









-
Apple (Malus domestica) is a globally significant commercial fruit, with China leading the world in cultivation area, total output, per capita share, and export volume. Despite this dominance, China's fruiting rate lags behind that of developed countries, primarily due to the limited adoption of high-density cultivation systems. Modern apple industries in China are increasingly transitioning to high-density cultivation, which relies heavily on dwarf rootstocks. Asexual propagation through stem cuttings is a preferred method for apple cultivation due to its efficiency and ability to produce genetically uniform plants. However, the formation of adventitious roots (ARs) from stem cuttings—a critical step for reestablishing water and nutrient uptake—remains a major bottleneck for mass propagation[1]. Grafting apple scions onto desirable rootstocks can enhance adaptability to diverse soil conditions and resistance to soilborne pests and diseases. Unfortunately, many apple rootstocks with favorable traits exhibit poor rooting ability from stem cuttings, underscoring the need to improve rooting efficiency to advance China's apple industry.
AR formation is a complex process involving three distinct stages: (1) induction (0–3 d), characterized by molecular reprogramming; (2) initiation (3–8 d), marked by AR primordia formation; and (3) emergence (after 8 d), when ARs break through the tissue[2]. In Arabidopsis, AR primordia originate from pericycle cells, while in apple, they arise from interfascicular cambium cells adjacent to phloem tissue[3−5]. This process is tightly regulated by a multitude of factors, including endogenous hormones, nutritional status, wounding, and environmental conditions. Previous studies have demonstrated that macronutrient availability—particularly nitrate and potassium—positively influences AR formation in apple rootstocks. Nitrate promotes AR initiation by enhancing cell division and differentiation, increasing indole-3-acetic acid (IAA) accumulation, reducing abscisic acid (ABA) and zeatin riboside (ZR) levels, and upregulating auxin transport and signaling genes (MdAUX1 and MdIAA23) as well as root development regulators (MdWOX11, MdLBD16, and MdLBD29)[2]. Potassium accelerates AR formation by stimulating auxin transport (MdPIN1, MdPIN2, and MdAUX1) and activating cell cycle and root regulatory genes (MdWOX11, MdLBD16, and MdLBD29), thereby promoting both initiation and emergence phases[6]. However, the role of phosphorus (P)—another essential macronutrient—remains largely unexplored.
P is vital for numerous physiological processes in plants, including energy transfer, root development, and hormone regulation[7,8]. As a key component of ATP (adenosine triphosphate), P ensures the energy required for cell division and elongation, which are critical for root initiation and growth. During AR formation, P supports the metabolic processes necessary for the development of root primordia and their subsequent[9,10]. Although direct reports on the effects of P on AR formation are limited, studies in other plant systems demonstrate that P availability modulates root initiation capacity, root elongation, and overall root system architecture through both hormonal and metabolic regulation. For example, P deficiency alters carbohydrate allocation and disrupts auxin-related transport or sensitivity, which can lead to impaired AR development[11−13]. Phosphate stress also induces root architectural changes via auxin-dependent and auxin-independent signaling pathways, with mediators like BIG activating pericycle cell division[14,15]. While auxin promotes early stages of AR formation, it can inhibit later stages[16]. This biphasic role of auxin is critical for regulating the timing and progression of AR development[17]. Other hormones, such as cytokinin (CK)[18,19] and ABA[20], generally inhibit AR formation, whereas brassinosteroids (BRs) have been shown to negatively affect rooting[21]. Thus, integrating current knowledge on P–hormone interactions and root development helps establish the physiological framework for understanding how P availability shapes AR formation in apple rootstocks.
This study aims to investigate the influence of P availability on AR formation in K2 apple rootstocks. In a preliminary experiment, the effects of low P (LP), medium P (MP), and high P (HP) levels were assessed, compared with a control (CK; containing the basal P level of standard MS medium), by measuring rooting percentage and number of ARs. Based on these results, LP was selected for detailed physiological and transcriptomic analyses due to its pronounced promotive effect on AR formation. It is hypothesized that LP conditions enhance AR formation by modulating hormonal pathways and regulating genes associated with hormone signaling. By elucidating the molecular and physiological mechanisms underlying LP's role in AR formation, this study provides valuable insights for improving the efficiency of apple asexual reproduction and supporting the sustainable development of China's apple industry.
-
In this study, stem cuttings of the K2 apple rootstock were used. The 'K2 apple' is a novel hybrid cultivar derived from a controlled cross between G.41 (a dwarfing Malus domestica rootstock from the Geneva® series) and Qiu Zi (Malus prunifolia [Willd.] Borkh.). Plant materials were obtained from Northwest A&F University, China. The preliminary experiment involved 144 four-week-old, morphologically homogeneous K2 apple rootstock stem cuttings grown on half-strength Murashige and Skoog (MS) medium containing sucrose (30 g/L), agar (7 g/L), IBA (1.2 mg/L), and pH adjusted to 5.8. The basal MS medium, containing approximately 0.04 g/L P derived from KH2PO4, was used as the CK and served as the true baseline.
To assess the effect of P availability on AR formation, three additional P treatments were prepared by supplementing the basal MS medium with 0.08 g/L (Low P, LP), 0.16 g/L (Medium P, MP), and 0.32 g/L (High P, HP) KH2PO4. These additions resulted in total P concentrations of 0.12 g/L (LP), 0.20 g/L (MP), and 0.36 g/L (HP), respectively. Although the LP treatment contains more P than the basal CK, it is referred to as 'Low P' relative to MP and HP to indicate the lowest supplemented level in the experimental gradient. This dose–response design allowed us to identify the optimal P concentration for promoting AR induction. A total of 36 stem cuttings were used per treatment. After 30 d of treatment, AR parameters—including rooting percentage and the number of ARs per cutting—were measured manually.
For the subsequent experiment, 420 cuttings were used, with 60 cuttings assigned to each LP and CK treatment at each time point. The study aimed to assess the effect of LP during AR formation, which is divided into three distinct stages in apple: 0–3 d (induction), 3–8 d (initiation), and post-8 d (emergence). The basal 0.5 cm segment of each stem cutting was designated as the rooting zone. Samples were collected at 0, 3, 8, and 16 d, rapidly frozen in liquid nitrogen, and preserved at −80 °C for subsequent analysis.
Anatomical and hormonal analyses
-
The harvested samples (0, 3, 8, and 16 d) were processed for fixation, paraffin embedding, and sectioning, following the previous method[4]. Additionally, endogenous hormones such as IAA, ABA, BR, ZR, gibberellic acid 3 (GA3), and methyl jasmonate (MeJA) were extracted and purified using an enzyme-linked immunosorbent assay (ELISA) at the Center for Plant Growth Regulators, China Agricultural University. Detailed methodology can be found in Dobrev & Kamınek[22].
RNA extraction and quality examination
-
Total RNA was isolated from the basal stem sections of LP- and CK-treated cuttings at 3 d using the RNAprep Pure Plant Kit (Tiangen, Beijing, China) in accordance with the manufacturer's instructions. RNA purity was evaluated using the Kaiao K5500® Spectrophotometer (Kaiao, Beijing, China). The RNA's integrity and concentration were determined using the RNA Nano 6000 Assay Kit on the Bioanalyzer 2100 system (Agilent Technologies, CA, USA).
