









-
Apple (Malus × domestica Borkh.) is a perennial deciduous fruit tree of the Rosaceae family, widely cultivated across temperate regions worldwide. Apples are economically significant and valued for their high nutritional content. According to the Food and Agriculture Organization (FAO), global apple production reached 86 million tons in 2020, with China accounting for 47% of this output, followed by the United States and Turkey as secondary producers. A typical raw apple comprises approximately 86% water and 14% carbohydrates, with minimal fat and protein content. Apple consumption is associated with numerous health benefits, including immune system support, cholesterol reduction, decreased diabetes risk, cardiovascular and cognitive health improvements, weight management, digestive regulation, and enhanced bone health.
Like other fruits such as pears, cherries, and plums, apples are enveloped by a peel composed of the cuticle, epidermis, and multiple layers of hypodermis[1−3]. The fruit peel is pivotal in determining overall fruit quality, influencing attributes such as appearance, texture, flavor, and shelf life. It also serves as the primary barrier against environmental stresses and pathogen invasion[4]. Peel thickness, in particular, affects consumer preference, mechanical resistance, and postharvest water loss[5,6]. While thicker peels generally confer greater resistance to physical damage and dehydration, thinner peels often correspond to higher sensory quality[7].
The peel's multi-layered structure plays critical roles in minimizing transpiration, preventing pathogen penetration, and shielding the fruit from mechanical injury, all while facilitating gaseous exchange[4,6]. Advances in imaging technologies—such as light microscopy (LM), scanning electron microscopy (SEM), and confocal laser scanning microscopy (CLSM)—have greatly enhanced our understanding of peel morphology and surface features like wax layers and microcracks in apples and other fruits[8−13].
Peel development is a tightly coordinated process involving cuticle deposition, cell wall reinforcement, and cellular differentiation[14−16]. These processes are regulated by a complex interplay of transcriptional networks, metabolic activities, hormone signaling pathways, and environmental cues[15]. Plant hormones such as auxin, gibberellin (GA), and abscisic acid (ABA) play crucial roles in orchestrating peel growth by modulating epidermal cell expansion, cuticle biosynthesis, and stress responses. Specifically, auxin and GA promote epidermal cell growth and cuticle formation[17,18], while ABA enhances cuticle integrity under abiotic stress by regulating cutin and wax biosynthesis[19]. Jasmonic acid (JA) and salicylic acid (SA) further influence cuticle properties and pathogen defense, contributing to peel resilience[20].
Transport proteins, particularly ABC transporters, have been implicated in cuticular lipid export and peel integrity in species such as tomato, and likely perform the same functions in apples[21]. The biosynthesis of phenylpropanoids and waxes is fundamental in determining cuticle composition and characteristics, impacting water retention and resistance to diseases[4]. Additionally, transcription factors like MdMYB93 regulate suberin deposition in russeted apple peels, thereby influencing peel structure, integrity, and functionality[22]. Flavonoids and other phenolic compounds also contribute to peel development and adaptation to environmental stresses[22,23].
Despite advances in the genetic and molecular understanding of apple fruit development, the mechanisms underlying natural variation in peel thickness remain poorly characterized. Most prior research has focused on cuticle and epicuticular wax composition[1−3], with few integrative studies addressing morphological, physiological, and transcriptomic factors that drive peel thickness variation.
In this study, a thin-peel apple mutant compared to a thick-peel cultivar was examined. By integrating physiological measurements, transcriptomic profiling, and morphological analyses, the aim was to elucidate the molecular, biochemical, and structural determinants of peel thickness. The present findings provide valuable insights into the gene regulatory networks and metabolic pathways underlying peel development, offering potential targets for genetic improvement of fruit texture, flavor, and postharvest performance in apple breeding programs.
-
Fruit samples were collected from an apple orchard in Liquan County, Xianyang City, Shaanxi Province, China, located at 108°27′40″ E, 34°50′02″ N. A natural short-spur mutant (sport) was discovered on a 21-year-old 'Fuji'/'Malus sieversii' apple tree in this orchard. The mutant was asexually propagated and grafted onto M9T337 dwarfing rootstocks. For the experiment, six uniform trees, with consistent growth vigor, were selected from both the mutant and cultivar-type groups. From each tree, four fruits were systematically sampled—two from the middle canopy on the north side and two from the south side, ensuring representative sampling. This sampling strategy yielded a total of 24 fruits per genotype (six trees × four fruits each). For subsequent biochemical analyses, composite samples were prepared by mixing peels from these collected fruits to ensure sufficient material and reduce individual fruit variability.
Microstructural analysis of peel
-
Peel discs (1 cm diameter × 0.5 mm thickness) were excised from the equatorial region of mutant and cultivar-type apples using a sterile cork borer, and immediately frozen in liquid nitrogen to preserve cellular ultrastructure. For cross-sectional analysis, frozen samples were cryo-fractured under liquid nitrogen, sputter-coated with 10 nm gold/palladium (Q150T ES Plus, Quorum Technologies), and imaged using two SEM systems: (1) Nano SEM-450 (FEI, Thermo Fisher Scientific, USA) at 5 kV for peel architecture; and (2) FlexSEM 1000 (Hitachi High-Tech, Japan) at 5 kV for high-resolution thickness measurements. Surface morphology was analyzed using intact peel samples air-dried at 25 °C, coated with gold/palladium, and imaged with the FlexSEM 1,000 at 5 kV. Peel thickness was quantified from ≥ 10 random locations per sample using Map3D software (Hitachi). The description can be found in the previous study by Li et al.[24].
Determination of structural components (NREL method)
-
Apple peels were thinly sliced (0.1–0.3 mm thickness) using a precision peeling knife, then subjected to enzyme deactivation at 105 °C for 30 min to prevent biochemical degradation. Samples were subsequently dried at 60 °C until a constant weight was achieved to ensure complete moisture removal. The dried material was ground using a laboratory mill and sieved through a 60-mesh screen to obtain homogeneous samples for analysis. Compositional analysis of cellulose, hemicellulose, and lignin was performed according to the NREL standard method[25] using UHPLC (Aminex HPX-87P column, 85 °C, 0.6 mL/min water mobile phase).
Fruit physical and quality attribute measurements
-
Fruit firmness was measured using a texture analyzer (TA.XT Plus, Stable Micro Systems, Surrey, UK) equipped with a 30 kg load cell, and a 2 mm diameter stainless steel puncture probe. The probe penetrated the fruit at 5 mm/s to a depth of 7 mm, with measurements taken on opposite sides (stem and calyx regions) after peel removal. Six biological replicates per genotype (mutant and cultivar-type) were analyzed, and firmness was recorded as the maximum force (N) required for puncture.
