzymes of the cell wall. (a) Firmness, (b) cellulose, (c) protopectin, and (d) water-soluble pectin (WSP) content after EBR treatment. Data are expressed as mean ± standard from three biological replicates. Different letters indicate significant differences (p < 0.05) between two different treatments at each sampling point.
Transcripts related to cell-wall degradation during ripening were found in response to EBR application (Supplementary Table S5). At first, the transcription level of two pectin methylesterases (VvPMEs: VIT_15s0048g00500 and VIT_12s0035g01900) was robustly induced by EBR treatment at two stages (Fig. 5e). The expression levels of VvXET (VIT_06s0061g00550), a prominent gene of xyloglucan endotransglucosylase family, exhibited significant induction (Fig. 5f). However, seven VvXETs (VIT_11s0052g01300, VIT_11s0052g01260, VIT_11s0052g01180, VIT_11s0052g01190, VIT_11s0052g01280, VIT_11s0052g01200 and VIT_11s0052g01270) demonstrated inhibited expression due to BRs application at all three stages (Fig. 5f), indicating VvXETs might have opposite role in BRs-mediated berry softening. Furthermore, 10 cellulose synthases (VvCES) (Fig. 5i) and three cellulases (VvCEL) (Fig. 5h), respectively, displayed down- and up-regulated expression responding to BRs application, which was consistent with the reduction in cellulose content (Fig. 5b). Additionally, six expansin (VvEXP) genes, which are responsible for the extension of cell walls, showed increased expression compared to the control groups (Fig. 5g). Moreover, aquaporins (VvAQP: VIT_03s0038g02520, VIT_13s0019g00330, VIT_06s0004g02850 and VIT_08s0007g04780), which are participated in the enlarge and soften of berry, were also up-regulated post-BRs application (Fig. 5j).
BRs application alters the transcription levels of multiple transcription factor families
-
BRs treatment had a strong effect on the expression of multiple transcription factor families, including bHLH, bZIP, ERF, MYB, NAC, and WRKY. Specifically, two bHLHs, four bZIPs, and two VvMYBs showed increased expression, whereas four bHLHs, three bZIPs, and five VvMYBs exhibited decreased expression in response to BRs treatment (Supplementary Table S6). Among them, the expression of bHLH, VvbHLH013 (VIT_11s0052g00100), was markedly inhibited at two stages of BRs treatment (Supplementary Table S6). Moreover, four VvMYB members (VIT_03s0180g00210, VIT_18s0001g11170, VIT_18s0001g09850 and VIT_07s0129g01050) belonging to the S22 subfamily also demonstrated down-regulated expression (Supplementary Table S6), indicating their potential role as negative regulators in BRs signaling during berry ripening. Furthermore, total a total of 10 VvNACs showed contrasting expression patterns under BRs treatment. Among them, two VvNACs (VIT_02s0012g01040 and VIT_01s0146g00280) displayed increased expression at two stages, whereas three VvNACs (VIT_15s0048g02280, VIT_17s0000g00770 and VIT_14s0068g01490) exhibited decreased expression at two stages responding to BRs application (Supplementary Table S6).
Interestingly, only one VvERF gene (VIT_05s0049g00510) displayed remarkably accumulated transcription levels on grape berry, and it was notably induced at 5 d post EBR treatment (Supplementary Table S6). However, eight VvERF genes (VIT_12s0028g03270, VIT_02s0025g04460, VIT_16s0100g00380, VIT_18s0001g03240, VIT_11s0016g03350, VIT_03s0063g00460, VIT_11s0016g00670 and VIT_16s0013g00990) had decreased expression at all time points (Supplementary Table S6). Additionally, six VvERF genes also exhibited down-regulated transcription levels at one or two time points following BRs application (Supplementary Table S6). Similarly, only one VvWRKY gene (VIT_12s0059g00880) exhibited up-regulated expression, whereas three VvWRKYs (VIT_09s0018g00240, VIT_04s0008g05760, and VIT_02s0025g01280) displayed down-regulated transcription level in all three-time points after BRs application (Supplementary Table S6). Seven and five VvWRKYs demonstrated down-regulated transcription levels in the first two phases, respectively (Supplementary Table S6). The results indicated that VvERFs and VvWRKYs might play negative regulatory roles in BRs-mediated berry ripening.