Construction of cDNA and RNA-seq libraries and sequencing
-
mRNA was purified using oligo(dT)-attached magnetic beads, eliminating rRNA and tRNA, with 2 μg of RNA per sample. The mRNA was fragmented into small pieces using a fragmentation buffer. First-strand cDNA was synthesized by random hexamer priming, followed by second-strand cDNA synthesis. RNA Index Adapters and A-Tailing Mix were added for end repair. The cDNA fragments were amplified by PCR, purified using Ampure XP beads, and resolved in an EB solution. The integrity of the PCR products was validated using an Agilent Technologies 2100 Bioanalyzer. These double-stranded PCR products were denatured and circularized using a splint oligo sequence, generating single-stranded circular DNA (ssCir DNA) as the final library. The libraries were amplified with phi29 to produce DNA nanoballs (DNBs), which were loaded onto a patterned nanoarray. Paired-end 150 bp base reads were generated on the T7 platform by Wuhan Benagen Technology Co., Ltd. (Wuhan, China).
RNA sequencing
-
The raw sequencing data were processed using FastQC (
www.bioinformatics.babraham.ac.uk/projects/fastqc ) to remove adapters and low-quality sequences. Clean reads were aligned to the Malus domestica reference genome (www.rosaceae.org/species/malus/malus_x_domestica ) using HISAT. Gene expression levels were quantified using the RSEM package and expressed as fragments per kilobase of transcript per million fragments mapped (FPKM). Differentially expressed genes (DEGs) were identified by comparing treatment samples with control samples using fold-change > 2.00 and probability ≥ 0.8, with a false discovery rate (FDR) adjusted p-value < 0.05 based on three biological replicates. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed using the clusterProfiler (version 3.8).RT-qPCR expression analysis
-
RT-qPCR validation of the selected DEGs involved in hormone signaling pathways was performed as described previously[23], using the apple actin gene as an internal control. Expression levels were calculated using the 2ΔΔCᴛ method[24]. Primer sequences are listed in Supplementary Table S1.
Statistical analysis
-
Graphical representations, heatmaps, and statistical analyses were performed using GraphPad Prism version 10.00. Data were analyzed by two-way ANOVA followed by Sidak's multiple comparisons test, with statistical significance set at p < 0.05. Results are presented as mean ± SD and denoted as follows: ns (p > 0.05), * p < 0.05, ** p < 0.01, and *** p < 0.001.
-
In the preliminary experiment, three P concentrations were tested against the CK to identify the most effective concentration for enhancing AR formation and development in K2 apple rootstock stem cuttings (Fig. 1a). The results showed that LP treatment significantly increased the rooting percentage compared to the CK, while HP treatment decreased the rooting percentage (Fig. 1). Furthermore, LP-treated cuttings also exhibited a 60% increase in AR numbers per cutting, while HP-treated cuttings showed a 70% reduction compared to the CK. These findings suggest that LP promotes AR formation, while HP inhibits it, likely due to P toxicity.
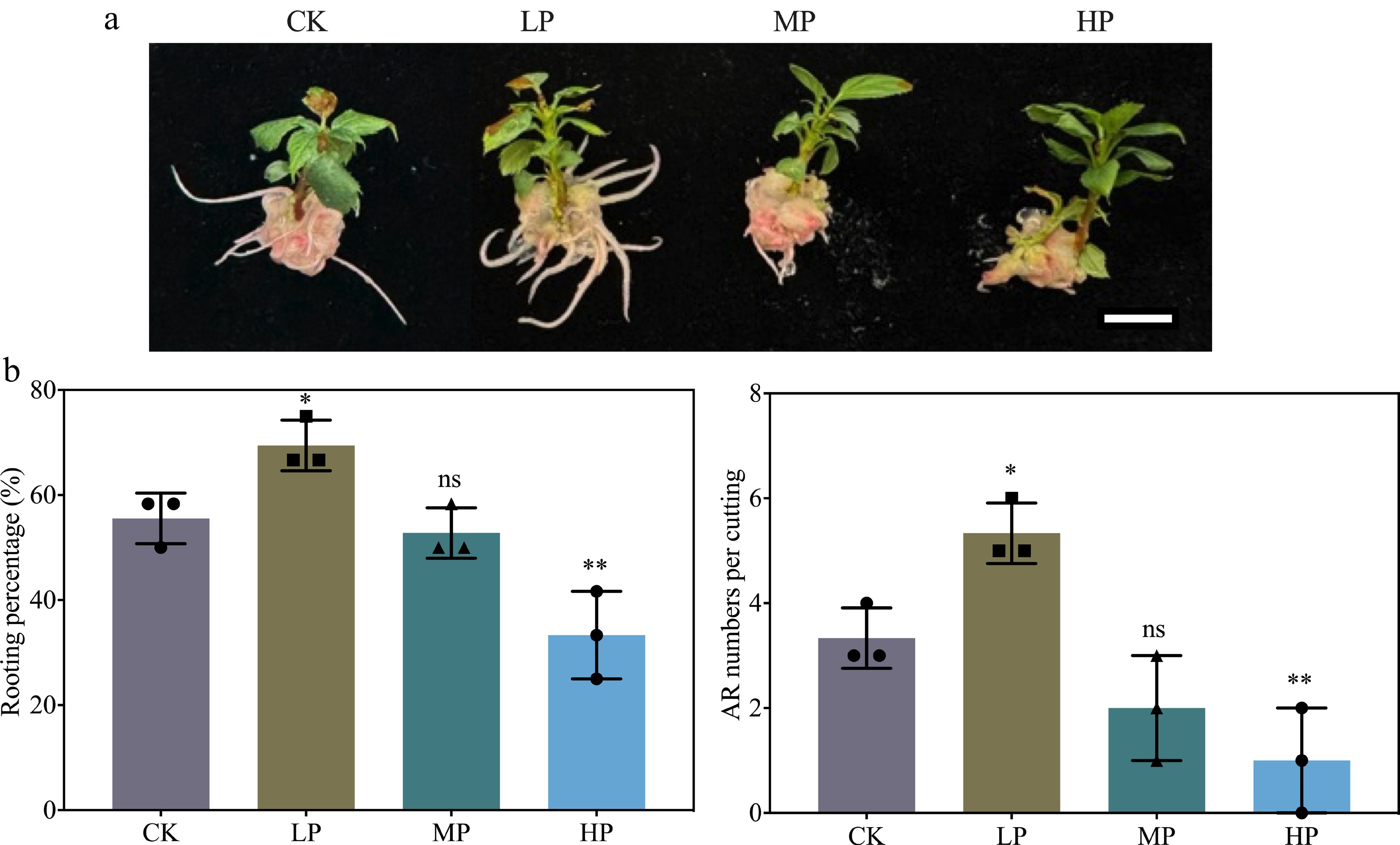
Figure 1.
Effect of graded phosphorus (P) concentrations on adventitious root (AR) formation in K2 apple rootstock stem cuttings. Stem cuttings were grown on half-strength MS medium with the basal P level (CK, 0.04 g/L) or supplemented with 0.08 g/L (Low P, LP; total P 0.12 g/L), 0.16 g/L (Medium P, MP; total P 0.20 g/L), or 0.32 g/L (High P, HP; total P 0.36 g/L) KH2PO4. Although the LP treatment contained more P than CK, it is termed 'Low P' relative to MP and HP in this dose–response design. Scale bar = 1 cm. AR morphological parameters, including rooting percentage and AR numbers per cutting, were measured after 30 d of treatment. Error bars indicate mean ± SD from three biological replicates. Different asterisks denote significance levels: ns, p > 0.05; * p < 0.05; ** p < 0.01.
Morphological and anatomical observation
-
Based on the preliminary results, LP was selected for further experiments to investigate its effect on AR formation at different stages, such as 0, 3, 8, and 16 d (Fig. 2). No significant phenotypic differences were observed between CK- and LP-treated cuttings at 0, 3, and 8 d. However, by 16 d, both groups exhibited AR outbreak, with LP-treated cuttings showing a more pronounced AR outbreak (Fig. 2). Anatomical analysis of the stem basal region at 0 d revealed the presence of component cells in both groups. By 3 d, mitotic activity was noticed in the cambial cells, and cell division started in the agminated differentiated cells. However, AR primordia appeared in both CK- and LP-treated cuttings, but LP-treated cuttings had a higher number of primordia, contributing to the higher AR numbers (Fig. 2). These findings suggest that LP enhances AR primordia formation, thereby promoting greater AR development, indicating a positive correlation between LP treatment and AR formation in K2 apple rootstock.