Transverse and longitudinal fruit diameters were determined using a vernier caliper, with five measurements per dimension at different angles, and the average was calculated. Soluble solids content (SSC, %) was assessed using an Atago PAL-1 digital pocket refractometer (Japan); juice from four fruit parts was extracted, and measurements were averaged. Data are presented as mean ± standard deviation (SD). Differences between the mutant-type and cultivar-type were assessed for statistical significance using an independent samples t-test in SPSS. A significance level of **p < 0.01 was applied.
RNA extraction and library preparation
-
Total RNA was extracted from apple peels of both mutant and cultivar-type varieties using Plant RNA Purification Reagent, followed by genomic DNA removal with Takara's rDNase-free treatment. RNA purity was verified using an ND-2000 spectrophotometer (NanoDrop Technologies), with acceptable samples meeting the criteria of OD260/280 = 1.8–2.2 and OD260/230 ≥ 2.0. RNA integrity was assessed using a 2100 Bioanalyzer (Agilent Technologies), where samples with RNA Integrity Number (RIN) ≥ 6.5 and 28S:18S ratio ≥ 1.0 were considered high-quality. Only RNA samples exceeding 10 μg were used for sequencing library construction, which was performed according to the protocol described by Fan et al.[26].
Differential gene expression analysis
-
Differentially expressed genes (DEGs) between groups were identified using the DESeq2 package, which employs a negative binomial distribution model to statistically evaluate gene expression differences. Additionally, the Benjamini-Hochberg method was employed to control the false discovery rate (FDR) by adjusting the p-values. DEGs were identified from the mutant-type and cultivated-type combination, applying thresholds of |log2(FoldChange)| ≥ 1 and padj ≤ 0.05. Principal Component Analysis (PCA) was performed on variance-stabilized transformed count data using the DESeq2 package, based on the 500 most variable genes. Heatmaps were generated in GraphPad Prism (v10.0.0) to visualize the log2 fold change values of the significant DEGs.
Functional enrichment analysis
-
Gene Ontology (GO) enrichment analysis was conducted using the clusterProfiler package, implementing the Wallenius non-central hypergeometric distribution to correct for gene length bias[27]. Additionally, the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was performed using the KEGG database to elucidate the biological functions and metabolic pathways associated with the identified DEGs[28]. The KEGG analysis leveraged its integrated molecular datasets to provide system-level insights into the gene functions (
www.genome.jp/kegg ). -
Visual inspection revealed pronounced phenotypic differences between the mutant and cultivar-type apples (Fig. 1). The mutant exhibited a thin-skinned peel with a distinct yellowish-red base coloration, overlaid with irregular red striations. Additionally, the surface displayed a high density of small, yellow lenticels (Fig. 1a). In contrast, the cultivar-type apple had a thicker peel with a more uniform reddish color and only faint yellow undertones. While lenticels were present, their distribution was limited, resulting in a smoother and more visually uniform peel surface compared to the mutant (Fig. 1b).
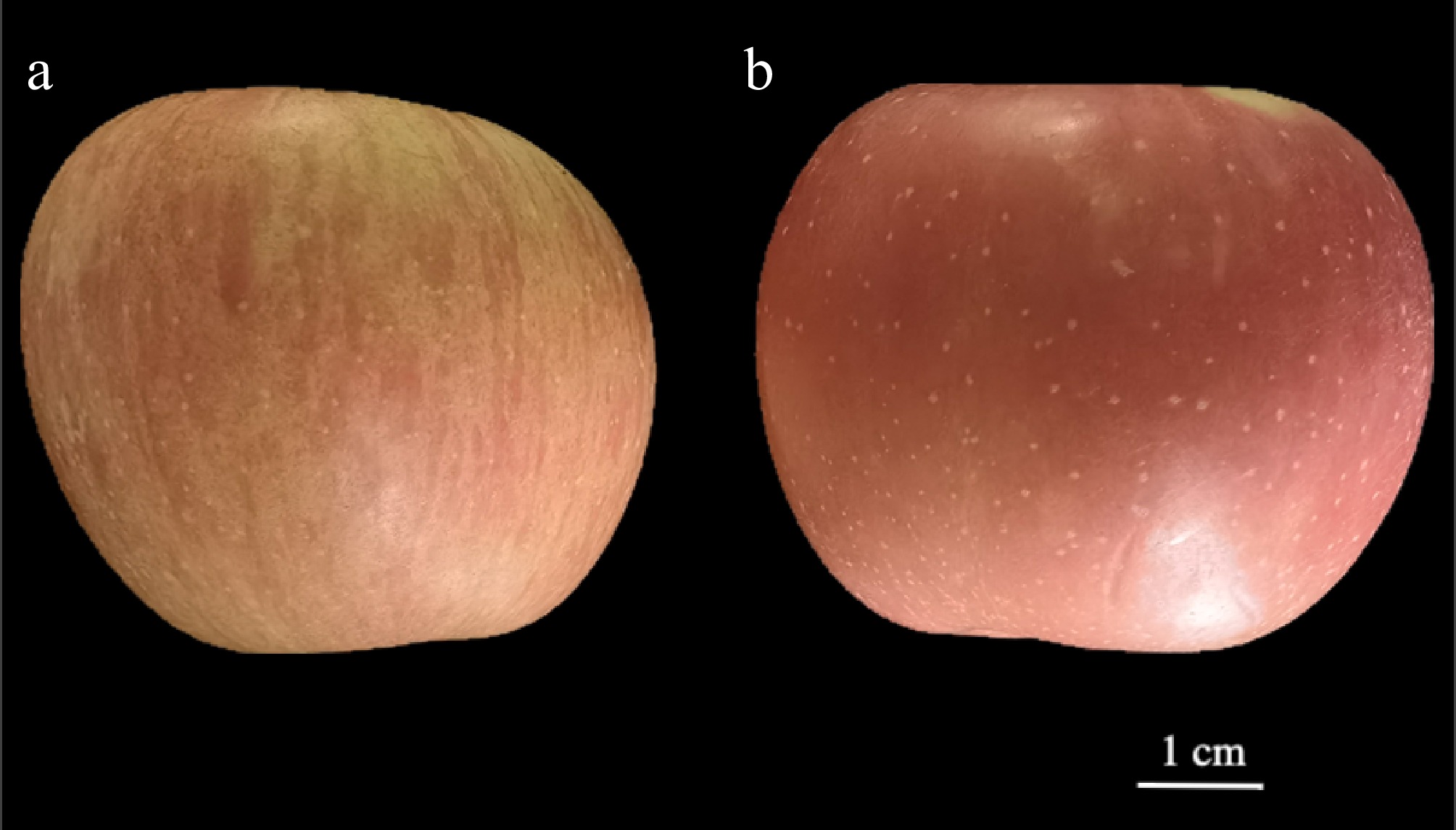
Figure 1.
