









-
Dandelion (Taraxacum officinale), a medicinal plant of significant nutritional and pharmacological value, exhibits hypolipidemic, antioxidant, anti-inflammatory, and antidiabetic properties[1]. All parts of the plant, including roots, leaves, flowers, and stems, are utilized in functional foods and traditional medicines. Notably, dandelion leaves are of particular interest due to their health benefits as salads globally[2] and as a vegetable food in northern China[3].
The growing consumer demand for plant-based health products has driven the rapid development of dandelion tea as a functional beverage, recognized for its potent antioxidant capacity[2,4]. However, despite its widespread consumption, key questions regarding how brewing parameters—especially steeping duration—affect its chemical stability and microbial safety remain unclear. Studies on tea often recommend short-term brewing (7–10 min) to optimize the extraction efficiency of bioactive compounds[5,6]. Nevertheless, prolonged steeping practices, such as overnight soaking, are still common, and there is a lack of experimental evidence to support the widespread claim that 'overnight tea is harmful'.
Using metabolomic analysis, key bioactive constituents in dandelion tea were identified, including gallic acid, chlorogenic acid, and flavonoids, which collectively underpin its therapeutic effects[7]. However, these compounds are heat-sensitive and prone to oxidation, suggesting that long-term steeping at high temperatures may reduce their nutritional value[8]. Furthermore, microorganisms have been identified in various types of tea, including green, black, and Pu-erh teas[9,10]. While some of these microorganisms can enhance the flavor and taste of tea, others can be detrimental to human health. For instance, common pathogens in tea, such as Bacillus spp., can accumulate over time in tea[11,12]. However, systematic microbial analysis of dandelion tea under time-gradient brewing conditions remains unexplored.
This study employed metabolomics analysis (UHPLC-QTOF-MS) and 16S rRNA microbiome sequencing of dandelion tea infusions to address two core questions concerning long-term brewing conditions. First, how do the chemical composition and antioxidant capacity of dandelion tea change over time under extended brewing conditions? Second, does long-term brewing, especially at room temperature, affect the microbial community structure, thereby impacting the quality of the tea and potentially posing health risks? Since there is no strict definition of 'overnight tea', brewing durations of 12, 24 h, and an extreme condition of 36 h were selected; although 36 h significantly exceeds typical consumption habits, it was included for mechanistic analysis. By analyzing temporal changes in both metabolite signatures and microbial populations, this work provides theoretical insights into mitigating health risks associated with dandelion tea consumption.
-
Dried dandelion leaf samples were obtained from Bozhou Yushuntang Biotechnology Co., Ltd (Anhui, China). A total of 5 g of dried leaf material was placed into a 500 mL beaker, followed by the addition of 500 mL of boiling water for infusion. To simulate typical consumption conditions, the beaker was left uncovered during the brewing process at room temperature (22–25 °C). Tea infusions were collected at 1, 12, 24, and 36 h post-infusion (labeled as H1, H12, H24, and H36, respectively). These tea infusions were included to evaluate time-dependent metabolic and microbial changes under extended steeping conditions. These samples were not intended to represent recommended consumption states but rather to identify potential degradation thresholds and safety concerns associated with prolonged infusion. Three biological replicates were prepared per time point, with 20 mL of tea liquor retrieved at each time point and immediately stored at –80 °C for subsequent metabolomic analyses. Concurrently, the remaining tea infusion at each time point was centrifuged, and 0.1 g of the resulting precipitate (pellets) was transferred into sterile centrifuge tubes and stored at –80 °C for microbiome analysis.
Although the 36 h steeping duration exceeds conventional consumer practices, this extended time point was included for two reasons. First, the concept of 'overnight tea' lacks a strict definition in public health discourse. A time gradient was therefore selected encompassing 12 h (a common overnight duration), 24 h (frequently used in informal long-soaking practices), and 36 h, which simulates extreme yet plausible consumer behaviors, such as prolonged room-temperature storage. Second, following the principles of food safety risk assessment proposed by national regulatory agencies (e.g., the China National Center for Food Safety Risk Assessment,
www.cfsa.net.cn ), toxicological studies should account for 'reasonably foreseeable maximum exposure scenarios'. Thus, including a 36 h condition allows for the assessment of potential risks under worst-case exposure conditions and provides more scientifically grounded recommendations.Metabolomic sequencing and analysis
-
The tea soup samples collected at 1, 12, 24, and 36 h were subjected to widely targeted metabolomic detection using a Shim-pack UFLC SHIMADZU CBM30A UPLC system coupled to a QTRAP® 4500+ triple quadrupole-linear ion trap mass spectrometer (AB Sciex, USA) at PANOMIX Biomedical Technology Co., Ltd. (Suzhou, China). For sample preparation, tea infusions were freeze-dried using a vacuum freeze-dryer (Scientz-100F), and 100 mg of lyophilized powder was ground with a mixer mill (MM 400, Retsch) at 30 Hz for 1.5 min using a zirconia bead. The powder was then extracted with 1.2 mL of 70% methanol by vortexing for 30 s every 30 min (six times total) and left overnight at 4 °C. After centrifugation at 12,000 rpm for 10 min, the supernatant was filtered through a 0.22 μm membrane (SCAA-104, ANPEL, Shanghai, China) before injection.
Chromatographic separation was performed on a Waters ACQUITY UPLC HSS T3 C18 column (1.8 µm, 2.1 mm × 100 mm) with a mobile phase consisting of water containing 0.1% formic acid (A) and acetonitrile containing 0.1% formic acid (B), using the following gradient: 95:5 A/B at 0 min; 5:95 A/B at 10.0 min; held to 11.0 min; returned to 95:5 A/B at 11.1 min and maintained until 15.0 min. The column temperature was maintained at 40 °C, the flow rate was 0.4 mL/min, and the injection volume was 2 μL.
The mass spectrometer was operated in both positive and negative ion modes using an electrospray ionization (ESI) source, with source temperature at 550 °C, ion spray voltage set at +5,500 V (positive) and –4,500 V (negative), and gas settings of 50 psi (GS1), 60 psi (GS2), and 25 psi (CUR). Collision-activated dissociation (CAD) was set to high. Data acquisition was performed in multiple reaction monitoring (MRM) mode using nitrogen as the collision gas, with compound-specific optimization of declustering potential (DP) and collision energy (CE). Instrument tuning and calibration were conducted using polypropylene glycol standards, and MRM transitions were scheduled according to metabolite retention times.
Metabolite identification was based on a self-built MS/MS spectral library provided by the service platform (PANOMIX Biomedical Technology Co., Ltd.), matching the acquired secondary fragmentation spectra against known reference spectra. During the annotation process, isotopic peaks, adduct signals (e.g., K+, Na+, NH4+), and fragment ions derived from larger molecules were excluded to avoid redundancy and misidentification.
Metabolite quantification was performed using the MRM mode of the triple quadrupole mass spectrometer, which selectively monitors specific precursor-product ion pairs for each compound. This approach enhances specificity by filtering precursor ions in Q1, inducing fragmentation in Q2, and selecting characteristic product ions in Q3, thereby improving quantification accuracy and reproducibility. Total peak area normalization was applied to eliminate systematic errors, followed by retention of metabolites exhibiting a coefficient of variation (CV) < 30% in quality control (QC) samples[13] for subsequent analyses. Multivariate statistical analyses, including principal component analysis (PCA), partial least squares-discriminant analysis (PLS-DA), and orthogonal PLS-DA (OPLS-DA), were performed using the Ropls package (v1.22.0) in R to reduce dimensionality and discriminate metabolic profiles among sample groups. Pearson correlation coefficients between samples were calculated via the cor function (v4.0.3) in R. To identify differentially abundant metabolites, statistical significance (p-value), variable importance in projection (VIP) scores from OPLS-DA, and fold change (FC) values were computed. Metabolites meeting the criteria of p-value < 0.05 and VIP > 1 were considered statistically significant. Trend analysis was subsequently performed using the online platform Bioinformatics (
www.bioinformatics.com.cn ). Kyoto Encyclopedia of Genes and Genomes (KEGG,www.genome.jp/kegg ) pathway enrichment analysis was conducted on the differential metabolites.Quantification of selected metabolites by UPLC-MS/MS
-
Selected compounds were quantified using UPLC-MS/MS (Sciex 4500). Frozen plant samples were ground into fine powder under liquid nitrogen. Approximately 200 mg of sample was extracted with 1 mL of 80% methanol containing 0.1% butylated hydroxytoluene (BHT) and 0.1% formic acid. After ultrasonic extraction and overnight incubation at 4 °C, the supernatant was centrifuged and filtered through a 0.22 μm membrane.