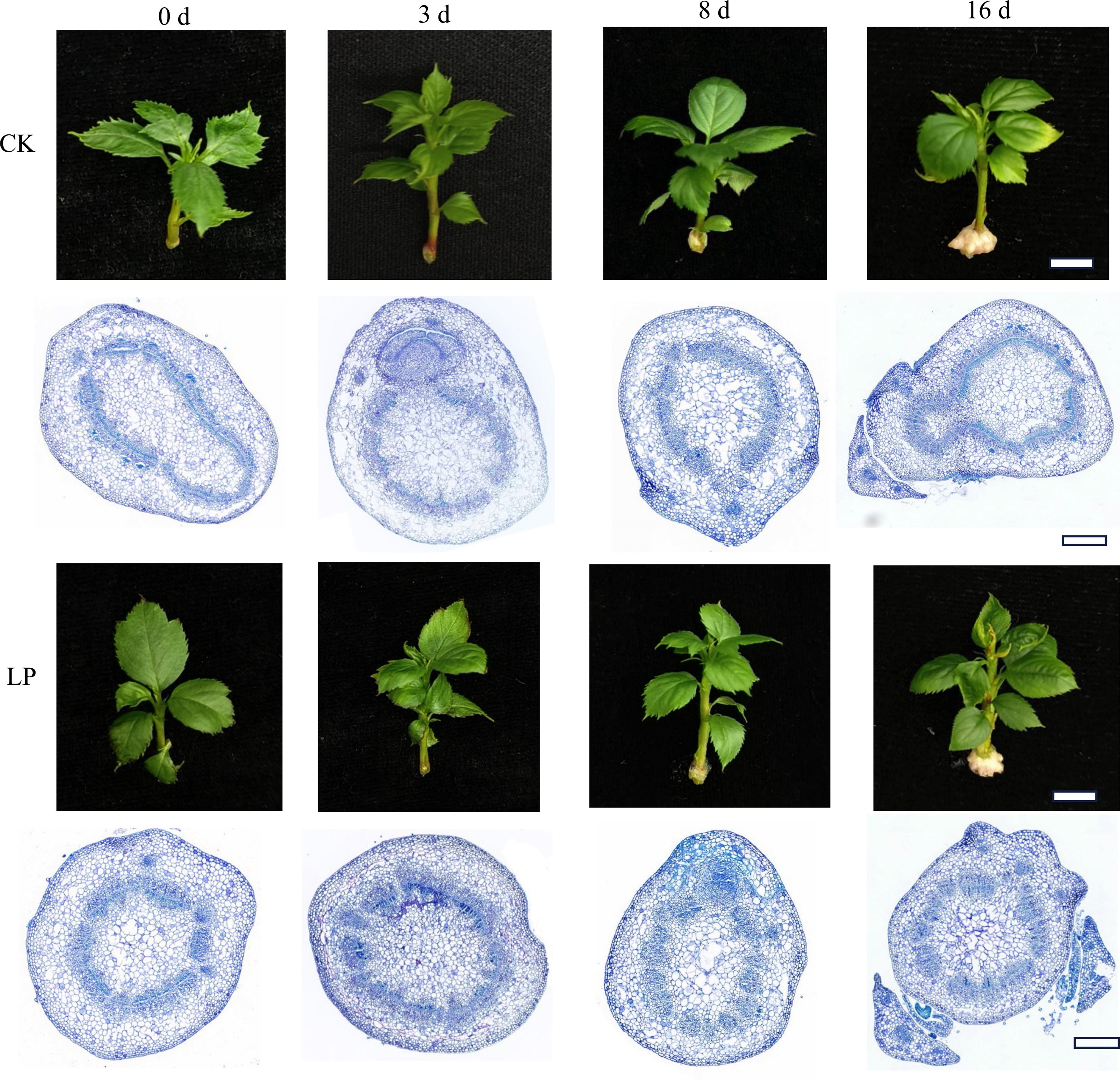
Figure 2.
Morphological and anatomical observations of adventitious root (AR) formation in K2 apple rootstock stem cuttings at different time points (0, 3, 8, and 16 d). Cuttings were grown on half-strength MS medium with the basal P level (CK, 0.04 g/L) or supplemented with 0.08 g/L KH2PO4 (Low P, LP; total P 0.12 g/L). Scale bars: 1 cm for morphological images and 500 μm for microscopic images.
Analysis of RNA-sequencing data
-
To gain deeper insights into the molecular mechanisms underlying the response to LP treatment during AR formation, messenger RNA (mRNA) analysis was performed at the crucial 3 d time point, which marks the initiation of molecular reprogramming required for adventitious rooting. Between 40,003,138 and 40,056,966 raw reads were generated across all libraries, with clean reads ranging from 40,003,064 to 40,056,926, indicating high-quality sequencing. Quality assessments showed that the Q20 and Q30 rates exceeded 98.247% and 95.173%, respectively, while the GC content consistently remained above 46.143% (Supplementary Table S2). Mapping analysis revealed that between 97.17% and 97.41% of total reads were successfully mapped, with 93.43% to 93.64% uniquely mapped and 3.68% to 3.77% showing multiple mappings. The percentage of unmapped reads ranged from 2.59% to 2.83% (Supplementary Table S3).
To further analyze gene expression, boxplots and density plots were generated to estimate the log10 FPKM values and assess the distribution of data for both sample groups, as shown in Supplementary Fig. S1a, S1b. Principal component analysis (PCA) revealed distinct separation between CK- and LP-treated K2 apple rootstock stem cuttings. The findings highlight notable variations in gene expression patterns, demonstrating substantial differences in the transcriptional profiles between the two treatment conditions (Supplementary Fig. S1c). In total, 39,249 genes were identified, including 1,184 genes uniquely found in CK-treated cuttings, 1,125 genes uniquely found in LP-treated cuttings, and 36,940 shared genes (Fig. 3a). To assess differential gene expression, a volcano plot, which revealed expression changes, identifying 1,009 DEGs, was generated. Among these, 383 genes were upregulated, and 626 genes were downregulated (Fig. 3b).