Peel morphology of (a) mutant, and (b) cultivar-type apples. Scale bar = 1 cm.
Morphological analysis of apple peel cells
-
To investigate the differences in peel structure between mutant and cultivar-type apples, SEM was utilized, analyzing cross-sectional and surface characteristics (Fig. 2a–f). Cross-sectional SEM images (Fig. 2a, b) revealed distinct differences in peel architecture. The mutant apple (Fig. 2a) exhibited a thinner, more porous peel with loosely arranged cell layers, while the cultivar-type apple (Fig. 2b) showed a thicker peel composed of densely packed epidermal and subepidermal tissues. These anatomical differences suggest a structural basis for the textural and storability variations observed between the two apple types. Quantitative analysis of peel thickness using the FlexSEM1000 (Fig. 2c, d) further supported these findings. The mutant peel measured approximately 18.0–20.2 µm in thickness, while the cultivar peel ranged from 31.3–33.7 µm, indicating nearly a 1.7-fold increase in thickness for the cultivar apple. This confirms the presence of a thin-peel phenotype in the mutant variety.

Figure 2.
Scanning electron microscopy (SEM) analysis of peel microstructure in mutant and cultivar apples. (a), (c), (e) SEM images of the mutant apple peel, showing both cross-sectional and surface structures at varying magnifications. The mutant type exhibits a thinner epidermal layer with a relatively smooth cuticle and less densely packed cell layers. (b), (d), (f) SEM images of the cultivar apple peel, revealing a thicker epidermal region with more compact cell layers and a more complex cuticular surface. The cultivar peel shows more prominent ridges and cuticle folds compared to the mutant type. Scale bars are indicated in each panel.
Surface microstructure and cuticle integrity
-
Surface SEM imaging at higher magnification revealed additional epidermal differences (Fig. 2e, f). The mutant apple peel (Fig. 2e) displayed a smoother surface, with fewer cuticular irregularities and reduced microcracking. In contrast, the cultivar-type peel (Fig. 2f) exhibited a rougher texture, with pronounced striations and numerous microcracks. Together, these observations confirm the thinner peel phenotype in the mutant apple and provide a microstructural basis for further physiological and genetic studies on peel development and its role in postharvest quality.
Physiological characteristics of mutant and cultivar apples
-
To further assess the fruit quality and structural composition of the two apple types, several physiological indicators were meaured, including size parameters, soluble solid content, firmness, and cell wall composition (Table 1). The mutant apple exhibited a significantly lower fruit size ratio (transverse to longitudinal diameter) compared to the cultivar, indicating a more rounded shape. While the soluble solids content was slightly higher in the mutant, this difference was not statistically significant. Conversely, the mutant fruit firmness was significantly lower than that of the cultivar, consistent with the observed softer texture (Table 1).
Table 1. Physiological characteristics of mutant-type and cultivar-type apple peel.
Investigated features Mutant type Cultivar type Fruit transverse and longitudinal diameters 1.12 ± 0.13 1.25 ± 0.09** Soluble solids content (%) 13.1 ± 0.6 12.7 ± 0.9 Firmness 3.46 ± 0.18 3.63 ± 0.22** Cellulose (%) 24.75 ± 0.25 25.05 ± 0.18** Acid-insoluble lignin (%) 21.20 ± 0.99 22.65 ± 0.49** Acid-soluble lignin (%) 1.90 ± 0.11 1.80 ± 0.08** Hemicellulose (%) 7.75 ± 0.07 7.80 ± 0.14** Data are mean ± SD (n = 24). ** Indicates a significant difference between genotypes at p < 0.01 (independent samples t-test). Analysis of cell wall composition revealed that the mutant apple had significantly lower levels of cellulose, acid-insoluble lignin, and hemicellulose, indicating substantially reduced structural rigidity. The mutant also had a significantly higher content of acid-soluble lignin (Table 1). These findings demonstrate that the mutant apple has a distinct physiological profile, characterized by a significantly softer texture and a comprehensive alteration of its cell wall composition. In contrast, the cultivar apple's greater firmness and structural fiber content suggest superior mechanical strength and postharvest durability.
Global transcriptomic profiling and differential gene expression between mutant and cultivated apple peels
-
To investigate the molecular mechanisms underlying peel thickness variation between mutant- and cultivar-type apples, mRNA sequencing analysis was performed. All samples yielded high-quality data, generating 42.30–52.61 million raw reads, with 41.50–51.77 million clean reads retained after quality filtering (1%–2% read loss), demonstrating excellent data integrity (Supplementary Table S1). Sequencing data quality exceeded standard thresholds, with Q20 > 98.45% and Q30 > 96.05% across all libraries, ensuring reliable downstream analysis (Supplementary Table S1). Read alignment metrics revealed robust mapping efficiency, with 82.55%–89.72% of reads successfully mapped to the reference genome (Supplementary Table S2). Notably, the vast majority of mapped reads (80.18%–87.10%) aligned uniquely, while a small fraction (2.37%–3.28%) exhibited multiple mappings, indicating minimal interference from repetitive genomic regions (Supplementary Table S2). The consistently high mapping rates and balanced unique/multi-mapping ratios across all samples confirm the suitability of these data for differential gene expression analysis.
Transcriptomic analysis revealed significant differences in gene expression between mutant-type (MT) and cultivated-type (CT) apple peels. PCA indicated a clear separation between MT and CT samples along the first principal component (PC1), which accounted for 50.42% of the total variance (Fig. 3a). This finding highlights distinct global transcriptomic profiles for the two groups. Fragments per kilobase of transcript per million mapped fragments (FPKM) density plots were generated to assess expression value distributions for each sample, showing consistent and comparable expression ranges across all biological replicates. This consistency indicates effective normalization and high-quality RNA-seq data (Fig. 3b).