Chromatographic separation was performed on a HYPERSIL GOLD C18 column (3 μm, 2.1 mm × 100 mm) at 35 °C with a flow rate of 0.3 mL/min. The mobile phases consisted of 0.1% formic acid in water (A) and acetonitrile (B), with a 10 min gradient program. Detection was carried out using electrospray ionization in both positive and negative modes under MRM acquisition. Compound identification was confirmed using authentic standards, and external standard curves were used to calculate concentrations based on peak areas.
16S rRNA sequencing and analysis
-
The microbial community analysis of dandelion tea samples involves DNA extraction from centrifuged pellets, followed by quantification using Nanodrop (Thermo Fisher Scientific, USA) and quality assessment via 1.2% agarose gel electrophoresis. Target regions of the 16S rRNA gene are amplified using barcoded primers, with subsequent purification and quantification of PCR products using the Quant-iT PicoGreen dsDNA Assay Kit (Invitrogen, Thermo Fisher Scientific, USA) measured on a BioTek FLx800 microplate reader (BioTek Instruments, USA). Sequencing libraries are prepared using Illumina TruSeq Nano DNA LT Library Prep Kit (Illumina, USA) and sequenced on the Illumina Novaseq 6,000 (Illumina, USA) for microbial analysis.
High-throughput raw sequencing data were initially screened for sequence quality. Passing sequences were assigned to libraries and samples according to index and barcode information, with barcode sequences removed. Sequence denoising or OTU clustering was performed following the QIIME2 software (v2019.4) pipeline. Taxonomic composition at various classification levels was analyzed to understand the overall microbial profile. Alpha diversity was evaluated for each sample based on ASV/OTU distribution, and rarefaction curves were generated to assess sequencing depth adequacy. At the ASV/OTU level, distance matrices were computed, and beta diversity differences between samples/groups were assessed using unsupervised clustering, ordination methods, and statistical tests. PICRUSt2 (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States) was employed to predict microbial metabolic functions, identify differential pathways, and determine the species contributing to specific pathways.
-
To investigate the effects of prolonged brewing on the bioactive compounds and microbial communities in dandelion tea, 5 g of dried dandelion leaves were brewed in 500 mL of boiling water (100 °C) and subjected to long-duration treatments with brewing times of 1, 12, 24, and 36 h (Fig. 1a, b). The tea infusion brewed for 1 h exhibited a deep color with high transparency and clarity. After 12 h of brewing, the tea color darkened slightly but remained clear and transparent. However, when brewed for 24 h (overnight), the tea color significantly lightened and the infusion became turbid with a notable decrease in transparency. After 36 h of brewing, the tea color further lightened, and turbidity increased, resulting in an almost opaque appearance (Fig. 1c, d).

Figure 1.
Comparison of tea infusion colors and sampling at different brewing times. (a), (b) Tea infusions after brewing for 1, 12, 24, and 36 h. (c) Comparison of tea infusion colors after brewing for 1, 12, 24, and 36 h. (d) Sampling of tea infusions after brewing for 1, 12, 24, and 36 h for metabolomic analysis.
For metabolomic analysis, 20 mL aliquots of tea infusions brewed for 1, 12, 24, and 36 h (labeled H1, H12, H24, and H36, respectively) were sampled. The remaining infusions were centrifuged, and the resulting pellets were collected for microbial community analysis (Fig. 1e, f).
Metabolomic analysis of tea infusions at different brewing times
-
To investigate the effects of different steeping durations on the metabolite composition of dandelion tea, LC-MS-based metabolomic profiling was conducted across all groups. As shown in the correlation matrix (Fig. 2a), strong intra-group correlations were observed (Pearson correlation coefficient > 0.98), indicating high experimental reproducibility and stable metabolic profiles. In contrast, inter-group correlations declined with increasing differences in steeping time, suggesting that brewing duration significantly affected the overall metabolomic composition.
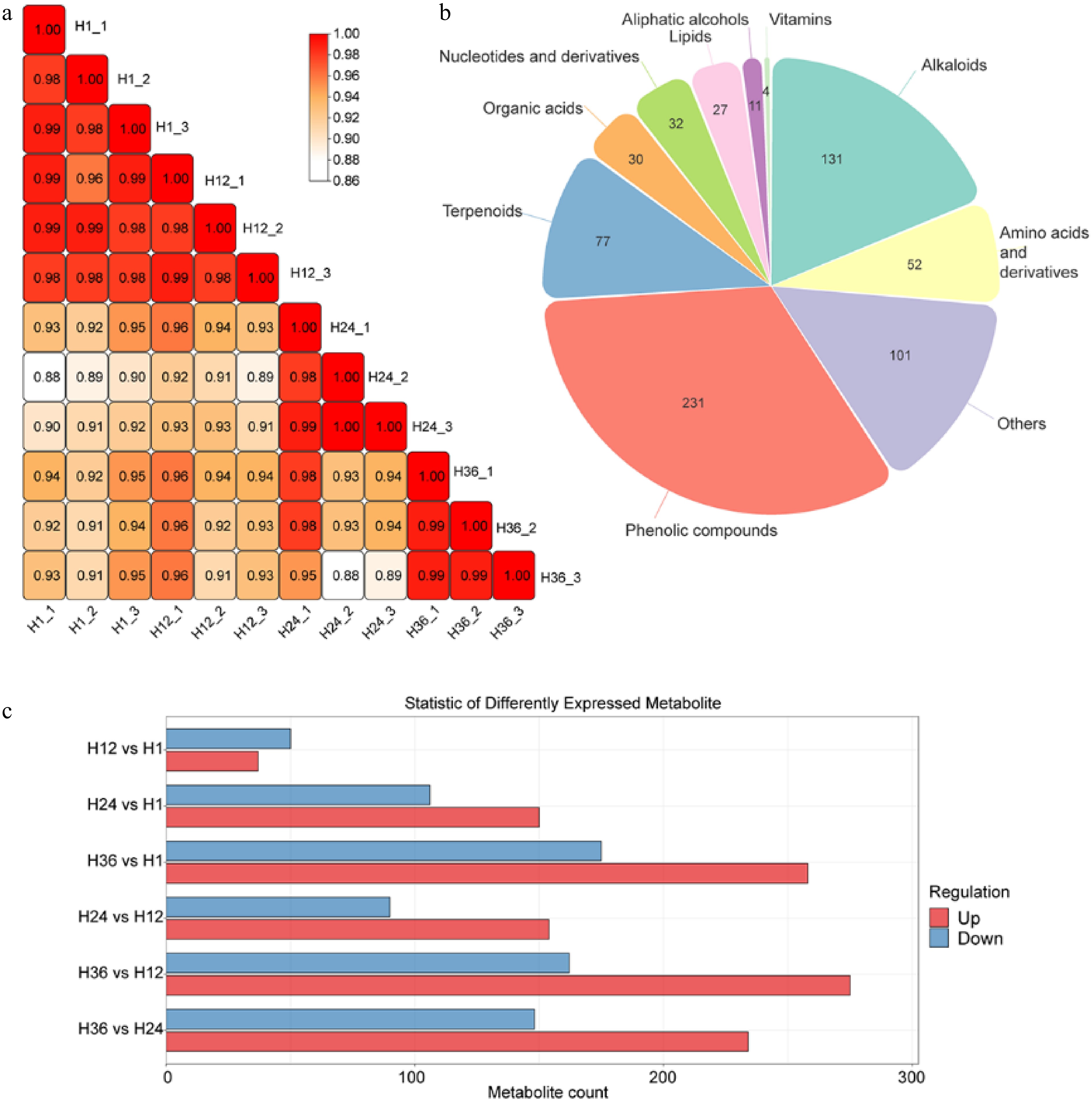
Figure 2.