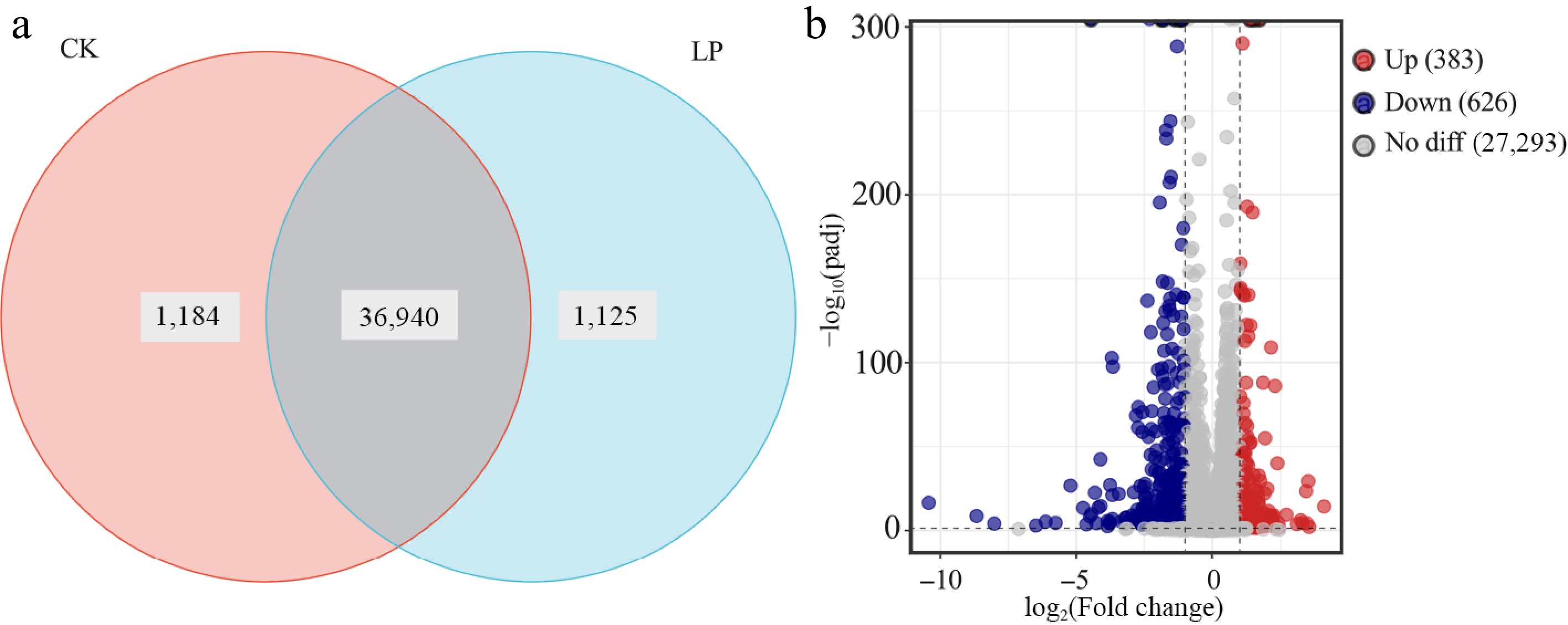
Figure 3.
Transcriptomic analysis of K2 apple rootstock stem cuttings on the 3rd day of adventitious root (AR) formation under low-phosphorus (LP, 0.08 g/L; total P 0.12 g/L) treatment compared with the control (CK, basal P level of standard MS medium, 0.04 g/L). (a) Venn diagram showing unique and shared genes between CK and LP-treated groups. (b) Volcano plot of differentially expressed genes (DEGs), with each dot representing a specific DEG: red dots indicate upregulated genes, blue dots indicate downregulated genes, and gray dots indicate non-significant genes.
Functional annotation of DEGs
-
GO enrichment analysis of DEGs revealed significant enrichment in various biological processes and functions. The highest number of DEGs were observed in categories such as carbohydrate metabolic processes, integral components of the membrane, cation binding, and oxidoreductase activity (Fig. 4a). Additionally, KEGG enrichment analysis highlighted several key pathways with a large proportion of enriched DEGs, including the biosynthesis of secondary metabolites, metabolic pathways, and plant hormone signal transduction (Fig. 4b). Furthermore, the KEGG pathway circle diagram identified five major categories, such as cellular processes, environmental information processing, genetic information processing, metabolism, and organismal systems. Notably, the metabolism and organismal systems categories contained the largest proportion of DEGs (Fig. 5).
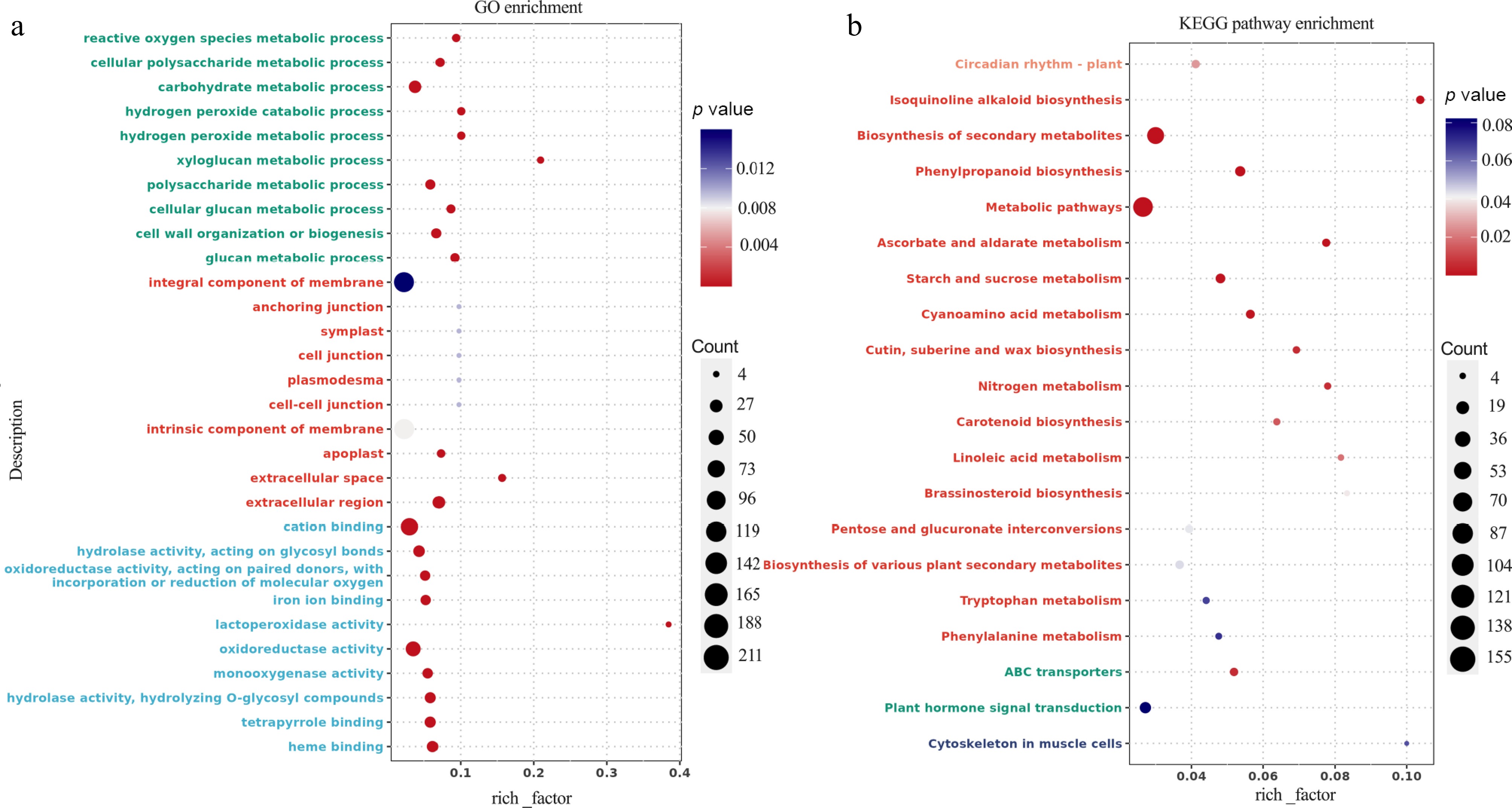
Figure 4.
Transcriptomic analysis of K2 apple rootstock stem cuttings on the 3rd day of adventitious root (AR) formation under low-phosphorus (LP, 0.08 g/L; total P 0.12 g/L) treatment compared with the control (CK, basal P level of standard MS medium, 0.04 g/L). (a) Gene Ontology (GO) enrichment analysis of differentially expressed genes (DEGs). (b) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of DEGs. The vertical axis shows the pathway names and GO terms, the horizontal axis represents the rich factor, the size of each point indicates the number of DEGs, and the color of the points corresponds to the p-value.