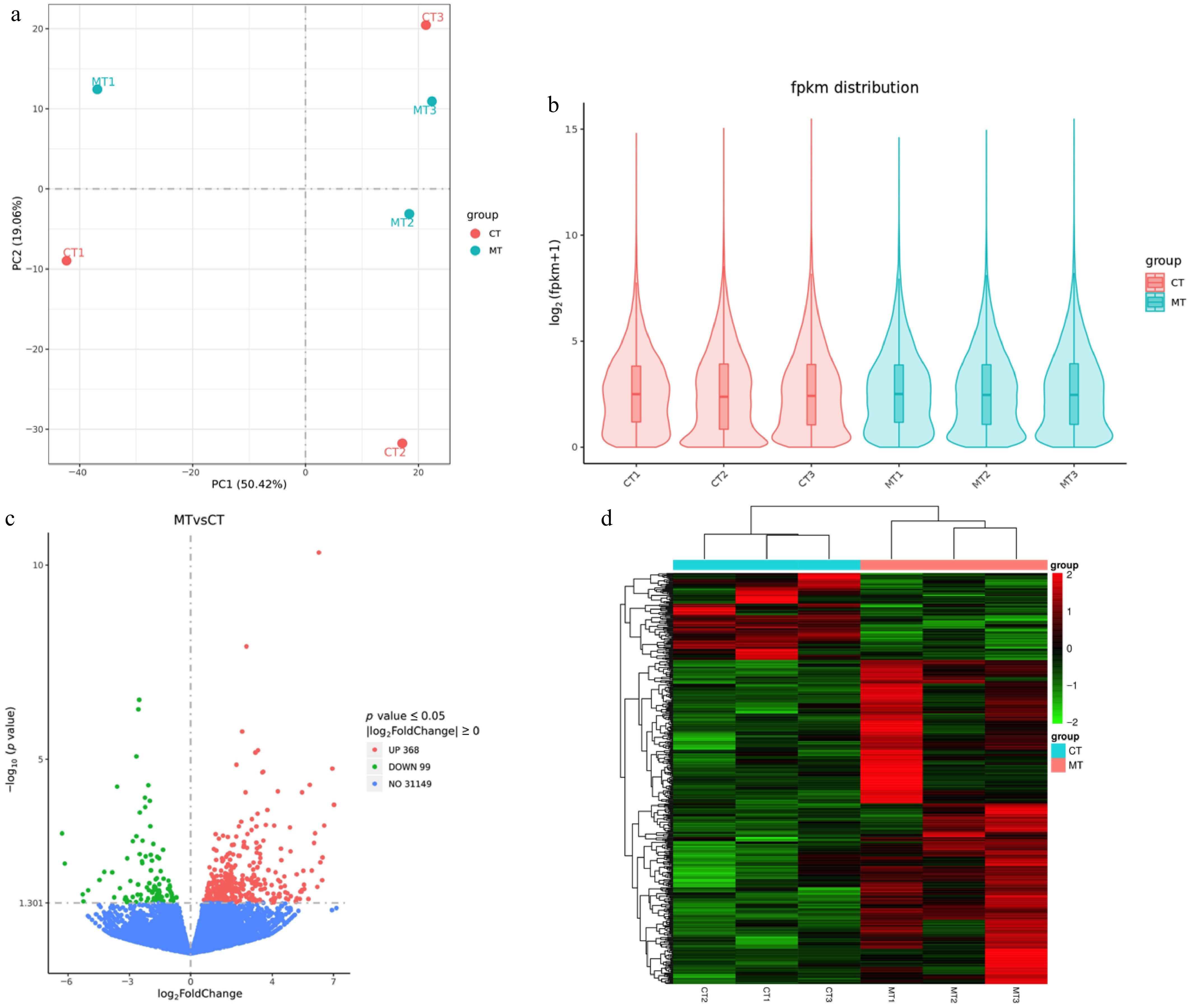
Figure 3.
Transcriptomic analysis of mutant-type (MT) and cultivated-type (CT) apple peels. (a) Principal component analysis (PCA) showing distinct clustering of MT and CT samples. (b) Violin plots displaying the distribution of fragments per kilobase of transcript per million mapped fragments (FPKM) values across all samples. (c) Volcano plot of differentially expressed genes (DEGs) between MT and CT groups. Red represents upregulated, green represents downregulated, and blue represents non-significant genes. (d) Heatmap of the DEGs, showing hierarchical clustering of expression profiles. Red indicates high expression, and green indicates low expression.
Differential expression analysis identified a total of 467 DEGs (FDR < 0.05, |log2FC| > 1), with 368 genes upregulated and 99 downregulated in MT compared to CT samples (Fig. 3c). The predominance of upregulated genes in MT suggests the activation of specific transcriptional programs associated with the mutant phenotype. Additionally, hierarchical clustering and heatmap visualization of DEGs revealed two major expression patterns (Fig. 3d): a large cluster of genes consistently upregulated in MT samples (approximately 78.8% of DEGs) and a smaller cluster enriched in CT samples (approximately 21.2% of DEGs). The clear segregation between MT and CT expression profiles further supports the PCA findings, emphasizing the robustness and reproducibility of group-specific transcriptomic signatures.
Functional enrichment analysis of DEGs
-
GO and KEGG were utilized to identify the biological processes and functions enriched in DEGs (Fig. 4a, b). GO enrichment analysis revealed significant enrichment of genes associated with metal ion transport, ion transport, and cation transport, underscoring their critical role in maintaining nutrient and ion homeostasis in apple peel, likely supporting cellular integrity and stress adaptation. Enriched terms related to the superoxide metabolic process and reactive oxygen species metabolic process suggest robust oxidative stress management mechanisms, consistent with the peel's function as a protective barrier against environmental challenges. Furthermore, the enrichment of transferase activities transferring glycosyl and hexosyl groups implies their involvement in cell wall modification and cuticle biosynthesis, processes essential for peel development, structural reinforcement, and pathogen defense (Fig. 4a).