Metabolomic profiling and comparative analysis of dandelion tea samples across steeping durations. (a) Pearson correlation matrix showing high within-group reproducibility and increasing divergence across groups with longer steeping times. (b) Pie chart representing the classification and number of identified metabolites. (c) Bar graph showing the number of differentially expressed metabolites (DEMs) between sample groups. Red and blue bars represent upregulated and downregulated metabolites, respectively.
Metabolite classification results (Fig. 2b) showed a total of 696 identified putative metabolites (Supplementary Table S1), primarily comprising phenolic compounds (231), alkaloids (131), terpenoids and flavonoids (77), amino acids and their derivatives (52), and organic acids (30). Additional minor constituents included nucleotides, lipids, vitamins, and aliphatic alcohols. Among them, phenolic compounds accounted for the largest proportion, representing the major functional foundation of dandelion tea's health-promoting properties.
Differential metabolite analysis (Fig. 2c) showed that steeping duration markedly altered the metabolic expression profile. Compared to the H1 group, only 87 differentially expressed metabolites (DEMs) were identified in the H12 group (37 upregulated, 50 downregulated). In contrast, 256 DEMs (150 upregulated/106 downregulated) and 433 DEMs (258 upregulated/175 downregulated) were identified in the H24 and H36 groups, respectively, indicating a progressive intensification of metabolite changes with prolonged brewing. Notably, upregulated metabolites substantially outnumbered downregulated ones, implying that certain heat-stable or water-soluble components may be increasingly released or enriched during long-term steeping. In addition, there are 244, 437, and 382 DEMs in the H24 vs H12, H36 vs H12, and H36 vs H24 groups, respectively.
Detection and comparison of secondary metabolites in overnight tea infusions
-
To comprehensively characterize the time-dependent metabolic dynamics of dandelion tea during different steeping durations, Mfuzz time-series clustering was performed on 562 DEMs, resulting in eight distinct expression trend clusters (Cluster 1–8) (Fig. 3a; Supplementary Table S2). Among them, Clusters 1, 2, and 6 represented metabolite groups that were continuously upregulated over time and showed marked enrichment after 12 h of steeping (overnight). These clusters collectively contained 282 metabolites, including phenolic compounds (70), terpenoids (47), organic acids, alkaloids, and other components. KEGG enrichment analysis (Fig. 3b) showed that these upregulated metabolites were predominantly involved in aromatic amino acid metabolism (e.g., phenylalanine, tyrosine, and tryptophan), alkaloid biosynthesis, ascorbate and aldarate metabolism, and the TCA cycle, suggesting their potential contributions to enhanced antioxidant capacity and flavor enrichment in the tea infusion during prolonged steeping.
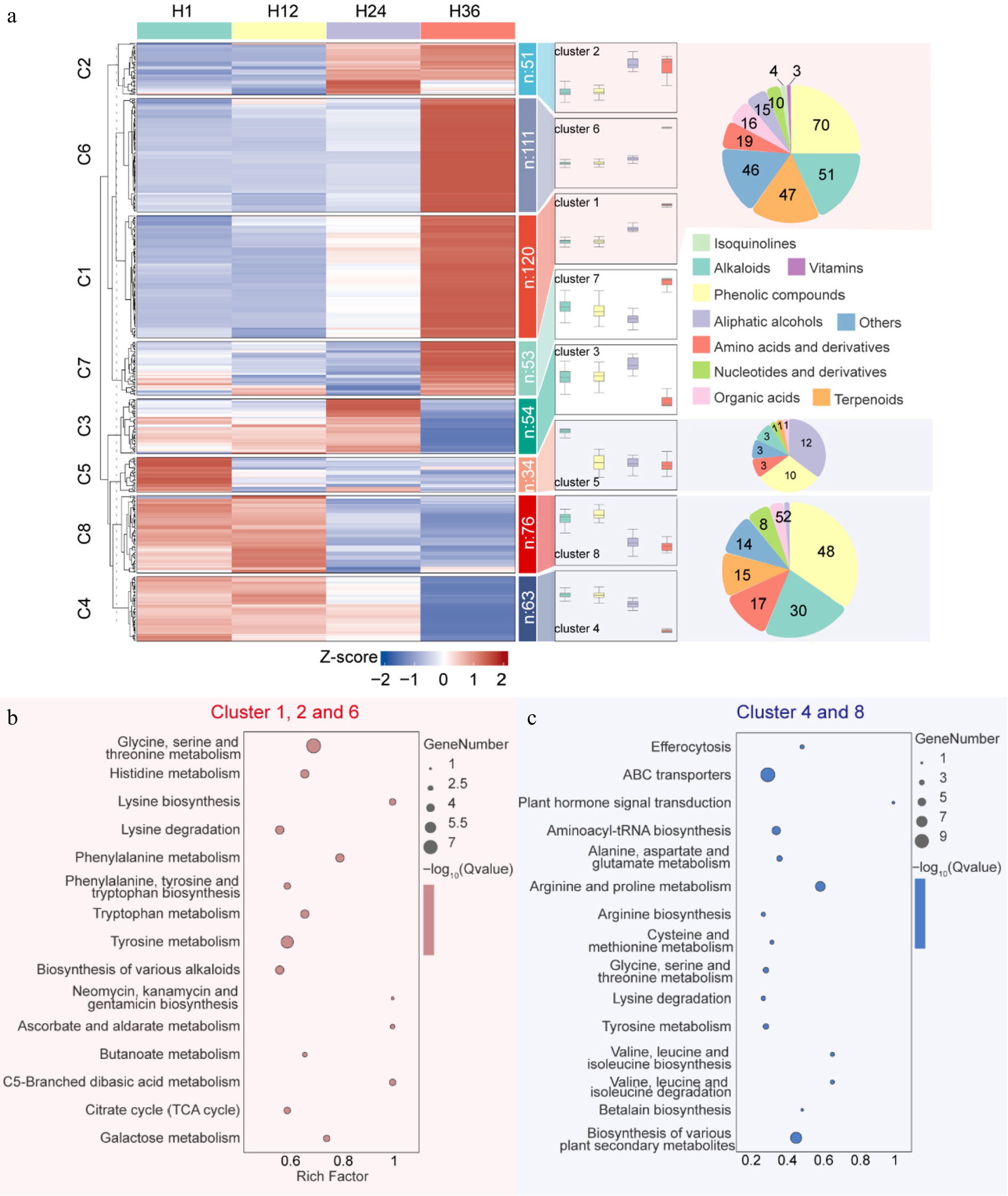
Figure 3.
Trend clustering and KEGG pathway enrichment of differentially expressed metabolites in dandelion tea across steeping durations. (a) Mfuzz time-series clustering of 696 metabolites across four steeping durations (H1, H12, H24, H36), grouped into eight dynamic expression clusters. Boxplots to the right show expression trends per cluster, while pie charts illustrate metabolite class composition. (b) KEGG pathway enrichment of upregulated metabolites in Clusters 1, 2, and 6. (c) KEGG enrichment of downregulated metabolites in Clusters 4 and 8.
In contrast, Cluster 5 represents a metabolite module that significantly downregulated after 1 h, mainly including 12 alcohols, ten phenolic compounds, etc.; Clusters 4 and 8 represent metabolite modules that significantly downregulated overnight (12 h), collectively containing 139 metabolites, predominantly phenolic compounds, alkaloids, amino acids and their derivatives, etc. These substances decreased significantly after 12 h. The corresponding KEGG analysis showed that these downregulated metabolites were enriched in pathways such as amino acid biosynthesis, plant hormone signal transduction, tRNA biosynthesis, and secondary metabolite biosynthesis (Fig. 3c), indicating that small molecules with nutritional value or regulatory effects in the tea infusion gradually leached out with prolonged brewing time, potentially affecting its overall functionality and nutritional health properties.