qRT-PCR validation of DEGs based on RNA-seq
-
Twelve DEGs were chosen for qRT-PCR identification between control and BR-treated berries to confirm the precision and credibility of sequencing results. Five genes (VvNCED, VvTAR1, VvCTR, VvPME, VvEXP) exhibited up-regulation due to EBR treatment. Seven genes (VvROT3, VvPRP1, VvERF058, VvLRR, VvCabBP, VvPUP and VvUbox) were down-regulated by BRs treatment (Supplementary Table S7). Expression verification revealed that although the fold-changes obtained by qRT-PCR and RNA-Seq did not align precisely, the overall expression profiles between the two techniques were largely congruent to each other (Fig. 6), implying that the RNA-Seq data are reliable and reproducible.
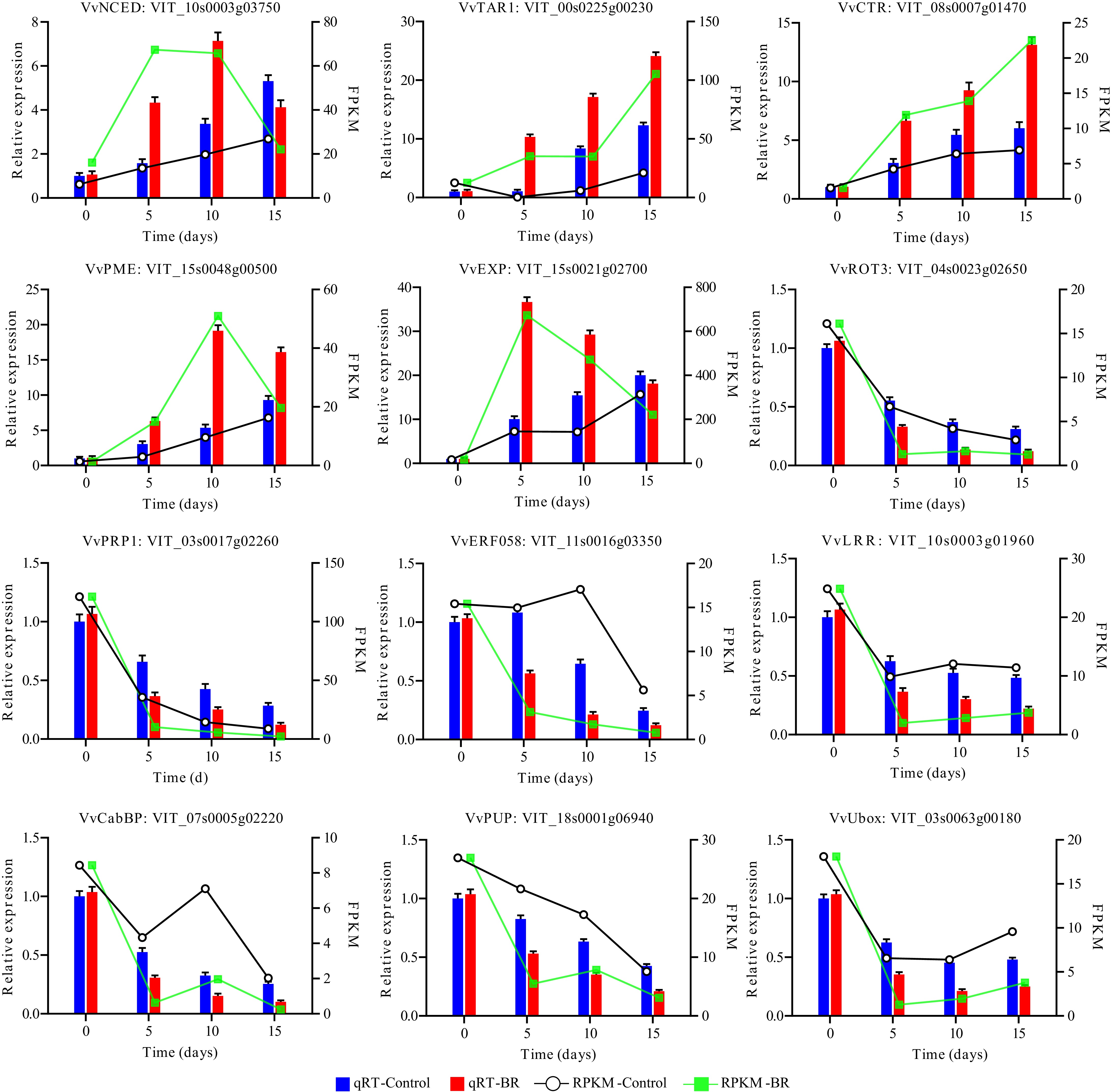
Figure 6.