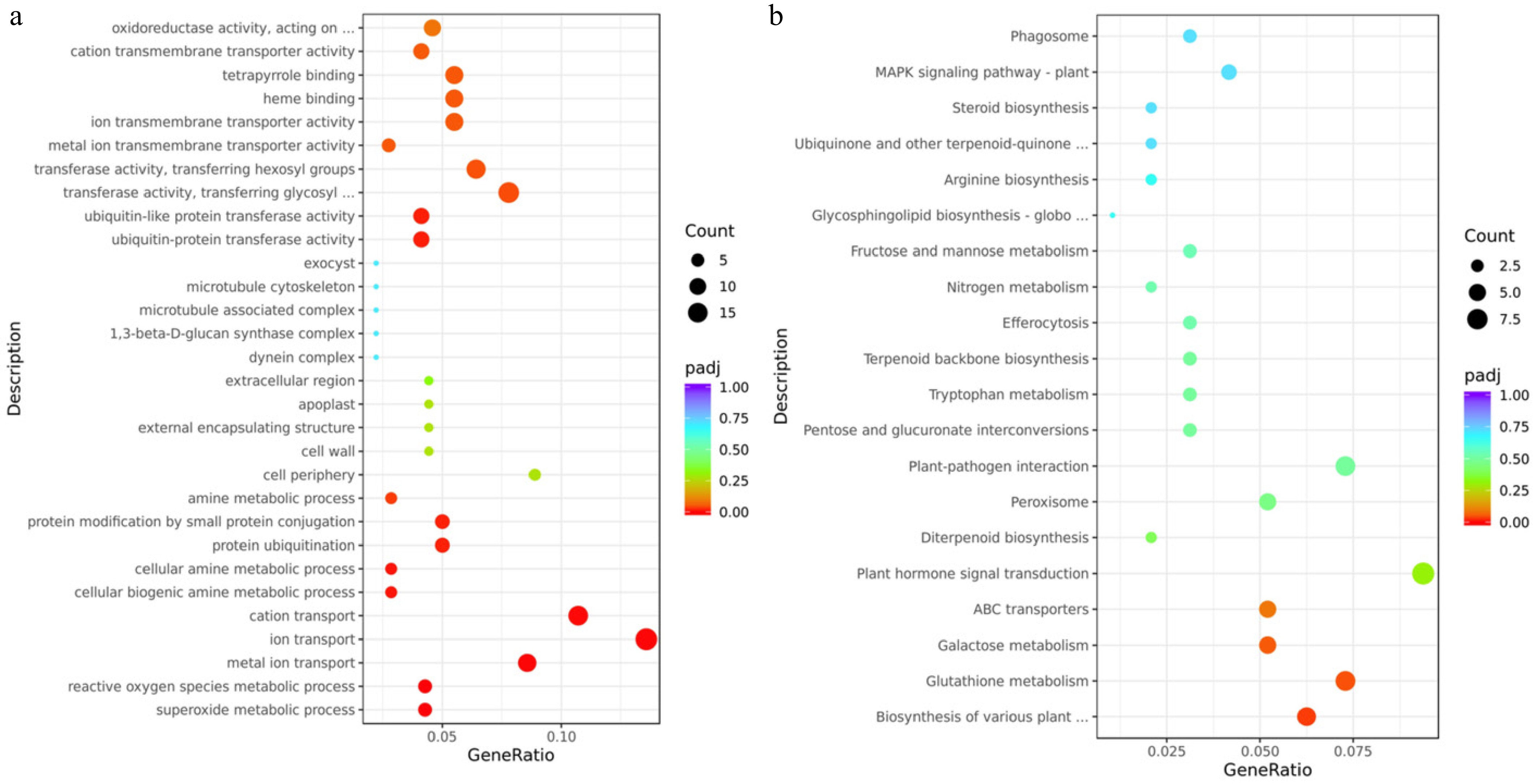
Figure 4.
Transcriptomic analysis of mutant-type (MT) and cultivated-type (CT) apple peels. (a) Gene Ontology (GO) enrichment analysis, and (b) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of differentially expressed genes (DEGs) between MT and CT peels. The vertical axis displays the names of enriched GO terms or KEGG pathways. The size of each dot represents the number of DEGs involved in the corresponding term or pathway (rich factor), while the color indicates the adjusted p-value (padj).
KEGG enrichment analysis DEGs revealed significant enrichment in several key pathways, including plant hormone signal transduction, biosynthesis of various plant secondary metabolites, glutathione metabolism, galactose metabolism, and ABC transporters (Fig. 4b). A total of 110 DEGs were associated with the top 30 enriched pathways (Table 2), among which 90 genes (81.90%) were upregulated, and 20 genes (18.10%) were downregulated. To gain deeper insights into peel-related biological processes, we focused on DEGs involved in plant hormone signaling and other pathways known to contribute to peel development, metabolism, and stress response for further analysis.
Table 2. KEGG pathways are related to hormone signaling and intracellular activity.
Pathway description KEGG ID Total DEGs Up Down Plant hormone signal transduction mdm04075 9 8 1 Glutathione metabolism mdm00480 7 5 2 Plant-pathogen interaction mdm04626 7 6 1 Biosynthesis of various plant secondary metabolites mdm00999 6 5 1 Galactose metabolism mdm00052 5 5 0 ABC transporters mdm02010 5 4 1 Peroxisome mdm04146 5 1 4 Biosynthesis of cofactors mdm01240 5 4 1 MAPK signaling pathway−plant mdm04016 4 4 0 Carbon metabolism mdm01200 4 3 1 Pentose and glucuronate interconversions mdm00040 3 2 1 Tryptophan metabolism mdm00380 3 3 0 Terpenoid backbone biosynthesis mdm00900 3 3 0 Efferocytosis mdm04148 3 2 1 Fructose and mannose metabolism mdm00051 3 3 0 Phagosome mdm04145 3 2 1 Biosynthesis of nucleotide sugars mdm01250 3 3 0 Phenylpropanoid biosynthesis mdm00940 3 2 1 Pyruvate metabolism mdm00620 3 2 1 Amino sugar and nucleotide sugar metabolism mdm00520 3 3 0 Starch and sucrose metabolism mdm00500 3 3 0 Glycolysis/gluconeogenesis mdm00010 3 2 1 Endocytosis mdm04144 3 2 1 Biosynthesis of amino acids mdm01230 3 3 0 Diterpenoid biosynthesis mdm00904 2 2 0 Nitrogen metabolism mdm00910 2 2 0 Arginine biosynthesis mdm00220 2 2 0 Ubiquinone and other terpenoid-quinone biosynthesis mdm00130 2 1 1 Steroid biosynthesis mdm00100 2 2 0 Glycosphingolipid biosynthesis−globo and isoglobo series mdm00603 1 1 0 Total number of DEGs 110 90 20 Percentages of up and down genes 100% 81.90% 18.10% DEGs related to hormone signal transduction
-
To elucidate hormonal regulation in apple peel development, DEGs involved in key plant hormone signaling pathways were analyzed, including those encoding hormone receptors and downstream response factors. This analysis focused on DEGs associated with auxin, GA, ABA, JA, and SA signaling pathways (Fig. 5). In the auxin signaling pathway, the auxin influx carrier gene (MdAUX1) was downregulated, while the GH3 family gene MdGH3.1, involved in auxin homeostasis, was upregulated. For GA signaling, only one DEG was identified: MdGID1C-like, encoding a gibberellin receptor, which was upregulated. In the ABA signaling pathway, both detected genes—MdPYR1, an ABA receptor, and MdPP2C56-like, a protein phosphatase 2C—were upregulated, suggesting enhanced ABA signaling activity (Fig. 5).