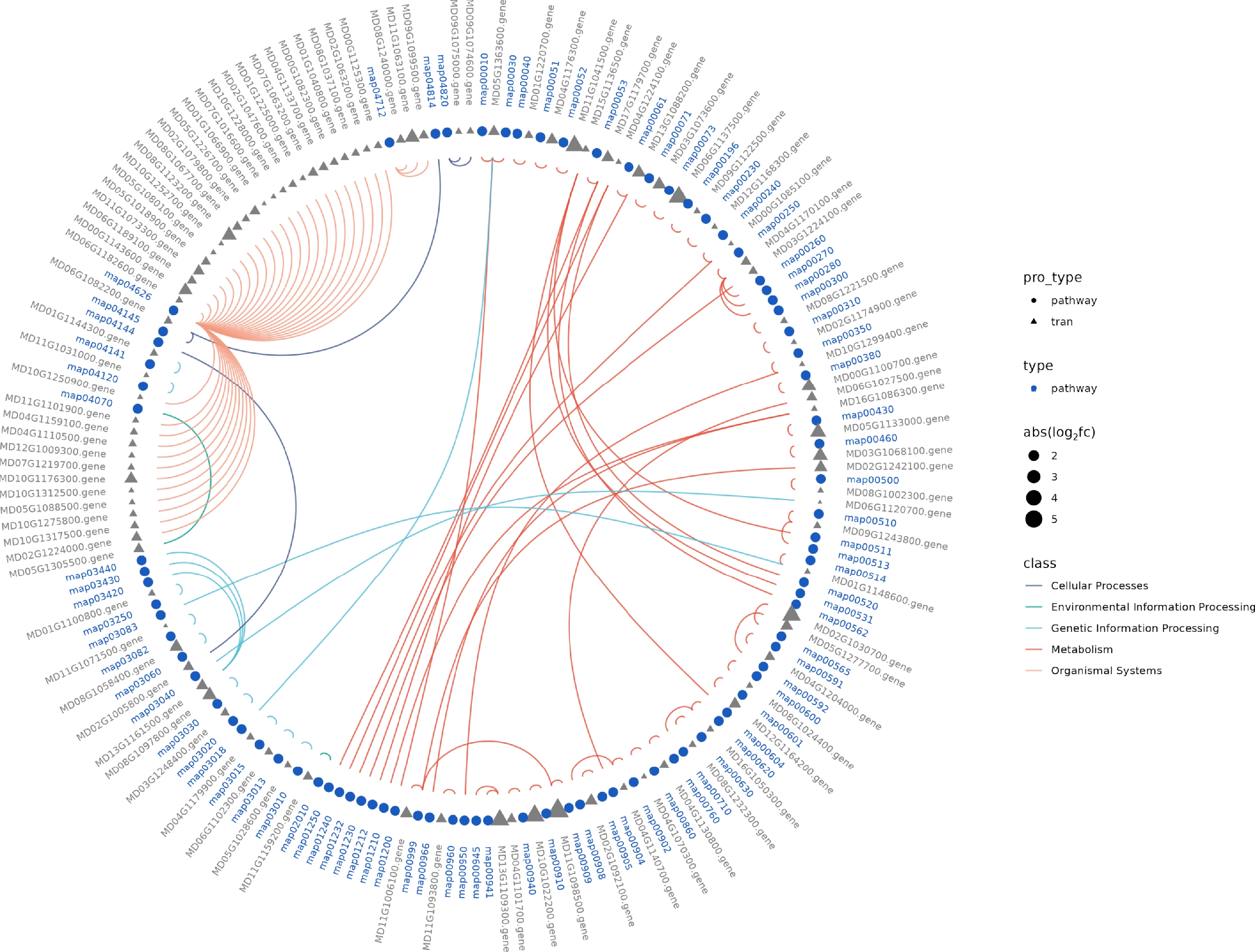
Figure 5.
Transcriptomic analysis of K2 apple rootstock stem cuttings on the 3rd day of adventitious root (AR) formation under low-phosphorus (LP, 0.08 g/L; total P 0.12 g/L) treatment compared with the control (CK, basal P level of standard MS medium, 0.04 g/L). The Kyoto Encyclopedia of Genes and Genomes (KEGG) circle illustrates different pathway types and their associated differentially expressed genes (DEGs) across the classifications.
Quantification of endogenous hormones and analysis of hormone signaling pathway-related DEGs
-
Previous studies have shown that endogenous hormones and DEGs associated with hormone signaling pathways play a significant role in AR formation in apples[23]. Therefore, this study analyzed the levels of endogenous hormones (at 0, 3, 8, and 16 d) and DEGs related to various hormone signaling pathways (at 3 d, a crucial time point for molecular reprogramming) to investigate the physiological and molecular responses induced by LP treatment.
The results showed that IAA levels were significantly improved at 3 d and decreased at 8 and 16 d in both CK- and LP-treated cuttings. At 3 and 8 d, LP-treated cuttings showed higher IAA levels than CK-treated cuttings. However, at 16 d, this trend reversed, with IAA levels being lower in LP-treated cuttings compared to CK-treated cuttings. Key DEGs involved in the auxin signaling pathway include auxin response factors (ARFs), auxin-responsive GH3 family proteins (GH3), small auxin upregulated RNAs (SAUR), and transmembrane kinase 1/4 (TMK 1/4). The expression of ARF1, GH3.1, SAUR1, and SAUR2 was downregulated, while the expression of SAUR3, TMK1/4a, and TMK1/4b was upregulated (Fig. 6a). For ZR, the content was consistently higher in CK-treated cuttings at all time points. DEGs related to CK signaling, including histidine-containing phosphotransmitter (APH) and type-B response regulators (B-ARR), were examined. The expression of APH1 was upregulated, while the expression of both B-ARR1 and B-ARR2 was downregulated (Fig. 6b). Similar to ZR, ABA levels were also higher in CK cuttings at all time points, although ABA content increased at 3 d but decreased at subsequent time points. In the ABA signaling pathway, key DEGs include abscisic acid receptors (PYR/PYL), where both PYR/PYL1 and PYR/PYL2 were downregulated (Fig. 6c).