Analysis of key upregulated and downregulated metabolites
-
Dandelion tea contains a wide array of major metabolites, including flavonoids, terpenoids, phenolic acids, and amino acids (along with their derivatives). To systematically characterize the dynamic changes in these compounds under different steeping durations, heatmap-based analyses were performed focusing on the upregulated and downregulated metabolite clusters. Among flavonoids, 20 compounds were significantly upregulated at 36 h (H36), including naringenin-5-methylether, dihydrokaempferol, kaempferol, daidzein-8-C-glucoside, and sophoricoside. Conversely, another subset of flavonoids exhibited a clear downward trend during prolonged steeping, such as eriodictyol-7-O-glucoside, naringenin, luteolin, apigenin, quercetin-3-O-xyloside, reynoutrin, and tectorigenin, which are typically associated with strong antioxidant properties (Fig. 4a).
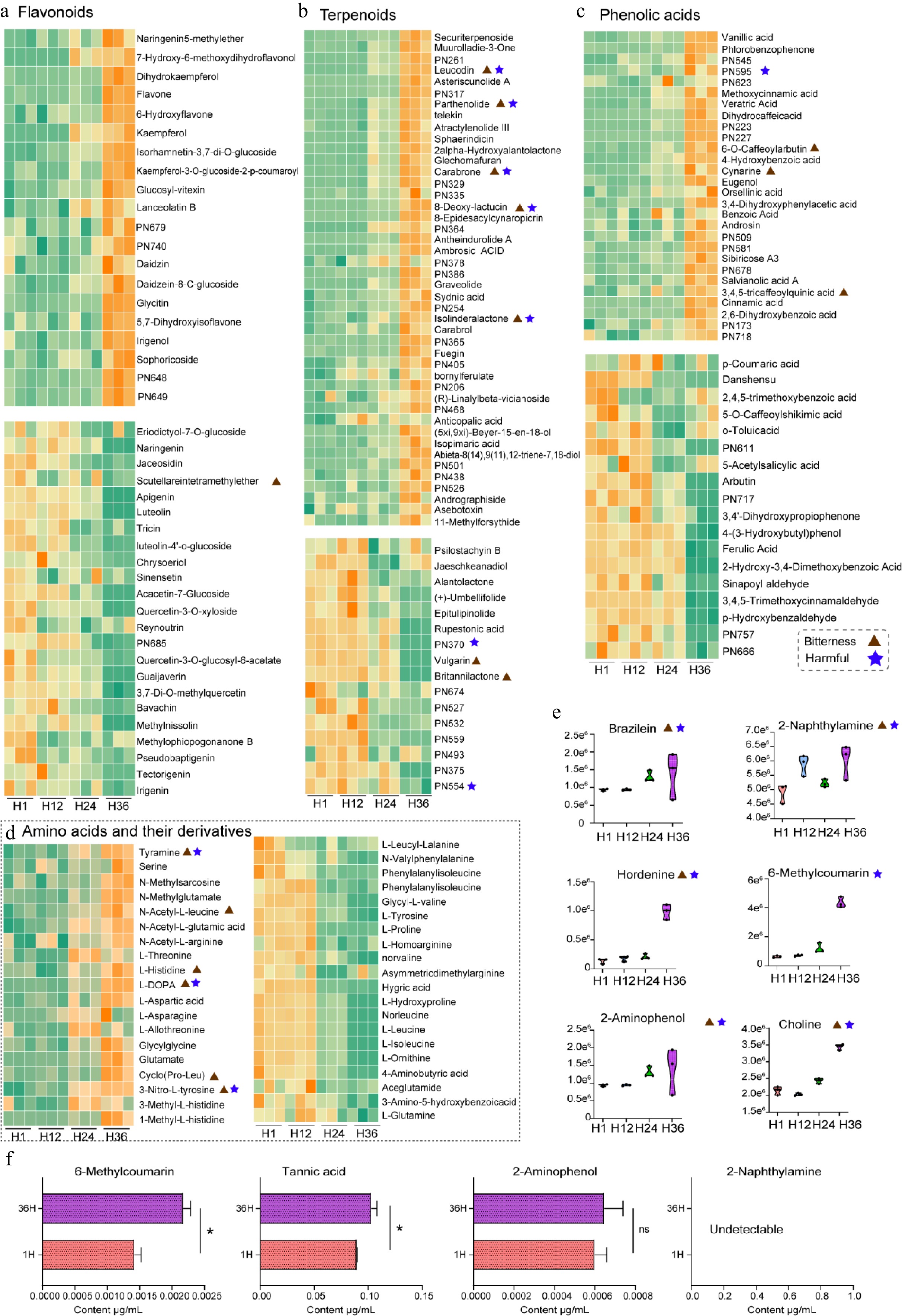
Figure 4.
Temporal variation of key metabolite classes in dandelion tea under different steeping durations (H1, H12, H24, H36). (a)–(d) Heatmaps display the relative abundance of representative metabolites across different steeping time points, classified into four major categories. (a) Flavonoids, (b) terpenoids, (c) phenolic acids, and (d) amino acids and their derivatives. Orange indicates relatively high abundance, while green denotes low abundance. Each row was individually normalized. (e) Violin plots depict the temporal accumulation patterns of selected harmful or bitter metabolites. (f) Quantitative UPLC-MS/MS analysis of selected compounds in dandelion tea infusions after 1 and 36 h of brewing. * p < 0.05.
With respect to terpenoids, a total of 45 compounds showed significant upregulation in the late steeping stages, while only 16 decreased (Fig. 4b). This suggests that extended steeping facilitates the release or stabilization of certain terpenoids. Notably, several of the accumulated terpenoids, such as leucodin, parthenolide, carabrone, 8-deoxy-lactucin, and isolinderalacton, are reported in the literature to possess high bioactivity but are also associated with bitter taste and potential toxicity.
For phenolic acids, 28 compounds were upregulated and 18 downregulated at later steeping stages (Fig. 4c). The accumulated phenolics included various bitter-tasting compounds such as 6-O-caffeoylarbutin, cynarine, and 3,4,5-tricaffeoylquinic acid. The downregulated subset was primarily composed of hydroxycinnamic acids, benzoic acid derivatives, phenolic aldehydes, phenols, and their glycosides, including p-coumaric acid, ferulic acid, danshensu, arbutin, 5-acetylsalicylic acid, and sinapoyl aldehyde.
Among amino acids and their derivatives, 19 were significantly upregulated and 20 downregulated (Fig. 4d). Notably, tyramine, L-DOPA, and 3-nitro-L-tyrosine—each of which increased significantly—have been linked to potential neurotoxicity and inflammatory responses. Furthermore, tyramine, L-histidine, L-DOPA, and cyclo (Pro-Leu) have been reported to contribute to bitter or umami-bitter flavor characteristics, suggesting that prolonged steeping may result in a shift from a fresh to a more bitter sensory profile. Conversely, the downregulated amino acids, including L-leucine, L-isoleucine, L-proline, L-tyrosine, and L-glutamine, are important for both nutritional quality and flavor; their depletion may diminish the health value and sensory appeal of the tea.
Additionally, violin plots showed that some other metabolites, including 2-naphthylamine, 2-aminophenol, hordenine, and 6-methylcoumarin, accumulated significantly at H24 and H36 compared to H1 and H12 (Fig. 4e). Based on further validation by quantitative UPLC-MS/MS analysis, several potentially harmful compounds exhibited changes in abundance during extended brewing. As shown in Fig. 4f, the level of 6-methylcoumarin increased from approximately 0.0014 µg/mL (1H) to 0.0021 µg/mL (36H); tannic acid increased from 0.089 µg/mL (1H) to 0.102 µg/mL (36H). Both showed statistically significant differences (*p < 0.05), suggesting accumulation over time. In contrast, 2-aminophenol was detected at comparable concentrations in both groups, with no significant difference. 2-naphthylamine was undetectable in both infusion conditions. These compounds have been documented as toxic or potentially carcinogenic, raising concerns about food safety risks associated with long-term steeping of dandelion tea.