Validation of 12 differentially expressed genes by qRT-PCR under EBR treatment in grapevine berry.
Transient overexpression in VvDWF4 promotes anthocyanin accumulation and fruit softening in strawberry fruit
-
Due to VvDWF4 being regulated in all three experimental stages, VvDWF4, a BR biosynthesis gene, was selected to confirm the significance of BRs during berry ripening by a strawberry transient expression system. Overexpression of VvDWF4 significantly promoted fruit coloration and anthocyanin accumulation (Fig. 7a, b), 6 d post-infiltration with VvDWF4, fruits began to exhibit a reddening color and became almost entirely red within 10 d post-infiltration, while the control fruits reached approximately intermediate red stage within the same period (Fig. 7a). The concentrations of total anthocyanin in VvDWF4-overexpression fruits were up to 2.4-times higher on the 10th day compared with the control fruit (Fig. 7b). Transient overexpression of VvDWF4 into strawberry fruit significantly enhanced the BRs content at 5 and 10 d of treatment (Fig. 7c), indicating an important role of VvDWF4 in BRs-induced anthocyanin accumulation. Furthermore, transient overexpression of VvDWF4 in strawberry increased the expression of both BRs biosynthetic genes (FaBR6OX1 and FaCPD) and the BRs receptor (FaBRI1) (Fig. 7d−f). Additionally, transient overexpression of VvDWF4 also promoted fruit softening (Fig. 7g). VvDWF4-overexpression fruit exhibited a significant increase in water-soluble pectin content and concurrent reduction in protopectin content when compared to the control fruit (Fig. 7h, i), resulting in rapid softening of the fruit.
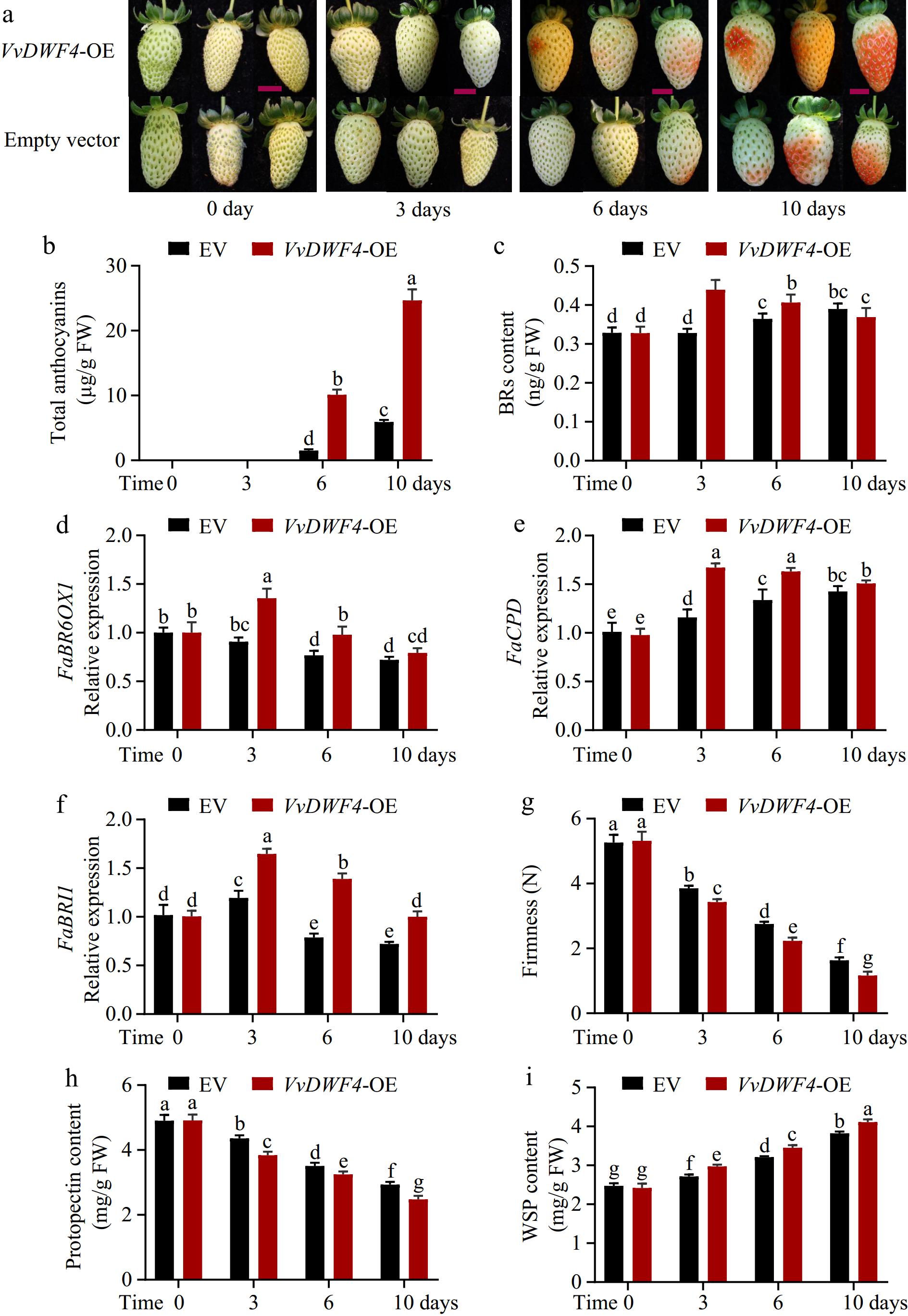
Figure 7.
Effects of transient overexpression of VvDWARF4 (VvDWF4) in strawberry fruits. (a) The phenotype of strawberry fruits after transient overexpression VvDWF4. The red bar indicates 5 cm. (b) Anthocyanin contents in grape shin. FW, fresh weight; EV, empty vector. (c) BRs content. The relative expression of brassinosteroid-6-oxidase (BR6OX1) (d), cytochrome P450 90A1 (CPD) (e), and BRASSINOSTEROID-INSENSITIVE 1 (BRI1) (f), respectively. (g) Firmness, (h) Protopectin, and (i) water-soluble pectin (WSP), content after transient overexpression VvDWF4. Data are expressed as mean ± standard from three biological replicate analysis, and different letters indicate significant differences (p < 0.05) between two different treatments at each sample.
-