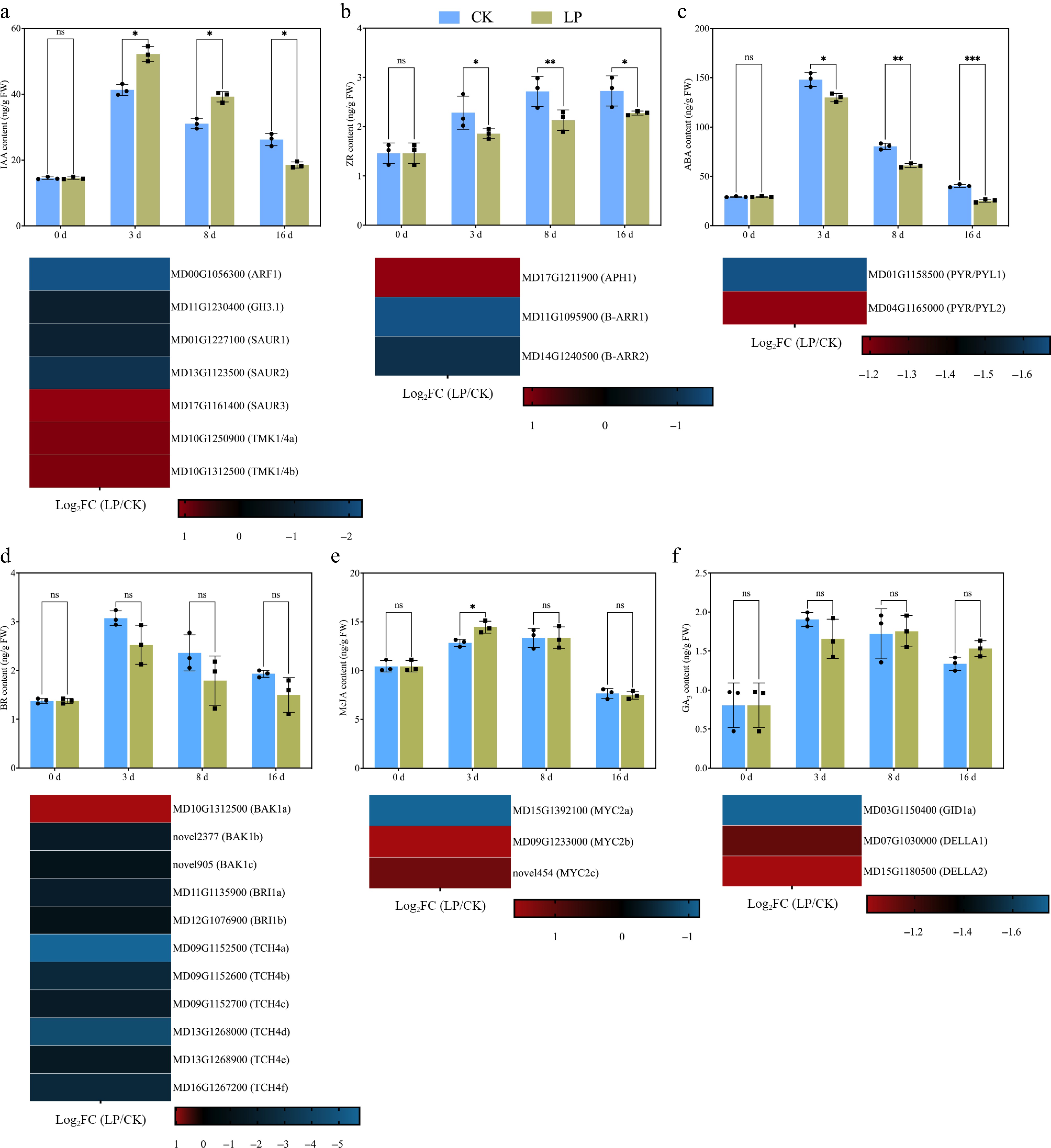
Figure 6.
Effect of low-phosphorus (LP, 0.08 g/L; total P 0.12 g/L) treatment compared with the control (CK, basal P level of standard MS medium, 0.04 g/L) on endogenous hormone levels in K2 apple rootstock stem cuttings during adventitious root (AR) formation at 0, 3, 8, and 16 d. Hormones measured include (a) indole-3-acetic acid (IAA), (b) zeatin riboside (ZR), (c) abscisic acid (ABA), (d) brassinosteroid (BR), (e) methyl jasmonate (MeJA), and (f) gibberellic acid 3 (GA3). Error bars refer to the average value ± SD from three biological replicates. Different asterisks indicate significant differences at ns p > 0.05, * p < 0.05, ** p < 0.01, and *** p < 0.001. Below each hormone level graph, the corresponding heatmap displays the expression of selected differentially expressed genes (DEGs) related to different hormone metabolisms and signaling pathways obtained from RNA-seq data (at 3rd d, which is crucial for the initiation of molecular reprogramming required for AR formation). The heatmaps show the log2FC values for genes across the pairwise comparison of CK_vs_LP, with the color gradient representing the gene expression levels.
BR exhibited a similar pattern to ABA, being higher in CK-treated cuttings at all time points. BR content improved at 3 d and decreased at 8 and 16 d. Key DEGs involved in the BR signaling pathway, such as brassinosteroid-associated kinase 1 (BAK1), brassinosteroid insensitive 1 (BRI1), and xyloglucan endotransglucosylase/ hydrolase family protein 4, were analyzed. Among these, BAK1a expression was upregulated, while the expression of all other genes was downregulated (Fig. 6d). Furthermore, MeJA was found to be higher at 3 d in response to LP treatment. DEGs involved in the JA signaling pathway, particularly MYC2-targeted component 2 (MYC2), showed that MYC2a expression was downregulated, while MYC2b and MYC2c were upregulated (Fig. 6e). No significant difference was found in GA3 contents in both cuttings at any time points. Key DEGs associated with GA signaling, gibberellin receptor (GID1), and DELLA family members, analysis showed the downregulation of GID1a, DELLA1, and DELLA2 (Fig. 6f). Finally, DEGs involved in the ethylene (ET) and salicylic acid (SA) signaling pathways were also analyzed. Key DEGs studied include nonexpressor of pathogenesis-related genes 1 (NPR1), pathogenesis-related protein 1 (PR-1), and ethylene response factor (ERF1/2). The expression of NPR1a, PR-1a, PR-1b, and ERF1/2a was downregulated (Supplementary Fig. S2). In addition, to provide a more intuitive overview of hormone-related DEGs under LP treatment, all identified DEGs within the major hormone signaling pathways were mapped onto the KEGG plant hormone signal transduction pathways, which are presented in Supplementary Fig. S3.
Validation of RNA-seq data using RT-qPCR
-
To validate the accuracy of the RNA-seq results, nine DEGs associated with various hormones and involved in key pathways related to AR formation were selected for RT-qPCR analysis. The RT-qPCR results showed that the expression patterns of most DEGs were consistent with the RNA-seq data, although the expression of B-ARR2 did not fully match the RNA-seq results (Fig. 7). Overall, these findings support the reliability of the RNA-seq dataset.
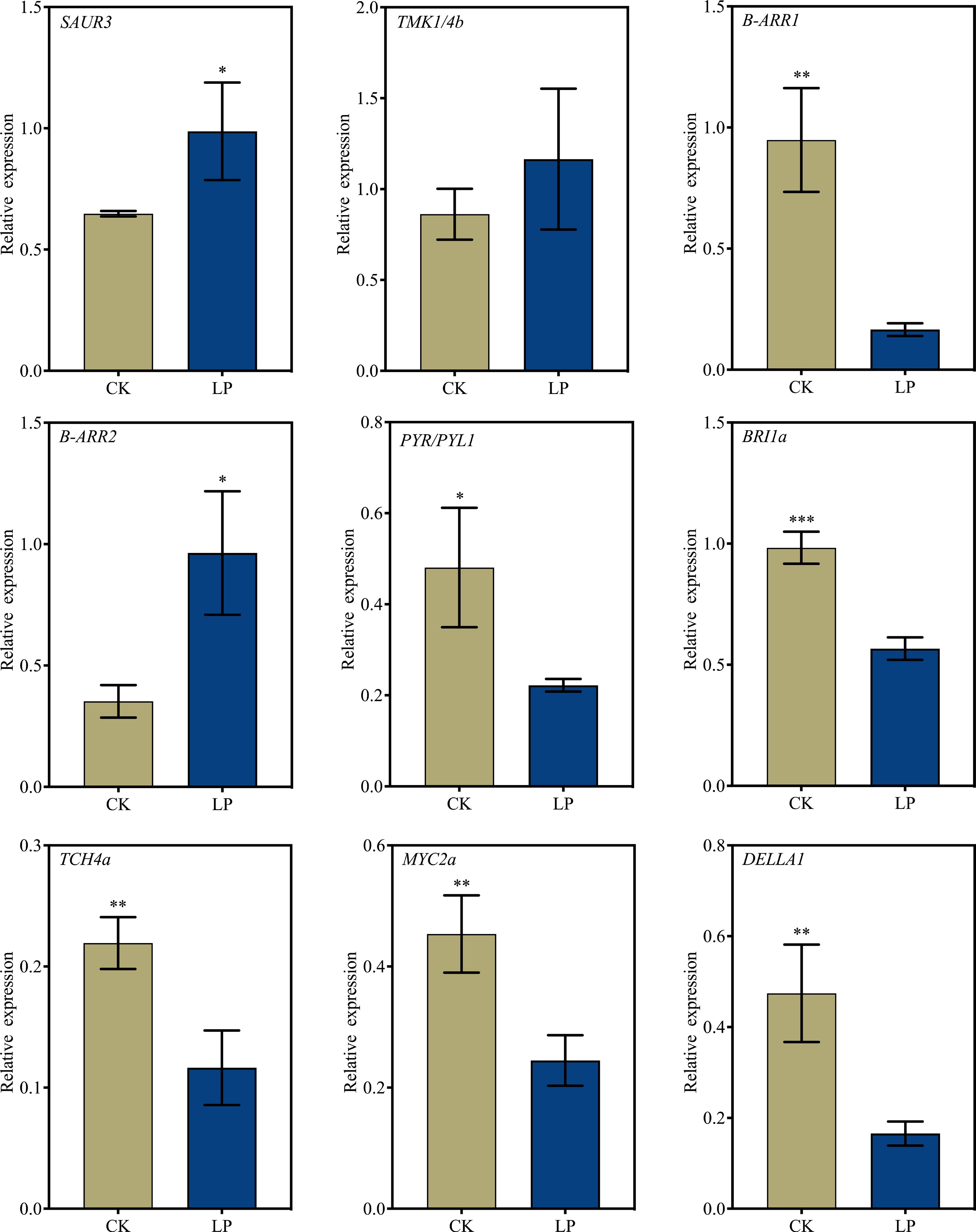
Figure 7.