Detection and comparison of microorganisms in overnight tea infusions
-
High-throughput 16S rRNA sequencing revealed that different steeping durations significantly influenced the taxonomic composition and diversity of microbial communities in dandelion tea. As shown in Fig. 5a, the number of taxonomic units across six hierarchical levels (from phylum to species) was highest in the H1 group, indicating the greatest microbial richness at the early stage of infusion (Supplementary Table S3). With prolonged steeping, the number of taxa gradually declined, reflecting a marked reduction in microbial diversity.
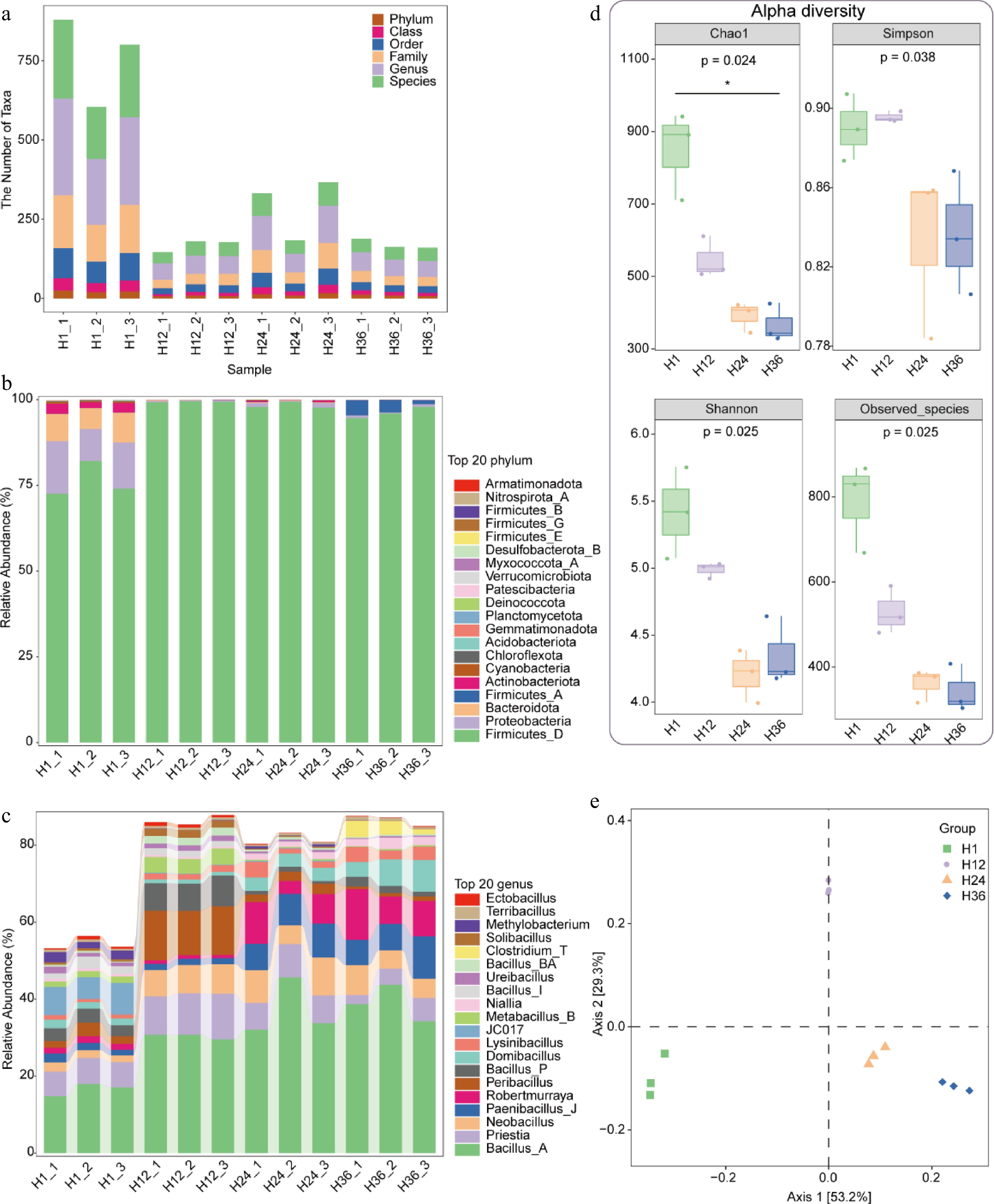
Figure 5.
Taxonomic composition and diversity of microbial communities in dandelion tea under different steeping durations. (a) Number of taxonomic units identified at six hierarchical levels (phylum, class, order, family, genus, and species) across samples steeped for 1 h (H1), 12 h (H12), 24 h (H24), and 36 h (H36). (b) Relative abundance of the top 20 bacterial phyla identified in tea infusions across different steeping durations. (c) Relative abundance of the top 20 bacterial genera. (d) Alpha diversity indices (Chao1, Simpson, Shannon, and Observed_species) of microbial communities in tea infusions. (e) Principal coordinate analysis (PCoA) based on Bray–Curtis distance demonstrates clear separation among samples from different steeping groups.
At the phylum level (Fig. 5b), Firmicutes_D, Proteobacteria, Bacteroidota, and Actinobacteriota were the dominant phyla in the H1 group. As steeping time increased, Firmicutes_D became progressively enriched in the H12, H24, and H36 groups, suggesting its ecological advantage during extended or overnight brewing. Notably, Firmicutes_A showed a significant increase in relative abundance in the H36 group. Genus-level analysis (Fig. 5c) further revealed that Bacillus, Robertmurraya, and Neobacillus were strongly enriched over time, indicating selective proliferation of specific bacterial genera under long-term steeping and room-temperature storage conditions. However, due to the limited resolution of the V3–V4 region of 16S rRNA sequencing, species-level identification was not feasible.
Alpha diversity indices (Fig. 5d) corroborated these findings. The Chao1, Shannon, and Observed species indices all declined significantly with increasing steeping time (p < 0.05), indicating a reduction in microbial richness, compositional complexity, and community evenness. Similarly, the Simpson index decreased, reflecting the emergence of dominant genera and a shift toward community homogenization. Principal coordinate analysis (PCoA) (Fig. 5e) further demonstrated clear separation among the different steeping groups along Axis 1 and Axis 2, with tight clustering within groups. These results confirm that steeping duration is a major factor shaping the structure and diversity of microbial communities in dandelion tea.
To further investigate the impact of prolonged steeping on the functional potential of microbial communities in dandelion tea, KEGG pathway enrichment analysis was performed based on predicted microbial functions (Fig. 6; Supplementary Table S4). Cluster analysis revealed that microbial functional profiles underwent substantial dynamic remodeling with increasing steeping time. Functional modules represented by C6, C3, and C4 exhibited a decline over time, with enriched pathways primarily related to terpenoid and polyketide metabolism, cofactor and vitamin metabolism, lipid metabolism, and pathways associated with human diseases. These results suggest that extended steeping may reduce the functional activity of microbial taxa responsible for the biosynthesis of secondary metabolites, vitamins, and lipids, thereby disrupting the homeostasis of bioactive compounds in the tea infusion and potentially weakening its health-promoting properties. In contrast, C1 and C8 modules represented functions that showed an increasing trend with prolonged steeping, mainly associated with carbohydrate metabolism and amino acid metabolism, reflecting basic metabolic activities. This observation implies that, although the capacity for the biosynthesis of functional compounds declines, fundamental energy metabolism activities are either maintained or selectively activated.