The exogenous application of plant hormones is a crucial method to enhance the quality attributes of grape berry, such as their size, color, flavor, and sensory traits[28]. BRs, which is known as the environmental-friendly hormone, have been extensively used to improve yield and confer resilience in responding to a range of stress factors[19]. BRs are implicated in the development and ripening process of both climacteric (tomato and mango) and non-climacteric (strawberry and grape) fruit[21,29−31]. Similarly, current findings demonstrate that epibrassinolide (EBR) treatment increased endogenous BR contents and promoted grape berry ripening (Fig. 1). However, the molecular mechanism of BRs-mediated berry ripening is not perfectly clear in grape. Therefore, we employed a combination of physiological and transcriptomic analyses to delve into the underlying biological processes and identify potential candidate genes involved in BR-mediated berry development and ripening.
BRs homeostasis was maintained via a feedback regulation during berry ripening in grape
-
It is well-established that endogenous BR contents undergo a significant increase during the onset of the berry ripening process[3]. During this research, the BR contents notably increased at the initial stage of EBR treatment, leading to accelerated grape berry ripening (Fig. 1b), indicating that BRs may serve as potent elicitors during grape berry ripening. As previously reported, BR homeostasis is primarily regulated by intricate feedback transcriptional regulation at various steps in the BR biosynthetic routine[32]. For example, four BRs biosynthetic genes in Arabidopsis, including AtDWF4, AtCPD, AtBR6ox1, and AtROT3, exhibited decreased expression in response to exogenous BL treatment but increased expression when subjected to the BR synthesis inhibitor BRZ[7,8]. Correspondingly, VvBR6OX1 and VvDWF1, two BR biosynthesis-related genes, also showed significant down-regulated transcription level responding to exogenous EBR application in grapes. Furthermore, transient overexpression in VvDWF4 from strawberry fruit significantly improved the accumulation of BR contents through up-regulating transcription level of BRs biosynthetic genes and positive signaling gene BRI1 (Fig. 7), indicating a positive correlation between fruit ripening and transcription level of BR biosynthetic genes, including GhDWF4 in upland cotton and SlCYP90B3 (SlDWF4) of tomato[33]. All the results illustrated BR homeostasis is maintained by the feedback regulatory expression in multiple genes associated with BR biosynthesis in grape.