RT-qPCR validation of selected differentially expressed genes (DEGs) to confirm the reliability of RNA-seq data. K2 apple rootstock stem cuttings were treated with low-phosphorus (LP, 0.08 g/L; total P 0.12 g/L) or grown on the control medium (CK, basal P level of standard MS medium, 0.04 g/L) and sampled on the 3rd day of adventitious root (AR) formation. Error bars indicate mean ± SD from three biological replicates. Different asterisks denote significance levels: ns, p > 0.05; * p < 0.05; ** p < 0.01; *** p < 0.001.
-
AR formation is a complex physiological and biological process influenced by multiple factors, many of which remain poorly understood, including the role of P. In the context of excision-induced AR formation, two key factors play a critical role: the physical wounding at the stem base and the absence of resources and signals typically provided by the mother plant[25,26]. Wounding activates a specialized developmental program in specific responsive cells, initiating de novo root formation[1]. In apples, this process occurs in three well-defined phases: the induction phase (0–3 d), the initiation phase (3–8 d), and the emergence phase (8–16 d)[2]. P availability has been shown to significantly influence root system architecture and AR formation across various plant species. For instance, in Arabidopsis, low P availability promotes the development of a highly branched root system, characterized by the stimulation of lateral roots (LRs) and root hairs, while inhibiting primary root (PR) growth[5,27−30]. Similarly, in species such as maize, rice, and common bean, AR development is enhanced under low P conditions[7,31]. Despite the significant importance of P in LR and PR development, its effects on AR formation in apples remained unexplored prior to this study.
The results of this study demonstrate that P availability plays a critical role in AR formation in K2 apple rootstock stem cuttings. LP treatment significantly enhanced AR formation, as evidenced by increased rooting percentage and AR numbers per cutting, while HP treatment had an inhibitory effect, likely due to P toxicity. These findings align with previous studies showing that low P availability promotes root branching and AR development in other plant species[7,31]. Anatomical analysis revealed that LP treatment stimulated cell division and primordia formation in the cambial and agminated differentiated cells, contributing to the increased AR formation observed in LP-treated cuttings. To further explore the physiological and molecular mechanisms underlying these phenotypes, endogenous hormone levels were assessed, and mRNA analysis was conducted. These analyses aim to provide a comprehensive understanding of the genetic and biochemical pathways involved in LP-mediated AR formation in apple rootstocks.
Hormones that promote AR formation under LP
-
P availability interacts with plant hormones, particularly auxins, which play a central role in AR formation. In this study, IAA content was significantly higher at 3 and 8 d in LP-treated cuttings compared to CK cuttings, but decreased at 16 d. This pattern aligns with the observed rooting phenotypes, where LP treatment promoted AR formation, consistent with the role of auxin as a stimulant in early AR stages (induction and initiation) and its reduced requirement during later emergence[10,16,32]. The decline in IAA at 16 d likely reflects the stage-specific demand: once root primordia have formed and ARs begin to emerge, auxin-mediated stimulation of cell division and differentiation is largely complete. Reduced IAA levels at this stage may help balance other hormones, which regulate the maturation and elongation of emerging roots. RNA-seq data revealed seven DEGs related to auxin signaling: four (ARF1, GH3.1, SAUR1, and SAUR2) were downregulated, and three (SAUR3, TMK1/4a, and TMK1/4b) were upregulated. While this pattern may seem inconsistent with the observed IAA levels and rooting phenotypes, the specific roles of these genes provide a plausible explanation. ARF1 and GH3.1, both negative regulators of auxin signaling, were expressed at lower levels in LP-treated cuttings. ARF1 represses auxin signaling by binding to auxin response elements (AuxREs) without activating transcription, inhibiting genes involved in cell division and differentiation[33,34]. Its downregulation likely enhanced AR formation by reducing repression of auxin-mediated processes. Similarly, GH3.1 conjugates free IAA to amino acids, reducing active auxin levels. Its lower expression in LP-treated cuttings likely allowed higher free auxin levels, promoting AR initiation[35,36]. In contrast, the upregulation of SAUR3 and TMK1/4 genes suggests a positive role in AR formation. SAUR3, which responds rapidly to auxin, likely facilitated cell elongation and division during early rooting stages[37]. TMK1/4 genes mediate non-canonical auxin signaling, regulating cell wall loosening, pH, and cytoskeletal reorganization—processes critical for cell division and primordia formation[38]. Their upregulation aligns with the observed rooting phenotypes, suggesting compensatory roles in AR development.
Hormones that inhibit AR formation under LP
-
CKs, like auxins, play a crucial role in the regulation of AR formation, particularly during the early stages, where cell division is essential. However, CKs often exhibit an antagonistic relationship with auxin, which can influence the establishment of AR founder cells[1,25] and negatively affect AR formation[18]. In this study, ZR levels were consistently higher in CK-treated cuttings at all time points, aligning with its role as a negative regulator of AR formation. This is consistent with previous findings in cucumber, where ZR was shown to inhibit AR formation in the root xylem[39] as well as in apples[2]. Elevated CK levels are known to restrict root growth by limiting the root meristem region through CRE1/AHK3/B-ARR signaling[40−42]. In Arabidopsis, exogenous CK application inhibited root initiation, and this effect could not be reversed by auxin application, highlighting the strong inhibitory role of CKs in AR formation[43,44]. CKs act directly on root founder cells to inhibit root initiation, and this inhibition is regulated independently of auxin signaling pathways[41,45,46]. In LP-treated cuttings, the expression of CK signaling genes revealed a pattern that supports the observed rooting phenotypes. The upregulation of APH1, a gene involved in CK biosynthesis, and the downregulation of B-ARR1 and B-ARR2, which are positive regulators of CK signaling, suggest a reduction in CK responsiveness in LP-treated cuttings. This modulation likely contributed to the enhanced AR formation by maintaining an optimal auxin-to-CK ratio, which is critical for root initiation[18]. B-ARRs are known to promote CK signaling, driving cell division and differentiation, while A-ARRs act as feedback inhibitors to prevent overactivation of CK pathways[47,48]. The downregulation of B-ARR1 and B-ARR2 in LP-treated cuttings may have reduced CK signaling, thereby alleviating its inhibitory effects on AR formation. The elevated ZR levels in CK-treated cuttings and the modulation of CK signaling genes in LP-treated cuttings highlight the antagonistic role of CKs in AR formation. The downregulation of B-ARR1 and B-ARR2, along with the upregulation of APH1, likely reduced CK signaling in LP-treated cuttings, promoting AR formation by maintaining an optimal auxin-CK balance.