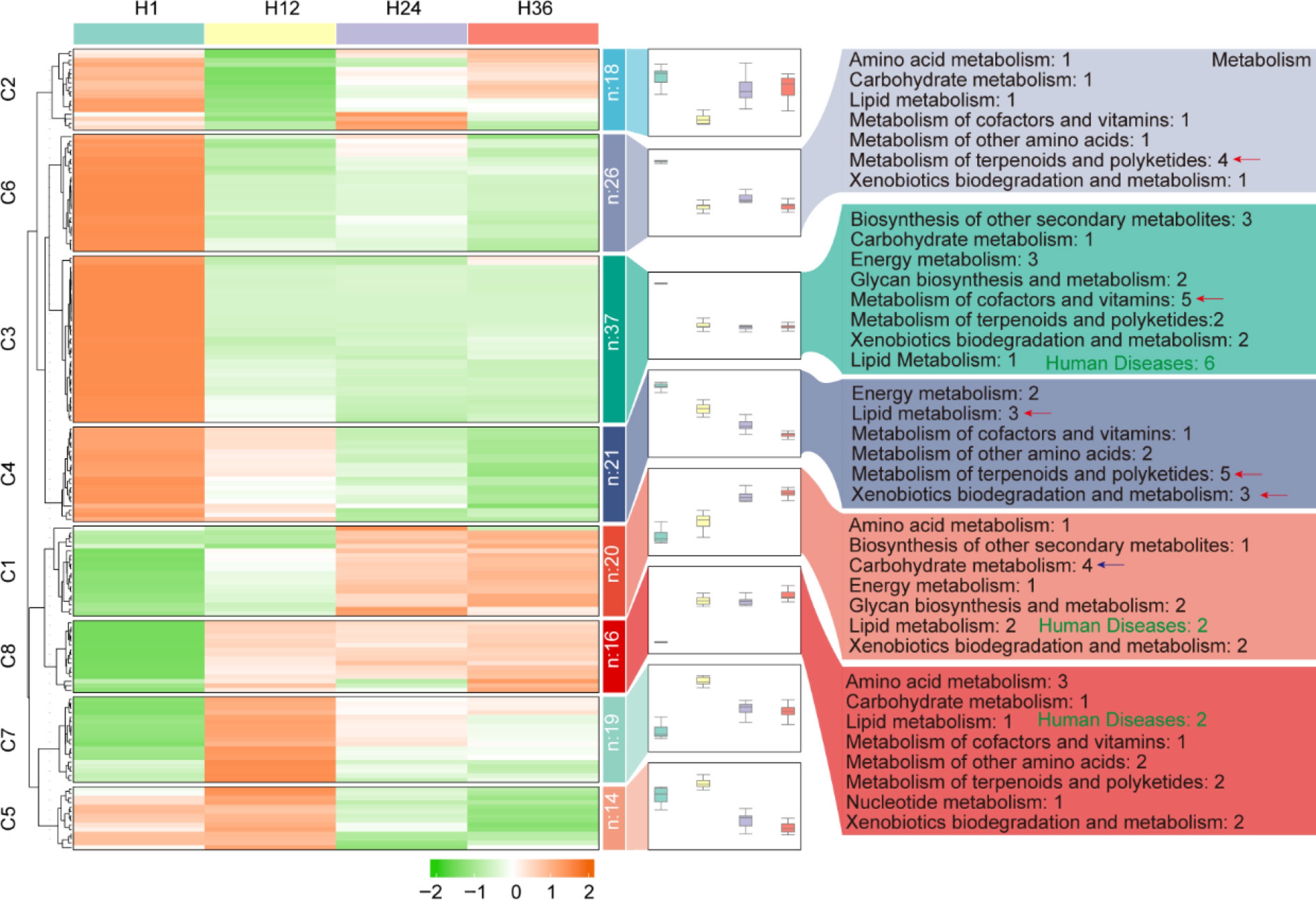
Figure 6.
KEGG functional pathway enrichment of predicted microbial communities across different steeping durations in dandelion tea. Heatmap clustering of microbial functional pathways across H1, H12, H24, and H36 samples. The right panel shows the number of pathways per functional group for each cluster.
Correlation analysis between metabolites and microbial communities
-
To investigate the potential associations between changes in microbial communities and the accumulation or degradation of metabolites during different steeping durations, a correlation network was constructed based on Spearman analysis (Fig. 7), with node sizes adjusted according to the degree value to reflect the relative importance of each element within the network. Overall, the analysis showed that upregulated metabolites were predominantly negatively correlated with microbial abundance. Notably, PN315 (Leucodin), PN193 (Hymecromone methyl ether), PN405 (1,2-Cyclohexanediol benzoate derivative), PN114 (6-methylcoumarin), PN364 ([4aR,9aR]-6,6-Dimethyl-pentaleno-pyran carboxylic acid), PN66 (Tyramine), PN162 (Tricarballylic acid), PN699 (D-[+]-Raffinose), and PN400 (2',6'-Dihydroxy-4'-methoxydihydrochalcone) exhibited high degree values, showing significant negative correlations with microbial taxa such as Firmicutes_D and Proteobacteria (Fig. 7a).
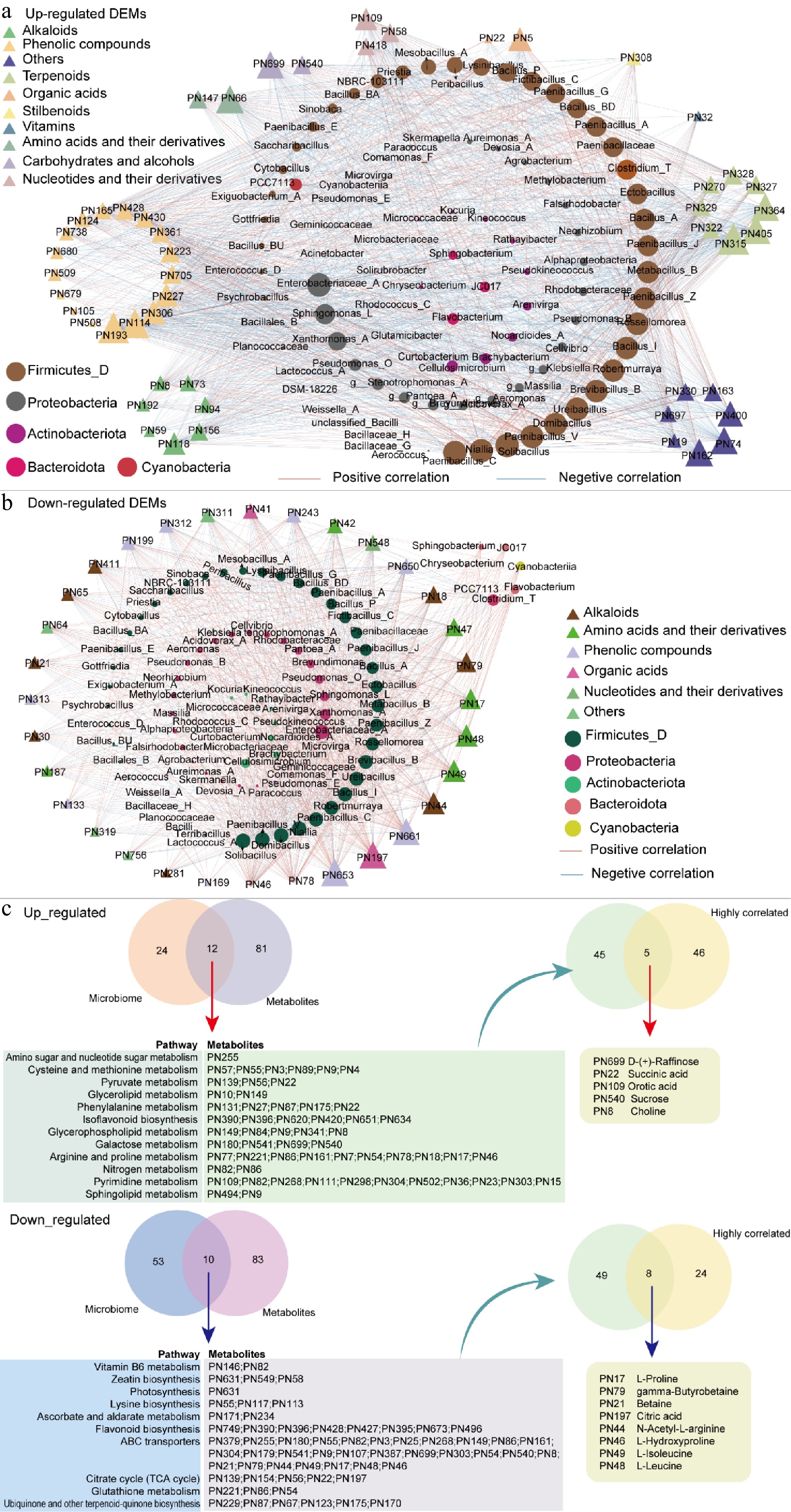
Figure 7.