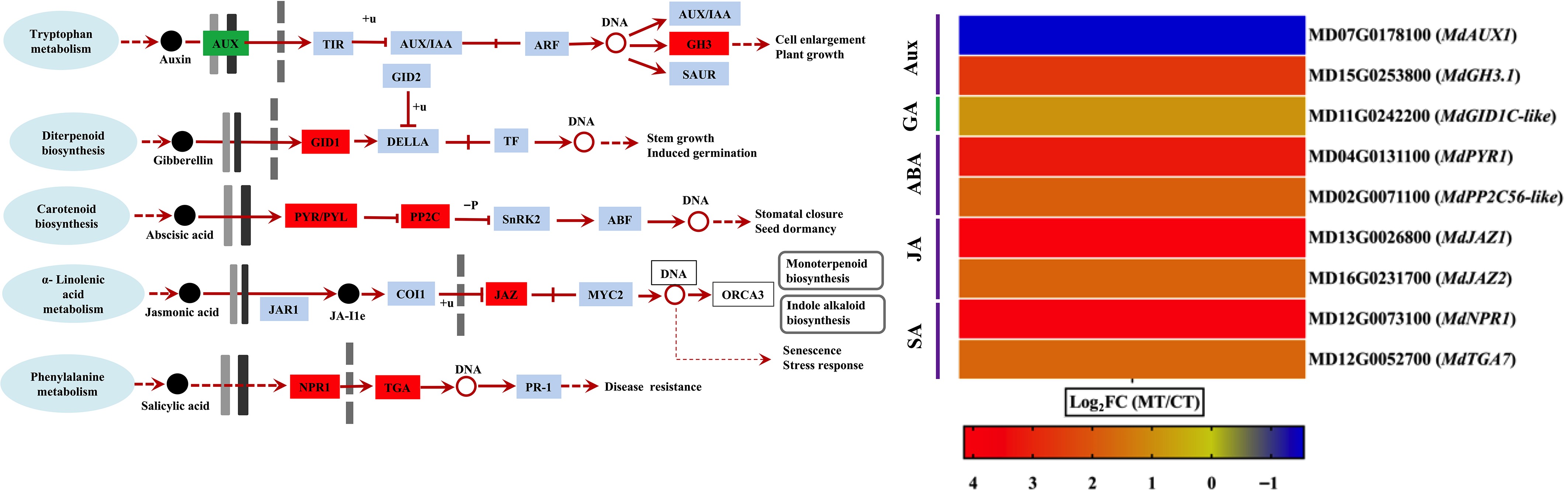
Figure 5.
Differential expression of hormone signaling-related genes in mutant-type (MT), and cultivated-type (CT) apple peels. The heatmap shows selected differentially expressed genes (DEGs) involved in major plant hormone signaling pathways, including auxin (AUX), gibberellin (GA), abscisic acid (ABA), jasmonic acid (JA), and salicylic acid (SA). Genes related to hormone receptors and downstream response regulators are included. Expression values are represented as log2 fold change (log2FC) between MT and CT. The color gradient indicates the relative expression level of each gene.
Within the JA signaling pathway, two DEGs were identified—MdJAZ1 and MdJAZ2—both encoding jasmonate ZIM-domain proteins, and both were upregulated, indicating activation of JA-mediated responses. Similarly, in the SA signaling pathway, the expression levels of MdNPR1 (Nonexpressor of Pathogenesis-Related Genes 1) and the MdTGA transcription factor were also upregulated, pointing to a potential role of SA in peel-related stress or defense responses (Fig. 5). These results collectively suggest that multiple hormone signaling pathways are transcriptionally modulated during apple peel development and may contribute to the phenotypic differences observed between mutant and cultivated apple types.
DEGs associated with specialized metabolism and transport pathways
-
Based on KEGG pathway enrichment, six pathways associated with peel physiology and stress response were selected for further analysis: biosynthesis of various plant secondary metabolites, glutathione metabolism, galactose metabolism, ABC transporters, glycosphingolipid biosynthesis, and terpenoid backbone biosynthesis. A number of key DEGs were identified within these pathways, reflecting transcriptional shifts between the mutant- and cultivated-type apple peels. In the biosynthesis of various plant secondary metabolites pathways, six DEGs were identified: MdCYP716 (β-amyrin 28-monooxygenase-like), MdNAS-like 1 and MdNAS-like 2 (nicotianamine synthase-like), MdCOR2-like (codeinone reductase-like), and MdSGT-like 1 and MdSGT-like 2 (scopoletin glucosyltransferase-like). All genes in this group were upregulated, except MdCOR2-like, which was downregulated (Fig. 6).
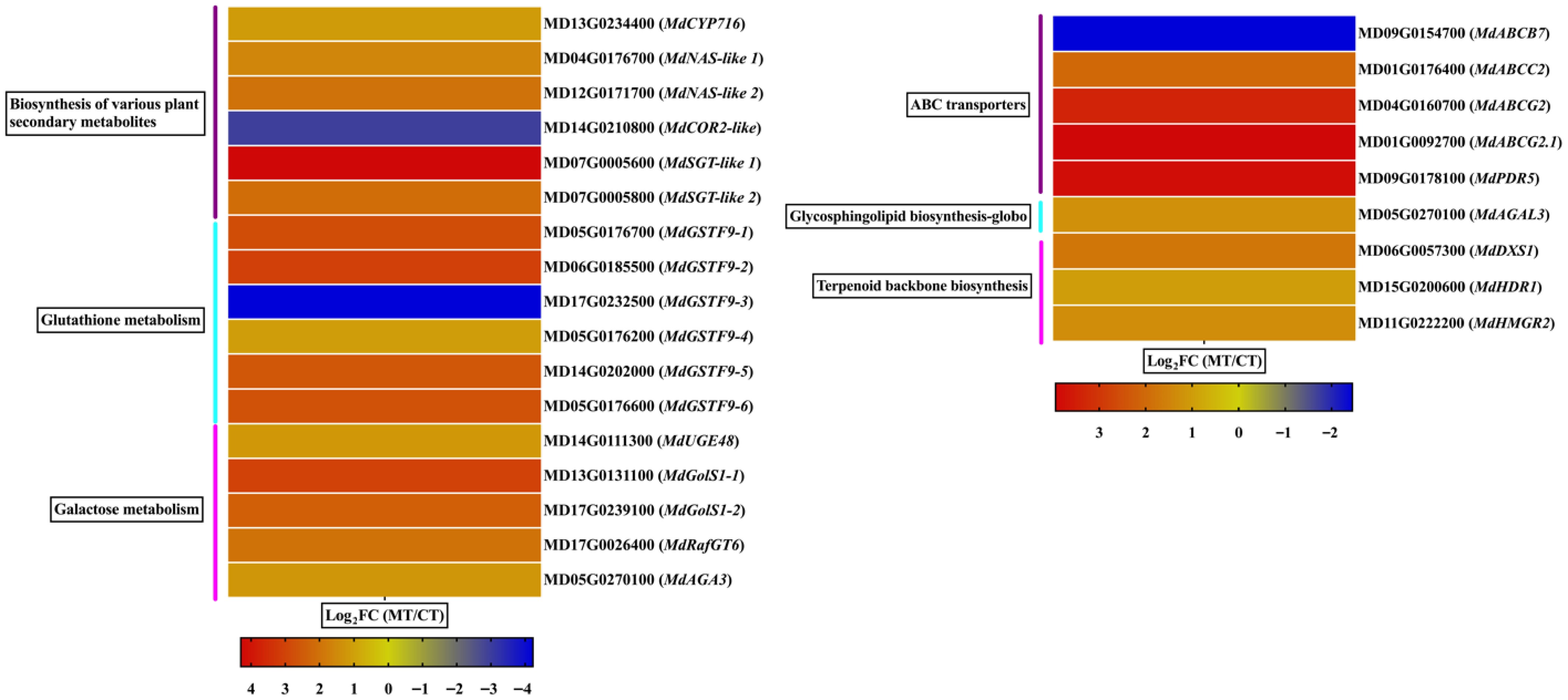
Figure 6.