BRs promote ripening through the coordination of multiple hormone cross-talk networks
-
Different plant growth regulators, like ethylene, ABA, and auxin, have participated in the regulation network of berry ripening of grapes[22,34]. For example, BRs application have been demonstrated to improve ethylene accumulation and fruit ripening of tomato and banana by amplifying the transcription levels of ACS and ACO genes[35]. Herein, EBR application down-regulated transcription level of two negative regulators of ethylene signaling, VvERS1 (VIT_07s0005g00850) and VvETR2 (VIT_05s0049g00090), whereas up-regulated the expression of the positive regulator of ethylene signaling, VvERF1 (VIT_05s0049g00510), suggesting that EBR application promotes ethylene signal transduction during berry ripening in grape. Similarly, BRs initiate a cascade of events leading to ripening by increasing ethylene levels in tomatoes[29].
BRs and ABA often exhibit antagonistic interactions in many plants' growth and stress response processes, including seed germination and dormancy[36], primary root development[37], and stomatal conductance[38]. For instance, high accumulation levels of BRs have appeared to suppress ABA-induced stomata closure by inhibiting ABA biosynthesis[38]. However, EBR treatment has been shown to increase ABA content and induce the transcription of ABA biosynthesis (VvNCED1, VvNCED2, and VvZEP) and signaling-related genes (VvSnRK2.6, VvPP2C4, VvABF1, and VvABF2) under water stress[39]. The current findings also reveal that EBR treatment activates ABA biosynthesis (VvNCED: VIT_10s0003g03750) and signaling-related genes (VvPYR/PYL: VIT_02s0012g01270, VIT_15s0046g01050, and VvABF1: VIT_12s0055g00420) while reducing the expression of 9 VvPP2Cs, which function as negative regulators within ABA signaling pathway (Fig. 4, Supplementary Table S4). Collectively, BR might exert a positive influence on ABA signaling during berry ripening, suggesting a more complex crosstalk mechanism between BR and ABA throughout the grape berry ripening process.
The crosstalk and interplay between BRs and auxin are complex and extensive, crucial for plant development. BRs and auxin work synergistically to regulate hypocotyl elongation and lateral root development[40]. BRs are also believed to be essential for fruit sets, reminiscent of auxin application. Interestingly, while exogenous BRs application promotes ripening[3], while auxin delays ripening[34]. Most auxin signaling-related genes, including three VvIAAs, seven VvARFs, and three VvSAUR, were significantly inhibited in response to EBR treatment (Fig. 4, Supplementary Table S4), indicating that BRs and auxin might exert opposing effects during the grape ripening process.
BRs accelerate berry color and softening by mediating anthocyanin- and cell wall- associated genes
-
In the current study, exogenous EBR application significantly enhanced berry coloration and anthocyanin concentration in grape berries (Fig. 1). Several researches have also indicated that BRs are involved in the development and ripening processes of different fruit crops, including tomato[29], mango[30], strawberry[31], persimmon[41], and jujube[42]. For example, EBR treatment was shown to regulate secondary metabolism by promoting the gene expression and enzyme activity of PAL in tomato[43]. Several studies in grapes have indicated that BRs treatment not only increases the activities of PAL and UFGT enzymes[19] but also stimulates transcript level of anthocyanin biosynthetic genes (VvCHI1, VvCHS2, VvCHS3, VvDFR, VvUFGT)[20]. The results depict that almost all anthocyanin structural and constitute genes were up-regulated responding to BR application in the first two stages, except for VvDFR (Fig. 3). These findings strongly indicated that exogenous BRs application activates the accumulation of anthocyanin through handling the transcription level of anthocyanin biosynthetic genes in grape.