Correlation network analysis between metabolites and microbial taxa during different steeping durations. (a) Network analysis showing the correlations between upregulated metabolites and microbial genera. (b) Network analysis focusing on downregulated metabolites and their associations with microbial taxa. Node size reflects the degree value, with larger nodes indicating higher connectivity and greater importance within the network. Red and blue edges represent positive and negative correlations, respectively. (c) Overlapping KEGG pathways and metabolites between the microbiome and metabolome (left). Five and eight metabolites showing strong correlations with microbial taxa (as in (a), (b), respectively).
Conversely, downregulated metabolites predominantly exhibited positive correlations with microbial taxa (Fig. 7b). Key metabolites, including PN46 (L-Hydroxyproline), PN78 (4-Guanidinobutanoic acid), PN653 (Luteolin-4'-O-glucoside), PN197 (Citric acid), PN661 (Aromatic ketone derivative), PN44 (5-Aminolevulinic acid), PN49 (L-Isoleucine), PN48 (L-Leucine), PN17 (L-Proline), and PN79 (gamma-Butyrobetaine), also showed high degree values.
To investigate the potential contribution of microbial metabolic activity to changes in the tea metabolome, microbiome-derived KEGG functional predictions (via PICRUSt2) were cross-referenced with metabolomic pathway enrichment results, and a set of shared metabolic pathways was identified (Fig. 7c, left). Differentially abundant metabolites involved in these shared pathways were further intersected with the highly correlated metabolites (|r| > 0.8) identified in Fig. 7a, b. As a result, five up-regulated candidate metabolites—D-(+)-raffinose, succinic acid, orotic acid, sucrose, and choline—were identified, which are not only significantly elevated during extended steeping but also showed strong correlations with microbial taxa (Fig. 7c). Similarly, from the overlapping downregulated pathways, eight highly correlated downregulated metabolites were identified, most of which were amino acids or their derivatives, including L-proline, L-isoleucine, L-leucine, N-acetyl-L-arginine, and gamma-butyrobetaine (Fig. 7c).
-
Dandelion leaves are rich in nutrients, such as proteins, fatty acids, amino acids, vitamins, and trace minerals, as well as functional components, including polysaccharides, flavonoids, and phenolic acids. A total of 24 compounds have been isolated from the aerial parts of dandelion, comprising six flavonoids, four sesquiterpenes, two sphingolipids, six glycerolipids, and six triterpenes/sterols[3,14]. Despite this phytochemical diversity, studies on the composition of dandelion leaf tea infusions remain limited.
In this study, metabolomic analysis identified 696 metabolites in the dandelion tea infusion, predominantly phenolic compounds (231), alkaloids (131), flavonoids/terpenoids (77), amino acids and derivatives (52), and organic acids (30), alongside trace functional components such as nucleotides, lipids, vitamins, and sterols. Major compounds included kaempferol, rutin, and glycosylated flavonoids, as well as amino acids like proline, leucine, and isoleucine. Notably, amino acids associated with the umami taste of tea[15,16], including theanine, glutamate, glutamine, and aspartate, were also detected. The infusion contains bitter-tasting phenolic acids (e.g., caffeic acid, chlorogenic acid) and sesquiterpene lactones (e.g., carabrone, 8-deoxy-lactucin), which impart sensory similarities to green or white teas but with a markedly bitter profile. While dandelion leaf extracts (DLE) exhibit anti-obesity potential as natural preventive agents[1,17], and root-derived triterpenes demonstrate anti-ageing effects via modulation of human dermal fibroblast proliferation[18], only one triterpenoid was detected in the tea infusion. This simplified composition, in contrast to the chemical complexity of concentrated extracts, highlights the tea's suitability as a daily functional beverage with moderate bioactive potency.
Time-dependent metabolite dynamics in dandelion tea
-
The quality of tea infusion is influenced by factors such as water temperature, water quality, and steeping time, among which steeping time plays an especially important role. Studies have found that there is a positive correlation between steeping time and the concentration of substances in tea infusion. The longer the steeping time, the higher the concentration of substances in the tea infusion, but the extraction rate per unit time decreases. In the initial stage of tea steeping (1–2 min), substances such as tea polyphenols, amino acids, caffeine, and water-soluble polysaccharides quickly dissolve into the tea infusion. With extended steeping, the extraction concentration of high-content substances such as tea polyphenols and caffeine in the tea infusion is higher than that of amino acids, thereby increasing the polyphenol-to-amino acid ratio and making the bitterness and astringency more pronounced[19,20]. Extended steeping time enhances bioactive compound extraction: gallic acid, epigallocatechin gallate, and caffeine levels rise significantly after 10 min, correlating with increased antioxidant capacity in white tea[6]. Similarly, black tea exhibits elevated phenolic compounds (e.g., phenolic compounds such as catechins, flavonoids, and phenolic acids) under prolonged high-temperature steeping, enhancing antioxidant activity[5]. Extending brewing time from 0 to 10 min increases total tea polyphenols, flavonoids, vitamin C, soluble sugars, catechins, and flavonoid glycosides in the infusion[21].
To evaluate changes in 'overnight tea' and address questions about its potential effects on human health, steeping time was extended to 36 h. Metabolomic analysis revealed a significant increase in 20 flavonoid substances (e.g., kaempferol) in the 36H group, while luteolin, naringenin, apigenin, and others declined. Among 45 terpenoids, most (e.g., leucodin, parthenolide) increased with prolonged steeping. These compounds exhibit anti-inflammatory and antioxidant properties but may also cause skin or gastrointestinal irritation[22−24]. Carabrone and 8-deoxy-lactucin, which are bitter-tasting sesquiterpenes, have been used as indicators of plant bitterness[25], and isolinderalactone has anti-tumor and antibacterial activities but also exhibits neurotoxicity and cytotoxicity[26,27]. These indicate compounds accumulated, intensifying bitterness and degrading flavor in prolonged steeping tea. Concurrently, amino acids and their derivatives (e.g., L-leucine, L-isoleucine, L-proline, L-tyrosine, and L-glutamine) declined, reducing the nutritional value and taste of dandelion tea. Notably, substances such as 2-naphthylamine, 2-aminophenol, hordenine, and 6-methylcoumarin were found in higher amounts in the H24 and H36 groups than in the H1 and H12 groups. These compounds, documented as toxic or potentially carcinogenic, adhere to strict safety thresholds such as OEHHA, Proposition 65, and EU Council Directive 88/388/EEC: 2-naphthylamine (< 0.4 µg/d, ideally non-detectable), 2-aminophenol (≤ 0.01 mg/kg under the total aromatic amines limit), and 6-methylcoumarin (≤ 2 mg/kg as a flavoring additive). In this study, all these analytes were undetectable or present at trace levels in samples brewed for ≤ 12 h, suggesting minimal safety risk under typical preparation conditions. However, prolonged steeping (36 h) resulted in measurable accumulation of several potentially harmful compounds. For example, 6-methylcoumarin reached ~2.1 mg/L, exceeding its regulatory threshold. These substances may be toxic or carcinogenic[28,29]. Thus, while prolonged steeping enriches certain beneficial compounds, it also amplifies bitterness and harmful substances, rendering overnight dandelion tea inadvisable for consumption. These findings suggest that short-term brewing (≤ 12 h) preserves nutritional quality and microbial safety. Therefore, it is recommended that dandelion tea be consumed within 12 h of brewing. For 12–24 h preparations, strict refrigeration at 4 °C is required to prevent amino acid degradation, microbial proliferation, and enzymatic toxin conversion. Particularly, steeping beyond 24 h generates toxicologically significant compounds and should be avoided.