ABA is a stress-responsive hormone that generally inhibits rooting by suppressing cell division and elongation, which is crucial for root initiation and development[20,49,50]. In this study, ABA levels were consistently higher in CK-treated cuttings at all time points, while LP-treated cuttings exhibited significantly lower ABA content. This observation is consistent with previous studies demonstrating that elevated ABA levels restrict root formation, whereas reduced ABA accumulation in LP-treated cuttings likely facilitated enhanced AR development, reinforcing the role of ABA as a negative regulator of rooting[2]. High ABA levels inhibit AR formation by disrupting auxin transport and signaling, reducing auxin accumulation at sites of root initiation, which is essential for AR development. The antagonistic interaction between ABA and auxin leads to decreased auxin availability, thereby inhibiting rooting[51−53]. In this study, the observed increase in IAA levels at 3 and 8 d in LP-treated cuttings, which correlated with enhanced AR initiation, further supports this mechanism. In the ABA signaling pathway, the downregulation of PYR/PYL1 and PYR/PYL2 suggests reduced ABA signaling, which may have alleviated ABA-mediated inhibition of AR formation. This is consistent with the known role of ABA in root development, where it negatively regulates cell division and elongation in the root meristem, thereby restricting root growth[51,54]. By reducing ABA signaling, LP-treated cuttings likely promoted cell division and elongation, facilitating the enhanced AR formation observed in this group.
BR plays a dual role in AR formation: they promote root development at low concentrations but inhibit it at high levels. In this study, BR levels were consistently higher in CK-treated cuttings at all time points, while LP-treated cuttings exhibited lower BR content. This aligns with previous findings that high BR levels inhibit AR formation in apple rootstocks[2]. BRs are known to interact with other hormonal pathways, particularly auxin signaling, which is critical for AR initiation. Excessive BRs can disrupt auxin transport and signaling, leading to reduced AR formation. Additionally, high BR levels can suppress cell division in root-forming tissues, further inhibiting AR development[21,55,56]. BRs and auxin often interact antagonistically during root development. High BR levels can interfere with auxin transport and signaling, which are critical for AR initiation. In this study, the lower BR levels in LP-treated cuttings likely alleviated this inhibitory effect, allowing auxin signaling to promote AR formation. This is supported by the observed increase in IAA levels at 3 and 8 d in LP-treated cuttings, which correlated with enhanced AR initiation. The expression of BR signaling genes further supports this mechanism, where only BAK1a was upregulated, while all other genes were downregulated in LP-treated cuttings. The downregulation of BRI1 and other BR signaling components suggests reduced BR responsiveness in LP-treated cuttings, which may have contributed to the enhanced AR formation by alleviating BR-mediated inhibition of auxin signaling and cell division.
-
LP treatment significantly enhanced AR formation in K2 apple rootstocks, as evidenced by increased rooting percentage and AR numbers. At the physiological and molecular level, LP promoted AR primordia formation by modulating endogenous hormone levels—increasing IAA while decreasing ZR, ABA, and BR—and altering the expression of key hormone-related genes, including ARF1 and GH3.1 (auxin signaling), B-ARR1 (CK signaling), PYR/PYL1 (ABA signaling), and BAK1a (BR signaling). These results provide important insights into the molecular and physiological processes underlying P-induced AR formation and offer potential strategies for improving adventitious rooting in apple rootstocks.
This work was financially supported by the Science and Technology Major Project of Xinjiang Production and Construction Corps (Grant No. 2023AB077), Shaanxi Zhaojin Xianheng Agricultural Science and Technology, the China Apple Research System (Grant No. CARS-27), the Young Talent Fund of University Association for Science and Technology in Shaanxi, China (Grant No. 20240218), and the Chinese Universities Scientific Fund (Grant No. 2452023005).
-
The authors confirm their contributions to the paper as follows: study conception and design: Tahir MM, Zhu K; data analysis: Li Q, Yu J, Jing M, Mao J; investigation: Bao L; supervision: Bao L; draft manuscript preparation: Tahir MM, Zhu K. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Muhammad Mobeen Tahir, Kexin Zhu
- Supplementary Table S1 Gene-specific primers used in RT-qPCR analysis.
- Supplementary Table S2 Summary of the sequencing statistics obtained from each sample.
- Supplementary Table S3 Summary of the total, multiple and uniquely mapped reads obtained from each sample.
- Supplementary Fig. S1 Transcriptomic analysis of K2 apple rootstock stem cuttings on the 3rd day of adventitious root (AR) formation under low-phosphorus (LP, 0.08 g/L; total P 0.12 g/L) treatment compared with the control (CK, basal P level of standard MS medium, 0.04 g/L).
- Supplementary Fig. S2 Transcriptomic analysis of K2 apple rootstock stem cuttings on the 3rd day of adventitious root (AR) formation under low-phosphorus (LP, 0.08 g/L; total P 0.12 g/L) treatment compared with the control (CK, basal P level of standard MS medium, 0.04 g/L).
- Supplementary Fig. S3 Transcriptomic analysis of K2 apple rootstock stem cuttings on the 3rd day of adventitious root (AR) formation under low-phosphorus (LP, 0.08 g/L; total P 0.12 g/L) treatment compared with the control (CK, basal P level of standard MS medium, 0.04 g/L).
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Tahir MM, Zhu K, Li Q, Yu J, Jing M, et al. 2026. Phosphorus-induced enhancement of adventitious root formation in K2 apple rootstocks: hormonal and gene expression insights. Fruit Research 6: e008 doi: 10.48130/frures-0025-0046 

Phosphorus-induced enhancement of adventitious root formation in K2 apple rootstocks: hormonal and gene expression insights
- Received: 02 September 2025
- Revised: 04 November 2025
- Accepted: 12 November 2025
- Published online: 09 February 2026
Abstract: Adventitious root (AR) formation is a crucial process for the clonal propagation of horticultural crops. Phosphorus (P), a key macronutrient, plays an essential role in various physiological and molecular processes involved in AR formation, including cell division and development. However, its role in apple rootstock remains underexplored. This study investigates the effects of three P treatments—low P (LP), medium P (MP), and high P (HP)—along with a control group (CK, containing the basal P level of standard MS medium)—on AR formation in K2 apple rootstock stem cuttings. The results demonstrated that LP treatment significantly increased both rooting percentage and the number of ARs per cutting compared to CK, whereas HP treatment inhibited both. Anatomical analysis revealed that LP-treated cuttings exhibited enhanced AR primordia formation, accompanied by increased endogenous levels of indole-3-acetic acid (IAA), while levels of zeatin riboside (ZR), abscisic acid (ABA), and brassinosteroid (BR) were reduced. Transcriptome analysis identified 1,009 differentially expressed genes (DEGs), including 383 upregulated and 626 downregulated, enriched in metabolic and hormone signaling pathways. LP altered the expression of key hormone-related genes—ARF1 and GH3.1 (auxin signaling), B-ARR1 (cytokinin signaling), PYR/PYL1 (ABA signaling), and BAK1a (BR signaling)—which may synergistically promote AR initiation and development. These results reveal the physiological and molecular basis of LP-enhanced AR formation and provide insights into nutrient–hormone interactions in root development.
-
Key words:
- Apple rootstock /
- Adventitious root (AR) formation /
- Phosphorus /
- Endogenous hormones /
- RNA-seq