Differential expression of genes involved in specialized metabolism and transport pathways in mutant-type (MT), and cultivated-type (CT) apple peels. The heatmap illustrates selected differentially expressed genes (DEGs) associated with key KEGG pathways, including the biosynthesis of various plant secondary metabolites, glutathione metabolism, galactose metabolism, ABC transporters, glycosphingolipid biosynthesis, and terpenoid backbone biosynthesis. Expression values are represented as log2 fold change (log2FC) between MT and CT. The color gradient indicates relative gene expression levels.
The glutathione metabolism pathway included six DEGs: MdGSTF9-1 through MdGSTF9-6 (glutathione S-transferase F9-like). All genes were upregulated except MdGSTF9-3, which showed downregulation. In the galactose metabolism pathway, all five identified DEGs were upregulated. These included MdUGE48 (UDP-glucose 4-epimerase), MdGolS1-1 and MdGolS1-2 (galactinol synthase 1), MdRafGT6 (galactinol–sucrose galactosyltransferase), and MdAGA3 (alpha-galactosidase 3) (Fig. 6). In ABC transporters, five DEGs were detected. One gene, MdABCB7, was downregulated, whereas MdABCC2, MdABCG2, MdABCG2.1, and MdPDR5 were upregulated, indicating a shift in transporter activity potentially related to hormone and metabolite movement across membranes (Fig. 6).
Regarding glycosphingolipid biosynthesis, only one DEG was identified: MdAGAL3 (alpha-galactosidase 3), which was upregulated. Similarly, all DEGs identified in terpenoid backbone biosynthesis were upregulated, including MdDXS1 (1-deoxy-D-xylulose-5-phosphate synthase 1), MdHDR1 (4-hydroxy-3-methylbut-2-enyl diphosphate reductase 1), and MdHMGR2 (3-hydroxy-3-methylglutaryl-CoA reductase 2) (Fig. 6), suggesting increased terpenoid precursor production. These findings collectively indicate that both metabolic and transport-related pathways are transcriptionally reprogrammed in mutant apple peels, likely contributing to phenotypic differences in peel thickness, composition, and stress adaptability.
-
Apple fruit is rich in vitamins and polyphenols, which contribute to its well-known nutritional value and health benefits[29,30]. This nutritional richness underpins the popular saying, 'An apple a day keeps the doctor away'. The apple peel consists of multiple outer layers—namely, the cuticle, epidermis, and hypodermis—that together protect the inner fleshy tissues rich in water, sugars, organic acids, and micronutrients[2,3]. These outer layers serve as a crucial barrier against environmental stressors during both growth and storage[31−33]. As such, peel thickness strongly influences fruit quality and shelf life, affecting not only storability but also consumer-relevant attributes such as appearance, taste, and texture.
In this study, a multidisciplinary approach was employed—combining morphological, physiological, and transcriptomic analyses—to compare a thin-peel apple mutant with a thick-peel cultivar. The integrated results reveal significant structural, biochemical, and molecular differences, offering novel insights into the biological mechanisms governing peel development and their implications for fruit quality.
Structural basis of peel thickness variation
-
Morphological observations and SEM confirmed that the mutant peel was significantly thinner (18.0–20.2 µm) than that of the thick-peel cultivar (31.3–33.7 µm). The mutant peel also exhibited fewer epidermal and hypodermal layers, a smoother surface, and reduced microcracking. These structural differences align with previous reports that associate higher epidermal layering and cuticular complexity with improved water retention and mechanical resistance[4,6,8], though the specific mechanisms in thin-peel mutants remain underexplored. Environmental factors and orchard management practices, such as irrigation regimes, can also influence cuticle structure. For instance, excessive irrigation can cause turgor-driven stretching of epidermal cells, leading to thinner cuticles and more surface microcracking, whereas deficit irrigation promotes thicker, more continuous cuticular layers[34−36]. Similarly, high relative humidity during storage can induce epidermal expansion and microcracking[37].
Differences in epidermal structure among apple cultivars have been documented[1,3,32,38]. Some studies suggest that surface roughness in apple and other fruits results from irregular cuticle deposition or variation in cuticle thickness among epidermal cells[39,40]. In apples, excessive microcracking and limited epicuticular wax production contribute to surface roughness[10,32,33]. Although thinner peels with fewer microcracks may enhance visual appeal and reduce transpiration to some extent, they also confer less resistance to mechanical damage during handling[9,16]. The smoother, less striated surface observed in the mutant peel likely reflects changes in epidermal cell architecture and cuticle deposition. These traits are often governed by lipid transport, cutin biosynthesis, and epidermal cell patterning, as previously reported in tomato and apple[20,21]. Our findings suggest that such structural changes contribute to the mutant's softer mouthfeel and altered surface texture.
Physiological evaluation of peel differences
-
Physiological evaluations revealed that the thin-peel mutant had significantly lower firmness than the thick-peel cultivar, a textural trait often associated with consumer preference for eating quality[7]. This reduced firmness can be directly explained by the comprehensively modified cell wall architecture of the mutant. It was found a significant reduction in all major structural components analyzed—cellulose, acid-insoluble lignin, and hemicellulose—compared to the cultivar. This coordinated downregulation of key structural polymers points to a systemic alteration in the cell wall biosynthetic pathway, rather than a targeted adjustment of a single component. Such a global reduction is consistent with patterns of large-scale cell wall remodeling during fruit softening but suggests a distinct genetic or developmental origin in this mutant[14]. The significantly lower deposition of these core components provides a clear mechanistic basis for the mutant's softer texture and reduced structural integrity. These results refine the current understanding of how cell wall composition dictates fruit quality and postharvest performance[5], They demonstrate that a pleiotropic effect on multiple cell wall polymers can underlie a pronounced phenotype, highlighting a potential regulatory node that could be targeted for breeding apples with tailored texture and durability.