Brewing duration governs microbial diversity, metabolic activity, and safety in dandelion tea
-
Microbial metabolic activities during tea processing critically influence flavor profiles, color characteristics, product variety, and safety parameters. In black tea, a symbiotic fermentation system involving acetic acid bacteria and yeast communities generates diverse metabolites (e.g., alcohols, aldehydes, acids, and esters) that define kombucha's sensory quality and health properties[30]). Pu-erh tea fermentation is dominated by bacterial communities such as Erwinia, Achromobacter, Bacillus, and Staphylococcus[31], with Aspergillus niger driving initial fungal fermentation via polysaccharide metabolism[32]. Later stages favor thermophilic species (e.g., Bacillus, Arxula adeninivorans), which accelerate fermentation and enhance quality[12,31]). As a distinct non-fermented tea, dandelion tea's microbial composition remains unexplored. This study investigated microbial community dynamics in dandelion tea across brewing durations using 16S rRNA sequencing. The results demonstrated a progressive decline in microbial diversity with extended brewing time, peaking in richness during short-term infusions (0–1 h) and reaching minimal levels at 36 h. Short-term brewing was dominated by Firmicutes_D, Proteobacteria, Bacteroidota, and Actinobacteriota, with Proteobacteria (including pathogenic strains such as Escherichia coli and Salmonella) alongside beneficial taxa. Prolonged brewing enriched thermotolerant genera within Firmicutes_D, particularly Bacillus_A, Robertmurraya, and Neobacillus. These Bacillus-associated species, recognized as functional probiotics, produce flavor-enhancing enzymes (e.g., polyphenol oxidases) and modulate gut microbiota and immunity[31]. It should be noted that the 16S rRNA sequencing approach does not allow definitive species-level resolution within the Bacillus genus. As such, the dominance of Bacillus spp. during prolonged steeping cannot be conclusively attributed to either beneficial strains (e.g., B. subtilis) or potentially pathogenic ones (e.g., B. cereus). This taxonomic ambiguity warrants cautious interpretation, and future studies employing full-length 16S or metagenomic sequencing are needed to more accurately assess species composition and associated food safety risks.
Integrated metabolomic-microbiome analysis linked extended brewing to diminished microbial activity in critical pathways, including terpenoid/polyketide metabolism, cofactor/vitamin synthesis, and lipid metabolism. This functional decline suggests impaired biosynthesis of bioactive compounds such as secondary metabolites and vitamins, potentially compromising the tea's health-promoting efficacy. To further investigate this connection, KEGG pathway overlaps between the microbiome and metabolome were integrated, and specific up- and down-regulated metabolites associated with microbial activity. Notably, D-(+)-raffinose, succinic acid, orotic acid, sucrose, and choline were significantly elevated and are commonly linked to microbial carbohydrate conversion and fermentation[33−35]. In contrast, amino acids such as L-leucine, L-isoleucine, N-acetyl-L-arginine, and gamma-butyrobetaine showed reduced abundance and are likely depleted due to microbial uptake and catabolism[36]. These findings suggest that prolonged brewing not only restructures the microbial community but also reshapes the metabolite landscape via microbial transformation and nutrient competition.
In addition, overnight infusions exhibited accumulation of harmful substances, which were inversely correlated with microbial viability, suggesting reduced microbial capacity for detoxification or metabolic conversion. Notably, amino acid content positively correlated with microbial activity, underscoring microbial roles in beneficial compound formation. These findings collectively highlight that brewing duration serves as a key determinant of dandelion tea's microbial ecology and functional outcomes. While short-term infusions maintain a balance between microbial diversity and bioactive compound production, prolonged brewing favors thermotolerant Bacillus dominance at the expense of metabolic efficacy, ultimately affecting safety and health attributes.
-
This study demonstrates that prolonged brewing (> 24 h) significantly alters the metabolic and microbial profiles of dandelion tea. Metabolomic analysis revealed progressive shifts in composition: while heat-stable compounds (e.g., specific flavonoids, terpenoids) accumulated over time, critical nutrients (e.g., amino acids, antioxidative phenolics) declined, accompanied by the release of potentially harmful metabolites (e.g., 2-naphthylamine, leucodin). Concurrently, 16S rRNA sequencing indicated reduced microbial diversity and functional activity, with dominance of Bacillus genera under extended steeping. Microbial functional decline correlated with disrupted biosynthesis of vitamins, lipids, and secondary metabolites, diminishing the tea's health-promoting properties. Critically, metabolite-microbe correlation networks highlighted negative associations between enriched toxins and microbial viability. These findings establish that dandelion tea should not be steeped for more than 12 h at room temperature (22–25 °C); in particular, steeping for longer than 24 h should be avoided. Future studies should optimize brewing parameters to balance bioactive compound retention and microbial safety.
-
The author confirms contributions to the paper as follows: conceptualization, writing and editing: Jin Y, Lin C; sampling, data analysis: Jin Y. All authors reviewed and approved the final version of the manuscript.
-
The raw sequence data reported in this paper have been deposited in the Genome Sequence Archive in the National Genomics Data Center, China National Center for Bioinformation/Beijing Institute of Genomics, Chinese Academy of Sciences (GSA: CRA026169) and are publicly accessible at https://ngdc.cncb.ac.cn/gsa. All data generated or analyzed during this study are included in this published article and its supplementary information files, further inquiries re available from the corresponding author on reasonable request.
-
We thank Suzhou Panomix Biomedical Technology Co., Ltd. for conducting the metabolomic and 16S rRNA sequencing. We acknowledge Shandong Guocangjian Biotechnology Co.,Ltd. for their technical assistance in performing the UPLC-MS/MS analyses.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 696 putative metabolites identified in dandelion tea.
- Supplementary Table S2 Differential metabolites identified across distinct expression modules.
- Supplementary Table S3 Summary table of microbial taxonomic units at each classification level.
- Supplementary Table S4 KEGG functional pathway enrichment analysis of bacterial communities across different expression modules.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Jin Y, Lin C. 2025. Safe to steep? Temporal shifts in microbial viability and functional metabolites define the safety of prolonged dandelion tea. Medicinal Plant Biology 4: e029 doi: 10.48130/mpb-0025-0029 

Safe to steep? Temporal shifts in microbial viability and functional metabolites define the safety of prolonged dandelion tea
- Received: 18 July 2025
- Revised: 01 August 2025
- Accepted: 11 August 2025
- Published online: 29 August 2025
Abstract: Dandelion (Taraxacum officinale) tea possesses significant nutritional and pharmacological value, but the impact of brewing time on its chemical stability and microbial safety remains unclear. This study investigated changes in chemical composition, antioxidant capacity, and microbial community structure in dandelion tea under different brewing durations (1, 12, 24, and 36 h) using metabolomic analysis and 16S rRNA microbiome sequencing. The results showed that prolonged brewing significantly altered the metabolic profile of dandelion tea, with certain heat-stable or water-soluble components being increasingly released or enriched. However, extended brewing led to the loss of nutritional components, a decline in flavor quality, and the potential release of harmful substances. Microbial analysis revealed a decrease in microbial diversity with longer brewing times, with specific bacterial genera (such as Bacillus) becoming dominant. Long-term brewing may reduce the functional activity of microbes responsible for the biosynthesis of secondary metabolites, vitamins, and lipids, thereby disrupting the balance of bioactive compounds and potentially weakening the health-promoting properties of the tea. Therefore, it is recommended to consume dandelion tea brewed for less than 12 h to ensure its nutritional value and safety.
-
Key words:
- Taraxacum officinale /
- Secondary metabolites /
- Metabolomics /
- 16S rRNA sequencing /
- Medicinal plant