Fruit texture and softening are critical factors for fruit shelf life and storage, primarily attributed to cell wall disassembly during ripening[44]. BRs have been implicated in berry ripening and softening. For example, BR treatment has been shown to accelerate ethylene production, enhance respiration rate, facilitate fruit coloring, and improve the development and softening of the fruit during the ripening process of mango[30]. In tomatoes, overexpressing SlCYP90B3 which is known as an important enzyme involved in BR biosynthesis, elevated the levels of bioactive BRs, promoting the ripening process of various fruits, including accelerated softening, increased soluble sugar, and enhanced flavor volatile concentrations[33].
Fruit softening is tightly linked to the degradation of cell-wall-related constituents, such as pectin and cellulose. In this study, the application of EBR significantly increased WSP content while reducing protopectin and cellulose contents, leading to berry softening in grapevine (Fig. 5), which is in line with recent studies in persimmon fruit[41]. In addition, transient overexpression of VvDWF4 in strawberry fruit also enhanced fruit softening by elevating WSP content and decreasing protopectin levels (Fig. 7). The process of cell wall degradation involves the role of multiple degrading enzymes of cell-wall. In persimmon, EBR application accelerated fruit softening by up-regulating the expression of genes related to cell-wall degrading pathway, including DkPG1, DkPL1, DkPE2, and DkEGase1, and increasing the activities of wall-degrading enzyme, including polygalacturonase, pectate-lyase, and endo-1,4-beta-glucanase, ultimately leading to fruit softening[41]. Correspondingly, various gene families encoding cell wall-degrading enzymes, including VvPME, VvXET, VvCEL, VvEXP, and VvAQP, were extensively regulated by BRs treatment (Fig. 5). These findings strongly support the positive association between BRs and grapevine berry ripening, either by impacting the genes about cell-wall degrading enzymes or through crosstalk network with other hormones.
As a non-toxic and ecologically friendly plant hormone, BRs have been shown to promote the growth, yield, and quality, stress tolerance of plants, and it is also effective at very low doses and degrade easily[45−48]. For example, BRs-treated mangoes outperformed the control group in terms of quality, including fruit weight, yield, the content of total soluble solids, total sugar, titratable acid, and β-carotene[45−48]. In addition, the application of BRs can reduce the dependence on chemical fertilizers and pesticides, thereby reducing environmental pollution[49,50].
The high cost of BRs may limit their widespread use in the world. However, in order to reduce the cost of BRs synthesis and improve the stability when applied, many types of BRs analogs that were commercially synthesized have been developed to make them more affordable[49,50]. These synthetic analogs are used in many countries to improve the performance and yield of agricultural and horticultural crops[49,50].
In summary, the application of BRs in large-scale industrial production has potential, especially in improving crop yield and quality. However, cost and application technology are key factors to consider during implementation. The cost problem can be partially solved by using more cost-effective synthetic analogs, while the precise application of the technology requires further research and development.
-
The findings of this study provide compelling evidence that the exogenous BRs positively regulate numerous ripening processes, including sugar and anthocyanin accumulation, color development, and berry softening. Transcriptome analysis has unveiled the intricate crosstalk network among BRs and distinct plant hormones, including ethylene, abscisic acid, auxin, and cytokinin, in elevating the ripening process from grapes. The BR-induced color development and anthocyanin accumulation are positively linked with the up-regulation of anthocyanin biosynthetic genes. Moreover, epibrassinolide (EBR) treatment elevates WSP content while reducing protopectin and cellulose levels, ultimately leading to berry softening by modulating the genes responsible for cell wall-degrading enzymes. These findings offer valuable insights into the potential benefits of utilizing BRs to enhance the visual and nutritional qualities of grape berries without compromising yield.