Transcriptomic changes underlying peel development
-
RNA-seq analysis revealed clear transcriptional reprogramming in the mutant peel, identifying 467 DEGs, most of which were upregulated. Many of these DEGs were linked to plant hormone signaling pathways, suggesting that peel development is tightly controlled by hormonal crosstalk. Auxin signaling was altered in the mutant, with downregulation of MdAUX1 (auxin influx carrier) and upregulation of MdGH3.1, involved in auxin homeostasis. This attenuation of auxin signaling in our apple mutant provides a plausible molecular explanation for the observed reduction in epidermal cell layers and overall peel thinning. While similar auxin-mediated epidermal patterning has been reported in tomato[17], and auxin homeostasis is known to coordinate fruit development[18], the present data specifically implicates the downregulation of MdAUX1, and upregulation of MdGH3.1 as key regulatory events in apple peel development.
GA signaling was enhanced via upregulation of MdGID1C-like, a GA receptor gene. It is hypothesized that in apple, enhanced GA perception may promote epidermal cell expansion without proliferation, leading to the characteristic thinner, softer peel—a phenotype distinct from the well-characterized role of GA in promoting internode elongation. Concurrently, the upregulation of ABA signaling components (MdPYR1 and MdPP2C56-like) suggests a compensatory mechanism in the apple mutant where a thinner peel might be counterbalanced by a potentially stronger cuticle, a concept supported by studies in Arabidopsis[19,41]. Thus, increased ABA activity may help compensate for reduced physical thickness by enhancing cuticle functionality. JA and SA pathways (upregulation of MdJAZ1, MdJAZ2, MdNPR1, and MdTGA) likely enhance peel resilience through defense-linked cuticle remodeling[19], and MYB93-mediated suberin deposition in apple[22]. Future research using hormone application assays in apple fruit could directly test these hypotheses.
Metabolic and transport pathways supporting peel remodeling
-
DEGs enriched in metabolic pathways provided further insights into peel remodeling. Upregulation of genes involved in glutathione metabolism and galactose metabolism suggests enhanced ROS detoxification and sugar interconversion, crucial for maintaining cell homeostasis in thinner peels[4,23,42,43]. Terpenoid backbone biosynthesis is a well-known pathway for producing secondary metabolites like terpenes, which contribute to aroma, flavor, and structural properties (e.g., cuticular waxes) in fruit peels[4,44]. Meanwhile, increased expression of ABC transporter and glycosphingolipid biosynthesis genes, including MdABCG2 and MdPDR5, highlights the importance of these transport systems for lipid trafficking to the cuticle in apple, a process critical for maintaining membrane integrity and cuticle formation[21]. Interestingly, several secondary metabolite biosynthetic genes were also upregulated in the mutant. While specific flavonoid content was not quantified in this study, such specialized metabolites may play roles in peel pigmentation, antioxidant defense, and pathogen resistance[22,23]. Future work should focus on validating these genes and quantifying their associated metabolite accumulation.
-
This study provides a comprehensive investigation into the morphological, physiological, and transcriptomic differences between a thin-peel apple mutant and a thick-peel cultivated apple. Morphological and microstructural analyses confirmed that the mutant peel is significantly thinner, with reduced cell layering and fewer surface microcracks. Physiological measurements confirmed that the mutant fruit has a significantly softer texture, a key consumer trait, which is mechanistically explained by its significantly reduced deposition of structural cell wall components, including cellulose, lignin, and hemicellulose. At the molecular level, transcriptome profiling revealed substantial reprogramming of gene expression in the mutant peel, including the upregulation of genes involved in hormone signaling, secondary metabolism, and membrane transport. The enrichment of pathways such as auxin, GA, ABA, and JA signaling, along with the biosynthesis of specialized metabolites and ABC transporters, suggests a coordinated regulatory network underlying peel development, composition, and stress adaptability. Collectively, these findings elucidate the molecular basis of peel thickness variation in apples and provide valuable insights for breeding programs aimed at improving fruit quality traits such as texture, flavor, and postharvest performance. Future studies focusing on functional validation of candidate genes and regulatory elements will further enhance our understanding of peel development mechanisms and their agronomic relevance.
This study was supported by the Experimental Demonstration of Apple Green and Simplified Fertilization Technology 2023 Provincial Agricultural Special Project (Billion-Yuan Fruit Industry Program) Department of Agriculture of Shaanxi Province, Young Breeding Experts Cultivation Support Plan of Yangling Demonstration Zone, and Construction Project of 'PhD + Vocational Students' Workshop at Yangling Vocational & Technical College (Project No. BG202004).
-
The authors confirm their contributions to the paper as follows: the main author: Zhang W; participated in the experimental design: Zhang W, Tahir MM; conducted experiments and performed data analysis: Zhang W, Han Y, Ma Y, Ma W, Yuan J; participated in paper writing: Zhang W, Tahir MM. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Summary of the sequencing statistics obtained from each sample.
- Supplementary Table S2 Summary of the total, multiple, and uniquely mapped reads obtained from each sample.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang W, Han Y, Ma Y, Ma W, Yuan J, et al. 2026. Physiological and molecular insights into peel thickness variation between mutant and cultivated type apples. Fruit Research 6: e001 doi: 10.48130/frures-0025-0037 

Physiological and molecular insights into peel thickness variation between mutant and cultivated type apples
- Received: 31 July 2025
- Revised: 27 September 2025
- Accepted: 30 October 2025
- Published online: 20 January 2026
Abstract: The apple peel plays a vital role in determining fruit quality and postharvest performance. In this study, the morphological, physiological, and transcriptomic differences between a thin-peel mutant and a thick-peel cultivated apple were comprehensively investigated. Morphological analysis, including scanning electron microscopy (SEM), revealed that mutant-type apple peels were significantly thinner (18.0–20.2 µm) and more porous than those of the cultivated type (31.3–33.7 µm), with reduced cell layering and fewer cuticular microcracks. Physiologically, mutant-type apples exhibited significantly lower firmness and reduced deposition of key structural components (cellulose, acid-insoluble lignin, and hemicellulose), features that are directly associated with a softer texture and altered postharvest potential. Transcriptomic profiling identified 467 differentially expressed genes (DEGs), comprising 368 upregulated, and 99 downregulated. Functional enrichment analyses highlighted the activation of multiple hormone signaling pathways, including auxin, gibberellin, abscisic acid, jasmonic acid, and salicylic acid, suggesting hormonal modulation of peel development. Additionally, key metabolic and transport pathways were enriched with upregulated DEGs in mutant-type apple peels, including the biosynthesis of secondary metabolites, glutathione and galactose metabolism, ABC transporters, glycosphingolipid biosynthesis, and terpenoid backbone biosynthesis. These transcriptional changes point to enhanced metabolic activity and membrane transport functions in the mutant-type apple peels. Together, these findings offer novel insights into the structural and molecular basis of peel thickness variation in apples and provide a foundation for breeding programs aimed at improving peel-related fruit quality traits.
-
Key words:
- Apple /
- Peels /
- Mutant-type /
- Cultivar-type /
- SEM /
- RNA-seq