This work was supported by the Key Research and Development Plan of Shandong Province (Grant Nos 2022LZGCQY019, 2023TZXD015 and 2022TZXD0011), the National Natural Science Foundation of China (NSFC) (Grant Nos 32102353, 32202449, and 32202430), the Natural Science Foundation of the Jiangsu Higher Education Institutions of China (Grant No. 24KJB210009), the Natural Science Research Project of Anhui Educational Committee (Grant No. 2024AH051983), the Natural Science Foundation of Jiangsu (Grant No. BK20190542), Shandong Provincial Natural Science Foundation (Grant Nos ZR2021QC005 and ZR2022QC018), Construction of genetic transformation of Grape (Grant No. SDAG2021A03), Inner Mongolia Science Technology Plan (Grant No. 2022YFDZ0029), the Unveiling and Commanding project of the West Coast new area of Qingdao (2022-23) and Qingdao Agricultural University Enterprise Cooperation Projects (Grant No. 660/2424191).
-
The authors confirm contribution to the paper as follows: Project administration: Zhu X, Leng X, Gong D; formal analysis, data curation: Ren Y, Xu X, Ma Y; resources: Zhao P, Sun X, Zhang L, Zhang Z, Cui Z, Li Z; writing-review & editing: Zhu X, Leng X, Haider M; funding acquisition: Zhu X, Leng X; validation: Ren Y, Xu X, Ma Y; visualization: Liu Y, Zhu X, Leng X; investigation: Liu Y, Ji X; methodology: Liu Y, You R, Ji X; writing-original draft: Liu Y. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
-
Authors contributed equally: Yuanxia Liu, Ruyue You
- Supplementary Table S1 The primers sequences of differentially expressed genes for qRT-PCR.
- Supplementary Table S2 RNA sequencing assembly information.
- Supplementary Table S3 Effects of BR treatment on the expression of anthocyanin biosynthetic related genes.
- Supplementary Table S4 Effects of BR treatment on the expression of phytohormone biosynthesis and signaling pathways genes.
- Supplementary Table S5 Effects of BR treatment on the expression of cell wall associated genes.
- Supplementary Table S6 Effects of BR treatment on the expression of transcription factors.
- Supplementary Table S7 The FPKM for the qRT-PCR validation.
- Supplementary Table S8 Effects of BR treatment on the expression of sugar and organic acid metabolism related genes.
- Supplementary Fig. S1 Principal component analysis based on the expression data from whole normalized gene.
- Supplementary Fig. S2 Venn diagrams and volcano plots displaying the numbers of differentially expressed genes (DEGs) in 2,4-epibrassinolide (EBR) treated berries, according to the developmental stage.
- Supplementary Fig. S3 Statistics of 2,4-epibrassinolide (EBR) induced KEGG pathway enrichment among three developmental stages.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of China Agricultural University, Zhejiang University and Shenyang Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu Y, You R, Gong D, Zhao P, Ji X, et al. 2025. Effect of brassinosteroids on grape berry ripening by coordinating transcriptomic and metabolic analysis. Food Innovation and Advances 4(2): 253−265 doi: 10.48130/fia-0025-0024 

Effect of brassinosteroids on grape berry ripening by coordinating transcriptomic and metabolic analysis
- Received: 12 October 2024
- Revised: 03 March 2025
- Accepted: 04 March 2025
- Published online: 26 June 2025
Abstract: Brassinosteroids (BRs) are essential for the regulation of plant growth, development, and stress responses. However, a comprehensive understanding of the transcriptional regulatory network governed by BRs that orchestrates grape berry ripening remains limited. Herein, the application of exogenous epibrassinolide (EBR) increased endogenous BRs content while down-regulating the expression of BRs biosynthetic genes, indicating feedback transcriptional regulation of BRs biosynthesis. EBR treatment accelerated coloring and enhanced anthocyanin accumulation, which was closely associated with the up-regulation of anthocyanin biosynthetic genes. In addition, EBR significantly increased water-soluble pectin (WSP) content and concurrently reduced protopectin and cellulose levels, resulting in berry softening. The study also delved into the intricate cross-talks between BRs and other plant growth regulators, for instance, ethylene, abscisic acid (ABA), auxin, and cytokinin. Moreover, transient overexpression of VvDWF4 (CYP90B1 steroid 22-alpha-hydroxylase) in strawberry increased BRs content, leading to anthocyanin accumulation and fruit softening. These findings provide compelling evidence that the pre-véraison application of BRs provide an effective strategy for enhancing grape berry quality.
-
Key words:
- Grape /
- Brassinosteroids /
- Anthocyanin /
- Softening /
- Ripening /
- VvDWF4